
Sara Dennis-Pacheco a, José Raúl Romo-León a, *, Alejandro E. Castellanos-Villegas a, María Cristina Peñalba-Garmendia a y Lara Alejandrina Cornejo-Denman b
a Universidad de Sonora, Departamento de Investigaciones Científicas y Tecnológicas, Luis Donaldo Colosio s/n, entre Sahuaripa y Reforma, Edificio 7G, Centro, 83000 Hermosillo, Sonora, México
b Dalhousie University, School for Resource and Environment Studies, Kenneth C. Rowe Management Building, 6100 University Ave, PO BOX 15000, Halifax, Nova Scotia, B3H 4R2 Canadá
*Autor para correspondencia: jose.romo@unison.mx (J.R. Romo-León)
Recibido: 5 diciembre 2022; aceptado: 9 octubre 2023
Resumen
La Reserva Jaguar del Norte es una propiedad privada ubicada en la sierra sonorense, dentro de una compleja matriz paisajística, con un amplio legado de uso ganadero. Esta región, identificada como prioritaria para la conservación, dispone de pocos datos sobre las trayectorias de cambio en la vegetación, un elemento clave para evaluar el impacto de las áreas naturales protegidas. Este trabajo presenta información acerca del efecto de las acciones con fines de conservación, sobre la composición de la vegetación tras el establecimiento de la reserva en el año 2003. Para ésto, se caracterizaron las comunidades vegetales presentes y se generaron clasificaciones supervisadas de cobertura con imágenes satelitales de mediana y alta resolución para realizar un análisis cambio de la cobertura vegetal (2003-2018). Las precisiones generales de las clasificaciones fueron iguales o superiores a 0.75, valor que aumentó al utilizar imágenes de mayor resolución espacial. El uso combinado de diferentes resoluciones espaciales presentó ventajas para entender las transiciones de composición entre diferentes comunidades vegetales. Las tendencias encontradas indicaron un aumento del matorral subtropical y una disminución del pastizal inducido, lo que refleja transformaciones hacia una mayor diversidad de especies y una reducción de posibles amenazas para la biodiversidad a nivel paisaje.
Palabras clave: Conservación; Ecología del paisaje; Percepción remota
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Vegetation composition after the establishment of a natural protected area in northwestern Mexico
Abstract
The Northern Jaguar Reserve is a private property located in the Sonoran mountains, within a complex landscape matrix with a wide legacy of livestock use. This region, identified as priority for conservation, has scarce data on vegetation change trajectories, a key element to assess the impact of natural protected areas. This work presents information about the effect of actions for conservation purposes on the vegetation composition after the establishment of the reserve in 2003. The plant communities were characterized, and land cover supervised classifications were generated with medium- and high-resolution satellite images to carry out a cover vegetation change analysis (2003-2018). General accuracies of the classifications were equal to or greater than 0.75, increasing when using images of higher spatial resolution. Combining different spatial resolutions presents advantages for understanding compositional transitions between different plant communities. The trends indicated an increase in subtropical scrub and a decrease in induced grassland, reflecting transformations towards a greater diversity of species and a reduction in potential threats to biodiversity at the landscape level.
Keywords: Conservation; Landscape ecology; Remote sensing
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
Actualmente, los profundos cambios en la estructura y funcionamiento de los ecosistemas han contribuido a la pérdida de biodiversidad (WWF, 2020), impulsada principalmente por la desaparición de hábitats, a través de cambios de uso y cobertura de suelo (Jantz et al., 2015; Rimal et al., 2019). Ante esta situación, se han buscado estrategias de conservación que consideren múltiples escalas espaciales, en áreas con amplios gradientes físicos y biológicos, con características ambientales únicas (Poiani et al., 2000). Con el objetivo de preservar la biodiversidad en sus diferentes niveles de organización (ecosistemas, poblaciones y genes) (Borda-de Água, 2019), se han establecido áreas naturales protegidas (ANP), donde se regulan las actividades antropogénicas, para evitar la degradación a diferentes escalas estructurales y funcionales en la naturaleza (Ferreira et al., 2013; Henle et al., 2014; Kumar-Verma, 2016). Las ANP son una herramienta de conservación que cumple diversos objetivos, tales como salvaguardar el funcionamiento de los ecosistemas, la biodiversidad y los servicios ambientales (Bezaury-Creel y Gutiérrez-Carbonell, 2009). Si bien todas estas áreas tienen la característica común de ser espacios físicos naturales, en donde los ambientes originales no han sido significativamente alterados, su categoría se designa según los propósitos principales y los organismos administradores que la dirigen, por ejemplo, el gobierno de un país, organizaciones no gubernamentales o predios de propiedad privada (Conanp, 2019). En las últimas décadas, el establecimiento, manejo y preservación de ANP, se han convertido en la principal política de conservación de sitios naturales, tanto en México, como en el resto del mundo (Durand y Jiménez, 2010).
La designación de estos sitios plantea escenarios ideales para observar cómo el tipo de manejo que se ejerce sobre un área puede repercutir en la estructura y procesos naturales de las comunidades a través del tiempo (Kamwi et al., 2017). Específicamente, comprender cómo ocurren los cambios en la composición de la vegetación dentro de estas áreas, conforma una importante contribución a nuestro entendimiento del efecto de las ANP en la matriz paisajística, y en la tendencia de cambio natural propia de regiones que registran poca o nula actividad humana (Guerra et al., 2019). Más aún, analizar una ANP de reciente creación y su evolución durante sus primeros años de establecimiento, presenta la oportunidad única de realizar análisis continuos y a largo plazo acerca de diferentes procesos ecológicos que ocurren en sus comunidades vegetales (Gaston et al., 2008; Coetzee, 2017).
Los cambios en la composición de la vegetación tienen efectos claros sobre la biodiversidad a diferentes escalas, por lo que es un enfoque relevante para considerar en proyectos que se encaminan a la conservación (Freitas y Mantovania, 2017). Al modificarse la composición de los diferentes elementos que interactúan en el paisaje, se pueden generar efectos positivos, negativos o neutrales, afectando las características estructurales y funcionales de los ecosistemas, que se encuentran en constante retroalimentación de materia y energía (Ramírez-Arce et al., 2022). Algunas consecuencias de estos cambios en la composición de la vegetación son las alteraciones en la provisión de servicios ecosistémicos (Rimal et al., 2019), modificaciones en diversos mecanismos biofísicos y biogeoquímicos (Duveiller et al., 2018), así como en el clima a escala local, regional y global (Arneth, 2015).
Si bien es común que los cambios de uso y cobertura de suelo sean concebidos como la pérdida de funciones y efectos negativos en el medio ambiente (pérdida de vegetación natural), este fenómeno también hace referencia a la recuperación de la vegetación a partir de sitios degradados, lo cual conlleva efectos positivos en términos de biodiversidad (Turner y Gardner, 2015). En el caso de las ANP, las restricciones de uso de suelo y las acciones enfocadas a la restauración, pueden promover variaciones espacio-temporales en las comunidades vegetales, observables y cuantificables, que a su vez brinden información útil para su adecuada gestión (Arneth, 2015). En estudios previos se han encontrado tendencias de recuperación en las comunidades vegetales tras la implementación de programas de conservación, donde antes había un régimen de aprovechamiento (Kamwi et al., 2017). En contraste, se ha observado que, bajo condiciones deficientes de gestión con fines de conservación, el incremento en la práctica de actividades productivas humanas ha provocado la pérdida de vegetación nativa y el aumento de sitios desprovistos de vegetación o con especies exóticas (Ramachandra et al., 2018). Específicamente, los usos de suelo ganaderos han mostrado tener efectos negativos en la diversidad de las comunidades vegetales (McDonald et al., 2019).
El estudio integral de procesos ecológicos que ocurren al interior de las ANP presenta desafíos, principalmente relacionados a su accesibilidad y a la escala espacial y temporal que se requiere para abordarlos desde la perspectiva de la conservación de especies y función de los ecosistemas (Gross et al., 2009; Wang et al., 2020). Sin embargo, en la actualidad es posible realizar seguimientos y detección sistemática de cambios en la cobertura de suelo, a nivel paisaje, mediante instrumentos de teledetección especializados (Duan et al., 2020; Willis, 2015). Los progresos en la tecnología relacionada con los sensores montados en satélites permiten realizar observaciones puntuales a lo largo del tiempo (Kumar et al., 2015), lo que hace posible llevar a cabo monitoreos continuos de las transformaciones de uso y cobertura de suelo (Bertzky et al., 2012; Fuente et al., 2019). Aunque las aplicaciones de percepción remota y teledetección son muy diversas, estas herramientas han encontrado un nicho especial en el monitoreo de la vegetación (Houborg et al., 2015). El monitoreo remoto de las diferentes comunidades vegetales, es primordial para detectar tendencias y variaciones tras un cambio de régimen de uso (Koch et al., 2017; Sokolov et al., 2020). Además, permite evaluar la efectividad de la gestión con fines de conservación, sin perturbar el paisaje en regiones ecológicamente importantes (Wang et al., 2020).
En México, gran parte del territorio está cubierto por zonas áridas, semiáridas y subhúmedas, las cuales comprenden más de 50% del territorio nacional (Conaza, 1994; UACh, 2011). Los paisajes de estas zonas cuentan con diferentes ecorregiones, con una variada geología y biogeografía y, por consiguiente, comunidades bióticas únicas (Maestre et al., 2021). Sin embargo, estas regiones han sido poco estudiadas y, por ende, sus características ecológicas son menos comprendidas que las de otras regiones del país (p. ej., regiones tropicales o regiones boscosas) (Briones et al., 2018; Flores-Olvera, 2011). En estudios relacionados con la ecología del paisaje, las tendencias de cambio en ecosistemas de zonas áridas, semiáridas y subhúmedas son a menudo desestimadas, en comparación con las de estudios realizados en ecosistemas húmedos (Arroyo-Rodríguez et al., 2017; Hernández-Moreno et al., 2021; Zhang et al., 2022). Más aún, existen pocos estudios en donde se evalúe el impacto de las áreas protegidas sobre la extensión y condición de la vegetación en este tipo de regiones con climas más extremos (Hernández-Moreno et al., 2021; Sharma et al., 2020; Zhang et al., 2022).
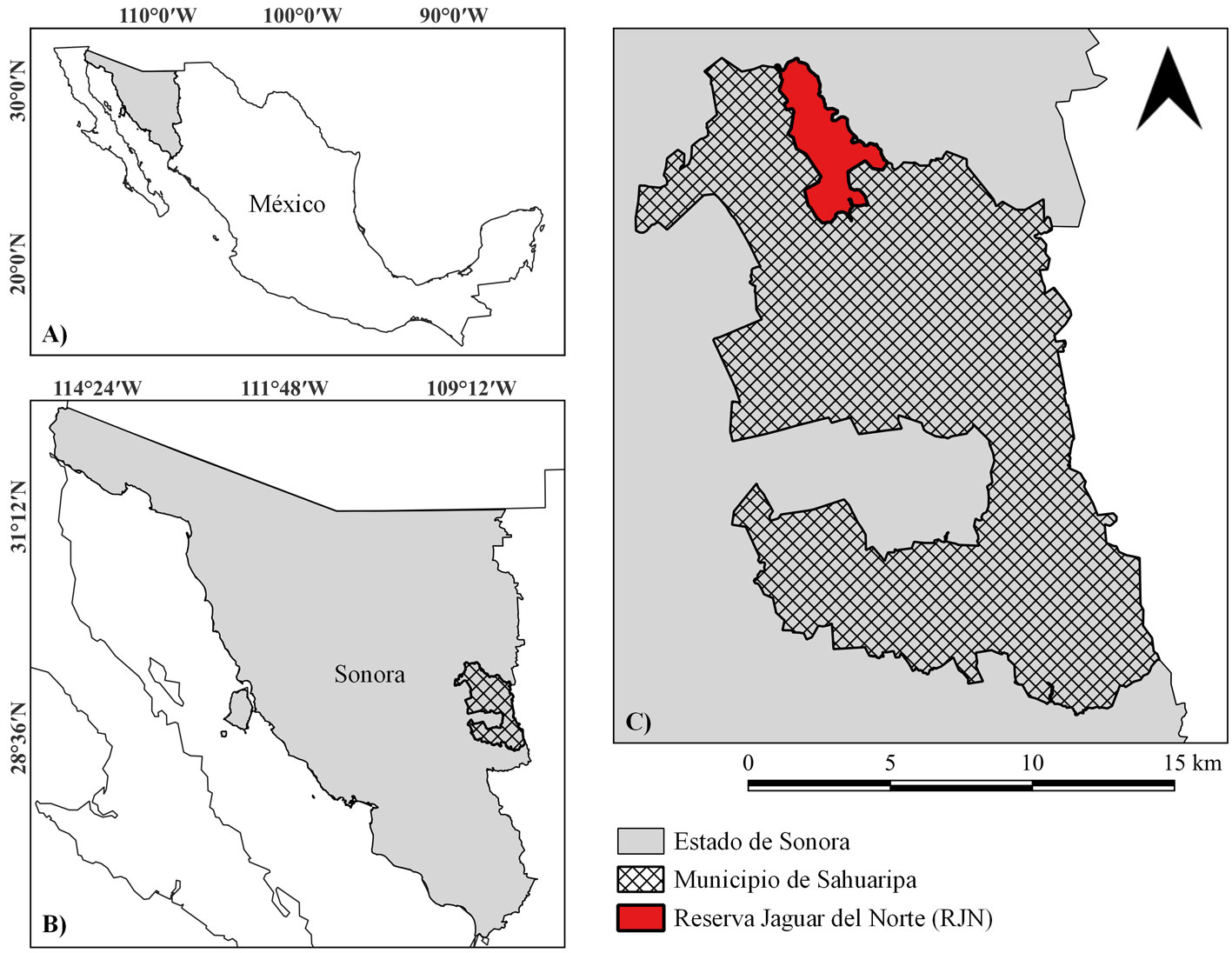
La Reserva Jaguar del Norte (RJN) es una ANP privada, ubicada en el noroeste de México (estado de Sonora). El área que comprende actualmente fue constituida a partir de la compraventa de ranchos ganaderos, adquiriendo su primera porción, en el año 2003, por Naturalia A.C., una asociación para la conservación, que tiempo después sumó esfuerzos con Northern Jaguar Proyect, asociación sin fines de lucro con acción binacional, dedicada a la protección de los jaguares y su hábitat. A partir de ese momento, se concreta lo que hoy en día es la Reserva Jaguar del Norte. Al estar ubicada en las inmediaciones de la sierra Madre Occidental, esta ANP cuenta con una compleja topografía que alberga una gran diversidad de vegetación. La RJN se encuentra dentro de una región terrestre prioritaria para la conservación de la biodiversidad de México, por su alta riqueza ecosistémica y específica (Arriaga, 2000).
Hoy en día, la RJN cuenta con aproximadamente 23,300 ha para fines de conservación, con el objetivo principal de proteger a la población de jaguar (Panthera onca) más septentrional en el continente americano, a través del mantenimiento y preservación de su hábitat en una extensa área en el norte de México (Northern Jaguar Project, 2022). Debido a su reciente creación y a sus características biofísicas, esta ANP presenta condiciones ideales para observar las trayectorias y tendencias que ha seguido la vegetación, después del cambio de régimen de uso y tras la implementación de acciones específicas para la conservación, las cuales se han centrado, principalmente, en la exclusión de ganado bovino, la remoción activa de especies inducidas, en específico de zacate buffel (Cenchrus ciliaris L.), y la regeneración natural de sitios con un amplio legado de uso de suelo con ganadería extensiva.
Comprender la dinámica de la vegetación a largo plazo es clave para la gestión sustentable de los servicios ecosistémicos y recursos naturales (Maass et al., 2010), ya que conocer las trayectorias de cambio de la vegetación, permite obtener un mejor entendimiento acerca de los procesos ecológicos, a múltiples escalas espaciotemporales, derivados de las acciones para la conservación. En consecuencia, el presente estudio tiene como objetivo principal analizar la dinámica en la composición de la vegetación dentro de la RJN, en los primeros años posteriores a su establecimiento, en el año 2003. Para lograrlo, se utilizaron aproximaciones que combinan el uso de técnicas de percepción remota en conjunto con análisis espacial y trabajo de campo. Los objetivos particulares del presente trabajo son: 1) caracterizar las comunidades vegetales presentes dentro de la RJN, obteniendo datos in situ de cobertura por estratos de vegetación y especies, además de estimaciones de diversidad de especies por comunidad vegetal y 2) la generación de un análisis de clasificación y cambio de cobertura de suelo a nivel paisaje entre los años 2003 y 2018, utilizando aproximaciones de teledetección, con diferentes resoluciones espaciales.
Materiales y métodos
La Reserva Jaguar del Norte es una ANP de privada, ubicada en el centro-este de Sonora, México (29°16’, 29°32’ N, 109°13’, 109°1’ O), en el municipio de Sahuaripa (fig. 1). Actualmente, cuenta con una extensión de 23,300 ha (233 km2). Debido a su ubicación, adyacente a la sierra Madre Occidental, el clima y la vegetación en el área son variables, como consecuencia del gradiente de elevación que presenta (450 a 1,200 m snm). El tipo de clima va de árido-cálido a templado, con una precipitación media anual de 574.6 a 600.8 mm, en un régimen biestacional (80% en verano y 20% en invierno) (Arriaga et al., 2000; Lin-Quintana, 2017). En cuanto a la vegetación, se pueden encontrar desde matorrales espinosos en zonas bajas, hasta bosques de encino en las zonas altas (INEGI, 2014). La compleja matriz paisajística que comprende la RJN permite identificar 7 comunidades vegetales y zonas de suelo desnudo. En este estudio, de las diferentes comunidades vegetales identificadas, el pastizal natural fue la única que no fue muestreada en campo, por lo que solo se consideró en las clasificaciones supervisadas y posterior análisis de cambio.
Para describir a las comunidades vegetales presentes en la RJN, se diseñó un esquema de clases como el propuesto por Anderson et al. (1976), que consistió en visitar sitios en campo para caracterizar a las diferentes comunidades vegetales o “clases”, y en cada una de ellas se realizaron muestreos estratificados, siguiendo el método de línea de Canfield (1941). En cada comunidad vegetal se seleccionaron 4 sitios y en cada uno se dispuso un transecto lineal de 30 m. Para cada transecto se registraron las medidas de inicio y fin de las coberturas individuales de árboles y arbustos interceptados por la línea (considerando solamente a aquellos individuos superpuestos a otros, es decir, el estrato superpuesto horizontal más alto). Por último, se estableció el porcentaje de cobertura para cada una de las especies individuales y para el conjunto de especies leñosas (árboles y arbustos).
Para evaluar la diversidad de especies se estimó la riqueza y dominancia de las comunidades vegetales mediante el cálculo de los primeros 3 números de Hill, 2 de los cuales utilizan transformaciones matemáticas de los índices originales de Shannon y Simpson (Hill, 1973). Adicionalmente, se siguieron criterios similares a los métodos de Braun-Blanquet (1932), donde se estimó visualmente la cobertura herbácea con 3 cuadrantes de 2 m2 cada uno, dispuestos en forma equidistante al inicio (0 m), parte media (15 m) y final (30 m) de cada transecto lineal mencionado anteriormente (12 cuadrantes por comunidad vegetal; 18 m2). Los muestreos de campo se realizaron durante el mes de septiembre del año 2021.
La delimitación de la RJN fue provista por su equipo de manejo en formato “shapefile”. Las imágenes satelitales para realizar las clasificaciones de cobertura de suelo a nivel paisaje se obtuvieron de los sensores Landsat 5 TM y Landsat 8 OLI (30 m de resolución espacial), para fechas correspondientes a 2003, 2010 y 2018. Esto se realizó por medio de la plataforma de Earth Explorer, del Servicio Geológico de los EUA (USGS, 2022). Adicionalmente, para el año 2018, se obtuvieron imágenes del sensor PlanetScope (3 m de resolución espacial), mediante la plataforma Planet Explorer (Planet Team, 2017). Para seleccionar los años de adquisición de las imágenes satelitales se buscó una representación temporal equidistante entre las fechas de captura y que la precipitación de esos años fuera similar, con el fin de tomar en cuenta la variabilidad espectral ante la respuesta fenológica de la vegetación. Para esto último, también se descargaron 2 imágenes por año, correspondientes a la temporada húmeda y seca en la región (Romo et al., 2014). Los datos de precipitación fueron obtenidos del satélite TRMM (Tropical Rainfall Measuring Mission, 2011), información con la cual se calcularon los montos mensuales y anuales de lluvia para el área de estudio.
Por último, como parte de las bases de datos utilizadas dentro del proceso de clasificación, se obtuvo un modelo de elevación digital (DEM, por sus siglas en inglés) para la zona. Esta información fue obtenida del conjunto de datos de elevación del USGS y ajustado a la resolución espacial de las imágenes satelitales utilizadas (30 y 3 m).
Las variables utilizadas en las clasificaciones de Landsat se derivaron para cada uno de los compuestos de reflectancias, calculando componentes principales, transformación de Tasseled Cap (Collins y Woodcock, 1996; Fung y LeDrew, 1987), texturas de terreno (Alhaddad et al., 2007; Asner et al., 2002), índice de vegetación de diferencia normalizada (NDVI) (Tucker, 1979), índice de vegetación mejorado (EVI) (Huete et al., 2002; van Leeuwen et al., 1999) e índice de vegetación ajustado al suelo (SAVI) (Huete, 1988). Derivado del modelo de elevación digital, se obtuvieron capas de información de la elevación media sobre el nivel del mar, pendiente y orientación del terreno. Las variables utilizadas para la clasificación de las imágenes de PlanetScope también se derivaron para cada uno de los compuestos de reflectancias, calculando texturas del terreno, índices de vegetación (NDVI, EVI y SAVI), además de capas de pendiente y orientación, derivadas del modelo de elevación digital. Las variables derivadas para las clasificaciones, independientemente del sensor del que provinieran las imágenes, se obtuvieron para ambas temporadas (húmeda y seca), excepto la elevación, la pendiente y la orientación, que no dependen del momento de adquisición de las imágenes.
Se utilizaron aproximaciones con modelos pertenecientes a la familia de árboles de clasificación y regresión (CART) (Breiman et al., 1984; Tso y Mather, 2009). Específicamente, las clasificaciones se realizaron con el algoritmo C5.0 de reclusión múltiple, el cual tiene la bondad de crear varios modelos de respuesta y seleccionar el de mejor ajuste. Todas las clasificaciones se ejecutaron en el Software R, con interfase RStudio (R Core Team, 2021).
Para cada una de las clases de cobertura de suelo se obtuvieron entre 55 y 266 puntos de entrenamiento (266 en encinar, 256 en matorral subtropical, 253 en matorral xerófilo, 252 en mezquital, 260 en pastizal natural, 250 en pastizal inducido, 257 en vegetación de arroyos y 55 en suelo desnudo), los cuales fueron utilizados para generar muestras de las variables derivadas para cada uno de los sitios específicos donde se conociera con certeza la presencia de cada una de las clases. Los puntos de entrenamiento fueron colectados con GPS por el equipo técnico de la RJN y complementados, en minoría, mediante la selección de puntos adicionales en la aplicación Google Earth Pro (1,849 puntos de entrenamiento en total). Los datos de entrenamiento fueron divididos aleatoriamente en 2 particiones: entrenamiento y validación (evaluación de la precisión alcanzada), los cuales representaron 70 y 30% del total de los datos por clase, respectivamente.
Por último, para cada una de las clasificaciones generadas, se obtuvo la precisión general de la clasificación y se complementó con el índice de Kappa (Cohen, 1960), que representa la comparación de la precisión general con la precisión de la probabilidad aleatoria esperada. Además, se obtuvieron las precisiones de usuario y de productor para cada una de las clases de cobertura, con el fin de visualizar el desempeño del algoritmo. La precisión de usuario indica la probabilidad de que las clases asignadas representen la realidad, mientras que la precisión de productor hace referencia a la probabilidad de que la muestra de entrenamiento sea clasificada correctamente (Story, 1986).
Tabla 1
Descripción general de las clases de cobertura presentes en el área de estudio en 2018. Información modificada de INEGI (2014). Al final de la descripción de cada clase se indican, entre paréntesis y separados por una diagonal, los porcentajes de cobertura del estrato leñoso (arbóreo y arbustivo) y herbáceo. Abreviatura NE: variables no estimadas.
| Clase | Descripción |
| Encinar | Bosques formados predominantemente por encinos (Quercus spp.). Se encuentran en forma de grandes parches o franjas extendidas, usualmente en laderas de zonas altas, sin embargo, también se registra en cañadas y laderas de menor elevación, excepto en zonas con climas áridos (72/34) |
| Matorral subtropical | Arbustos o árboles bajos, inermes o espinosos, con cobertura densa, que se desarrollan entre zonas subhúmedas y matorrales de zonas áridas y semiáridas (61/53) |
| Matorral xerófilo | Arbustos bajos, con presencia dispersa de cactáceas, pastos y árboles caducifolios, adaptados a medios secos, con escasos periodos de humedad. Los individuos suelen presentar espinas, tallos fotosintéticos y hojas de tamaño pequeño (68/51) |
| Mezquital | Vegetación mayoritariamente arbórea y espinosa, donde predominan los mezquites (Prosopis spp.). Se encuentran en sitios semiáridos y subhúmedos, sobre suelos planos y profundos, donde puede existir agua subterránea (58/71) |
| Pastizal natural | Áreas dominadas por un estrato herbáceo, especialmente compuesto por gramíneas nativas. Se pueden encontrar en áreas extensas de planicies o laderas altas, con terrenos poco o nada accidentados de zonas semiáridas, con clima seco (NE) |
| Pastizal inducido | Praderas dominadas casi en su totalidad por el pasto exótico y con potencial invasor Cenchrus ciliaris (zacate buffel). Éste se desarrolla en suelos medianamente profundos de mesetas y laderas poco inclinadas (33/74) |
| Vegetación de arroyos | Vegetación que sigue el curso de los arroyos y ríos, incluso sin presencia de agua permanente. Los árboles de hasta 30 m de altura, acompañados por arbustos y herbáceas, que cubren gran porcentaje del suelo (87/69) |
| Suelo desnudo | Áreas desprovistas de vegetación, como posible consecuencia de secuelas por uso humano de suelo o por la deposición arenosa del curso de ríos y arroyos (NE) |
Con el fin de analizar la trayectoria, ubicación y extensión de los cambios en la cobertura de suelo en los años analizados, se realizaron análisis de cambio post clasificación (Romo-León et al., 2014). Para lo anterior, se eligieron las clasificaciones generadas con Landsat TM y Landsat OLI, para los años de 2003 y 2018, respectivamente. Para ambos años se utilizaron los mapas de cobertura a nivel paisaje productos de las clasificaciones y se sometieron a un análisis de posprocesamiento en el programa QGIS, utilizando el complemento PCA4CD-PCA para construir la capa de detección de cambios (Jensen, 2005). El análisis de cambio obtenido permite calcular el área y porcentaje de cambio para cada una de las clases presentes entre los años analizados.
Resultados
Las comunidades vegetales en el área de estudio se dividen en 7 clases de cobertura vegetal: encinar, matorral subtropical, matorral xerófilo, mezquital, pastizal natural, pastizal inducido y vegetación de arroyos (tabla 1). En la tabla 1 se observa que la cobertura del estrato leñoso (árboles y arbustos) presentó un mayor porcentaje en la vegetación de arroyos (87%), seguida por el encinar, el matorral xerófilo, matorral subtropical, mezquital y, por último, pastizal inducido (33%). Para el estrato herbáceo, el pastizal inducido fue la comunidad que mayor porcentaje de cobertura presentó (74%), dominado casi en su totalidad por zacate buffel (Cenchrus ciliaris). El mezquital y la vegetación de arroyo también presentaron alta cobertura en el estrato herbáceo, seguidos por el matorral subtropical, matorral xerófilo y, por último, el encinar con 34% de cobertura.
En cuanto a la cobertura de suelo por especies individuales, el porcentaje de cobertura leñosa por especie varió según la comunidad vegetal. En algunas clases, especies características de dicha la comunidad fueron dominantes en cobertura, como en el caso del encinar, con 30% de cobertura de Quercus chihuahuensis Trel., el mezquital, con 52% de cobertura de Prosopis velutina Wooton o la vegetación de arroyo, con 38% de cobertura de sicomoro (Platanus wrightii S. Watson). En el resto de las clases muestreadas, los dominantes en cobertura fueron el huizache (Vachellia campechiana (Mill.) Seigler et Ebinger) o el mezquite (Prosopis velutina) y ambos fueron las especies más comunes entre clases, pues se encontraron en 5 y 4 tipos de vegetación muestreados, respectivamente. Por el contrario, hay especies que se presentan exclusivamente dentro de una comunidad vegetal: i) en encinar, Condalia correllii M.C. Johnst., Quercus albocincta Trel. y Senegalia occidentalis (Rose) Britton et Rose, ii) en matorral subtropical, Croton sonorae Torr., Opuntia fulgida Engelm., Erythrostemon caladenia (Standl.) Gagnon et G.P. Lewis, Fouquieria macdougalii Nash, Jatropha cardiophylla (Torr.) Müll. Arg., Mimosa distachya Cav., Solanum grayi Rose, iii) en matorral xerófilo, Encelia farinosa A. Gray ex Torr., Lycium andersonii A- Gray, Parthenium tomentosum DC., Randia sonorensis Wiggins, iv) en mezquital, Lycium fremontii A. Gray y Phaulothamnus spinescens A. Gray, v) en pastizal inducido, Ayenia jaliscana S. Watson, Bursera fagaroides (Kunth) Engl., Bursera laxiflora S. Watson y vi) en vegetación de arroyos, Ambrosia ambrosioides (Delpino) W.W. Payne, Baccharis salicifolia (Ruiz et Pav.) Pers., Guazuma ulmifolia Lam., Platanus wrightii, Sabal uresana Trel., Salix bonplandiana Kunth, Sapindus saponaria L. y Senegalia greggii (A. Gray) Britton et Rose.
El matorral subtropical fue la clase con mayor riqueza de especies leñosas, seguido por matorral xerófilo y vegetación de arroyo (tabla 2). En cambio, el mezquital registró la menor riqueza, seguido por encinar y pastizal inducido. La riqueza de las comunidades se relacionó negativamente con la dominancia de 1 o pocas especies, ya que las comunidades con un bajo número de especies también presentaron una mayor dominancia (q1 y q2, menor número de especies abundantes y muy abundantes, respectivamente). Además, las comunidades con menos riqueza de especies también mostraron un número menor de especies restringidas a ellas (mezquital, encinar y pastizal inducido). Por el contrario, las comunidades vegetales con mayor riqueza de especies presentaron una menor dominancia y un mayor número de especies restringidas (matorral subtropical, vegetación de arroyos y matorral xerófilo).
Tabla 2
Diversidad estimada a partir de los 3 primeros números de Hill (1973). q1, Riqueza de especies; q2, especies abundantes; q3, especies muy abundantes).
| Clase | q1 | q2 | q3 |
| Encinar | 8 | 5.9 | 4.7 |
| Matorral subtropical | 17 | 8.2 | 4.7 |
| Matorral xerófilo | 15 | 9.7 | 7.1 |
| Mezquital | 6 | 2.7 | 1.8 |
| Pastizal inducido | 8 | 3.1 | 1.9 |
| Vegetación de arroyos | 11 | 7.9 | 5.9 |
Al analizar la precipitación estimada por el sensor satelital TRMM, se encontró que la precipitación fue similar en 2003, 2010 y 2018, con valores entre 668 y 768 mm. Estos 3 años presentaron la característica de encontrarse a una separación temporal que permite tener una buena representación de los posibles cambios ocurridos a partir del establecimiento de la RJN. Una vez determinado lo anterior, se solicitaron las imágenes de Landsat y PlanetScope. Las fechas para las cuales se solicitaron cada una de las imágenes se muestran en la tabla 3.
Una vez derivadas las variables necesarias, se realizaron las clasificaciones mediante entrenamiento supervisado de cada una de las 8 clases de cobertura de suelo presentes en el área de estudio. Como resultado del proceso de clasificación supervisada se generaron 4 mapas de cobertura de suelo para el área de la RJN. Para los mapas correspondientes a 2003, 2010 y 2018 de los sensores Landsat TM y OLI (resolución espacial de 30 m), se obtuvieron precisiones generales superiores a 0.75, con índices de Kappa iguales o superiores a 0.72 (tabla 4). Así mismo, se obtuvieron las precisiones por clase (precisión de usuario y productor), que variaron según el año de clasificación y clase de cobertura (tabla 4). En general, la representación visual de estos mapas muestra una concordancia en la forma en que se distribuyen las clases espacialmente, pero también es posible observar discrepancias en la representatividad de éstas al paso del tiempo (fig. 2).
Tabla 3
Selección de fechas de imágenes satelitales utilizadas para realizar las clasificaciones supervisadas.
| Año | Temporada | Fecha | Satélite | Sensor |
| 2003 | Seca | 01/05/2003 | Landsat 5 | Thematic Mapper (TM) |
| Húmeda | 06/09/2003 | |||
| 2010 | Seca | 20/05/2010 | Landsat 5 | Thematic Mapper (TM) |
| Húmeda | 25/09/2010 | |||
| 2018 | Seca | 08/04/2018 | Landsat 8 | Operational Land Imager (OLI) |
| Húmeda | 17/10/2018 | |||
| 2018 | Seca | 04 y 05/04/2018 | PlanetScope | Dove Classic |
| Húmeda | 28/09/2018 |
Tabla 4
Precisiones para cada una de las clasificaciones de cobertura elaboradas. La adición de las letras “PS” al año 2018, indica que corresponde a resultados derivados de la clasificación de imágenes de PlanetScope; el resto son derivados de las clasificaciones de Landsat.
| Clase | 2003 | 2010 | 2018 | 2018PS | ||||
| Usuario | Productor | Usuario | Productor | Usuario | Productor | Usuario | Productor | |
| Encinar | 0.72 | 0.80 | 0.83 | 0.82 | 0.90 | 0.89 | 0.97 | 0.98 |
| Matorral subtropical | 0.76 | 0.63 | 0.78 | 0.76 | 0.84 | 0.87 | 0.98 | 0.98 |
| Matorral xerófilo | 0.75 | 0.73 | 0.75 | 0.76 | 0.66 | 0.76 | 0.96 | 0.96 |
| Mezquital | 0.77 | 0.61 | 0.76 | 0.75 | 0.70 | 0.71 | 0.96 | 0.96 |
| Pastizal natural | 0.75 | 0.81 | 0.78 | 0.74 | 0.80 | 0.82 | 0.98 | 0.96 |
| Pastizal inducido | 0.68 | 0.81 | 0.83 | 0.85 | 0.90 | 0.81 | 0.96 | 0.98 |
| Suelo desnudo | 0.83 | 0.94 | 0.81 | 0.81 | 0.82 | 0.56 | 1.00 | 0.99 |
| Vegetación de arroyos | 0.88 | 0.87 | 0.87 | 0.92 | 0.94 | 0.88 | 0.99 | 0.99 |
| Precisión general | 0.75 | 0.80 | 0.81 | 0.97 | ||||
| Índice de Kappa | 0.72 | 0.77 | 0.78 | 0.96 |
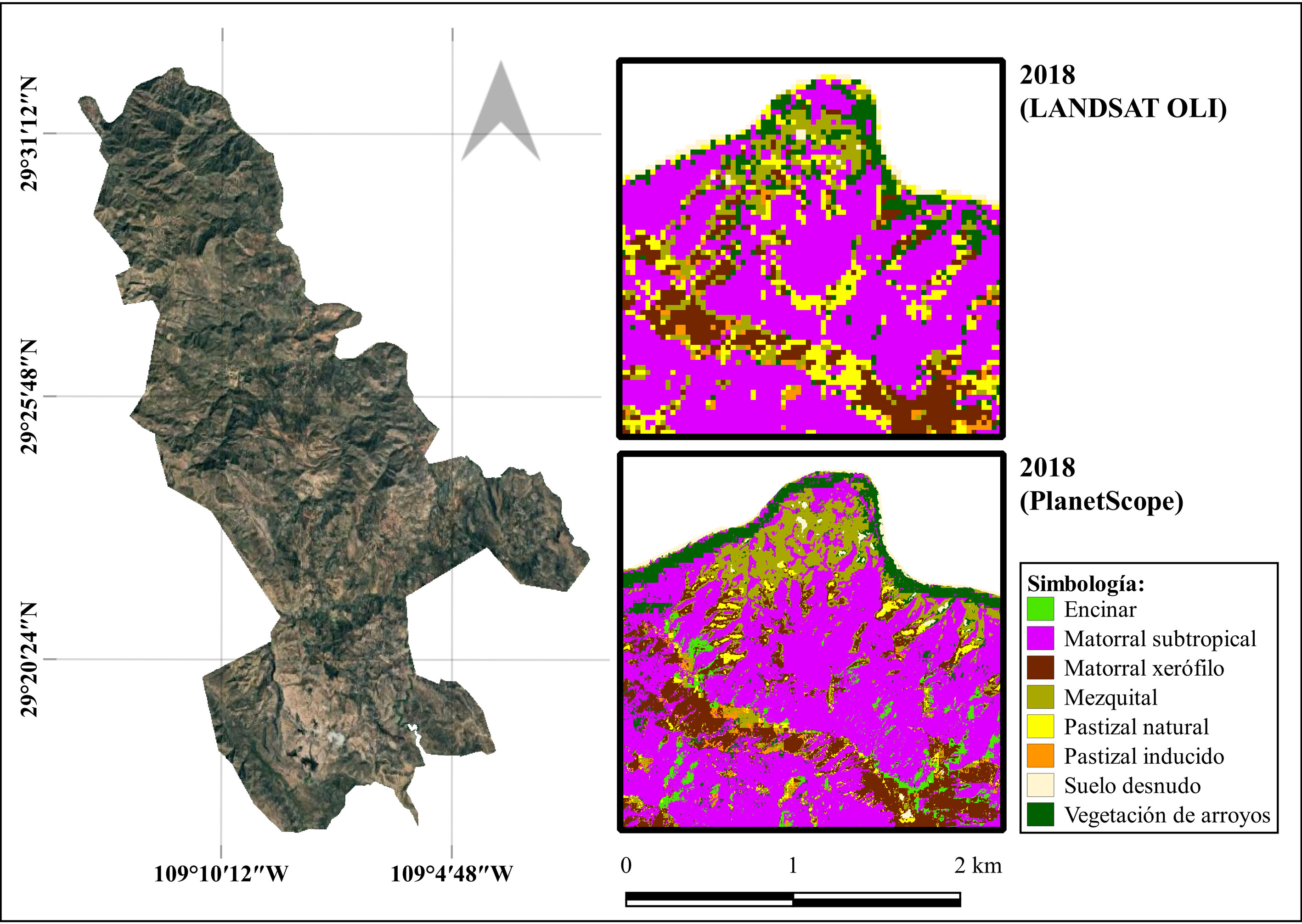
La precisión general e índice de Kappa, resultantes de la clasificación de cobertura con imágenes de PlanetScope para el año 2018 (resolución espacial de 3 m), fueron muy superiores a los obtenidos en la clasificación de Landsat OLI para el mismo año, así como para las clasificaciones de Landsat para 2003 y 2010, realizadas con una resolución espacial 10 veces menor (tabla 4). En la figura 3 es posible apreciar las diferencias visuales que existen entre ambas clasificaciones, al hacer un acercamiento en una porción del área de estudio.
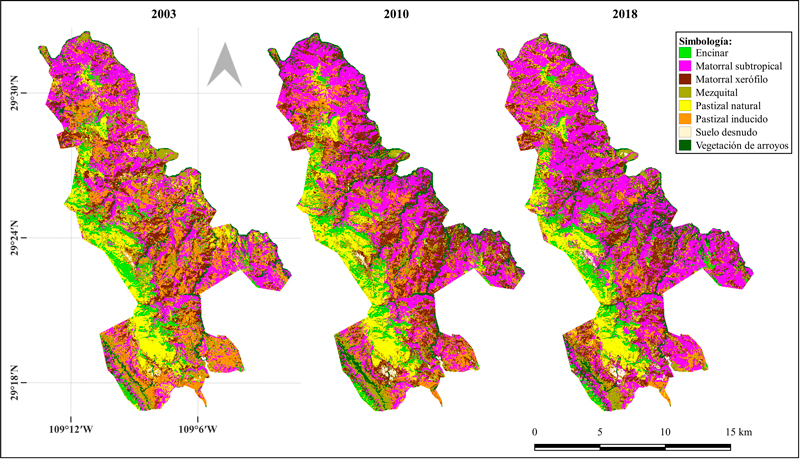
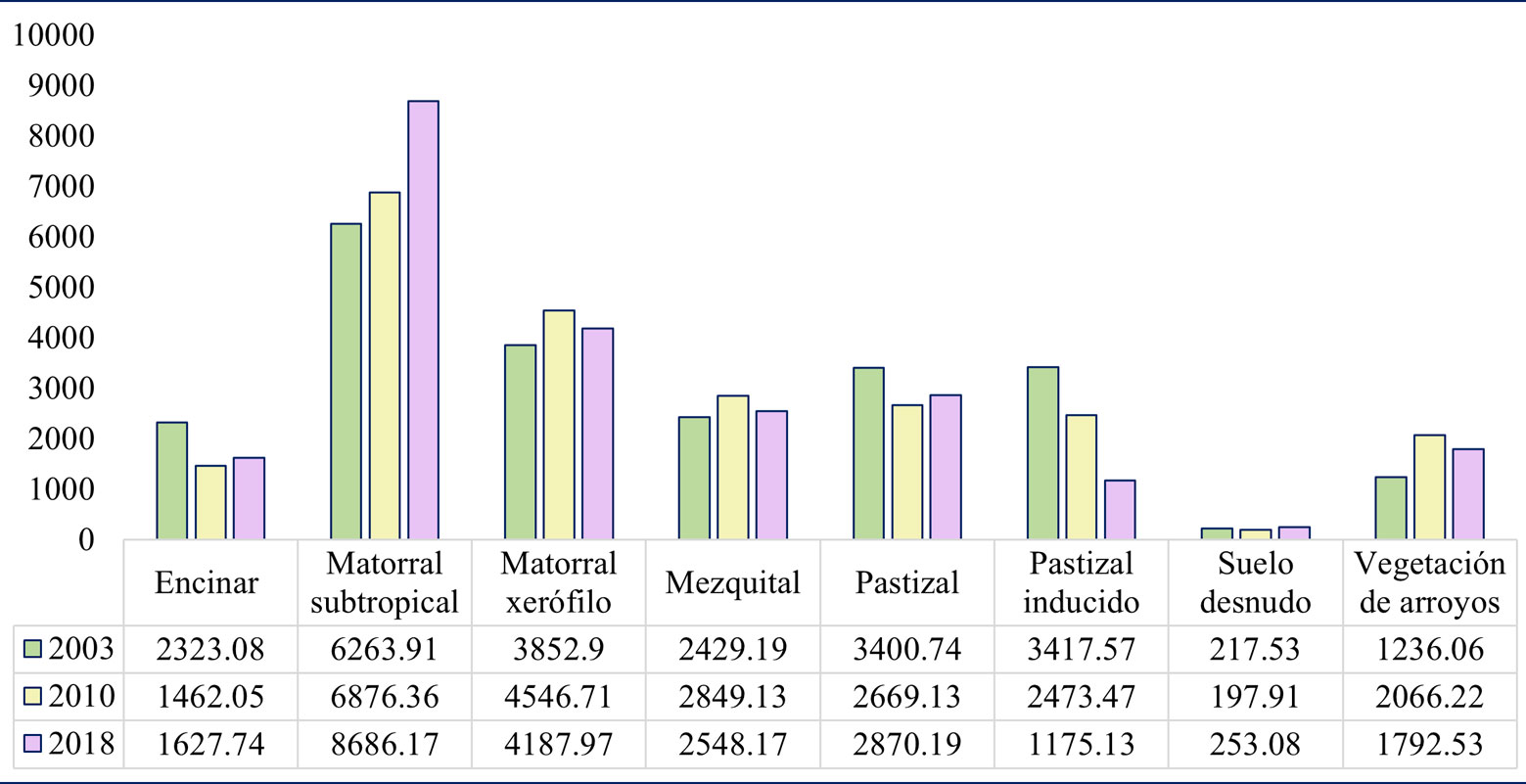
Los resultados de cobertura por clase a nivel paisaje para las clasificaciones elaboradas con imágenes de Landsat, muestran que todas las clases de cobertura presentan cambios entre el lapso de 2003 y 2018 (fig. 4). Las clases con mayores proporciones de área dentro de la RJN durante los 3 años analizados son el matorral subtropical y el matorral xerófilo, lo que se mantuvo en los 3 años analizados, mientras que las clases con menor representación son el suelo desnudo, el encinar y la vegetación de arroyos. El matorral subtropical aumentó su cobertura, pasando de 6,263.91 ha en el 2003 a 8,686.17 ha en el 2018. Por el contrario, las áreas con pastizales inducidos tendieron a disminuir, pasando de 3,417.57 a 1,175.13 ha (fig. 4).
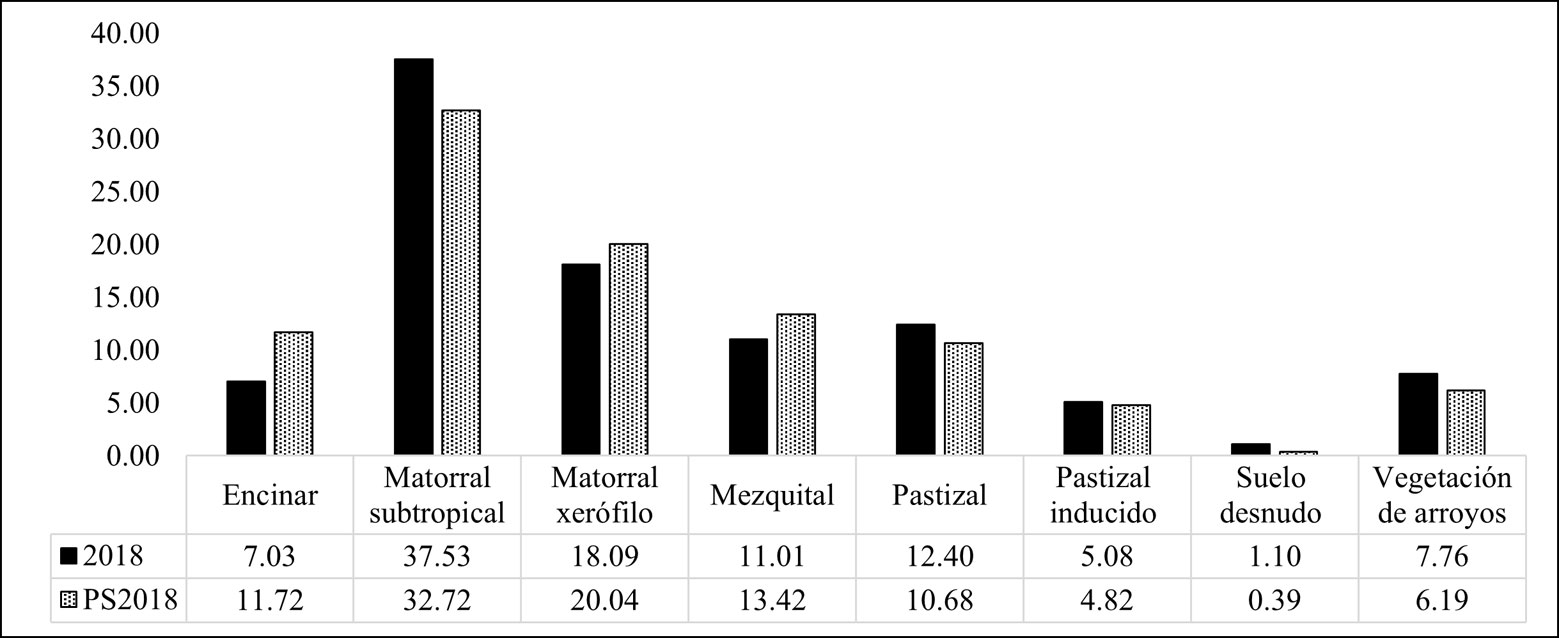
Por otro lado, las clasificaciones realizadas para el año 2018 derivadas de imágenes con diferente resolución espacial, muestran porcentajes similares de área por clase (fig. 5). Aunque para la mayoría de las clases de cobertura mantienen un porcentaje de área equivalente entre ambas resoluciones, las clases de matorral subtropical y encinar presentan mayores diferencias. En la primera de ellas, la clasificación con imágenes de alta resolución indicó una cobertura de área casi 5% menor que la resultante con imágenes de mediana resolución. Así mismo, el encinar mostró una diferencia de casi 5% de mayor cobertura al utilizar imágenes de alta resolución (fig. 5).
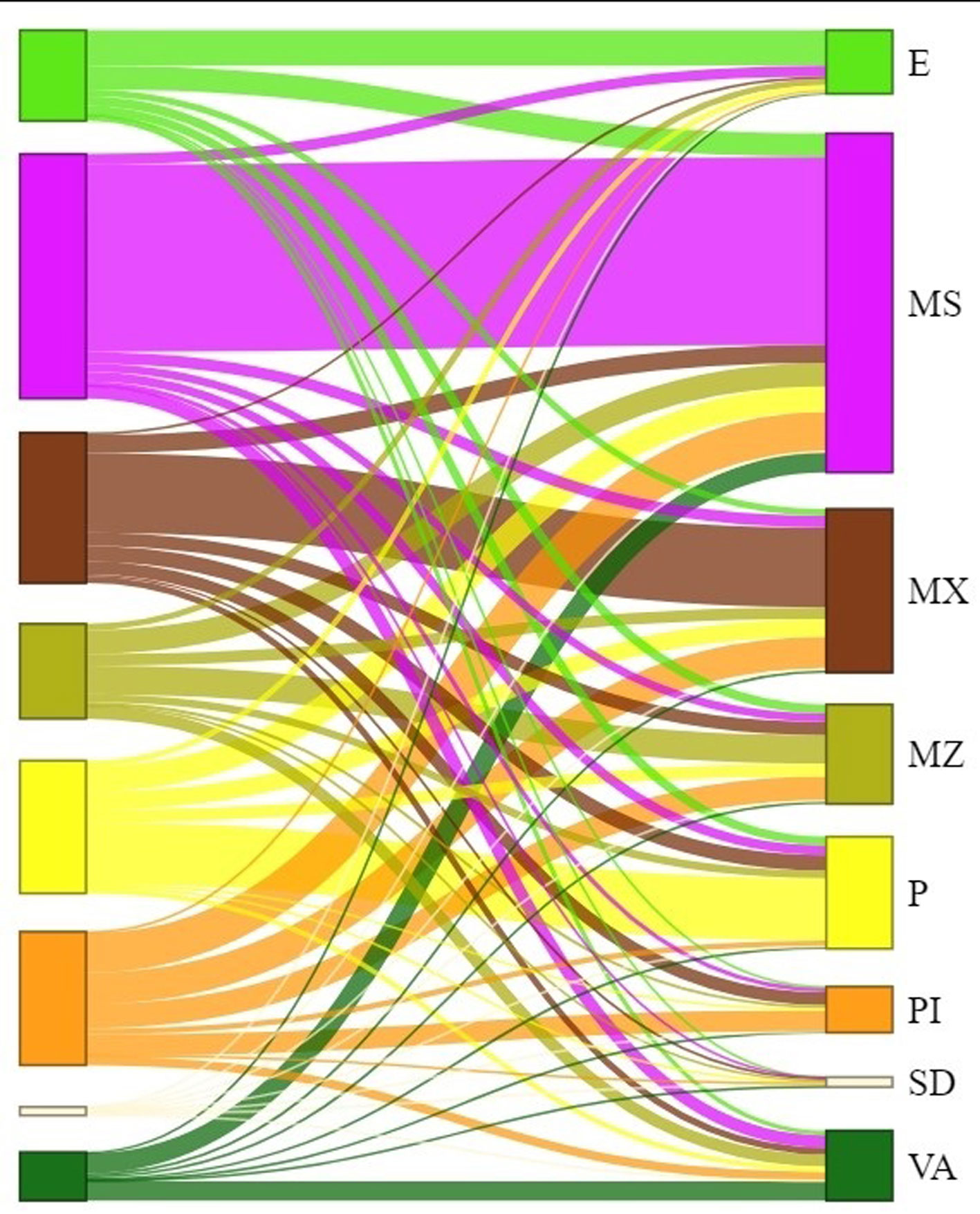
Tras realizar el análisis de cambio para las clasificaciones con imágenes de Landsat 2003 y 2018, fue posible detectar que todas las clases de cobertura se modificaron (tabla 5, fig. 6). Los cambios por ganancia y pérdida de cobertura se presentaron de manera diferencial entre clases (tabla 5). Las clases que fueron mayormente clasificadas como la misma en el 2018 fueron el matorral subtropical (76.74%) y el matorral xerófilo (52.96%). En el extremo opuesto, la clase de pastizal inducido fue la que menor cobertura mantuvo (15.91%; tabla 5).
En cuanto a los cambios de cobertura en el paisaje de una clase a otra, los resultados indican que, en mayor o menor medida, hubo correspondencia de área transformada entre todas ellas (tabla 5, fig. 6). El matorral subtropical fue la clase que menor porcentaje de área transformó a otras clases de cobertura, en el lapso del 2003 al 2018 (menos del 30%). En contraste, todas las demás clases de cobertura presentaron cambios en alrededor de 10% de su área o más, misma que se transformó en otras clases. El matorral subtropical fue la clase que más aporte de área obtuvo del resto de las clases presentes en el área de estudio. Esto último ocurrió, particularmente, con las comunidades de mezquital, pastizal inducido y suelo desnudo, que si bien también ganaron área proveniente de todas las demás clases (incluso de matorral subtropical), su mayor porcentaje de pérdida de área fue transformada en matorral subtropical.
Este mismo ejercicio también se realizó analizando primero los porcentajes de área obtenidos para la clasificación del año 2018, comparándolos posteriormente con los obtenidos para la de 2003. Al hacerlo de esta forma, las porciones de área identificadas en el 2018 como encinar, matorral subtropical y pastizal natural, en su mayoría coincidieron en su clasificación en 2003. En cambio, para el resto de las clases de cobertura, el área ocupada en 2018 fue, en su mayoría, catalogada como una clase diferente en el 2003. Lo anterior quiere decir que más de 50% del área identificada (por clase) en el 2018 como matorral xerófilo, mezquital, pastizal inducido, suelo desnudo y vegetación de arroyos, fue asignada a otras clases de cobertura en 2003. Las transformaciones de cobertura en el paisaje mencionadas se ejemplifican visualmente en la figura 6.
Discusión
La cantidad de superficie representada por cada clase de vegetación muestra patrones y tendencias que son relevantes en la gestión de un área para la conservación. Si bien todas las comunidades vegetales analizadas en este trabajo presentaron cambios de cobertura a nivel paisaje, solo 2 mostraron tendencias claras de disminución y aumento: el pastizal inducido y el matorral subtropical, respectivamente. Lo anterior puede atribuirse a resultados derivados del tipo de gestión en el área de estudio.
Tabla 5
Extensión en hectáreas y porcentaje del área total calculados para cada clase en el lapso 2003-2018.
| Clase 2003 | Clase 2018 | Área (ha) | Cobertura a nivel paisaje (%) |
| Encinar | Encinar | 914.49 | 39.37 |
| Matorral subtropical | 603.54 | 25.98 | |
| Matorral xerófilo | 177.3 | 7.63 | |
| Mezquital | 239.22 | 10.3 | |
| Pastizal natural | 230.58 | 9.93 | |
| Pastizal inducido | 29.7 | 1.28 | |
| Suelo desnudo | 10.71 | 0.46 | |
| Vegetación de arroyos | 117.54 | 5.06 | |
| Matorral subtropical | Encinar | 269.73 | 4.31 |
| Matorral subtropical | 4,806.99 | 76.74 | |
| Matorral xerófilo | 297.27 | 4.75 | |
| Mezquital | 206.1 | 3.29 | |
| Pastizal natural | 240.75 | 3.84 | |
| Pastizal inducido | 115.11 | 1.84 | |
| Suelo desnudo | 4.14 | 0.07 | |
| Vegetación de arroyos | 323.82 | 5.17 | |
| Matorral xerófilo | Encinar | 57.51 | 1.49 |
| Matorral subtropical | 463.86 | 12.04 | |
| Matorral xerófilo | 2,040.57 | 52.96 | |
| Mezquital | 339.21 | 8.8 | |
| Pastizal natural | 396.18 | 10.28 | |
| Pastizal inducido | 329.04 | 8.54 | |
| Suelo desnudo | 59.13 | 1.53 | |
| Vegetación de arroyos | 167.4 | 4.34 | |
| Mezquital | Encinar | 165.69 | 6.82 |
| Matorral subtropical | 605.7 | 24.93 | |
| Matorral xerófilo | 299.43 | 12.33 | |
| Mezquital | 735.84 | 30.29 | |
| Pastizal natural | 200.79 | 8.27 | |
| Pastizal inducido | 80.01 | 3.29 | |
| Suelo desnudo | 25.2 | 1.04 | |
| Vegetación de arroyos | 316.53 | 13.03 | |
| Pastizal | Encinar | 146.61 | 4.31 |
| Matorral subtropical | 657.54 | 19.34 | |
| Matorral xerófilo | 457.11 | 13.44 | |
| Mezquital | 318.24 | 9.36 | |
| Pastizal natural | 1,589.85 | 46.75 | |
| Pastizal inducido | 48.51 | 1.43 | |
| Tabla 5. Continúa | |||
| Clase 2003 | Clase 2018 | Área (ha) | Cobertura a nivel paisaje (%) |
| Suelo desnudo | 47.7 | 1.4 | |
| Vegetación de arroyos | 135.18 | 3.98 | |
| Pastizal inducido | Encinar | 42.66 | 1.25 |
| Matorral subtropical | 1,008.99 | 29.52 | |
| Matorral xerófilo | 808.29 | 23.65 | |
| Mezquital | 623.07 | 18.23 | |
| Pastizal natural | 144.45 | 4.23 | |
| Pastizal inducido | 543.87 | 15.91 | |
| Suelo desnudo | 18.99 | 0.56 | |
| Vegetación de arroyos | 227.25 | 6.65 | |
| Suelo desnudo | Encinar | 1.35 | 0.62 |
| Matorral subtropical | 23.04 | 10.59 | |
| Matorral xerófilo | 51.93 | 23.87 | |
| Mezquital | 27.63 | 12.7 | |
| Pastizal natural | 22.23 | 10.22 | |
| Pastizal inducido | 11.97 | 5.5 | |
| Suelo desnudo | 71.19 | 32.73 | |
| Vegetación de arroyos | 8.19 | 3.76 | |
| Vegetación de arroyos | Encinar | 29.7 | 2.4 |
| Matorral subtropical | 516.51 | 41.79 | |
| Matorral xerófilo | 56.07 | 4.54 | |
| Mezquital | 58.86 | 4.76 | |
| Pastizal natural | 45.36 | 3.67 | |
| Pastizal inducido | 16.92 | 1.37 | |
| Suelo desnudo | 16.02 | 1.3 | |
| Vegetación de arroyos | 496.62 | 40.18 |
El pastizal inducido destacó por su escasa cobertura leñosa, con la situación inversa del estrato herbáceo, ampliamente dominado por la gramínea exótica, el zacate buffel. Este pasto fue introducido en el noroeste de México en la década de los 50 y establecido de forma masiva a partir de 1970, con la finalidad de incrementar la producción forrajera (Castellanos-Villegas et al., 2002; Ibarra-Flores et al., 2009). Tras su establecimiento en la región, se han realizado investigaciones enfocadas en determinar sus efectos sobre las comunidades naturales locales. Diversos estudios han advertido de su potencial dañino como especie invasora (Castellanos-Villegas et al., 2002; Jackson, 2005; Choza-Farías, 2021), lo que puede traducirse en posibles amenazas para la biodiversidad nativa, al verse mermada la capacidad de resiliencia de los ecosistemas en los cuales se establece (Morales-Romero et al., 2019; Siller-Clavel, 2022). Estos efectos se han documentado en estudios que indican una menor riqueza de especies (Olsson et al., 2012) y cubierta nativa vegetal (Abella et al., 2012; De la Barrera, 2008). La tendencia notoria hacia una menor superficie de área ocupada por esta comunidad vegetal, a escala de paisaje, es un aspecto positivo para los fines de conservación que se persiguen en la RJN y, posiblemente, sea una consecuencia de las medidas tomadas por los responsables en su gestión, como la remoción activa de zacate buffel (Cenchrus ciliaris) y el fomento a la regeneración natural de sitios con un amplio legado de uso de suelo con ganadería extensiva.
Por otro lado, el matorral subtropical, además de presentar una tendencia de aumento, fue la comunidad vegetal con mayor representatividad de área en el paisaje durante los 3 años analizados. Esta clase también presentó la mayor riqueza de especies, con baja dominancia y un alto número de especies presentes exclusivamente en esa comunidad vegetal, lo que podría aumentar la estabilidad, productividad y resiliencia a escala de paisaje (Isbell et al., 2015; Oehri et al., 2020). De esta forma, la tendencia de transformación hacia matorral subtropical a nivel paisaje, específicamente cuando se trata, en gran medida, de pastizal inducido, no parece negativa. Sin embargo, esto no debe tomarse como un argumento definitivo, ya que las transformaciones en la composición de la vegetación repercuten en el funcionamiento de las comunidades, por lo que resulta necesario estudiar con mayor profundidad los posibles efectos de estos cambios a nivel paisaje. Es relevante destacar que Rzedowski (1978) afirmó que las comunidades vegetales que describen Rzedowski y McVaugh (1966), bajo el nombre de matorral subtropical, posiblemente representen, al menos parcialmente, fases sucesionales estables del bosque tropical caducifolio. Los resultados del presente estudio permiten relacionar la naturaleza sucesional de este tipo de vegetación en la reserva estudiada.
El huizache (Vachellia campechiana) y el mezquite (Prosopis velutina) estuvieron presentes en la mayoría de las comunidades vegetales muestreadas, donde tendieron a dominar el estrato leñoso. Lo anterior puede deberse a que ambas son especies que tienen menos requerimientos para su establecimiento, lo que las vuelve más tolerantes a variaciones sutiles (grado de exposición solar, tipo de suelo, temperatura, humedad), que ocurren entre hábitats de diferentes comunidades vegetales (Miller et al., 2015). Por el contrario, aquellas especies presentes solo en una comunidad vegetal podrían presentar requerimientos más específicos. A largo plazo, las primeras podrían tener ventajas al tolerar mejor algunas condiciones adversas como la sequía y el aumento de la temperatura, mismas que se pronostican en un escenario de cambio climático para el desierto de Sonora en el futuro (Caracciolo et al., 2016; Piccinelli et al., 2020; Seager et al., 2007).
En cuanto al desempeño de las clasificaciones, las realizadas con imágenes de Landsat obtuvieron precisiones generales e índices de Kappa que reflejan un buen desempeño al discriminar bien entre las clases de cobertura presentes en la RJN (Landis y Koch, 1977). Las imágenes de Landsat han sido ampliamente recomendadas para el monitoreo de la cobertura terrestre a gran escala, debido a su accesibilidad continua desde 1972 (Ma et al., 2017; Willis, 2015). Además, forma parte de los satélites y sensores más utilizados con fines de monitoreo de áreas protegidas (Duan et al., 2020). Sin embargo, el uso de imágenes provenientes de sensores de mayor resolución espacial se está popularizando cada vez más (Belward y Skøien, 2015). Aunque existe una correlación positiva entre el tamaño de las áreas de estudio y las resoluciones espaciales utilizadas, en la mayoría de las ocasiones se ha visto que las imágenes de alta resolución espacial son ventajosas para obtener mayores precisiones de clasificación y, por consiguiente, una representación más cercana a su distribución real en el paisaje (Ma et al., 2017).
En este trabajo, la precisión general e índice de Kappa resultantes para la clasificación de PlanetScope, con una resolución 10 veces más fina que la clasificación resultante de imágenes de Landsat, tuvo muy buen desempeño (Landis y Koch, 1977). Además, al hacer comparaciones entre clasificaciones generadas con imágenes de diferentes resoluciones, pero para el mismo año y utilizando el mismo algoritmo y puntos de entrenamiento, el incremento en la precisión se explica, esencialmente, por la diferencia en la resolución espacial y el aumento en capacidad que esto supone para detectar sutiles variaciones en el espacio físico. Esto sugiere que la utilización de alta resolución en ejercicios de clasificación supervisada en paisajes heterogéneos tiene un desempeño muy superior, en comparación con resoluciones más bajas (en este caso de 30 m). Sin embargo, algunos estudios han demostrado que una alta resolución no significa, necesariamente, una alta precisión, ya que las clasificaciones derivadas de estas imágenes suelen tener una mayor variación intraclase, lo que puede complicar la diferenciación entre las mismas (Lv et al., 2022). Particularmente, en el caso de este estudio, es probable que una buena colecta de puntos de entrenamiento (representativa al interior de cada clase y espectralmente homogénea) beneficiara el uso de una alta resolución espacial.
Finalmente, es importante tener en cuenta que los resultados de precisión obtenidos en las clasificaciones no deben tomarse como definitivos y estáticos, ya que estos mapas se vuelven obsoletos y, eventualmente, dejan de representar los procesos que actúan en el paisaje (Robinson et al., 2019). Sin embargo, en este estudio permitieron valorar los efectos de un programa de manejo en un periodo específico (2003-2018), mostrando un resultado aparentemente positivo. Por último, se recomienda ampliamente la colecta de información directamente del sitio de estudio, en combinación con el uso de imágenes satelitales con resoluciones espaciales medias y finas, ya que esta práctica brinda una mejor perspectiva de los vínculos existentes entre los procesos ecológicos que se presentan en el paisaje a diferentes escalas.
Agradecimientos
SDP agradece al Consejo Nacional de Ciencia y Tecnología, por el apoyo brindado en forma de una beca de maestría. JRRL quiere agradecer a Northern Jaguar Project, por el patrocinio y apoyo al proyecto registrado en la Universidad de Sonora como USO313007014. Así mismo, se agradece al personal de la Reserva Jaguar del Norte, por su apoyo en colecta de datos en campo y la información geoespacial proporcionada. Se agradece también a los colaboradores, revisores anónimos y al editor asociado de la revista, Guillermo Ibarra Manríquez, por sus valiosas aportaciones y revisiones. Por último, a la Universidad de Sonora y al Posgrado en Biociencias por su espacio y medios de trabajo.
Referencias
Abella, S. R., Chiquoine, L. P. y Backer, D. M. (2012). Ecological characteristics of sites invaded by buffelgrass (Pennisetum ciliare). Invasive Plant Science and Management, 59, 443–453. https://doi.org/10.1614/IPSM-D-12-00012.1
Alhaddad, B. I., Burns, M. C. y Cladera, J. R. (2007). Texture Analysis for correcting and detecting classification structures in urban land uses; Metropolitan area case study – Spain. Urban Remote Sensing Joint Event. París, Francia. https://doi.org/10.1109/URS.2007.371811
Anderson, J. F., Hardy, E. E., Roach, J. T. y Witmer, R. E. (1976). A land use and land cover classification system for use with remote sensor data. Washington, USA: U.S. Geological Survey. https://doi.org/10.3133/pp964
Arneth, A. (2015). Uncertain future for vegetation cover. Nature, 524, 44–45. https://doi.org/10.1038/524044a
Arriaga, L., Espinoza, J. M., Aguilar, C., Martínez, E., Gómez, L. y Loa, E. (2000). Regiones terrestres prioritarias de México. Ciudad de México: Conabio.
Arroyo-Rodríguez, V., Moreno, C. y Galán-Acedo, C. (2017). La ecología del paisaje en México: logros, desafíos y oportunidades en las ciencias biológicas. Revista Mexicana de Biodiversidad, 8, 42–51. https://doi.org/10.1016/j.rmb.
2017.10.004
Asner, G. P., Keller, M., Pereira, R. y Zweede, J. C. (2002). Remote sensing of selective logging in Amazonia: assessing limitations based on detailed field observations, Landsat ETM+, and textural analysis. Remote Sensing of Environment, 80, 483–496. https://doi.org/10.1016/S0034-4257(01)00326-1
Belward, A. S. y Skøien, J. O. (2015). Who launched what, when and why; trends in global land-cover observation capacity from civilian earth observation satellites. ISPRS Journal of Photogrammetry and Remote Sensing, 103, 115–128. https://doi.org/10.1016/j.isprsjprs.2014.03.009
Bertzky, B., Corrigan, C., Kemsey, J., Kenney, S., Ravilious, C., Besançon, C. et al. (2012). Protected Planet Report 2012: tracking progress towards global targets for protected areas. Cambridge, UK: United Nations Environment Programme.
Bezaury-Creel, J. E. y Gutiérrez-Carbonell, D. (2009). Áreas naturales protegidas y desarrollo social en México. En J. Sarukhán, P. Koleff, J. Carabias, J. Soberón, R. Dirzo, J. Llorente-Bousquets et al. (Eds.). Capital natural de México vol. II. Estado de conservación y tendencias de cambio (pp. 385–431). Ciudad de México: Conabio.
Borda-de Água, L. (2019). The importance of scaling in biodiversity. En E. Casetta, J. Marques-da Silva y D. Vecchi (Eds.), From assessing to conserving biodiversity, conceptual and practical challenges (pp. 107–122). Cham, Switzerland: Springer. https://doi.org/10.1007/978-3-030-10991-2_5
Braun-Blanquet, J. (1932). Plant Sociology, the study of plant communities. New York: McGraw-Hill Book Company.
Breiman, L., Friedman, J. H., Olshen, R. A. y Stone, C. G. (1984). Classification and regression trees. California: Taylor & Francis. https://doi.org/10.1201/9781315139470
Briones, O., Burquez, A., Martínez-Yrízar, A., Pavón, N. P. y Perroni, Y. (2018). Biomasa y productividad en las zonas áridas mexicanas. Madera y Bosques, 24, e2401898. https://doi.org/10.21829/myb.2018.2401898
Canfield, R. H. (1941). Application of the line interception method in sampling range vegetation. Journal of Forestry, 39, 388–394.
Caracciolo D., Istanbulluoglu, E., Valerio-Noto, L. y Collins, S. L. (2016). Mechanisms of shrub encroachment into Northern Chihuahuan Desert grasslands and impacts of climate change investigated using a cellular automata model. Advances in Water Resources, 91, 46–62. https://doi.org/10.1016/j.advwa
tres.2016.03.002
Castellanos-Villegas, A. Yanes, G. y Valdez-Zamudio, D. (2002). Drougth-tolerant exotic buffelgrass and desertification. En B. Tellman (Ed.), Weeds across borders: Proceedings of a North American Conference Held in Tucson, Arizona (pp. 99–112), Arizona: Arizona-Sonora Desert Museum.
Choza-Farías, S., Romo-León, J. R. y Castellanos-Villegas, A. E. (2021). Análisis de la respuesta productiva ante la variabilidad climática en tipos de vegetación exótica y nativa del Desierto Sonorense. Revista Chapingo, 20, e2021203. https://doi.org/10.5154/r.rchsza.2021.20.3
Coetzee, B. W. T. (2017). Evaluating the ecological performance of protected areas. Biodiversity and Conservation, 26, 231–236. https://doi.org/10.1007/s10531-016-1235-2
Cohen, J. (1960). A coefficient of agreement for nominal scales. Educational and Psychological Measurement, 20, 37–46. https://doi.org/10.1177/001316446002000104
Collins, J. B. y Woodcock, C. E. (1996). An assessment of several linear change detection techniques for mapping forest mortality using multitemporal landsat TM data. Remote Sensing of Environment, 56, 66–77. https://doi.org/10.1016/0034-4257(95)00233-2
Conaza (Comisión Nacional de las Zonas Áridas). (1994). Mezquite (Prosopis spp.). Cultivo alternativo para las zonas áridas y semiáridas de México. Ciudad de México: Conaza.
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2019). Resiliencia Áreas Naturales Protegidas: soluciones naturales a retos globales. Ciudad de México: Conanp – PNUD en México y Proyecto Resiliencia.
De la Barrera, E. (2008). Recent invasion of buffel grass (Cenchrus ciliaris) of a natural protected area from the southern Sonoran Desert. Revista Mexicana de Biodiversidad, 79, 385–392. https://doi.org/10.22201/ib.20078706e.2008.002.562
Duan, P., Wang, Y. y Peng, Y. (2020). Remote sensing applications in monitoring of protected areas: A Bibliometric Analysis. Remote Sensing, 12, 772. https://doi.org/10.3390/rs12050772
Durand, L. y Jiménez, J. (2010). Sobre áreas naturales protegidas y la construcción de no-lugares, Notas para México. Revista Lider, 16, 59–72.
Duveiller, G., Hooker, J. y Cescatti, A. (2018). A dataset mapping the potential biophysical effects of vegetation cover change. Scientific Data, 5, 180014. https://doi.org/10.1038/sdata.2018.14
Ferreira, S., Daemane, M., Deacon, A., Sithole, H. y Bezuidenhout, H. (2013). Efficient evaluation of biodiversity concerns in protected areas. International Journal of Biodiversity, 2013, 298968. https://doi.org/10.1155/2013/298968
Flores-Olvera, H. (2011). Las zonas áridas y semiáridas de México, las menos exploradas. Ciudad de México: Boletín UNAM-DGCS-763.
Freitas, J. R. y Mantovania, W. (2017). An overview of the applicability of functional diversity in Biological Conservation. Brazilian Journal of Ecology, 78, 517–524. https://doi.org/10.1590/1519-6984.09416
Fuente, B., Weynants, M., Bertzky, B., Giacomo, D., Mandrici, A., Bendito, E. et al. (2019). Land productivity dynamics in and around protected areas globally from 1999 to 2013. Plos One, 15, e0224958. https://doi.org/10.1371/journal.pone.0224958
Fung, T. y LeDrew, E. (1987). Application of principal components analysis to change detection. Photogrammetric Engineering and Remote Sensing, 53, 1649–1658.
Gaston, K., Jackson, S., Cantú-Salazar, L. y Cruz-Piñón, G. (2008). The ecological performance of Protected Areas. Annual Review of Ecology Evolution and Systematics, 39, 93–113. https://doi.org/10.1146/annurev.ecolsys.39.110707.173529
Gross, J., Goetz, S. y Cihlar, J. (2009). Application of remote sensing to parks and protected area monitoring: Introduction to the special issue. Remote Sensing of Environment, 113, 1343–1345. https://doi.org/10.1016/j.rse.2008.12.013
Guerra, C. A., Rosa, I. M. D. y Pereira, H. M. (2019). Change versus stability: are protected areas particularly pressured by global land cover change? Landscape Ecology, 34, 2779–2790. https://doi.org/10.1007/s10980-019-00918-4
Henle, K., Potts, S., Kunin, W., Matsinos, Y., Simila, J., Pantis, J. et al. (2014). Scaling in Ecology and Biodiversity Conservation. Sofía, Bulgaria: Pensoft Publishers. https://doi.org/10.3897/ab.e1169
Hernández-Moreno, M. M., Téllez-Valdés, O., Martínez-Meyer, E., Islas-Saldaña, L. A., Salazar-Rojas, V. M. y Macías-Cuéllar, H. (2021). Distribución de la cobertura vegetal y del uso del terreno del municipio de Zapotitlán, Puebla, México. Revista Mexicana de Biodiversidad, 92, e923649. https://doi.org/10.22201/ib.20078706e.2021.92.3649
Hill, M. O. (1973). Diversity and evenness: a unifying notation and its consequences. Ecology, 54, 427–432. https://doi.org/10.2307/1934352
Houborg, R., Fisher, J. B. y Skidmore, A. K. (2015). Advances in remote sensing of vegetation function and traits. International Journal of Applied Earth Observation and Geoinformation, 43, 1–6. https://doi.org/10.1016/j.jag.2015.06.001
Huete, A. R. (1988). A soil-adjusted vegetation index (SAVI). Remote Sensing of Environment, 25, 295–309. https://doi.org/10.1016/0034-4257(88)90106-x
Huete, A., Didan, K., Miura, T., Rodriguez, E. P., Gao, X. y Ferreira, L. G. (2002). Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sensing of Environment, 83, 195–213. https://doi.org/10.1016/s0034-4257(02)00096-2
Ibarra-Flores, F. A., Martín-Rivera, M. H., Denogean-Ballesteros, F. G. y Aguirre-Murrieta, R. (2009). Buffelgrass, cattle, and the Sonoran Desert. En T. R. Van Devender, F. J. Espinosa-García, B. L. Harper-Lore y T. Hubbard (Eds.), Invasive plants on the move: controlling them in North America (pp. 375–381). Tucson, AZ: Arizona-Sonora Desert Museum.
INEGI (Instituto Nacional de Estadística y Geografía). (2014). Guía para la interpretación de cartografía: uso del suelo y vegetación: escala 1:250,000: serie V. Ciudad de México: INEGI.
Isbell, F., Craven, D., Connolly, J., Loreau, M., Schmid, B., Beierkuhnlein, C. et al. (2015). Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature, 526, 574–577. https://doi.org/10.1038/nature15374
Jackson, J. (2005). Is there a relationship between herbaceous species richness and buffel grass (Cenchrus ciliaris)? Austral Ecology, 30, 505–517. https://doi.org/10.1111/j.1442-9993.2005.01465.x
Jantz, S. M., Barker, B., Brooks, T. M., Chini, L. P., Huang, Q., Moore, R. M. et al. (2015). Future habitat loss and extinctions driven by land-use change in biodiversity hotspots under four scenarios of climate-change mitigation. Conservation Biology, 29, 1122–1131. https://doi.org/10.1111/cobi.12549
Jensen, J. R. (2005). Introductory digital image processing: a remote sensing perspective. New Jersey, USA: Pearson.
Kamwi, J., Kätsch, C., Graz, F., Chirwa, P. y Manda, S. (2017). Trends in land use and land cover change in the protected and communal areas of the Zambezi Region, Namibia. Environmental Monitoring and Assessment, 189, 242. https://doi.org/10.1007/s10661-017-5934-2
Koch, M., Schröder, B., Günther, A., Albrecht, K., Pivarci, R., Jurasinski, G. et al. (2017). Taxonomic and functional vegetation changes after shifting management from traditional herding to fenced grazing in temperate grassland communities. Applied Vegetation Science, 20, 259–270. https://doi.org/10.1111/avsc.12287
Kumar, N., Yamaç, S. y Murugan, A. (2015). Applications of remote sensing and GIS in natural resource management. The Andaman Science Association, 20, 1–6.
Kumar-Verma, A. (2016). Biodiversity: its different levels and values. International Journal on Environmental Sciences, 7, 143–145.
Landis, J. R. y Koch, G. G. (1977). The measurement of observer agreement for categorical data. Biometrics, 33, 159−174. https://doi.org/10.2307/2529310
Lin-Quintana, L. E. (2017). Análisis de la disponibilidad de agua de lluvia en el Estado de Sonora (Tesis). Universidad Nacional Autónoma de México, Ciudad de México.
Lv, Z., Liu, T., Benediktsson, J. A. y Falco, N. (2022). Land cover change detection techniques: very-high-resolution optical images: a review. IEEE Geoscience and Remote Sensing Magazine, 10, 44–63. https://doi.org/10.1109/MGRS.2021.3088865
Ma, L., Li, M., Ma, X., Cheng, L., Du, P. y Liu, Y. (2017). A review of supervised object-based land-cover image classification. ISPRS Journal of Photogrammetry and Remote Sensing, 130, 277–293. https://doi.org/10.1016/j.isprsjprs.2017.06.001
Maass, J. M., Jardel, E. J., Martínez-Yrízar, A., Calderón-Aguilera, L. E., Herrera, J. y Castillo, A. (2010). Las áreas naturales protegidas y la investigación ecológica de largo plazo en México. Ecosistemas, 19, 69–83.
Maestre, F. T., Benito, B. M., Berdugo, M., Concostrina-Zubiri, L., Delgado-Baquerizo, M., Eldrige, D. J. et al. (2021). Biogeography of global drylands. New Phytologist, 231, 540–558. https://doi.org/10.1111/nph.17395
McDonald, S., Reid, N., Smith, R., Waters, C., Hunter, J. y Rader, R. (2019). Comparison of biodiversity and ground cover between a commercial rotationally grazed property and an adjacent nature reserve in semi-arid rangeland. Austral Ecology, 45, 60–69. https://doi.org/10.1111/aec.12829
Miller, J. E. D., Damschen, E. I., Harrison, S. P. y Grace, J. B. (2015). Landscape structure affects specialists but not generalists in naturally fragmented grasslands. Ecology, 96, 3323–3331. https://doi.org/10.1890/15-0245.1
Morales-Romero, D., López-García, H., Martínez-Rodríguez, J. y Molina-Freaner, F. (2019). Documenting a plant invasion: the influence of land use on buffelgrass invasion along roadsides in Sonora, Mexico. Journal of Arid Environments, 164, 53–59. https://doi.org/10.1016/j.jaridenv.2019.01.012
Northern Jaguar Project. (2022). Northern Jaguar Reserve. Recuperado el 20 noviembre, 2014 de: https://www.northernjaguarproject.org/northern-jaguar-reserve/
Oehri, J., Schmid, B., Schaepman-Strub, G. y Niklaus, P. A. (2020). Terrestrial land-cover type richness is positively linked to landscape-level functioning. Nature Communications, 11, 154. https://doi.org/10.1038/s41467-019-14002-7
Olsson, A. D., Betancourt, J. L., Crimmins, M. A. y Marsh, S. E. (2012). Constancy of local spread rates for buffel grass (Pennisetum ciliare L.) in the Arizona Upland of the Sonoran Desert. Journal of Arid Environments, 87, 136–143. https://doi.org/10.1016/j.jaridenv.2012.06.005
Piccinelli, S., Brusa, G. y Cannone, N. (2020). Climate warming accelerates forest encroachment triggered by land use change: A case study in the Italian Prealps (Triangolo Lariano, Italy). Catena, 195, 104870. https://doi.org/10.1016/j.catena.2020.104870
Planet Team. (2017). Planet application program interface: in space for Life on Earth. San Francisco, CA. https://api.planet.com/
Poiani, K. A., Richter, B. D., Anderson, M. G. y Richter, H. E. (2000). Biodiversity conservation at multiple scales: functional sites, landscapes, and networks. Bioscience, 50, 33–146, https://doi.org/10.1641/0006-3568(2000)050[0133:BCAMSF]2.3.CO;2
R Core Team. (2021). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ramachandra, T. V., Setturu, B. y Gupta, N. (2018). Modelling landscape dynamics with LST in protected areas of Western Ghats, Karnataka. Journal of Environmental Management, 15, 1253–1262. https://doi.org/10.1016/j.jenvman.2017.08.001
Ramírez-Arce, D. G., Ochoa-Ochoa, L. M. y Lira-Noriega, A. (2022). Effect of landscape composition and configuration on biodiversity at multiple scales: a case study with amphibians from Sierra Madre del Sur, Oaxaca, Mexico. Landscape Ecology, 37, 1973–1986. https://doi.org/10.1007/s10980-022-01479-9
Rimal, B., Sharma, R., Kunwar, R., Keshtkar, H., Stork, N. E., Rijal, S. et al. (2019). Effects of land use and land cover change on ecosystem services in the Koshi River Basin, Eastern Nepal. Ecosystem Services, 38, 100963. https://doi.org/10.1016/j.ecoser.2019.100963
Robinson, C., Hou, L., Malkin, K., Soobitsky, R., Czawlytko, J., Dilkina, B. et al. (2019). Large scale high-resolution land cover mapping with multi-resolution data. IEEE/CVF Conference on Computer Vision and Pattern Recognition (CVPR) (pp. 12718–12727). Long Beach, CA, USA. https://doi.org/10.1109/CVPR.2019.01301
Romo-León, J. R., van Leeuwen, W. J. D. y Castellanos-Villegas, A. (2014). Using remote sensing tools to assess land use transitions in unsustainable arid agro-ecosystems. Journal of Arid Environments, 106, 27–35. https://doi.org/10.1016/j.jaridenv.2014.03.002
Rzedowski, J. (1978). Vegetación de México. Ciudad de México: Limusa.
Rzedowski, J. y McVaugh, R. (1966). La vegetación de Nueva Galicia. BioStor, 9, 1–123.
Seager R., Ting, M., Held, I., Kushnir, Y., Lu, J., Vecchi, G. et al. (2007). Model projections of an imminent transition to a more arid climate in southwestern North America. Science, 316, 1181–1184. https://doi.org/10.1126/science.1139601
Sharma, R., Eklund, J., Barnes, M., Geldman, J., Schleicher, J., Pressey, R. L. et al. (2020). The impact of terrestrial protected areas on vegetation extent and condition: a systematic review protocol. Environmental Evidence, 9, 8. https://doi.org/10.1186/s13750-020-00191-y
Siller-Clavel, P., Badano, E. I., Villarreal-Guerrero, F., Prieto-Amparán, J. A., Pinedo-Álvarez, A., Corrales-Lerma, R. et al. (2022). Distribution Patterns of Invasive Buffelgrass (Cenchrus ciliaris) in Mexico Estimated with Climate Niche Models under the Current and Future Climate. Plants, 11, 1160. https://doi.org/10.3390/plants11091160
Sokolov, A., Sokolova, G., Bairambekov, S. y Boeva, T. (2020). Change in species composition of vegetation on various-aged set-aside lands of the Volga Delta. E3S Web of Conferences, 64, 07015. https://doi.org/10.1051/e3sconf/202016407015
Story, M. H. (1986). Accuracy assessment: a user’s perspective. Photogrammetric Engineering and Remote Sensing, 52, 397–399.
Tropical Rainfall Measuring Mission (TRMM). (2011). TRMM (TMPA/3B43) Rainfall Estimate L3 1 month 0.25 degree x 0.25 degree V7, Greenbelt, MD, Goddard Earth Sciences Data and Information Services Center (GES DISC). Recuperado el 07 abril, 2020 de: https://disc.gsfc.nasa.gov/datasets/TRMM_3B43_7/summary
Tso, B. y Mather, M. P. (2009). Classification methods for remotely sensed data. Washington, D.C.: CRC Press.
Tucker, C. J. (1979). Red and photographic infrared linear combinations for monitoring vegetation. Remote Sensing of Environment, 8, 127–150. https://doi.org/10.1016/0034-4257(79)90013-0
Turner M. G. y Gardner, R. H. (2015). Landscape Ecology in theory and practice: pattern and process. New York: Springer.
UACh (Universidad Autónoma de Chihuahua). (2011). Actualización de la delimitación de las zonas áridas, semiáridas y subhúmedas de México, a escala regional. Reporte final de proyecto de investigación. Ciudad de México: Universidad Autónoma Chapingo.
USGS (United States Geological Survey). (2022). EarthExplorer. Recuperado el 13 junio, 2020, de: https://earthexplorer.usgs.gov/
van Leeuwen, W. J., Huete, A. R. y Laing, T. W. (1999). MODIS vegetation index compositing approach: A prototype with AVHRR data. Remote Sensing of Environment, 69, 264–280. https://doi.org/10.1016/s0034-4257(99)00022-x
Wang, Y., Lu, Z., Sheng, Y. y Zhou, Y. (2020). Remote sensing applications in monitoring of protected areas. Remote Sensing, 12, 1370. https://doi.org/10.3390/rs12091370
Willis, K. S. (2015). Remote sensing change detection for ecological monitoring in United States protected areas. Biological Conservation, 182, 233–242. https://doi.org/10.
1016/j.biocon.2014.12.006
WWF (World Wide Fund for Nature). (2020). Living Planet Report 2020 – Bending the curve of biodiversity loss. Gland, Switzerland: WWF.
Zhang, Y., Tariq, A., Hughes, A. C., Hong, D., Wei, F., Sun, H. et al. (2022). Challenges and solutions to biodiversity conservation in arid lands. Science of The Total Environment, 857, 159695. https://doi.org/10.1016/j.scitotenv.2022.159695
Zhou, Q. (2016). Digital elevation model and digital surface model. En D. Richardson, N. Castree, M. F. Goodchild, A. Kobayashi, W. Liu, y A. R. Marston (Eds.), The International Encyclopedia of Geography (pp. 1–17). Oxford, UK: Wiley-Blackwell.


