Defining the phylogeographic relationship between cis- and trans-Andean populations of Dendrocincla fuliginosa and Xenops minutus in Colombia
Enrique Arbeláez-Cortés *
Grupo de Estudios en Biodiversidad, Escuela de Biología, Facultad de Ciencias, Universidad Industrial de Santander, Carrera 27 Calle 9, Bucaramanga, Santander, Colombia
*Corresponding author: enriquearbelaez@gmail.com (E. Arbeláez-Cortés)
Abstract
Due to the position of Colombia between Central and South America, and the presence of several ecosystems in its territory, phylogeographic information from populations across this country is fundamental to understand the evolutionary history of widespread Neotropical species. The Andes have long been noted for their influence isolating lowland species into cis- and trans-Andean populations. However, detailed sampling across cis-Andean zones adjacent to the Andes (i.e., Orinoquia) has been lacking in avian phylogeographic studies. Information from DNA sequences, from 2 passerine birds: Dendrocincla fuliginosa and Xenops minutus is presented herein; to depict their phylogeographic patterns, focusing on the relationship between cis- and trans-Andean populations from Colombia. The analyses, regarding Colombian samples indicated that cis-Andean populations (i.e., Orinoquia and Amazon) are not closely related, rather the Orinoquia populations are more closely related to trans-Andean populations in both species. These relationships suggest that populations on both sides of the Andes were connected in the recent past (less than 1 myA). I propose that phylogeographic differentiation in lowland species in this region is not only explained by the presence of the Andean mountains, but also based on the ecological shifts between major ecosystems such as Amazonia and Orinoquia.
Keywords: Amazonia; Birds; Orinoquia; Lowlands; mtDNA; nDNA; South America
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Definiendo las relaciones filogeográficas entre poblaciones cis- y trans-andinas de Dendrocincla fuliginosa y Xenops minutus en Colombia
Resumen
Por su posición entre Centroamérica y Sudamérica, y por presentar varios ecosistemas, la información filogeográfica de poblaciones de Colombia es fundamental para entender la historia evolutiva de especies neotropicales ampliamente distribuidas. Los Andes se consideran como factor de separación de especies de tierras bajas en poblaciones cis- y trans-andinas. Pero un muestreo detallado de zonas cis-andinas adyacentes a los Andes (i.e., Orinoquia) ha estado ausente de los estudios filogeográficos con aves. Se presenta aquí información de secuencias de ADN de 2 aves paseriformes: Dendrocincla fuliginosa y Xenops minutus, para describir sus patrones filogeográficos, enfocando dicha información en la relación entre poblaciones cis- y trans-andinas en Colombia. Los análisis indican que poblaciones cis-andinas (i.e., Orinoquía y Amazonía) no están estrechamente relacionadas entre sí, sino que las poblaciones de la Orinoquía están más relacionadas con poblaciones trans-andinas en ambas especies. Estas relaciones sugieren que las poblaciones en ambos lados de los Andes estuvieron conectadas en el pasado reciente (menos de 1 MA). Se propone que la diferenciación filogeográfica en especies de tierras bajas en esta región no puede explicarse solo por la presencia de las montañas andinas, sino también con base en cambios ecológicos entre ecosistemas mayores como la Amazonía y la Orinoquía.
Palabras clave: Amazonía; Aves; Orinoquía; Tierras bajas; ADNmt; ADNn; Sudamérica
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
Phylogeographic studies of widespread lowland Neotropical birds usually have indicated a strong genetic differentiation among allopatric populations (Cheviron et al., 2005; Fernandes et al., 2014; Harvey & Brumfield, 2015; Marks et al., 2002; Miller et al., 2008; Nyári, 2007; Rheindt et al., 2009). For lowland species, these studies have usually considered the Andes mountains as a barrier to gene flow between populations to the West (trans-Andean) and to the East (cis-Andean). For species such as Automolus ochrolaemus, A. subulatus (Schultz et al., 2017), Hylophylax naevius/ H. naevioides (Fernandes et al., 2014), Lepidothrix coronata (Cheviron et al., 2005), and Myiopagis viridicata (Rheindt et al., 2009), a clear phylogeographic differentiation between cis- and trans-Andean populations has been found. However, other species such as Glyphorynchus spirurus (Marks et al., 2002), Schiffornis turdina (Nyári, 2007), and Mionectes oleagineus (Miller et al., 2008), exhibit a lower degree of genetic differentiation between trans- and cis-Andean populations. Therefore, recent dispersion (or population connection) across (or around) the Andes, originally proposed by Haffer (1967), has been considered to explain such phylogeographic relationships between cis- and trans-Andean populations. Results of these single-species studies, agree with those studies including several lowland bird species (Smith, Harvey et al., 2014; Smith, McCormack et al., 2014), indicating that phylogeographic relationships among populations in Neotropical lowlands are complex. Connection across the Andes as a cause underlying the actual patterns of species distributions has also been analyzed using geographic models of potential ranges of bird species, and there is evidence of historic ecological continuity across low-lying passes that could promote population connectivity between cis- and trans-Andean regions (Cadena et al., 2016).
Despite advances in the knowledge of the evolution of lowland Neotropical birds, phylogeographic patterns of these lineages have been derived mainly from information from Mexico, Central America, and Brazil; but a gap often exists in phylogeographic information from Colombia. Given the geographic location of Colombia between Central and South America, their complex landscape, and the dynamic history of their environment (Cadena et al., 2007, 2016; Flantua et al., 2007; Haffer, 1967; Nores, 2004); more taxon and geographic sampling across this country is crucial to reconstruct the evolutionary history of Neotropical organisms (Avendaño et al., 2017). To test if Colombian populations adhere to the established cis/trans Andean dichotomy, phylogeographic patterns of 2 widespread lowland species that range on both sides of the Andes and inhabit forests were analyzed: Dendrocincla fuliginosa and Xenops minutus (Furnariidae).
Dendrocincla fuliginosa (Vieillot, 1818) ranges from southeastern Honduras to eastern Brazil, mainly below 1,300 m asl, but occasionally up to 2,000 m asl. This species inhabits humid and savanna forests in lowlands and foothills, generally in mature forests, but it also tolerates secondary vegetation and plantations. Dendrocincla fuliginosa forages for invertebrates and small vertebrates on vertical trunks at low- and mid-levels, usually alone but also follows ant swarms (as facultative) and mixed-species flocks (Del Hoyo et al., 2003; Restall et al., 2006). This species comprises 11 subspecies; 4 of which occur in Colombia (Gill & Donsker, 2018). Molecular information indicated that the diversification of D. fuliginosa began around 4 myA (Derryberry et al., 2011; Weir & Price, 2011).
Materials and methods
Eighteen tissue samples of D. fuliginosa (17 localities from 8 Colombian departments) and 13 tissue samples of X. minutus (12 localities from 9 Colombian departments), covering their main distribution ranges in Colombia (Appendix 1) were used. DNA was isolated from frozen tissues using Qiagen DNeasyTM kit (Qiagen Inc., Valencia, CA, USA), following the manufacturer protocol. I amplified the mitochondrial (mtDNA) gene subunit 2 of the nicotinamide adenine dinucleotide dehydrogenase (ND2), using primers H1056U and L5215U (H. Vázquez-Miranda in litt.) and the nuclear (nDNA) locus 20454 using primers 20454F and 20454R (Backström et al., 2008). All PCR trials were performed in a final volume of 15 μl, containing 1x PCR buffer, 1.5 mM MgCl2, 0.2 mM each dNTP, 0.4-0.5 μM each primer, 1-2 unit DNA polymerase, and around 50-100 ng of DNA. PCR conditions were the same used for those loci in Arbeláez-Cortés et al. (2014).
PCR products were purified using Exo-SAP-ITTM (GE Healthcare Bio-Sciences Corp. Piscataway, NJ, USA), and the sequences were obtained by ABI Prism BigdyeTM v3.1 (Qiagen Inc., Valencia, CA, USA) terminator chemistry in an ABI 3730XL automated sequencer located in an overseas facility (University of Washington, USA). All PCR products were exported from Colombia under the respective permission of Colombian authorities (see Acknowledgments). Sequences were edited and manually aligned using BioEdit (Hall, 1999). For sequences of nDNA, I inspected chromatograms to detect double peaks, which were edited by coding them following a standard IUPAC ambiguity code. Double peaks were scanned across all individuals to check for accuracy and consistency in their identification. The allele phase of the nDNA locus was resolved using a coalescent-based Bayesian method of the Phase algorithm (Stephens & Donnelly, 2003; Stephens et al., 2001) in DNAsp v.5 (Librado & Rozas, 2009) employing default settings. The resulting highest-probability haplotypes for further analyses were used. All sequences are deposited in GenBank (MN486096-MN486154).
For the mtDNA locus ND2, additional sequences of other countries from GenBank were obtained. For D. fuliginosa, 17 sequences from Brazil, 3 from Suriname, 2 from Peru, 2 from Venezuela, 2 from Panama, 1 from Ecuador, and 1 from Trinidad and Tobago were obtained (Claramunt et al., 2010; Naka et al., 2012; Weir et al., 2009). For X. minutus, 21 sequences were obtained from Guyana, 10 from Belize, 9 from Panama, 8 from Brazil, 2 from Venezuela, 1 from Ecuador, and 1 from Paraguay (Derryberry et al., 2011; Naka et al., 2012, M.J. Miller, 2008 unpublished). For each of these GenBank sequences, I obtained the geographical information available either from: their metadata, the database of the biological collection which harbors the sample, the supplementary material of the cited papers, or from other papers in which several of these individuals have been included (C.W. Burney, 2009 unpublished; Smith et al., 2014). Only GenBank sequences for which there was at least country information were included. I georeferenced localities without information using GeoNames (GeoNames, 2013) or Google Earth (Google, 2010). In the cases where only the information for the country or state was available, I used central geographic coordinates to depict their position on a map only for illustrative purposes. I included ND2 sequences from closest congeneric species, Xenops rutilans and Dendrocincla anabantina (Derryberry et al., 2011) as outgroups. A list of samples with GenBank accession numbers is provided in appendix 1. Because the size of the sequences obtained and the ones available from GenBank varied, I trimmed the ND2 alignments to 625 bp, for D. fuliginosa, and to 622 bp for X. minutus. For the nDNA locus 20454 I aligned 285 bp for D. fuliginosa and 381 bp for X. minutus. However, full sequences were deposited in GenBank.
It should be noted that Weir and Price (2011) indicated that D. fuliginosa is paraphyletic. These authors found that the eastern South American populations are distantly related from the western populations. For this reason, I analyzed both the whole set of sequences available for D. fuliginosa and a subset from its western range, but I focus my results and discussion on the latter. Here, I refer to these populations as D. fuliginosa, even though the information in Weir and Price (2011) indicates that they can be considered under a different name.
BEAST version 1.7.4 (Drummond et al., 2012; Drummond et al., 2012) was used to reconstruct phylogenetic relationships among ND2 haplotypes. The best-fit model for each alignment was selected using Akaike’s information criterion in Modeltest, version 3.7 (Posada & Crandall, 1998). I ran BEAST for 200,000,000 steps, sampling every 2,000 steps, using a Yule speciation tree prior, a UPGMA starting tree, and a strict molecular clock with a mutation rate of 2.9 × 10-8 substitutions ⁄ site ⁄ year for ND2 according to rates previously reported in passerine birds (Lerner et al., 2011). After this analysis, TreeAnnotator, version 1.7.4 (Rambaut & Drummond, 2012) was used to generate a tree file with 25% burn-in and a posterior probability limit of 0.5. Sequences of the nDNA locus, available just for Colombia, were examined using Network (Bandelt et al., 1999) to generate an allele network depicting their relationships.
I acknowledge that the estimates of time trees depend on the selected substitution rate for the loci. For instance, looking broadly for externally calibrated rate estimates of the ND2 rate in birds, one finds variation ranging from 1.94% to 12.3% per my (Arbogast et al., 2006; Benham et al., 2015; Fuchs et al., 2011; Johnson & Weckstein, 2011; Patel et al., 2011; Weir & Schluter, 2008). However, my principal aim is to depict the phylogeographic pattern of both species in northwestern South America and not to detail divergence times among clades. The only temporal issue discussed here is the differentiation between cis-Andean (Orinoquia) and trans-Andean populations for both species to compare it with information on the Andes orogenic formation. Because the substitution rate I used is towards the low end of estimated rates for birds it would lead to ancient dates.
Xenops minutus (Sparrman, 1788) ranges from southern Mexico to southeastern South America (below 1,000 m asl, but locally up to 1,500 m asl), and inhabits tropical lowland forests where it forages for arthropods, usually alone or in mixed-species flocks, climbing along branches and woody vines in low- to mid-level vegetation (Del Hoyo et al., 2003; Restall et al., 2006). This species comprises 10 subspecies; 5 of which occur in Colombia (Gill & Donsker, 2018). Molecular information suggests that the split of X. minutus from its sister species occurred around 10 myA (Derryberry et al., 2011). Harvey & Brumfield (2015) used genome-wide single nucleotide polymorphisms to investigate the phylogeography of this taxon, and found the presence of at least 3 deeply divergent clades that seem to represent different species.
For both species, the Andes have been considered as a barrier involved in their diversification (Harvey & Brumfield, 2015; Weir & Price, 2011), but these studies lack Colombian samples. Smith et al. (2014) included Colombian samples of Xenops minutus and D. fuliginosa in a phylogeographic study, but analyzed general patterns across multiple species. The aim here is to depict the phylogeographic patterns of both species, focusing on defining relationships among cis- and trans-Andean populations from Colombia.
Results
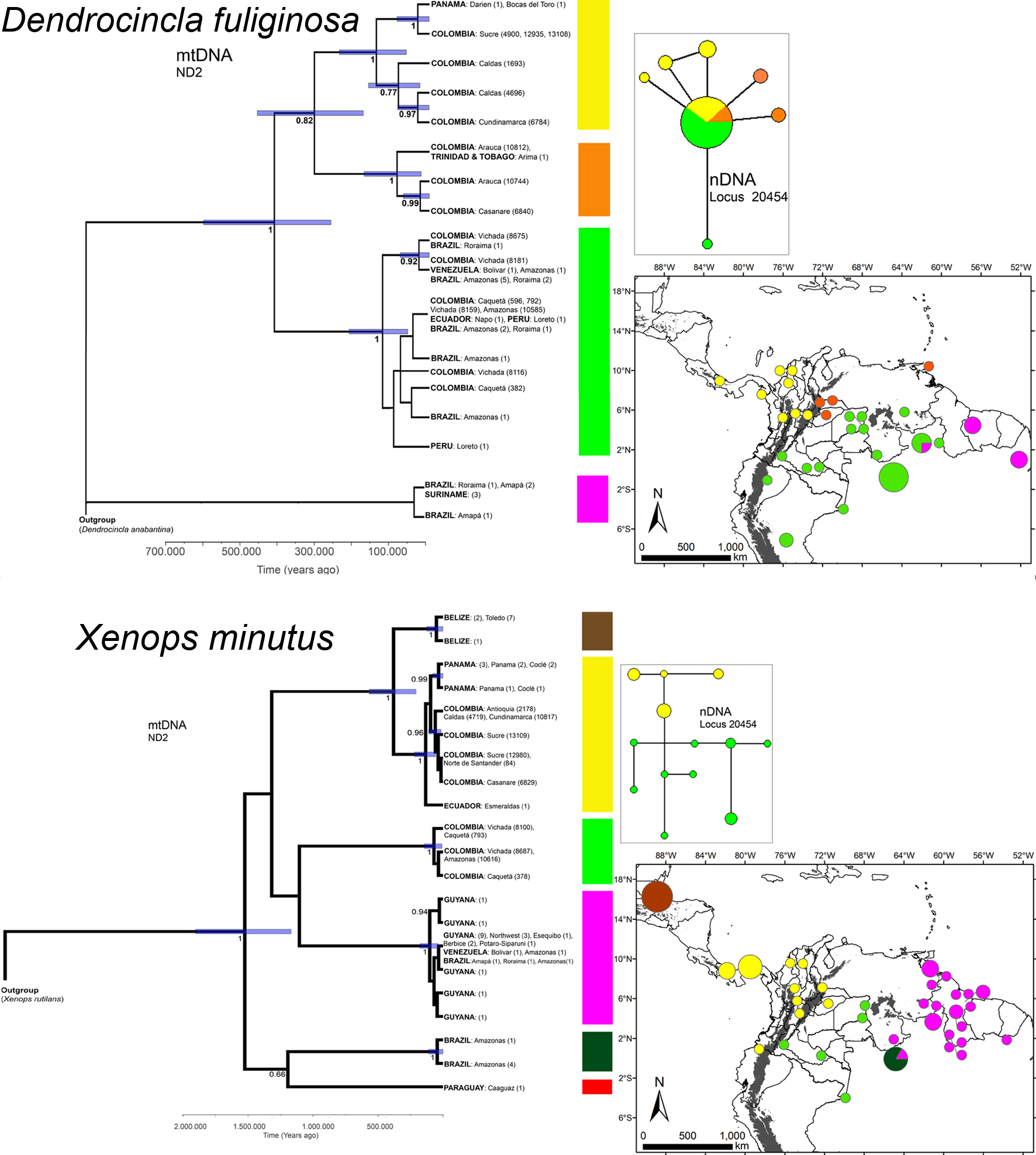
I gathered new mtDNA sequences (ND2) of 29 individuals of D. fuliginosa and X. minutus from Colombia, which I combined along with 82 sequences, publically available, from other regions (Appendix 1). mtDNA phylogeographic structure were found in both species across their ranges (Fig. 1). The 20454 nDNA locus, examined only for Colombian samples, presented different alleles on both species, and geographic structure seems to be present in X. minutus but not in D. fuliginosa (Fig. 1). The phylogenetic trees indicated that there are 6 mtDNA lineages for X. minutus and 4 mtDNA lineages for D. fuliginosa (Fig. 1).
The 4 D. fuliginosa mtDNA lineages exhibited phylogeographic structure (Fig. 1). In addition to the differentiated lineage from eastern South America, a widespread mtDNA lineage was found including the Amazonian individuals from Brazil, Venezuela, Colombia, Ecuador, and Peru. This Amazonian lineage was sister to the clade including 2 sister lineages, one of these includes the Colombian individuals from the Orinoquia and 1 individual from Trinidad and Tobago, while the trans-Andean individuals from Colombia and Panama comprised the other lineage. The Orinoquian individuals (cis-Andean) were sister to the trans-Andean lineage, rather than with the Amazon (cis-Andean) lineage. The network representation of nDNA alleles (Fig. 1) did not show such structure. However, only the most common allele of the 20454 nDNA locus was shared by individuals from the 3 mtDNA lineages, while the other 6 alleles were exclusive of different mtDNA lineages (Fig. 1). Besides these general patterns in D. fuliginosa, it is also notable that the trans-Andean individuals showed geographic differences in their mtDNA haplotypes. For instance, Colombian inter Andean valley’s individuals did not share haplotypes with Caribbean individuals, and these Caribbean individuals in spite to be close to Panama individuals harbor a different haplotype (Fig. 1).
The 6 X. minutus mtDNA lineages also exhibited phylogeographic structure (Fig. 1). One individual from Paraguay (not depicted in the map in figure 1) and 5 individuals from the Amazon in Brazil (but without locality information) comprise a clade with posterior probability (pp) = 0.66 that is sister to the poorly supported clade (pp = 0.34) including the remaining individuals. One mtDNA lineage including individuals from Venezuela, Brazil, and Guyana appeared as sister (pp = 0.81) to a lineage including individuals from Colombian Amazon. On the other hand, the lineage with individuals from Belize is a sister to a lineage including individuals from Panama, western Ecuador, Colombia (trans-Andean and Orinoquia), and northwestern Venezuela. Colombian individuals from Orinoquia (cis-Andean) are again clustered with trans-Andean individuals and not with the other cis-Andean individuals from the Amazon. This result for X. minutus is paralleled by the Network analysis of the nDNA (Fig. 1) that showed that individuals from the Orinoquia (cis-Andean) shared alleles with other Colombian trans-Andean individuals, while individuals from the Colombian Amazon (cis-Andean) not share alleles with the remaining Colombian individuals. As in the case of D. fuliginosa, the trans-Andean individuals of X. minutus also showed geographic differences in their haplotypes. The Belize haplotypes were well differentiated, while the Ecuador, Panama, and Colombia individuals did not share haplotypes among them. In addition, Colombian individuals from the inter-Andean valleys harbor 1 haplotype that is different from those found in the Caribbean Colombian individuals.
The BEAST analysis (Fig. 1) indicated that divergence of mtDNA lineages in D. fuliginosa began approximately 0.5 my (95% HPD between 266,000 and 608,000 years ago), while divergence of lineages in X. minutus was older, beginning approximately 1.5 my (95% HPD between 1.17 and 1.9 myA). Haplotypes within each mtDNA lineage of D. fuliginosa appear to have coalesced during the last 150,000 years, whereas the mtDNA lineages of X. minutus coalesced during the last 370,000 years. Regarding the Colombian haplogroups, the lineage of D. fuliginosa in the Orinoquia diverged from its sister trans–Andean lineage approximately 300,000 years ago, and both of them diverged from the Amazon lineage around 400,000 years ago (Fig. 1). In the case of X. minutus, the divergence of the 2 lineages present in Colombia occurred probably more than 1 million years ago, but the alleles comprising a clade for the Orinoquia (Andean) and trans-Andean samples likely coalesced less than 1 my.
Discussion
Molecular information for X. minutus and D. fuliginosa was analyzed, and I agree with other authors (e.g., Avendaño et al., 2017; Cadena et al., 2007) in considering the inclusion of samples from Colombia as necessary to draw conclusions about recent biological diversification in the Neotropics, which was the focus of this paper for 2 species of passerine birds. Both species exhibited phylogeographic structure in their whole geographic distribution, suggesting the occurrence of historic processes (i.e., vicariance) separating their populations. Regarding Colombian samples both species show a clear separation of individuals from the Amazon (cis-Andean), while individuals from the Orinoquia (cis-Andean) and the trans-Andean lowlands are more related. These patterns agree with the biogeographic regionalization of the Neotropics which indicates that the Orinoquia and trans–Andean lowlands conform a biogeographic dominion, which is different from the Amazon (Morrone, 2014), but contrast with the idea of the Andes as a barrier for lowland species.
The uplift of the Andes as a process associated with differentiation between Orinoquia (Andean) and trans-Andean populations of D. fuliginosa and X. minutus can be ruled out because such mountains reached their actual elevation in Colombia 2.7 myA (Gregory-Wodzicki, 2000), while populations of D. fuliginosa and X. minutus on both sides probably diverged less than 0.5 myA (95% HPD between 450,000 – 175,000 years ago for D. fuliginosa, and less than 250,000 years ago for X. minutus). However, I do not discard the presence of the Andes as related to genetic differentiation, in the sense that its mountains have facilitated isolation of populations that were connected recently (a possible scenario for D. fuliginosa) or have promoted differentiation in the trans-Andean zone followed by dispersal over (or around) these mountains (a possible scenario for X. minutus). Another possible scenario for X. minutus is that their populations in the Orinoquia and the trans-Andean populations have been connected throughout their history because they are part of the same mtDNA lineage.
Close relationships between Orinoquia and trans–Andean lowlands have been also revealed by biogeographical and ecological analyses of other animal taxa (Lozano-Zambrano & Fernández, 2007; Sigrist & Carvalho, 2009). Similarly, this relationship between cis- and trans-Andean populations has been found in other phylogeographic studies. For instance, in monkey species (Lynch et al., 2015) and another bird (Chrysomus icterocephalus, Cadena et al., 2011). Weir & Price (2011) also noted the recent separation of D. fuliginosa ridgwayi (trans-Andean) from D. f. meruloides (cis–Andean). Several additional works have found sister relationships between populations in Central America or Choco, and populations in Amazonia (e.g., Marks et al., 2002; Miller et al., 2008; Nyári, 2007; Rheindt et al., 2009), but they did not include Colombian samples that I show here are key to a complete understanding of these lineages’ phylogeographic patterns.
Close phylogeographic relationships between cis– and trans-Andean populations are not new for vertebrates (Lynch et al., 2015; Nyári, 2007). However, another result is more novel. I found that the major phylogeographic break for D. fuliginosa and X. minutus in Colombia is between Orinoquia and Amazonia (both cis-Andean regions), and not among cis- and trans-Andean populations. This phylogeographic break (Amazon/Orinoquia) has been also found in monkeys (Lagothrix lagotricha, Botero et al., 2015), and is a noteworthy result, suggesting that ecological differences between Orinoquia and Amazon could be involved in maintaining genetic isolation, which is in line with Brumfield (2012) who considers a secondary role of barriers in the Neotropics in structuring variation that arose elsewhere, and that could be in secondary contact (in this case) in the border between Amazon and Orinoquia. An alternative explanation for this phylogeographic pattern is to consider a major role for the ecological shifts between Orinoquia and Amazonia as the reason for genetic differentiation in these species. Whatever the explanation (either as a zone of division or as a zone of contact); the Amazon/Orinoquia transition must be considered as a necessary factor to explain genetic differentiation in widespread lowland taxa in the Neotropics.
In spite of the large genomic information used by Harvey and Brumfield (2015), which allowed understand several issues of the history of X. minutus, the lack of Colombian individuals in their analyses made impossible to appreciate that individuals from Orinoquia (cis-Andean) and from the trans-Andean zone are included in the same lineage. However, Harvey and Brumfield (2015) found evidence of migration from the Amazon into the trans-Andean populations. The close relationship of cis- and trans-Andean populations of birds inhabiting forests suggests that lowlands on both sides of the Andes in northwestern South America have (or recently had) a connection. This observation agrees with the scenario proposed by Haffer (1967), who considered that during glacial periods of the Pleistocene, the climate in northern Colombia was more humid, and then these lowlands were forest-covered much more extensively than they are at present, which provided a broad connection between cis- and trans-Andean lowland forests. This effect was increased by a lower sea level (about 100 m), which promoted the emergence of a large area of land in northern South America (Haffer, 1967; Nores, 2004). For Ateles E. Arbeláez-Cortés / Revista Mexicana de Biodiversidad 91 (2020): e912984 7 https://doi.org/10.22201/ib.20078706e.2020.91.2984
monkeys, this scenario has been considered to explain their distribution in cis– and trans-Andean zones (Lynch et al., 2015). However, a northern connection between cis– and trans-Andean regions is not the only way supported by analyses of species ranges in lowland species (Cadena et al., 2016). In addition, Avendaño et al. (2013) presented evidence of recent crossings of cis-Andean bird species into the trans-Andean lowlands, apparently promoted by human modifications to the landscape, suggesting that ecological changes could facilitate the interchange of some species between both zones.
Considering only the trans-Andean samples, the differences in their mtDNA haplotypes are noteworthy, particularly these observed between the inter-Andean valleys and the Caribbean Colombian populations that probably occurred during the last 250,000 years. This result parallels the result for C. icterocephalus that also shows different haplotypes between the Magdalena valley and the Caribbean lowlands (Cadena et al., 2011), but contrasts with the lack of genetic structure found in the monkey Ateles hybridus and other bird species in this same area (Link et al., 2015; Sandoval et al., 2017). Differences found in the 2 species analyzed here could be explained by historic processes of isolation among trans-Andean populations due to climatic fluctuations. For instance, the repeated change of dry and wet climatic periods during the Pleistocene (Haffer, 1967) and fluctuations in sea level in the Caribbean coastal zone (Nores, 2004) could be related to population isolation in trans-Andean lowlands, thus allowing the origin of the genetic patterns observed. Further phylogeographic studies are warranted to depict detailed phylogeographic patterns in this zone and to test the role of these paleoclimatic processes in the genetic differentiation of populations.
It is also noteworthy that trans-Andean samples show genetic differences which contrast with an apparent lack of genetic differences among Amazonian samples. For instance, western Amazonian individuals clustered together, despite that they are widespread and represent localities in 2 or more endemism areas (Cracraft, 1985). Colombian individuals of both species, in this study, from the Napo and Imeri endemism areas shared haplotypes, indicating that these areas are not separable based on the molecular information. For D. fuliginosa individuals from Venezuela (Guiana area of endemism) and Peru (Inambari) harbor haplotypes like the ones found in the Colombian Amazon (Imerí and Napo). These results suggest a more active (and recent) genetic differentiation in the trans-Andean lowlands than in the Amazon.
The Colombian Orinoquia (cis-Andean) and Caribbean (trans-Andean) are 2 gaps in biodiversity knowledge (Arbeláez-Cortés, 2013). The Caribbean and the inter-Andean valleys in Colombia have undergone major anthropogenic changes in their landscapes (Armenteras et al., 2011; Etter et al., 2006), while the Orinoquia is undergoing significant changes in its natural ecosystems (Armenteras et al., 2005; Romero-Ruiz et al., 2012). Therefore, it is urgent to better understand the biodiversity in these zones to have scientific support for its conservation. Further works should test if the close phylogeographic relationship between trans-Andean and Orinoquia (cis-Andean) populations regarding the Amazon (cis-Andean) populations is a generality for birds and other taxa. If such pattern is common, then the genetic differences among populations in this region of the Neotropics will not be explained only by the presence of Andean mountains or large rivers, but also by the ecological shift between ecosystems such as those represented by Amazonia and Orinoquia or by an historical barrier to gene flow between ecosystems that has shifted.
Acknowledgments
This work was supported by the Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Colombia, under funds allocated to Colecciones Biológicas; and by the Vicerrectoría de Investigación y extensión of Universidad Industrial de Santander, Colombia under grant “Proyecto de investigación capital semilla 2300”. I also thank the family Arbeláez-Cortés and A. G. Navarro-Sigüenza for their support. L. A. Sánchez González, N. Trujillo-Arias, and two anonymous reviewers made valuable comments and corrections that improved this work. A special acknowledgment to the specimen and tissue collectors of the material used in this study. Samples were exported to sequencing overseas under: “Permiso para la exportación de especímenes de la diversidad biológica no listados en los apéndices de la convención CITES número 385 de 2014 de la Autoridad Nacional de Licencias Ambientales, Colombia”.
Appendix 1. List of individuals of D. fuliginosa and X. minutus with GenBank accession numbers and locality information for samples and sequences included in this work. Source of the sequences information is indicated.
|
Taxon |
Collection number |
GenBank number: ND2 / 20454 |
Country |
Department or State |
County |
Locality |
Source |
|
D. fuliginosa |
STRI TR-DFU1 |
GU215371 |
Trinidad and Tobago |
Arima |
Simla Research Station |
Weir et al. (2009) |
|
|
D. fuliginosa |
IAvH-CT 4900, IAvH-A 13106 |
MN486096 / MN486131 |
Colombia |
Sucre |
San Onofre |
Corregimiento La Barce, Santuario de Fauna y Flora El Corchal El Mono Hernandez |
This study |
|
D. fuliginosa |
IAvH-CT 13108, IAvH-A 15248 |
MN486112 / MN486143 |
Colombia |
Sucre |
Toluviejo |
Corregimiento El Cañito, Monte de Los Navas |
This study |
|
D. fuliginosa |
IAvH-CT 12935, IAvH-A 15075 |
MN486102 / MN486141 |
Colombia |
Sucre |
Colosó |
Estación Primatologica, Montes de María |
This study |
|
D. fuliginosa |
IAvH-CT 12997 |
/ MN486142 |
Colombia |
Sucre |
This study |
||
|
D. fuliginosa |
STRI JTW253 |
GU215373 |
Panama |
Bocas del Toro |
Valle de Risco |
Not locality information-Valle de Risco |
Weir et al. (2009) |
|
D. fuliginosa |
STRI JTW744 |
GU215374 |
Panama |
Darien |
Puerto Piña |
Not locality information-Puerto Piña |
Weir et al. (2009) |
|
D. fuliginosa |
IAvH-CT 10744, IAvH-A 14722 |
MN486109 / MN486139 |
Colombia |
Arauca |
Arauquita |
Extracciòn de Arenas Remolinos, Rìo Arauca |
This study |
|
D. fuliginosa |
IAvH-CT 10812, IAvH-A 14795 |
MN486107 / MN486140 |
Colombia |
Arauca |
Arauquita |
Plataforma Cosecha G |
This study |
|
D. fuliginosa |
AMNH 11848 |
JQ445335 |
Venezuela |
Bolivar |
Cerro Guaiquinima |
Naka et al. (2012) |
|
|
D. fuliginosa |
IAvH-CT 4696, IAvH-A 13282 |
MN486098 / MN486130 |
Colombia |
Caldas |
Vereda La Miel |
This study |
|
|
D. fuliginosa |
IAvH-CT 6840, IAvH-A 14175 |
MN486110 / MN486133 |
Colombia |
Casanare |
Pore |
Corregimiento Altamira, Vereda Altamira La Esperanza |
This study |
|
D. fuliginosa |
IAvH-CT 6784, IAvH-A 14146 |
MN486104 / MN486132 |
Colombia |
Cundinamarca |
Puerto Salgar |
Not locality information-Puerto Salgar |
This study |
|
Taxon |
Collection number |
GenBank number: ND2 / 20454 |
Country |
Department or State |
County |
Locality |
Source |
|
D. fuliginosa |
FMNH FM391298 |
GU215370 |
Brazil |
Amapá |
Not locality information-Amapá |
Weir et al. (2009) |
|
|
D. fuliginosa |
FMNH 391303 |
JQ445317 |
Brazil |
Amapá |
Not locality information-Amapá |
Naka et al. (2012) |
|
|
D. fuliginosa |
IAvH-CT 382, IAvH-A 11047 |
MN486099 / MN486126 |
Colombia |
Caquetá |
Parque Nacional Natural Serranía de Chiribiquete, Río Mesay |
This study |
|
|
D. fuliginosa |
INPA A1940 |
JQ445330 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
D. fuliginosa |
INPA A650 |
JQ445327 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
D. fuliginosa |
INPA A1230 |
JQ445333 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
D. fuliginosa |
INPA A1165 |
JQ445331 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
D. fuliginosa |
INPA A694 |
JQ445328 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
D. fuliginosa |
INPA A1220 |
JQ445323 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
D. fuliginosa |
INPA A2007 |
JQ445321 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
D. fuliginosa |
INPA A1596 |
JQ445320 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
D. fuliginosa |
INPA A1938 |
JQ445319 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
D. fuliginosa |
IAvH-CT 596, IAvH-A 11276 |
MN486105 / MN486127 |
Colombia |
Caquetá |
Parque Nacional Natural Serranía de Chiribiquete Río Sararamano, afluente del río Messay |
This study |
|
|
D. fuliginosa |
STRI EC-DFU1 |
GU215372 |
Ecuador |
Napo |
Jatun Sacha |
Not locality information-Jatun Sacha |
Weir et al. (2009) |
|
Taxon |
Collection number |
GenBank number: ND2 / 20454 |
Country |
Department or State |
County |
Locality |
Source |
|
D. fuliginosa |
IAvH-CT 10585, IAvH-A 14562 |
MN486111 / MN486138 |
Colombia |
Amazonas |
Leticia |
Reserva Forestal Rio Calderon, Estacion Biologica El Zafire |
This study |
|
D. fuliginosa |
LSUMZ B6895 |
JQ445325 |
Peru |
Loreto |
Not locality information-Loreto |
Naka et al. (2012) |
|
|
D. fuliginosa |
LSUMZ B2723 |
JQ445326 |
Peru |
Loreto |
Not locality information-Loreto |
Naka et al. (2012) |
|
|
X. minutus |
UAM 24522; ABJ415 |
FJ175798 |
Belize |
Toledo District |
Not locality information-Toledo District |
M.J. Miller, 2008 unpublished document |
|
|
X. minutus |
UAM ABJ428 |
FJ175799 |
Belize |
Not locality information-Belize |
M.J. Miller (2008) unpublished document |
||
|
X. minutus |
UAM ABJ622 |
FJ175800 |
Belize |
Not locality information-Belize |
M.J. Miller (2008) unpublished document |
||
|
X. minutus |
UAM ABJ607 |
FJ175801 |
Belize |
Not locality information-Belize |
M.J. Miller (2008) unpublished document |
||
|
X. minutus |
UAM 24355; ABJ294 |
FJ175802 |
Belize |
Toledo District |
Not locality information-Toledo District |
M.J. Miller (2008) unpublished document |
|
|
X. minutus |
UAM 24347; ABJ266 |
FJ175803 |
Belize |
Toledo District |
Not locality information-Toledo District |
M.J. Miller (2008) unpublished document |
|
|
X. minutus |
UAM 14330; KSW3706 |
FJ175804 |
Belize |
Toledo District |
Not locality information-Toledo District |
M.J. Miller (2008) unpublished document |
|
|
X. minutus |
UAM 24348; ABJ270 |
FJ175805 |
Belize |
Toledo District |
Not locality information-Toledo District |
M.J. Miller (2008) unpublished document |
|
|
X. minutus |
UAM 24357; ABJ296 |
FJ175806 |
Belize |
Toledo District |
Not locality information-Toledo District |
M.J. Miller (2008) unpublished document |
|
Taxon |
Collection number |
GenBank number: ND2 / 20454 |
Country |
Department or State |
County |
Locality |
Source |
|
X. minutus |
UAM 24356; ABJ295 |
FJ175807 |
Belize |
Toledo District |
Not locality information-Toledo District |
M.J. Miller (2008) unpublished document |
|
|
X. minutus |
IAvH-CT 13109, IAvH-A 15249 |
MN486118 / MN486154 |
Colombia |
Sucre |
Toluviejo |
Corregimiento El Cañito, Monte de Los Navas |
This study |
|
X. minutus |
IAvH-CT 12980, IAvH-A 15120 |
MN486123 / MN486153 |
Colombia |
Sucre |
Colosó |
Estación Primatologica, Montes de María |
This study |
|
X. minutus |
UAM 20495; MJM410 |
FJ175809 |
Panama |
Panama |
Cerro Azul: ANAM Station |
M.J. Miller (2008) unpublished document |
|
|
X. minutus |
UAM KSW4392 |
FJ175810 |
Panama |
Not locality information-Panama |
M.J. Miller (2008) unpublished document |
||
|
X. minutus |
UAM 20492; MJM407 |
FJ175812 |
Panama |
Not locality information-Panama |
M.J. Miller (2008) unpublished document |
||
|
X. minutus |
UAM MJM1462 |
FJ175814 |
Panama |
Not locality information-Panama |
M.J. Miller (2008) unpublished document |
||
|
X. minutus |
UAM 22105; MJM675 |
FJ175815 |
Panama |
Panama |
Not locality information-Cerro Azul: ANAM Station |
M.J. Miller (2008) unpublished document |
|
|
X. minutus |
UAM 22110 |
FJ175816 |
Panama |
Panama |
Not locality information-Cerro Azul: ANAM Station |
M.J. Miller (2008) unpublished document |
|
|
X. minutus |
UAM 20338; MJM244 |
FJ175808 |
Panama |
Cocle |
Molejon, Finca Moreno |
M.J. Miller (2008) unpublished document |
|
|
X. minutus |
UAM 20326; MJM232 |
FJ175811 |
Panama |
Cocle |
Molejon, Finca Moreno |
M.J. Miller (2008) unpublished document |
|
Taxon |
Collection number |
GenBank number: ND2 / 20454 |
Country |
Department or State |
County |
Locality |
Source |
|
X. minutus |
UAM 20350 |
FJ175813 |
Panama |
Cocle |
Molejon, Finca Moreno |
Molejon, Finca Moreno |
M.J. Miller (2008) unpublished document |
|
X. minutus |
USNM 14628 |
JQ445992 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
AMNH DOT11942 |
JQ445967 |
Venezuela |
Bolivar |
Not locality information-Bolivar |
Naka et al. (2012) |
|
|
X. minutus |
USNM 9164 |
JQ445982 |
Guyana |
Northwest |
Not locality information-Northwest |
Naka et al. (2012) |
|
|
X. minutus |
USNM 9333 |
JQ445983 |
Guyana |
Northwest |
Not locality information-Northwest |
Naka et al. (2012) |
|
|
X. minutus |
IAvH-CT 84, IAvH-A 10842 |
MN486114 / |
Colombia |
Norte de Santander |
Toledo |
Vereda El Diamante Río Negro Parque Nacional Natural Tamá |
This study |
|
X. minutus |
IAvH-CT 2178 |
MN486113 |
Colombia |
Antioquia |
Amalfi |
Vereda Las Animas, Bosque Las Animas, Cuenca de la quebrada Las Animas |
This study |
|
X. minutus |
USNM 14183 |
JQ445989 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
USNM 14260 |
JQ445990 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
USNM 4266 |
JQ445979 |
Guyana |
Berbice |
Not locality information-Berbice |
Naka et al. (2012) |
|
|
X. minutus |
USNM 4331 |
JQ445980 |
Guyana |
Berbice |
Not locality information-Berbice |
Naka et al. (2012) |
|
|
X. minutus |
IAvH-CT 4719, IAvH-A 13305 |
MN486115 / MN486146 |
Colombia |
Caldas |
Vereda La Miel |
This study |
|
|
X. minutus |
IAvH-CT 6829, IAvH-A 14164 |
MN486117 / MN486147 |
Colombia |
Casanare |
Pore |
Vereda Altamira, La Esperanza |
This study |
|
X. minutus |
USNM 5132 |
JQ445981 |
Guyana |
Essequibo |
Not locality information-Essequibo |
Naka et al. (2012) |
|
|
X. minutus |
IAvH-CT 8687, IAvH-A 12848 |
MN486124 / MN486150 |
Colombia |
Vichada |
Cumaribo |
Corregimineto Santa Rita, Parque Nacional Natural El Tuparro, Bosque de Guaipé |
This study |
|
Taxon |
Collection number |
GenBank number: ND2 / 20454 |
Country |
Department or State |
County |
Locality |
Source |
|
X. minutus |
USNM 15759 |
JQ445993 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
USNM 14525 |
JQ445991 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
LSUMZ B48433 |
JQ445975 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
LSUMZ B48452 |
JQ445976 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
LSUMZ B48478 |
JQ445977 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
UKNHM 1225 |
JQ445973 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
UKNHM 1276 |
JQ445974 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
IAvH-CT 8100, IAvH-A 14317 |
MN486121 / MN486148 |
Colombia |
Vichada |
Cumaribo |
Selva de Mataven,Caño Cajaro |
This study |
|
X. minutus |
IAvH-CT 8152, IAvH-A 14370 |
MN486125 / MN486149 |
Colombia |
Vichada |
Cumaribo |
Selva de Mataven,Caño Matavén |
This study |
|
X. minutus |
IAvH-CT 10817, IAvH-A 14208 |
MN486116 / MN486152 |
Colombia |
Cundinamarca |
Anapoima |
Club campestre Mesa de Yeguas |
This study |
|
X. minutus |
ANSP 7407 |
JQ445968 |
Guyana |
Potaro-Siparuni |
Not locality information-Potaro-Siparuni |
Naka et al. (2012) |
|
|
X. minutus |
INPA A1709 |
JQ445972 |
Brazil |
Roraima |
Not locality information-Roraima |
Naka et al. (2012) |
|
|
X. minutus |
USNM 12223 |
JQ445987 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
USNM 12772 |
JQ445988 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
AMNH:DOT8845 |
JF975328 |
Venezuela |
Amazonas |
Not locality information-Amazonas |
Derryberry et al. (2011) |
|
|
X. minutus |
FMNH 391346 |
JQ445969 |
Brazil |
Amapá |
Not locality information-Amapá |
Naka et al. (2012) |
|
|
X. minutus |
USNM 11810 |
JQ445986 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
USNM 10412 |
JQ445984 |
Guyana |
Not locality information-Guyana |
Naka et al. (2012) |
||
|
X. minutus |
USNM 10887 |
JQ445985 |
Guyana |
North West |
Not locality information-Northwest |
Naka et al. (2012) |
|
Taxon |
Collection number |
GenBank number: ND2 / 20454 |
Country |
Department or State |
County |
Locality |
Source |
|
X. minutus |
IAvH-CT 793, IAvH-A 11395 |
MN486119 / MN486145 |
Colombia |
Caquetá |
San José de Fragua |
Corregimiento La Esmeralda |
This study |
|
X. minutus |
LSUMZ:B11948 |
JF975329 |
Ecuador |
Esmeraldas |
El Placer |
Not locality information-El Placer |
Derryberry et al. (2011) |
|
X. minutus |
IAvH-CT 378, IAvH-A 11043 |
MN486120 / MN486144 |
Colombia |
Caquetá |
Parque Nacional Natural Serranía de Chiribiquete, Río Mesay |
This study |
|
|
X. minutus |
AMZ 118 (MPEG 59473) |
JQ445964 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
X. minutus |
AMZ 119 (MPEG 59474) |
JQ445965 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
X. minutus |
AMZ 325 (MPEG 59475) |
JQ445966 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
X. minutus |
INPA A71 |
JQ445970 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
X. minutus |
INPA A1610 |
JQ445971 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
X. minutus |
JAP 662 (MPEG 62739) |
JQ445978 |
Brazil |
Amazonas |
Not locality information-Amazonas |
Naka et al. (2012) |
|
|
X. minutus |
IAvH-CT 10616, IAvH-A 14593 |
MN486122 / MN486151 |
Colombia |
Amazonas |
Leticia |
Reserva Forestal Rio Calderon, Estacion Biologica El Zafire |
This study |
|
X. minutus |
LSUMZ:B25938 |
JF975330 |
Paraguay |
Caaguaz |
Not locality information-Caaguaz |
Derryberry et al. (2011) |
References
Arbeláez-Cortés, E. (2013). Knowledge of Colombian biodiversity: published and indexed. Biodiversity and Conservation, 22, 2875–2906. https://doi.org/10.1007/s10531-013-0560-y
Arbeláez-Cortés, E., Roldán-Piña, D., & Navarro-Sigüenza, A. G. (2014). Multilocus phylogeography and morphology give insights into the recent evolution of a Mexican endemic songbird: Vireo hypochryseus. Journal of Avian Biology, 45, 253–263. https://doi.org/10.1111/j.1600-048x.2013.00335.x
Arbogast, B. S., Drovetski, S. V., Curry, R. L., Boag, P. T., Seu-tin, G., Grant, P. R. et al. (2006). The origin and diversification of Galapagos mockingbirds. Evolution, 60, 370–382. https://doi.org/10.1554/03-749.1
Armenteras, D., Rodríguez, N., Retana, J., & Morales, M. (2011). Understanding deforestation in montane and lowland forests of the Colombian Andes. Regional Environmental Change, 11, 693–705. https://doi.org/10.1007/s10113-010-0200-y
Armenteras, D., Romero, M., & Galindo, G. (2005). Vegetation fire in the savannas of the Llanos Orientales of Colombia. World Resource Review, 17, 628–647.
Avendaño, J. E., Arbeláez-Cortés, E., & Cadena, C. D. (2017). On the importance of geographic and taxonomic sampling in phylogeography: A reevaluation of diversification and species limits in a Neotropical thrush (Aves, Turdidae). Molecular Phylogenetics and Evolution, 111, 87–97. https://doi.org/10.1016/j.ympev.2017.03.020
Avendaño, J. E., Cortés-Herrera, J. O., Briceño-Lara, E. R., & Rincón-Guarín, D. A. (2013). Crossing or bypassing the Andes: a commentary on recent range extensions of cis-Andean birds to the West of the Andes of Colombia. Orinoquia, 17, 208–214. https://doi.org/10.22579/20112629.18
Backström, N., Fagerberg, S., & Ellegren, H. (2008). Genomics of natural bird populations: a gene-based set of reference markers evenly spread across the avian genome. Molecular Ecology, 17, 964–980. https://doi.org/10.1111/j.1365-294x.2007.03551.x
Bandelt, H. J., Forster, P., & Röhl, A. (1999). Median-joining networks for inferring intraspecific phylogenies. Molecular Biology and Evolution, 16, 37–48. https://doi.org/10.1093/oxfordjournals.molbev.a026036
Benham, P. M., Cuervo, A. M., McGuire, J. A., & Witt, C. C. (2015). Biogeography of the Andean metaltail hummingbirds: contrasting evolutionary histories of tree line and habitat-generalist clades. Journal of Biogeography, 42, 763–777. https://doi.org/10.1111/jbi.12452
Botero, S., Stevenson, P. R., & Di Fiore, A. (2015). A primer on the phylogeography of Lagothrix lagotricha (sensu Fooden) in northern South America. Molecular Phylogenetics and Evolution, 82, 511–517. https://doi.org/10.1016/j.ympev.2014.05.019
Brumfield, R. T. (2012). Inferring the origins of lowland Neotropical birds. Auk, 129, 367–376. https://doi.org/10.1525/auk.2012.129.3.367
Cadena, C. D. (2007). Testing the role of interspecific competition in the evolutionary origin of elevational zonation: An example with Buarremon brush-finches (Aves, Emberizidae) in the Neotropical mountains. Evolution, 61, 1120–1136. https://doi.org/10.1111/j.1558-5646.2007.00095.x
Cadena, C. D., Gutiérrez-Pinto, N., Dávila, N., & Chesser, R. T. (2011). No population genetic structure in a widespread aquatic songbird from the Neotropics. Molecular Phylogenetics and Evolution, 58, 540–545. https://doi.org/10.1016/j.ympev.2010.12.014
Cadena, C. D., Klicka, J., & Ricklefs, R. E. (2007). Evolutionary differentiation in the Neotropical montane region: Molecular phylogenetics and phylogeography of Buarremon brush-finches (Aves, Emberizidae). Molecular Phylogenetics and Evolution, 44, 993–1016. https://doi.org/10.1016/j.ympev.2006.12.012
Cadena, C. D., Pedraza, C. A., & Brumfield, R. T. (2016). Climate, habitat associations and the potential distributions of Neotropical birds: Implications for diversification across the Andes. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 40, 275–287. https://doi.org/10.18257/raccefyn.280
Claramunt, S., Derryberry, E. P., Chesser, R. T., Aleixo, A., & Brumfield, R. T. (2010). Polyphyly of Campylorhamphus, and description of a new genus for C. pucherani (Dendrocolaptinae). Auk, 127, 430–439. https://doi.org/10.1525/auk.2009.09022
Cracraft, J. (1985). Historical biogeography and patterns of diversification within the South American areas of endemism. In P.A. Buckley, M.S, F., E.S. Morton, R.S. Ridgely & F.G. Buckley (Eds.) Ornithological Monographs (pp. 49–84). The American Ornithologist Union. https://doi.org/10.2307/40168278
Cheviron, Z. A., Hackett, S. J., & Capparella, A. P. (2005). Complex evolutionary history of a Neotropical lowland forest bird (Lepidothrix coronata) and its implications for historical hypotheses of the origin of Neotropical avian diversity. Molecular Phylogenetics and Evolution, 36, 338–357. https://doi.org/10.1016/j.ympev.2005.01.015
Del Hoyo, J., Elliot, A., & Christie, D. A. (2003). Handbook of the Birds of the World. Broadbills to Tapaculos. Lynx Edicions, Barcelona.
Derryberry, E. P., Claramunt, S., Derryberry, G., Chesser, R. T., Cracraft, J., Aleixo, A. et al. (2011). Lineage diversification and morphological evolution in a large-scale continental radiation: The Neotropical ovenbirds and woodcreepers (Aves: Furnariidae). Evolution, 65, 2973–2986. https://doi.org/10.1111/j.1558-5646.2011.01374.x
Drummond, A. J., Rambaut, A., & Suchard, M. (2012). BEAST v1.7.4 Bayesian Evolutionary Analysis Sampling Trees. Department of Computer Science University of Auckland, Institute of Evolutionary Biology University of Edinburgh, David Geffen School of Medicine University of California, Los Angeles, Available at: http://beast.bio.ed.ac.uk,
Drummond, A. J., Suchard, M. A., Xie, D., & Rambaut, A. (2012). Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Molecular Biology and Evolution, 29, 1969–1973. https://doi.org/10.1093/molbev/mss075
Etter, A., McAlpine, C., Wilson, K., Phinn, S., & Possingham, H. (2006). Regional patterns of agricultural land use and deforestation in Colombia. Agriculture, Ecosystems and Environment, 114, 369–386. https://doi.org/10.1016/j.agee.2005.11.013
Fernandes, A. M., Wink, M., Sardelli, C. H., Aleixo, A., & Ebach, M. (2014). Multiple speciation across the Andes and throughout Amazonia: the case of the spot-backed antbird species complex (Hylophylax naevius/Hylophylax naevioides). Journal of Biogeography, 41, 1094–1104. https://doi.org/10.1111/jbi.12277
Flantua, S. A., van Boxel, J., Hooghiemstra, H., & van Smaalen, J. (2007). Application of GIS and logistic regression to fossil pollen data in modelling present and past spatial distribution of the Colombian savanna. Climate Dynamics, 29, 697–712. https://doi.org/10.1007/s00382-007-0276-3
Fuchs, J., Chen, S., Johnson, J. A., & Mindell, D. P. (2011). Pliocene diversification within the South American Forest falcons (Falconidae: Micrastur). Molecular Phylogenetics E. Arbeláez-Cortés / Revista Mexicana de Biodiversidad 91 (2020): e912984 17 https://doi.org/10.22201/ib.20078706e.2020.91.2984
and Evolution, 60, 398–407. https://doi.org/10.1016/j.ympev.2011.05.008
GeoNames. (2013). GeoNames geographical database, Retrieved January 15, 2013, from: http://www.geonames.org/
Gill, F., & Donsker, D. (2018). IOC World Bird Names v 8.2, Retrieved September 30, 2018, from: http://www.worldbirdnames.org
Google. (2010). Google Earth. http://earth.google.com/web/.
Gregory-Wodzicki, K. M. (2000). Uplift history of the Central and Northern Andes: A review. Geological Society of America Bulletin, 112, 1091–1105. https://doi.org/10.1130/0016-7606(2000)112<1091:uhotca>2.3.co;2
Haffer, J. (1967). Speciation in Colombian forest birds west of the Andes. American Museum Novitates, 1–57.
Hall, T.A. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41, 95–98.
Harvey, M. G., & Brumfield, R. T. (2015). Genomic variation in a widespread Neotropical bird (Xenops minutus) reveals divergence, population expansion, and gene flow. Molecular Phylogenetics and Evolution, 83, 305–316. https://doi.org/10.1016/j.ympev.2014.10.023
Johnson, K. P., & Weckstein, J. D. (2011). The Central American land bridge as an engine of diversification in New World doves. Journal of Biogeography, 38, 1069–1076. https://doi.org/10.1111/j.1365-2699.2011.02501.x
Lerner, H. R., Meyer, M., James, H. F., Hofreiter, M., & Fleischer, R. C. (2011). Multilocus resolution of phylogeny and timescale in the extant adaptive radiation of Hawaiian Honeycreepers. Current Biology, 21, 1838–1844. https://doi.org/10.1016/j.cub.2011.09.039
Librado, P., & Rozas, J. (2009). DNAsp v5: a software for comprehensive analysis of DNA polimorphism data. Bioinformatics, 25, 1451–1452. https://doi.org/10.1093/bioinformatics/btp187
Link, A., Valencia, L. M., Céspedes, L. N., Duque, L. D., Cadena, C. D., & Fiore, A. D. (2015). Phylogeography of the critically endangered brown spider monkey (Ateles hybridus): Testing the riverine barrier hypothesis. International Journal of Primatology, 36, 530–547. https://doi.org/10.1007/s10764-015-9840-6
Lozano-Zambrano, F. H., & Fernández, F. (2007). Aproximación al análisis zoogeográfico de las hormigas cazadoras (Hymenoptera: Formicidae) de Colombia. Boletín del Museo de Entomología de la Universidad del Valle, 8, 22–31. https://doi.org/10.2307/3495802
Lynch, J. W., Cortés-Ortiz, L., Di Fiore, A., & Boubli, J. P. (2015). Special issue: comparative biogeography of Neotropical primates. Molecular Phylogenetics and Evolution, 82, 518–529. https://doi.org/10.1016/j.ympev.2014.09.027
Marks, B. D., Hackett, S. J., & Capparella, A. P. (2002). Historical relationships among Neotropical lowland forest areas of endemism as determined by mitochondrial DNA sequence variation within the Wedge-billed Woodcreeper (Aves: Dendrocolaptidae: Glyphorynchus spirurus). Molecular Phylogenetics and Evolution, 24, 153–167. https://doi.org/10.1016/s1055-7903(02)00233-6
Miller, M. J., Bermingham, E., Klicka, J., Escalante, P., Amaral, F. S. R. d., Weir, J.T. et al. (2008). Out of Amazonia again and again: episodic crossing of the Andes promotes diversification in a lowland forest flycatcher. Proceedings of the Royal Society B: Biological Sciences, 275, 1133–1142. https://doi.org/10.1098/rspb.2008.0015
Morrone, J. J. (2014). Biogeographical regionalisation of the Neotropical region. Zootaxa, 3782, 1–110. https://doi.org/10.11646/zootaxa.3782.1.1
Naka, L. N., Bechtoldt, C. L., Henriques, L. M. P., & Brumfield, R. T. (2012). The role of physical barriers in the location of avian suture zones in the Guiana Shield, northern Amazonia. The American Naturalist, 179, E115–E132. https://doi.org/10.1086/664627
Nores, M. (2004). The implications of Tertiary and Quaternary sea level rise events for avian distribution patterns in the lowlands of northern South America. Global Ecology and Biogeography, 13, 149–161. https://doi.org/10.1111/j.1466-882x.2004.00076.x
Nyári, Á. S. (2007). Phylogeographic patterns, molecular and vocal differentiation, and species limits in Schiffornis turdina (Aves). Molecular Phylogenetics and Evolution, 44, 154–164. https://doi.org/10.1016/j.ympev.2007.02.020
Patel, S., Weckstein, J. D., Patané, J. S. L., Bates, J. M., & Aleixo, A. (2011). Temporal and spatial diversification of Pteroglossus araçaris (Aves: Ramphastidae) in the neotropics: Constant rate of diversification does not support an increase in radiation during the Pleistocene. Molecular Phylogenetics and Evolution, 58, 105–115. https://doi.org/10.1016/j.ympev.2010.10.016
Posada, D., & Crandall, K. A. (1998). Modeltest: testing the model of DNA substitution. Bioinformatics, 14, 817–818. https://doi.org/10.1093/bioinformatics/14.9.817
Rambaut, A., & Drummond, A. J. (2012). TreeAnnotator v1.7.4. Institute of Evolutionary Biology, University of Edinburgh, Department of Computer Science, University of Auckland, Available as part of the BEAST package at http://beast.bio.ed.ac.uk/
Restall, R., Rodner, C., & Lentino, M. (2006). Birds of Northern South America: an identification guide. Yale University Press, New Haven.
Rheindt, F. E., Christidis, L., Cabanne, G. S., Miyaki, C., & Norman, J. A. (2009). The timing of Neotropical speciation dynamics: A reconstruction of Myiopagis flycatcher diversification using phylogenetic and paleogeographic data. Molecular Phylogenetics and Evolution, 53, 961–971. https://doi.org/10.1016/j.ympev.2009.09.001
Romero-Ruiz, M. H., Flantua, S. G. A., Tansey, K., & Berrio, J. C. (2012). Landscape transformations in savannas of northern South America: Land use/cover changes since 1987 in the Llanos Orientales of Colombia. Applied Geography, 32, 766–776. https://doi.org/10.1016/j.apgeog.2011.08.010
Sandoval, H., J., Gómez, J. P., & Cadena, C. D. (2017). Is the largest river valley west of the Andes a driver of E. Arbeláez-Cortés / Revista Mexicana de Biodiversidad 91 (2020): e912984 18 https://doi.org/10.22201/ib.20078706e.2020.91.2984
diversification in Neotropical lowland birds? Auk, 137, 168–180. https://doi.org/10.1642/auk-16-91.1
Schultz, E. D., Burney, C. W., Brumfield, R. T., Polo, E. M., Cracraft, J., & Ribas, C. C. (2017). Systematics and biogeography of the Automolus infuscatus complex (Aves; Furnariidae): Cryptic diversity reveals western Amazonia as the origin of a transcontinental radiation. Molecular Phylogenetics and Evolution, 107, 503–515. https://doi.org/10.1016/j.ympev.2016.12.023
Sigrist, M. S., & Carvalho, C. J. B. d. (2009). Historical relationships among areas of endemism in the tropical South America using Brooks Parsimony Analysis (BPA). Biota Neotropica, 9, 79–90. https://doi.org/10.1590/s1676-06032009000400009
Smith, B. T., Harvey, M. G., Faircloth, B. C., Glenn, T. C., & Brumfield, R. T. (2014). Target capture and massively parallel sequencing of ultraconserved elements for comparative studies at shallow evolutionary time scales. Systematic Biology, 63, 83–95. https://doi.org/10.1093/sysbio/syt061
Smith, B. T., McCormack, J. E., Cuervo, A. M., Hickerson, M. J., Aleixo, A., Cadena, C. D. et al. (2014). The drivers of tropical speciation. Nature, 515, 406–409. https://doi.org/10.1038/nature13687
Sparrman, A. (1788). Museum Carlsonianum, in quo novas et selectas aves, coloribus ad vivum brevique descriptione illustratas, suasu et sumtibus generosissimi possessoris. Holmiae: Ex Typographia Regia. Biodiversity Heritage Library. https://doi.org/10.5962/bhl.title.62901
Stephens, M., & Donnelly, P. (2003). A comparison of Bayesian methods for haplotype reconstruction from population genotype data. American Journal of Human Genetics, 73, 1162–1169. https://doi.org/10.1086/379378
Stephens, M., Smith, N. J., & Donnelly, P. (2001). A new statistical method for haplotype reconstruction from population data. The American Journal of Human Genetics, 68, 978–989. https://doi.org/10.1086/319501
Vieillot, L. J. P. (1818). Nouveau dictionnaire d’Histoire Naturelle appliquée aux arts , à l’agriculture, à l’économie rurale et domestique, à la médecine, etc. Paris. https://doi.org/10.5962/bhl.title.51404
Weir, J. T., Bermingham, E., & Schluter, D. (2009). The great American biotic interchange in birds. Proceedings of the National Academy of Sciences, 106, 21737–21742. https://doi.org/10.1073/pnas.0903811106
Weir, J. T., & Price, M. (2011). Andean uplift promotes lowland speciation through vicariance and dispersal in Dendrocincla woodcreepers. Molecular Ecology, 20, 4550–4563. https://doi.org/10.1111/j.1365-294x.2011.05294.x
Weir, J. T., & Schluter, D. (2008). Calibrating the avian molecular clock. Molecular Ecology, 17, 2321–2328. https://doi.org/10.1111/j.1365-294x.2008.03742.x


