
Samuel Nolasco a, b, Alejandro Valdez-Mondragón c, d, *
a Laboratorio de Aracnología, Laboratorio Regional de Biodiversidad y Cultivo de Tejidos Vegetales, Instituto de Biología, Universidad Nacional Autónoma de México, sede Tlaxcala, Ex-Fábrica San Manuel, San Miguel Contla, 90640 Santa Cruz Tlaxcala, Tlaxcala, Mexico
b Posgrado en Ciencias Biológicas, Centro Tlaxcala de Biología de la Conducta, Universidad Autónoma de Tlaxcala, Carretera Federal Tlaxcala- Puebla, Km. 1.5, 90062 Tlaxcala, Tlaxcala, Mexico
c Conacyt Research Fellow, Laboratorio de Aracnología, Laboratorio Regional de Biodiversidad y Cultivo de Tejidos Vegetales, Instituto de Biología, Universidad Nacional Autónoma de México, sede Tlaxcala, Ex-Fábrica San Manuel, San Miguel Contla, 90640 Santa Cruz Tlaxcala, Tlaxcala, Mexico
d Colección Nacional de Arácnidos, Departamento de Zoología, Instituto de Biología, Universidad Nacional Autónoma de México, 04510 México City, Mexico
*Corresponding author: lat_mactans@yahoo.com.mx (A. Valdez-Mondragón)
Received: 4 Dicember 2019; accepted: 10 March 2020
http://zoobank.org/urn:lsid:zoobank.org:pub:23829591-817F-4AFE-BF3D-A3012229D9E2
Abstract
A new species of daddy long–legs spiders of the genus Physocyclus Simon, 1893 of the globosus species group is described from the Baja California Peninsula desert, in the state of Baja California Sur, Mexico. The species is described based on adult males and females: Physocyclus xerophilus sp. nov. This is the sixth species described from Baja California Sur. The total number of species of Physocyclus increases to 34, including the new species described herein. Thirty-two of 34 species are distributed in Mexico, with one cosmopolitan: Physocyclus globosus. The states of Baja California Sur, Guerrero and Michoacán have the highest diversity of species in the country, with 8 species each.
Keywords: Arteminae; Taxonomy; Xerophytic scrub; Baja California Sur
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Sobre las arañas patonas del género Physocyclus (Araneae: Pholcidae) de México: descripción de una especie nueva de la península de Baja California
Resumen
Se describe una especie nueva de araña patona del género Physocyclus Simon, 1893 del grupo de especies globosus del desierto de la península de Baja California, en el estado de Baja California Sur, México. Physocyclus xerophilus sp. nov. se describe con base en machos y hembras adultos y es la sexta especie descrita para este estado. El número total de especies del género Physocyclus aumenta a 34, incluyendo la especie nueva descrita aquí; 32 de 34 especies están distribuidas en México, con una cosmopolita: Physocyclus globosus. Baja California Sur, Guerrero y Michoacán tienen la mayor diversidad de especies en el país, con 8 especies cada uno.
Palabras clave: Arteminae; Taxonomía; Matorral xerófilo; Baja California Sur
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
The spider family Pholcidae Koch, 1850 is the ninth largest in the order Araneae, with 1,743 species including the new species described herein, distributed in 94 genera (WSC, 2020). The family is subdivided into 5 subfamilies: Arteminae Simon, 1893, Modisiminae Simon, 1893, Ninetinae Simon, 1890, Pholcinae C.L. Koch, 1850 and Smeringopinae Simon, 1893 (Dimitrov et al., 2013; Eberle et al., 2018; Huber, 2011). The spider genus Physocyclus Simon, 1893 is classified in the subfamily Arteminae, which currently has 103 species in 9 genera.
Physocyclus is one of the most diversified and common pholcid spiders in Mexico, which currently includes 33 species (Jiménez & Palacios-Cardiel, 2013; Valdez-Mondragón 2010, 2013, 2014; WSC, 2020). The genus is distributed mainly in North and Central America, with most of the species from Mexico, including the synanthropic species Physocyclus dugesi Simon and Physocyclus globosus (Taczanowski), the latter with a cosmopolitan distribution influenced by human activities (Beatty et al., 2008; Huber & Kwapong, 2013; Valdez-Mondragón, 2010). Although some species are synanthropic, the common habitat is in dry and semi-arid climates, and most of the species have been collected in dry tropical deciduous forests and deserts, as the new species described here. In karstic zones, some species are found frequently in their sheet webs inside caves, on walls, holes in walls, amid large rocks on the ground, or among the karst formations (stalagmites, stalactites and columns) (Valdez-Mondragón, 2010, 2014).
Morphologically, their legs are long with relation to the body, which ranges from 3 to 5 mm, and show a ringed coloration on tibiae and femora. The body coloration varies between white and pale yellow, with an irregular brown dorsal pattern on the carapace, usually with 3 symmetrical spots on each side. However, the general overall shape and coloration is similar among the species, which indicating a conserved morphological pattern (Valdez-Mondragón, 2010, 2014).
Physocyclus is a monophyletic group based on morphological characters currently constituted by 2 species groups, globosus and dugesi, with 12 and 20 species respectively, including the new species (Valdez-Mondragón, 2013, 2014). The globosus group has a biogeographical distribution pattern in the Mesoamerican and Mexican Mountain biotic components, whereas the dugesi group has a biogeographical distribution in the Mesoamerican and Continental Nearctic biotic components (Valdez-Mondragón, 2013, 2014).
In this contribution we describe a new species of daddy long-legs spiders of the genus Physocyclus belonging to the globosus species group, based on adult males and females from the desert of the Baja California Peninsula in the state of Baja California Sur, Mexico.
Material and methods
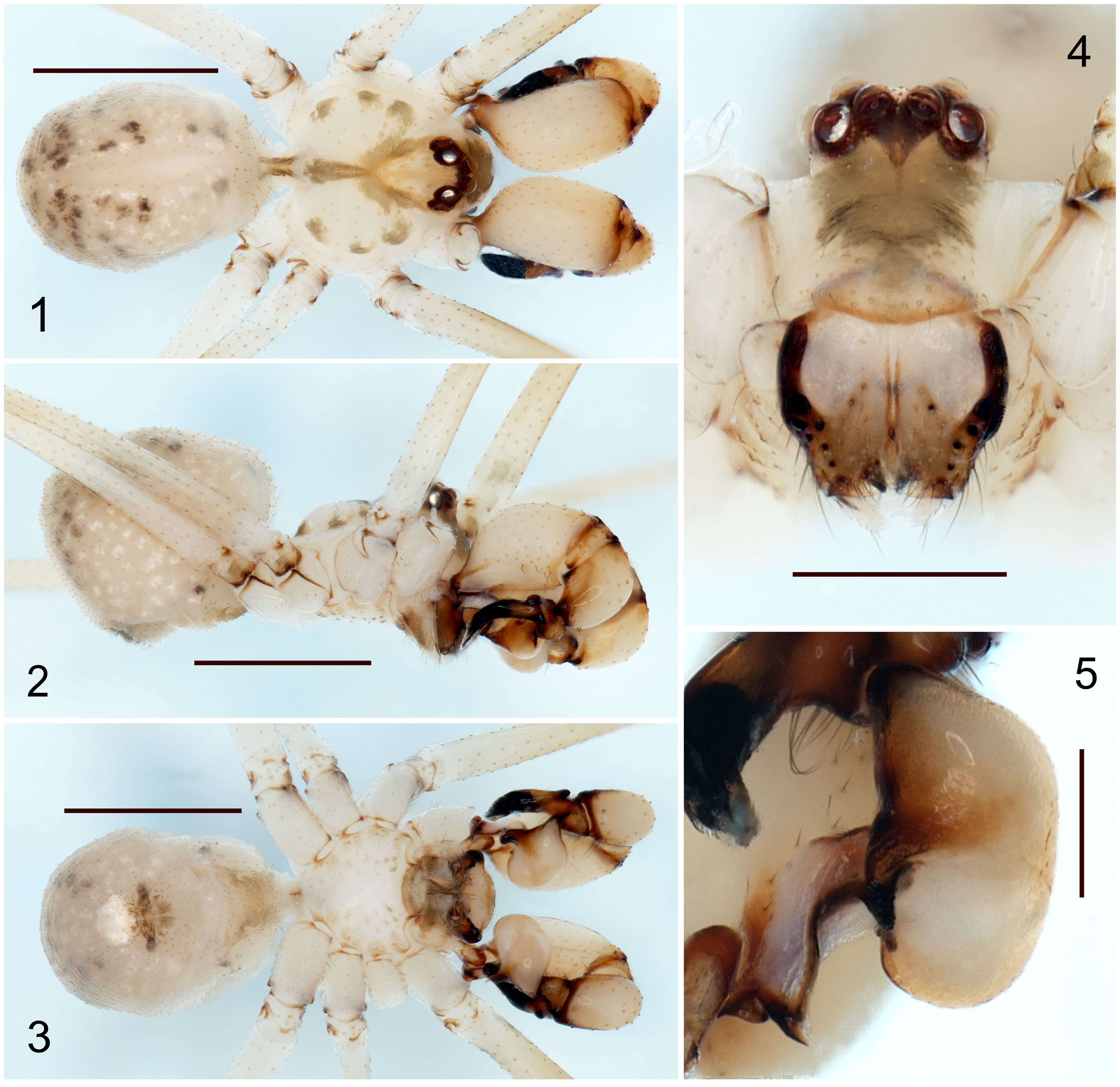
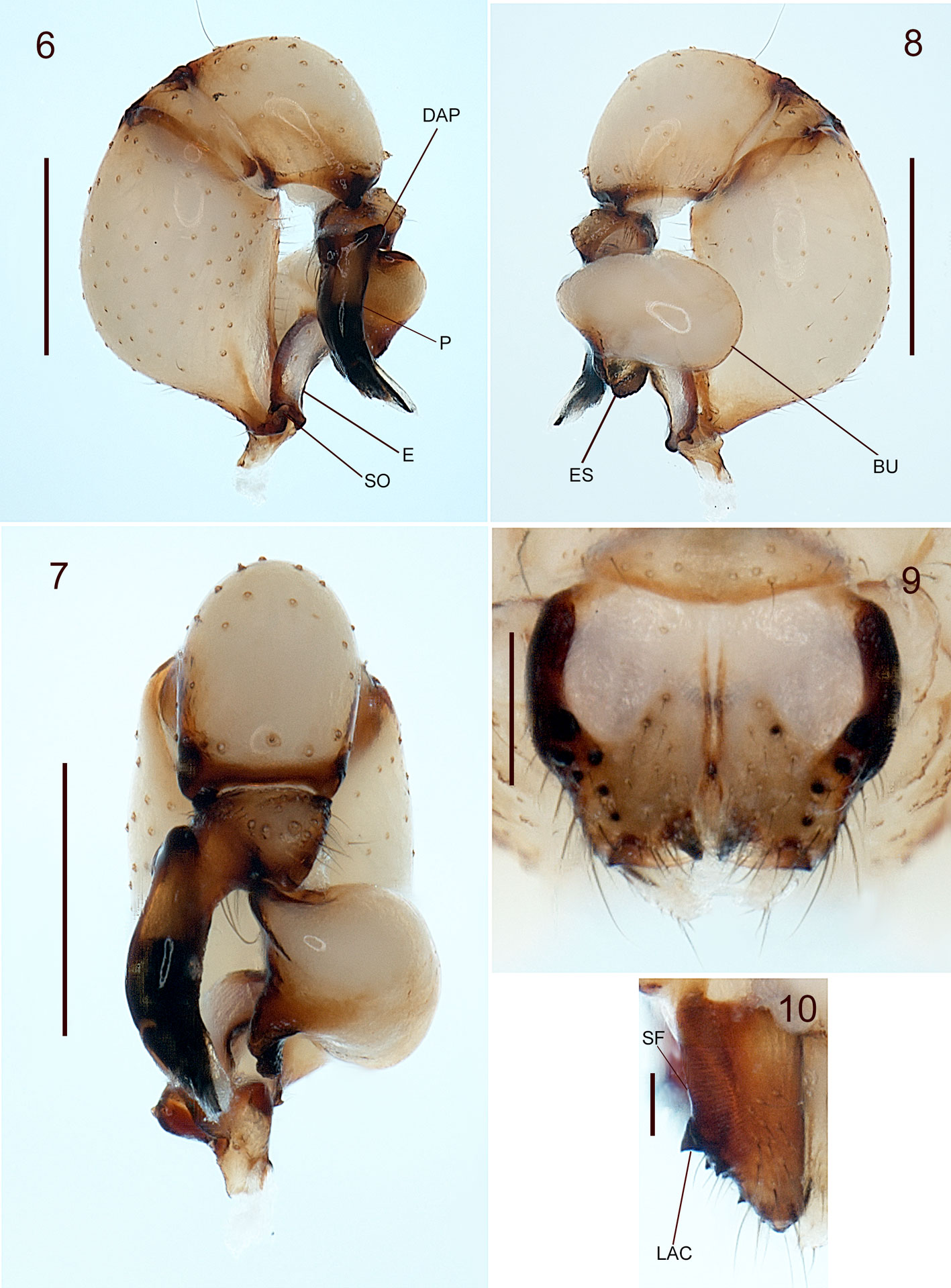
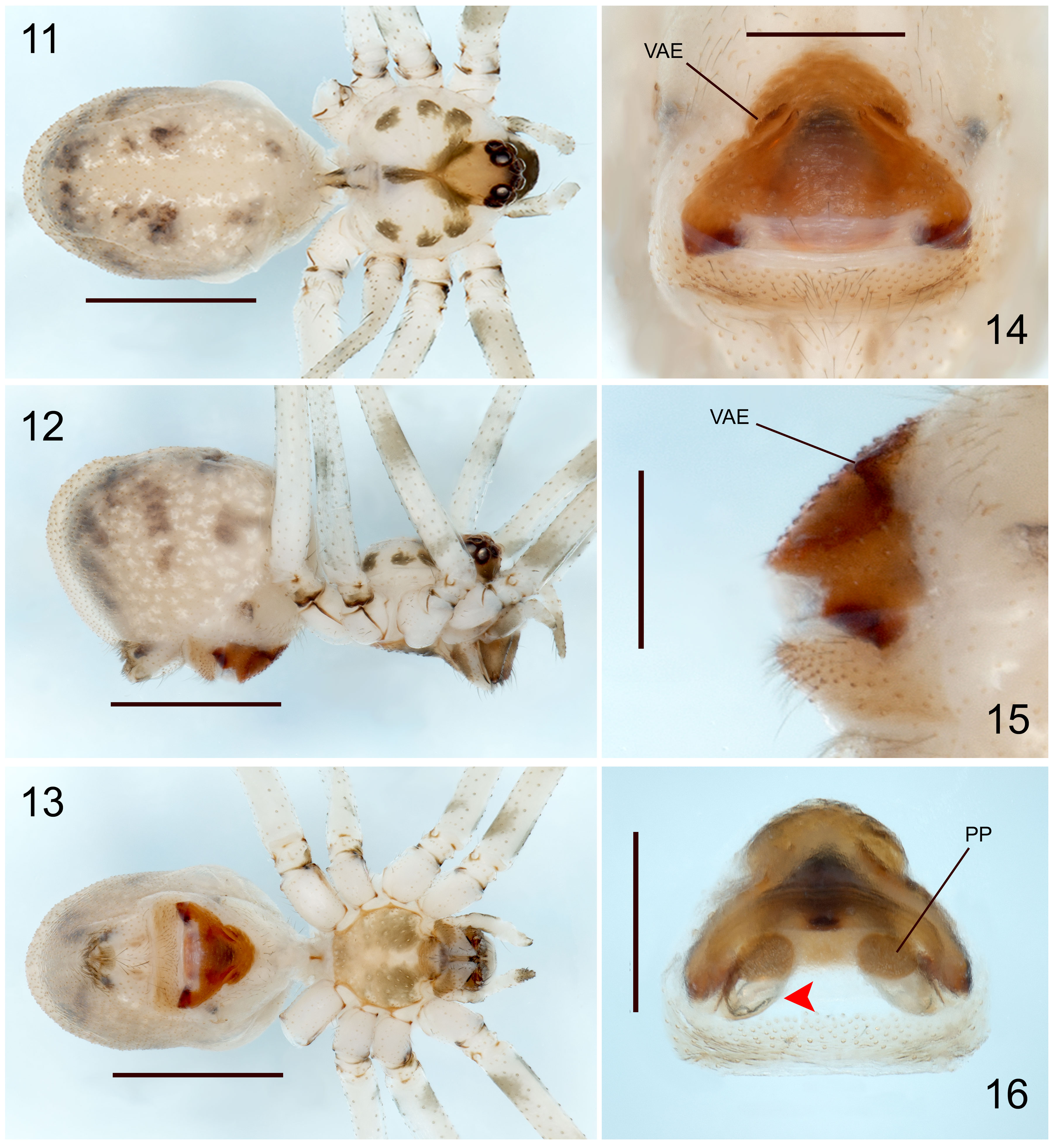
The specimens used for this study are preserved in ethanol (80%), and for future molecular studies in ethanol (96%), deposited at the Colección Nacional de Arácnidos (CNAN), Instituto de Biología, Universidad Nacional Autónoma de México, Mexico City (curator: Dr. Oscar F. Francke Ballvé). The specimens were collected under the Scientific Collector Permit FAUT-0309 from the Ministry of Environment and Natural Resources (Semarnat) granted to Dr. Alejandro Valdez Mondragón. All the specimens were measured and examined with a Zeiss Discovery Stereoscope V.8. Digital photographs were made using a Zeiss Axio Zoom V.16 microscope, and Axio Zoom Zen and Zen Pro software. All measurements are in millimeters (mm). The male palp and female epigynum were dissected and observed in ethanol (80%), and the epigynum was cleaned in potassium hydroxide (KOH, 10%) for a few minutes to remove all soft tissues around the pore plates. The map was done with QGIS v.3.10. The digital photographs and map were edited in Photoshop CS6 v.1.30×32. Morphological terminology follows Valdez-Mondragón and Francke (2015). Abbreviations: ALE, anterior lateral eyes; AME, anterior median eyes; PLE, posterior lateral eyes; PME, posterior median eyes; P, procursus; E, embolus; DAP, dorsal apophyses of procursus; SO, spermatic operculum; ES, embolic sclerites; BU, bulb of palp; SF,stridulatoy files of cheliceare; LAC, lateral apophyses of chelicerae; VAE, ventral apophysis of epigynum; PP, pore plates; L/d, tibia I lenght/diamater.
Description
Family Pholcidae C. L. Koch, 1850
Genus Physocyclus Simon, 1893
Type species. Physocyclus globosus (Taczanowski, 1874)
Physocyclus xerophilus sp. nov.
Figs. 1-16
http://zoobank.org/urn:lsid:zoobank.org:act:18DBFAA3-
D914-4DDC-9F27-7821C3E1246F
Diagnosis. Males of Physocyclus xerophilus sp. nov. resemble Physocyclus globosus in the number of sclerotized cones on the frontal part of the chelicerae (4-5 cones on each chelicerae) (Figs. 9, 10; Valdez-Mondragón, 2010: figs. 42-43). However, in the new species the lateral conical apophyses of the chelicerae are longer and sharper than P. globosus (Fig. 10; Valdez-Mondragón, 2010: Fig. 43). In retrolateral view, the procursus in P. xerophilus sp. nov. is thinner and curved, with a thin distal spine (Fig. 6); whereas in P. globosus the procursus is wider and conical, with a small, wide spine distally (Valdez-Mondragón, 2010: Fig. 44). The embolic sclerites (ES) in the male palp are on the bulb in P. xerophilus (Fig. 5), whereas the ES in P. globosus are located on the embolus (E) (Valdez-Mondragón, 2010: Fig. 45). In dorsal view, the embolus in P. xerophilus sp. nov. is wider and curved (Fig. 45), whereas the embolus in P. globosus is thinner and almost straight (Valdez-Mondragón, 2010: Fig. 45). Females of P. xerophilus sp. nov. resemble Physocyclus lautus Gertsch, 1971 in the shape of the epigynum (Figs. 14-16; Valdez-Mondragón, 2010: Figs. 67-69). However, in the new species the ventral apophyses of the epigynum (VAE) are shorter and in oblique position (Figs 14, 15), whereas in P. lautus the VAE are longer and conical (Valdez-Mondragón, 2010: Figs. 67, 69). The epigynum of P. lautus has paired W-shaped concavities in the middle (Valdez-Mondragón, 2010: Fig. 67), whereas in P. xerophilus sp. nov. those concavities are absent (Fig. 14). In dorsal view, the pore plates (PP) of the epigynum of P. xerophilus sp. nov. are almost circular (Fig. 16), whereas in P. lautus the PP are longer and oval (Valdez-Mondragón, 2010: Fig. 68). The PP in the new species have bag-shaped structures posteriorly, which are oval and smaller (arrow, Fig. 16), whereas in P. lautus they are longer and circular (Valdez-Mondragón, 2010: Fig. 68).
Male (holotype) (CNAN-T01365). Prosoma. Carapace light beige, with a Y-shaped light brown pattern around fovea, which extends towards the posterior part of carapace. Fovea longitudinal. Carapace with 3 irregular spots on each side (Fig. 1). Clypeus with a wide light brown mark lengthwise. Each chelicera with 4-5 sclerotized cones frontally (Fig. 4). Chelicerae with stridulatory files laterally (Figs. 4, 9, 10). Chelicerae with a lateral conical apophysis (Fig. 10). Sternum light beige, with small and inconspicuous light brown spots. Labium brown, wider than long, merged to the sternum. Endites brown prolaterally, beige retrolaterally, longer than wide (Fig, 3). Legs. All segments with beige coloration. Trochanters light brown retrolaterally with a notch distally (Figs. 2, 3). Femora and tibiae with brown rings distally. Opisthosoma. Globular, longer than wide and high, with numerous white and dark brown spots (Figs. 1-3). Spinnerets brown, with a short brown seta around (Fig. 3). Palp. Dorsal apophyses of procursus (DAP) conical (Fig. 6). Procursus (P) thin, with basal half dark brown and distal half dark, ending in a thin and long spine with a brush of pseudotrichia distally (Figs. 6-8). Femur without apophyses (Figs. 6, 8). In prolateral view, ES with oval shape located on the bulb (Figs. 5, 7, 8). E wide, slighly curved in retrolateral view, strongly curved on dorsal view (Figs. 5, 6). E with V-shaped apically (Fig. 5). Spermatic operculum (SO) on apical part of E (Fig, 6). Measurements. Total lenght 2.7. Carapace 1.18 long, 1.23 wide. Clypeus 0.6 long. Diameter AME 0.08, ALE 0.12, PME 0.10, PLE 0.11. Distance ALE-PME 0.05, PME-PME 0.15. Leg I: 27.19 (7.04+0.47+7.68+10.6+1.4). Tibia II: 5.18, tibia III: 3.32, tibia IV: 5.12; tibia I L/d: 51.2.
Female (paratype) (CNAN-T01366). Similar to the male, differences: Prosoma. Dorsal pattern of carapace darker brown than male (Fig. 11). Sternum brown, with a small white region on anterior part (Fig. 13). Labium and endites brown, darker than male (Fig. 13). Clypeus darker than male. Chelicerae without sclerotized cones nor lateral apophyses. Legs. Trochanters light brown retrolaterally, darker than male. Femora and tibiae with darker brown rings distally. Epigynum. Wider than long, bell-shaped, VAE small and conical (Figs. 14, 15). PP small, oval, with bag-shaped structures below them (Fig. 16). Measurements. Total lenght 3.04. Carapace 1.20 long, 1.23 wide. Clypeus 0.4 long. Diameter AME 0.08, ALE 0.11, PME 0.10, PLE 0.10. Distance ALE-PME 0.05, PME-PME 0.14. Leg I: 19.49 (5.18+0.46+5.50+7.25+1.). Tibia II: 3.60, tibia III: 2.28, tibia IV: 4.00; tibia I L/d: 30.5.
Variation. Male paratype smaller than male holotype, with lighter coloration. Chelicerae of the male paratype has darker coloration on basal part. Females with small variation in size. Female collected on Santipac Beach is smaller and with darker coloration in the epigynum than the females from the type locality. Females from Cocos Beach, Misión Mulegé, and the road to San Isidro-San José Comundú with small variaton in size. Males: (N = 2), tibia I: 6.64, 7.68. Females: same locality as holotype (N = 2), tibia I: 4.50, 5.81. 800 SE of Misión Mulegé (N = 1), tibia I: 5.75. Santipac Beach (N = 1), tibia 1: 4.55. Cocos Beach (N = 1), tibia I: 5.37. Comundú (N = 1), tibia I: missing.
Taxonomic summary
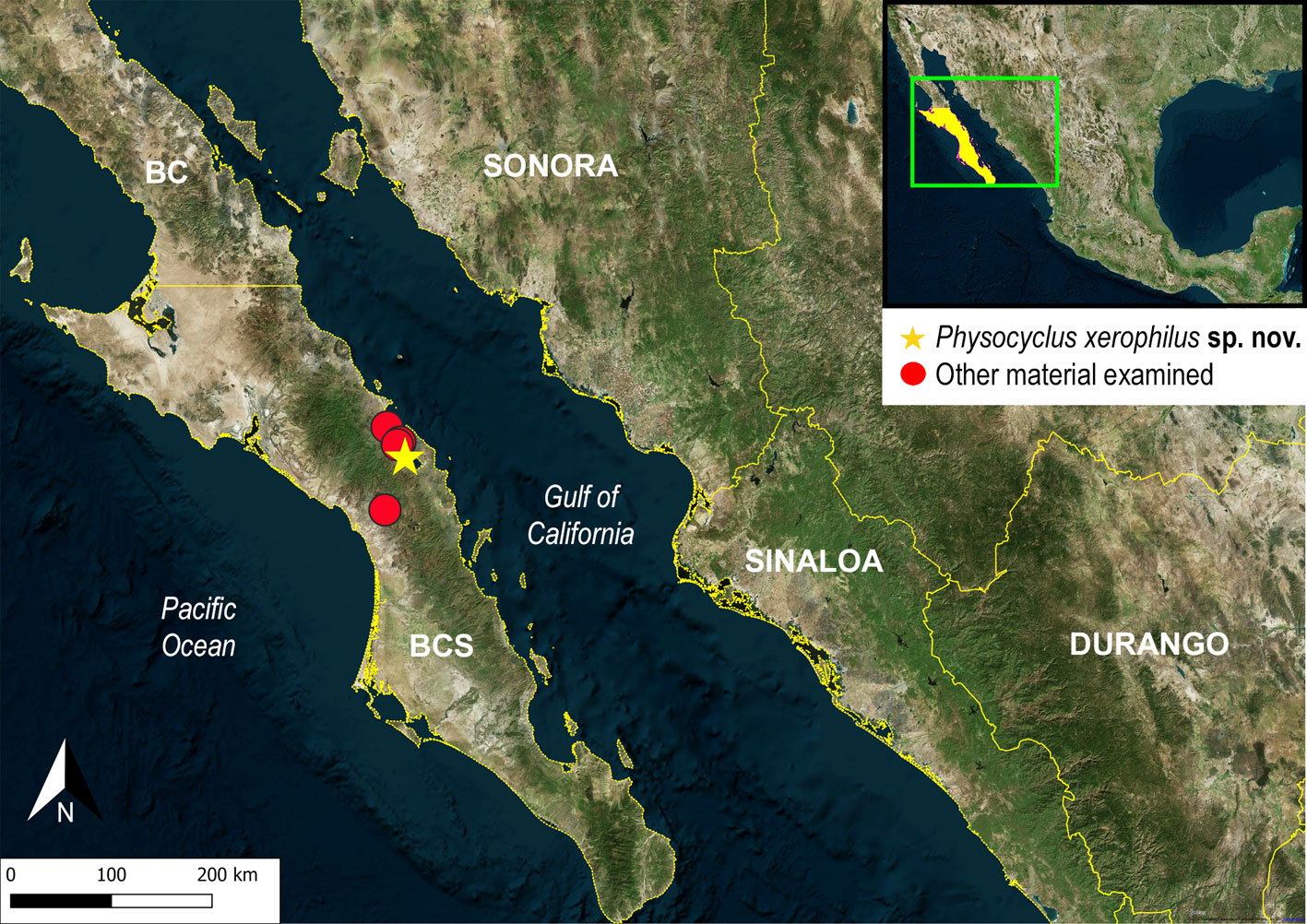
Type material. Mexico: Baja California Sur: 1 male holotype (CNAN-T01365) [21 July 2019; A. Valdez; P. Solís; D. Montiel; A. Cabrera Cols.], from close to “El Requesón” Beach (26.63539° N, -111.83173° W; 6 m), Mulegé Municipality. Paratypes: 1 female (CNAN-T01366); 1 male, 2 females (CNAN-T01367), same data as holotype.
Other material examined. Mexico: Baja California Sur: 2 females (Ara-0541) [21 July 2019; A. Valdez, P. Solís, D. Montiel, A. Cabrera Cols.], 800 m SE from Misión Mulegé (lat 26.88183°, lon -111.99310°; 9 m), Mulegé Municipality. 1 female (Ara-0542) [21 July 2019; A. Valdez, P. Solís, D. Montiel, A. Cabrera Cols.], Santipac Beach, Loreto-Santa Rosalía road (26.76198° N, -111.87842° W; 2 m), Mulegé Municipality. 1 female (Ara-0543) [29 July 2019; A. Valdez, P. Solís, D. Montiel, A. Cabrera Cols.], Cocos Beach (26.74279° N, -111.90151° W; 1 m), Mulegé Municipality. 1 female (Ara-0544) [30 July 2019; A. Valdez, P. Solís, D. Montiel, A. Cabrera Cols.], Km 1 San Isidro-San José Comundú road (26.22135° N, -112.01165° W; 137 m), Comundú Municipality.
Etymology. The specific name is a noun in apposition and refers to the vegetation type where the type locality of the species is: xerophilous scrub or desert.
Natural history. The specimens were collected on their sheet webs among and under large boulders on the ground in the different localities. The vegetation type of the zone is dry xerophilous scrub, with cacti and scrubby vegetation (Figs. 17, 18).
Distribution. Mexico: Baja California Sur (Fig. 19).
Acknowledgements
The second author thanks the program “Cátedras Conacyt” Consejo Nacional de Ciencia y Tecnología (Conacyt), Mexico, for scientific support for Project No. 59, Regional Laboratory for Biodiversity and Plant Tissue Cultivation (LBCTV), Institute of Biology, National Autonomous University of Mexico (UNAM) campus Tlaxcala. The second author also thanks SEP-Conacyt for their financial support for the Basic Science Project (Ciencia Básica) 2016, No. 282834. The first author thanks the Doctorate in Biological Sciences of the Tlaxcala Center for Behavior Biology (CTBC), Autonomous University of Tlaxcala (UATx) for their educational support, and the Conacyt for scholarship support in the Doctorate Program; to the Ministry for the Development of Agriculture and Livestock (SEFOA), and the Government of the state of Tlaxcala, for their considerations and support to do this research; and to Mayra R. Cortez-Roldán for her help with the distribution map. We also thank the students of the Laboratory of Arachnology (LATLAX), IBUNAM, Tlaxcala, for their help both in the field and in processing the material in the laboratory; Cheryl Harleston for the English language revision of the manuscript.

References
Beatty, J. A., Berry, J, W., & Huber, B. A. (2008). The pholcid spiders of Micronesia and Polynesia (Araneae: Pholcidae). Journal of Arachnology, 36, 1–25. https://doi.org/10.1636/h05-66.1
Dimitrov, D., Astrin, J. J., & Huber, B. A. (2013). Pholcid spider molecular systematics revisited, with new insights into the biogeography and evolution of the group. Cladistics, 29, 132–146. https://doi.org/10.1111/j.1096-0031.2012.00419.x
Eberle, J., Dimitrov, D., Valdez-Mondragón, A., & Huber, B. A. (2018). Microhabitat change drives diversification in pholcid spiders. BMC Evolutionary Biology, 18, 141. https://doi.org/10.1186/s12862-018-1244-8
Huber, B. A. (2011). Phylogeny and classification of Pholcidae (Araneae): an update. Journal of Arachnology, 39, 211–222. https://doi.org/10.1636/ca10-57.1
Huber, B. A., & Kwapong, P. (2013). West African pholcid spiders: an overview, with descriptions of five new species (Araneae, Pholcidae). European Journal of Taxonomy, 59, 1–44. https://doi.org/10.5852/ejt.2013.59
Jiménez, M. L., & Palacios-Cardiel, C. C. (2013). A new species of Physocyclus (Araneae: Pholcidae) from Mexico. Zootaxa, 3717, 96–99. https://doi.org/10.11646/zootaxa.3717.1.8
Valdez-Mondragón, A. (2010). Revisión taxonómica de Physocyclus Simon, 1983 (Araneae: Pholcidae) con la descripción de especies nuevas de México. Revista Ibérica de Aracnología, 18, 3–80.
Valdez-Mondragón, A. (2013). Morphological phylogenetic analysis of the spider genus Physocyclus (Araneae: Pholcidae). Journal of Arachnology, 41, 184–196. https://doi.org/10.1636/k12-33.1
Valdez-Mondragón, A. (2014). A reanalysis of the morphological phylogeny of the spider genus Physocyclus Simon, 1983 (Araneae: Pholcidae) with the description of a new species and description of the female of Physocyclus paredesi Valdez-Mondragón from México. Zootaxa, 3866, 202–220. https://doi.org/10.11646/zootaxa.3866.2.2
Valdez-Mondragón, A., & Francke, O. F. (2015). Phylogeny of the spider genus Ixchela Huber, 2000 (Araneae: Pholcidae) based on morphological and molecular evidence (CO1 and 16S), with a hypothesized diversification in the Pleistocene. Zoological Journal of the Linnean Society, 175, 20–58. https://doi.org/10.1111/zoj.12265
WSC (World Spider Catalog). (2020). World Spider Catalog. Version 21.0. Natural History Museum Bern, online at http://wsc.nmbe.ch, accessed on 10/07/2020


