
Angel Rolando Endara-Agramont a, *, Rosa Laura Heredia-Bobadilla a, Luis Antonio García-Almaraz a, Alma Abigail Luna-Gil a, Sergio Franco-Maass a, Víctor David Cibrián-Llanderal b
a Universidad Autónoma del Estado de México, Instituto de Ciencias Agropecuarias y Rurales, Instituto Literario Núm. 100, Colonia Centro, 50000 Toluca, Estado de México, México
b Colegio de Postgraduados Campus Montecillo, Cátedras Conacyt-Postgrado en Ciencias Forestales, Carretera México-Texcoco Km. 36.5, Montecillo, 56230 Texcoco, Estado de México, México
*Autor para correspondencia: arendaraa@uaemex.mx (A.R. Endara-Agramont)
Recibido: 22 abril 2022; aceptado: 5 agosto 2022
Resumen
Los muérdagos (Arceuthobium globosum y A. vaginatum) afectan los bosques de Pinus hartwegii, se distribuyen en las partes altas de las montañas de México (3,000-4,200 m snm). El objetivo del estudio fue determinar los factores asociados con la presencia y niveles de infestación de dichos muérdagos en 2 montañas de México. Para ello, se establecieron 1,622 sitios de muestreo en el Área de Protección de Flora y Fauna Nevado de Toluca y 634 en Monte Tláloc, donde se registró la presencia y grado de infestación de la regeneración e individuos adultos (≥ 7.5 cm de DN). Se obtuvieron datos de localización de los sitios de muestreo, altitud, pendiente, orientación de ladera, dominancia de los estratos herbáceo y arbustivo, presencia de individuos muertos y la infestación por descortezador (Dendroctonus adjunctus). Posteriormente, se realizó una prueba de correlación para identificar los factores relacionados con la presencia de muérdagos, además de un análisis de componentes principales. Esta investigación reveló que 37% de la superficie forestal se encuentra infestada por muérdagos enanos; la presencia de descortezadores y la orientación del sitio son los principales factores que inciden en la mayor infestación de muérdagos en los bosques del área de estudio.
Palabras clave: Coníferas; Parásitos; Pino de las alturas; Regeneración; Niveles de infestación
© 2022 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Factors associated with the spatial distribution of dwarf mistletoes in two Pinus hartwegii populations in Central México
Abstract
Dwarf mistletoes (Arceuthobium globosum and A. vaginatum) represent a threat to the Pinus hartwegii forests distributed in the high parts of mountains in Mexico (3,000-4,200 m asl). The objective of the study was to determine the factors associated with the presence and levels of infestation of such mistletoes in 2 mountains of Mexico. Two thousand, two hundred and fifty-four sampling sites were established, 1,622 in the Nevado de Toluca Flora and Fauna Protection Area and 634 in Monte Tláloc, to register the presence and degree of infestation of regeneration and adult individuals (≥ 7.5 cm DN). The data obtained in the sampling sites included geographical location, altitude, slope, slope orientation, the dominance of the herbaceous and shrubby strata, presence of dead or damaged individuals and infestation by bark beetle (Dendroctonus adjunctus). A correlation test was performed to identify the factors related with the presence of mistletoes, in addition to a principal component analysis. This study revealed that dwarf mistletoes infest 37% of the forest area; the presence of bark beetles and the site orientation are the main factors that influence on the infestation of mistletoes in the studied forests.
Keywords: Conifers; Parasites; High mountain pine; Regeneration; Infestation levels
© 2022 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introducción
Los muérdagos enanos son plantas hemiparásitas (parasitismo parcial) del género Arceuthobium; México es centro de diversidad del género, pues alberga 20 de las 42 especies reportadas en el mundo, y de éstas, 12 son endémicas (Geils et al., 2002; Queijeiro-Bolaños y Cano-Santana, 2016); en México parasitan por lo menos a 13 especies del género Pinus, lo cual sugiere una capacidad de infestación alta (Varo-Rodríguez et al., 2016).
Los bosques templados ocupan 15% del territorio mexicano y se distribuyen en 5 regiones principales: Península de Baja California, Sierra Madre Occidental, Sierra Madre Oriental, Sierra Madre del Sur, y Faja Volcánica Transmexicana (FVTM) (Sánchez-González, 2008). El Área de Protección de Flora y Fauna Nevado de Toluca (APFFNT) y el Parque Nacional Izta-Popo Zoquiapan y Anexas (PNIPZA) se localizan en la FVTM; dentro del PNIPZA se encuentra el monte Tláloc (MT), que constituye el séptimo pico más alto del país con una altitud de 4,125 m snm (Iwaniszewski, 1994) y el APFFNT, considerada la cuarta montaña más alta de México, alcanzando una altitud de 4,660 m snm (Montero-García, 2004). Tanto el MT como el APFFNT se caracterizan por la presencia de los bosques del pino de las alturas (Pinus hartwegii Lindl.).
Los bosques de P. hartwegii son de gran importancia ecológica, presentan tolerancia a temperaturas bajas extremas, marcan el límite altitudinal superior arbóreo (4,000-4,200 m snm), contribuyen en la captura de carbono y a la formación de suelo (Rojas et al., 2019), además de que constituyen el hábitat de diferentes especies de fauna endémica, como el teporingo (Romerolagus diazi Ferrari-Pérez) y ajolotes de arroyos de alta montaña (Ambystoma spp.), siendo así proveedores importantes de servicios ambientales e hidrológicos (Endara et al., 2013; Heredia-Bobadilla y Sunny, 2021; Luna-Gil, 2021; Queijeiro-Bolaños y Cano-Santana, 2016). Sin embargo, estos bosques se encuentran sometidos a constante presión antrópica debido a la cercanía con grandes complejos urbanos; esto ocasiona entre otras problemáticas, contaminación de suelo y cuerpos de agua, aumento de la tala ilegal, que reduce la variabilidad genética, además de que, como sugieren algunos estudios, facilita la dispersión de las semillas de muérdago (Endara y Herrera, 2017; Heredia Bobadilla et al., 2018). Las respuestas del bosque ante estos disturbios son cambios en la dinámica de las poblaciones de muérdagos enanos, por ejemplo, el incremento de los niveles de infestación (Endara et al., 2012; Queijeiro-Bolaños et al., 2013). Por otro lado, se conoce que los requerimientos ecológicos de Arceuthobium globosum Hawksworth et Wiens. y Arceuthobium vaginatum Willd. son similares, por lo que, pueden habitar en simpatría en la FVTM (Geils et al., 2002).
Se estima que en México las plantas parásitas se distribuyen en cerca de 1,800,000 ha de zonas forestales (Martínez-Salvador et al., 2015). De acuerdo con Ramírez y Porcayo (2009, 2010), la distribución de muérdagos enanos en el bosque de P. hartwegii es agregada y suele ser más abundante en hospederos jóvenes. En términos de infestación, Queijeiro-Bolaños y Cano-Santana (2016) reportan que A. vaginatum es más agresivo que A. globosum, aunque ambos provocan elevados niveles de infestación y presentan una distribución agregada. Queijeiro-Bolaños et al. (2013) reportan que factores fisiográficos como la altitud están relacionados positivamente con la distribución de A. vaginatum y A. globosum, mientras que los disturbios de origen antropogénico, como el desarrollo de basureros, actividades ganaderas e incendios, favorecen su presencia. La información sobre la distribución espacial de muérdagos en bosques de alta montaña es escasa debido a la complejidad de estos ecosistemas y las grandes superficies de distribución de estas plantas parásitas, por lo que el objetivo de este estudio fue conocer la distribución espacial de de A. vaginatum y A. globosum en los bosques de P. hartwegii del APFFNT y MT e identificar aquellos factores que estén correlacionados directamente con niveles de infestación por muérdagos; en este sentido se plantearon las siguientes preguntas de investigación: 1) ¿qué variables fisiográficas y bióticas influyen en la distribución espacial de los muérdagos enanos? 2) ¿la densidad del bosque influye en el aumento de los niveles de infestación? y 3) ¿la presencia de disturbios de origen antrópico favorece el aumento de los niveles de infestación?
Materiales y métodos
El estudio se realizó en los bosques de P. hartwegii del APFFNT y el MT, esta especie es una de las más representativas en las montañas de la FVTM por las condiciones en las que se desarrolla (Conanp, 2016), ya que tolera bajas temperaturas (hasta -30 °C) y determina el límite superior de la vegetación arbórea (Alfaro-Ramírez et al., 2017; Challenger y Soberón, 2008), siendo el bosque que se establece a mayor altitud en el mundo, en un intervalo de 3,000 a 4,200 m snm (Perry, 1991). En ambas montañas, hacia el límite inferior del bosque de P. hartwegi, se encuentran bosques mixtos de otras especies de pinos (P. montezumae Lamb., P. ayacahuite Ehrenb. ex Schltdl.), encinos (Quercus rugosa Née, Q. laurina Bondpland) y ailes (Alnus acuminata Kunth y A. jorullensis Kunth) (Astudillo-Sanchez et al., 2019; Bolaños-González et al., 2017). El presente estudio se enfocó en los bosques de alta montaña, es decir, aquellos presentes a partir de 3,500 m snm y hasta el límite superior de la vegetación (Endara et al., 2013) y en los que predomina Pinus hartwegii y, ocasionalmente, se pueden encontrar otras especies de coníferas como Abies religiosa Kunth Schltdl. et Cham., Pinus montezumae, P. ayacahuite y P. pseudostrobus Lindl.
El muestreo se realizó durante el periodo de enero 2016 a diciembre 2021. Se establecieron 1,622 sitios de muestreo (SM) en el APFFNT y 634 en el MT (fig. 1). Se instalaron SM circulares de 0.1 ha (35.6 m de diámetro) utilizando las curvas de nivel (espaciadas a cada 100 m altitudinales) como líneas de acceso al bosque; sobre cada curva se establecieron los SM con una equidistancia de 200 m (APFFNT) y 300 m (MT). La intensidad mínima de muestreo calculada, según Dauber (1995), para el Nevado de Toluca fue de 0.44 y la realizada fue de 1.25, para el monte Tláloc fueron 0.89 y 1.36, respectivamente. Es importante resaltar la importancia del muestreo por curvas de nivel, ya que un incremento en altitud de 100 m implica una reducción de entre 0.6 y 1.2 grados centígrados, es decir, un cambio radical en las condiciones ambientales (Mayer y Ott, 1991).
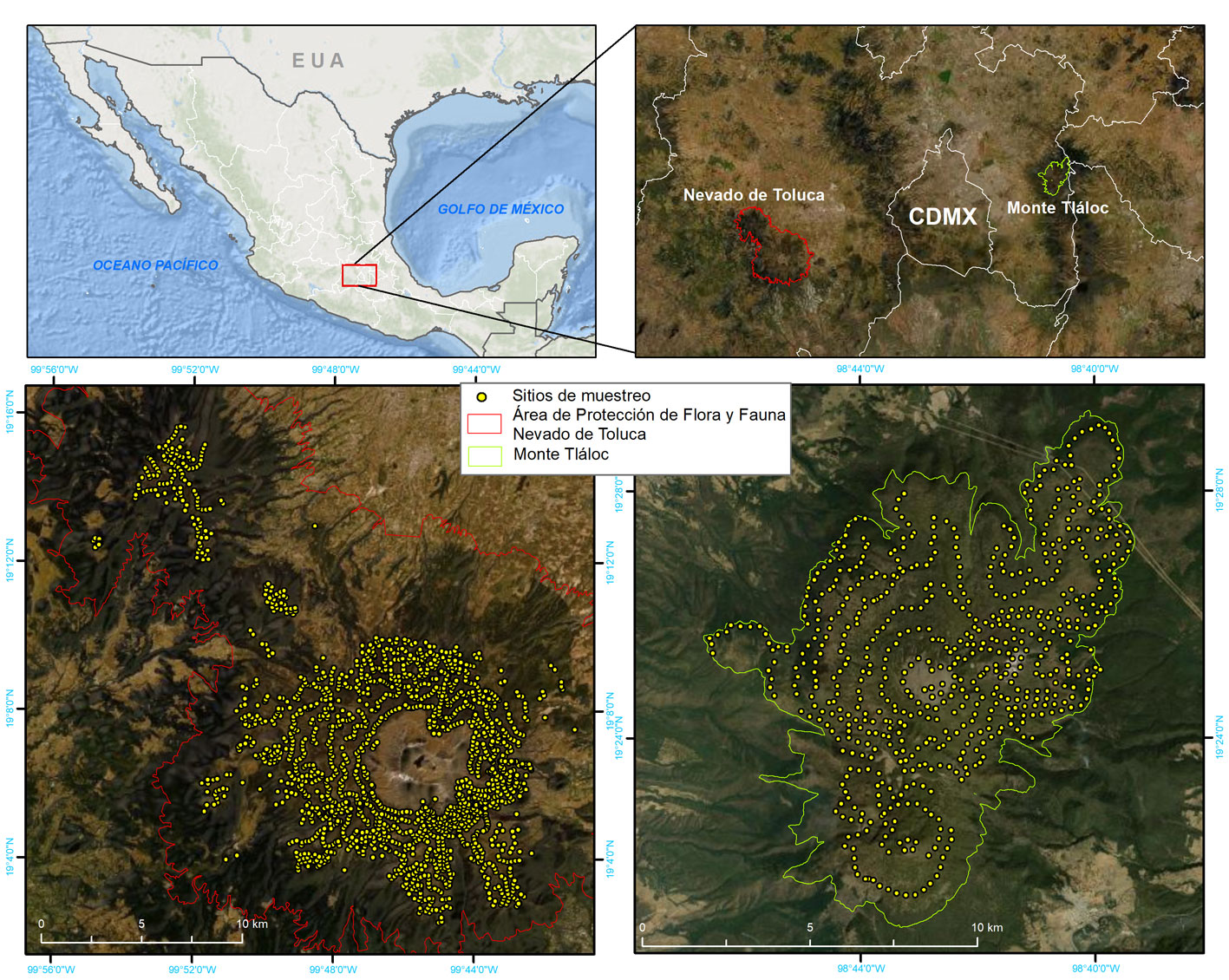
En cada SM se registraron 4 variables fisiográficas, 5 variables correspondientes a características de la vegetación y características que pudieran estar relacionadas con disturbios forestales; en este caso se registró la presencia de 7 tipos de actividades antrópicas (tabla 1), así como la categoría o edad de las especie arbóreas presentes en el SM, 6 variables dasonómicas y densidad del bosque (Endara et al., 2013; Rojas-García et al., 2019), el nivel de infestación de los árboles con muérdago, según el método de Hawksworth (1983) y también la presencia y nivel de infestación por descortezadores, según Billings y Espino (2005). Esta última requirió la recolecta de individuos adultos, mismos que fueron identificados en el laboratorio de sanidad forestal de la Protectora de Bosques del Estado de México (Probosque).
Se contabilizaron las plántulas, brinzales y latizales; a las categorías juveniles y fustales se les tomaron medidas dasométricas; con éstas últimas categorías se determinó la densidad del bosque (Endara et al., 2013; Rojas et al., 2019) (tabla 2).
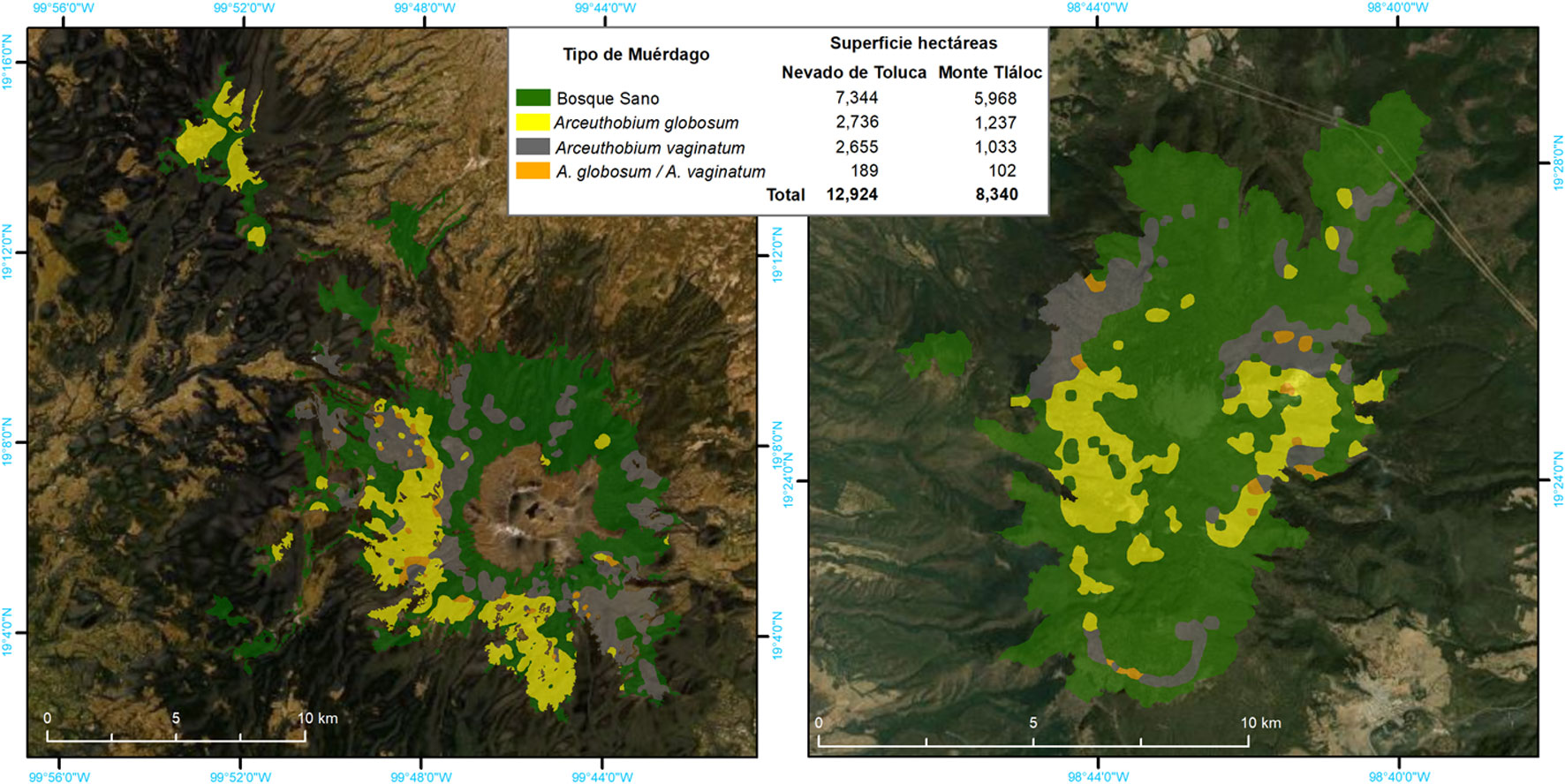
Con la información de sitios e individuos se generó una base de datos para visualizarla en el software ArcGIS ver.10.8, en el cual se aplicó la metodología de interpolación de datos del vecino natural (Sibson, 1982), que consiste en la generación de polígonos con base en información de los SM, utilizando el valor del vecino más cercano para formar superficies continuas y así representarlas por condición, sin importar el tipo distribución (Childs, 2004; Declercq, 1996; Lam, 1983; Pirotti y Tarolli, 2010). La generación del mapa de distribución se realizó con base en la presencia-ausencia de cada tipo de muérdago y los diferentes niveles de infestación por sitio.
Se realizaron gráficos descriptivos para analizar si existe una tendencia de establecimiento de muérdago por altitud (tabla 1) y con una prueba de regresión simple se analizó si la presencia de muérdago está relacionada con factores abióticos (presencia de caminos, extracción, incendios, pastoreo y/o tinas ciegas). Con una prueba de Anova y diferencias mínimas significativas de Fisher (LSD), se analizó si el nivel de infestación difiere por especie de muérdago, se espera que el mayor nivel de infestación sea para A. vaginatum, por ser la especie que tiene mayor distribución y también más hospederos (Queijeiro-Bolaños et al., 2013); para esta prueba se utilizaron los niveles de infestación por muérdago bajo 3 condiciones: muérdago amarillo (A. globosum), muérdago negro (A. vaginatum) y ambos (cuando en un mismo árbol se presentaron las 2 especies de muérdago) (Hawksworth, 1983). Se realizó una prueba de regresión simple para determinar si la densidad del bosque influye en el nivel de infestación por especie de muérdago (tabla 2). Para realizar esta prueba se utilizaron los valores de porcentaje de cobertura y nivel de infestación por cada una; estas pruebas se realizaron con el programa Statgraphics Centurion XVI v.16.1.03 (StatPoint Technologies Inc., 2009).
Se estableció qué tipo de variables bióticas —cobertura de copa, porcentaje herbáceo y arbustivo, infestación por descortezador y número de fustale— y abióticas —pendiente, altitud— tuvieron un mayor peso sobre el nivel de infestación por muérdago, con un análisis de componentes principales; se eligió la opción de extraer los componentes principales con base en la matriz de correlaciones, debido a las diferentes unidades en que se midieron las variables consideradas para el análisis. Las comparaciones se hicieron entre grupos de variables (tabla 1). Se realizaron gráficos de distribución diamétrica por categoría de individuos (fustales y juveniles) y, si éstos eran sanos o enfermos, para establecer qué categoría es más atacada por muérdago según el diámetro normal (DN, 1.3 metros de la base del fuste) (tabla 2). Para estos análisis se utilizó el programa SPSS Statistics 26 (IBM Corp., 2019).
Tabla 1
Variables de sitio; información colectada en cada Sitio de Muestreo (SM). Se indica entre paréntesis la forma en que se midió cada variable.
| Fisiográficas | Vegetación | Actividades antrópicas
(presencia/ ausencia) |
| Coordenadas (UTM)
Ubicación (ladera, pie, cima) Pendiente y azimut (grados) Altitud (m snm) |
Estructura (juvenil, senil, semillero, pastizal)
Cobertura de copa (%) Cobertura de suelo (%) Estrato herbáceo, arbustivo, afloramiento rocoso, suelo desnudo (%) Número de especies (arbóreas, arbustivas, herbáceas y rasantes) |
Camino
Cultivo Extracción Incendios Pastoreo Reforestación Tina ciega |
Tabla 2. Datos obtenidos para Pinus hartwegii a partir de la cuantificación de plántulas, brinzales, latizales, juveniles y fustales.
| Categoría
(intervalo de altura-diámetro) |
Variables dasonómicas |
| Plántula (< 30 cm altura)
Brinzal (≥ 30 cm < 1.5 m altura) Latizal (≥ 1.5 m altura < 2.5 cm DN) Juvenil (≥ 2.5 cm DN < 7.5 cm DN) Fustal (≥ 7.5 cm DN) Porcentaje de cobertura (el número de fustales/ha determinan la densidad del arbolado) |
Diámetro a la altura del pecho (DN)
Altura total Nivel de infestación para muérdagos enanos (Hawksworth, 1983) Nivel de infestación por descortezador (Billings y Espino, 2005) Daños por rayos Muertos |
Resultados
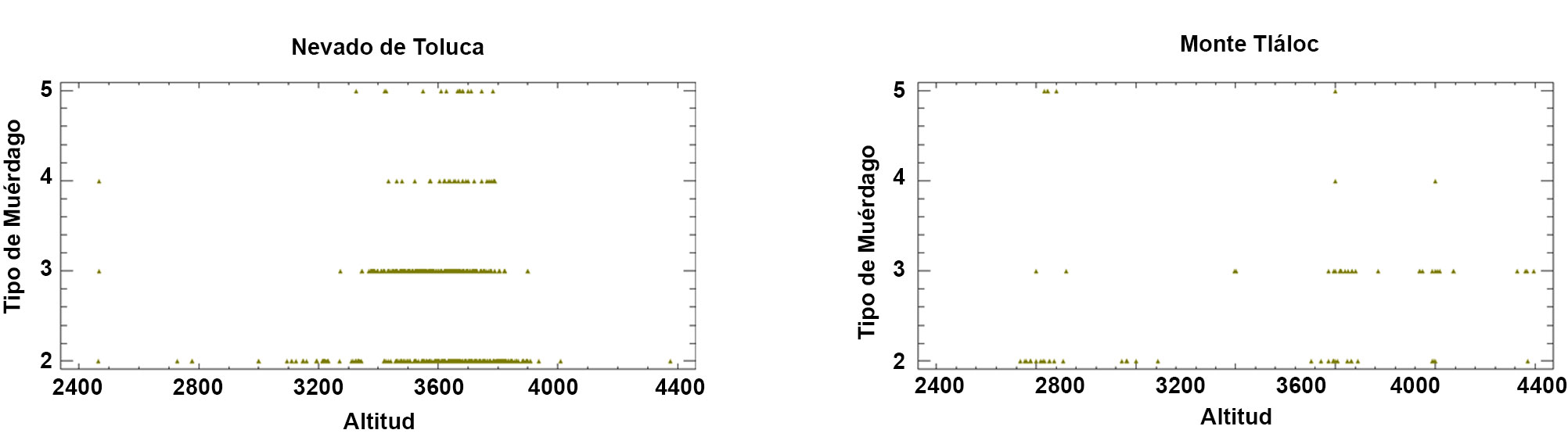
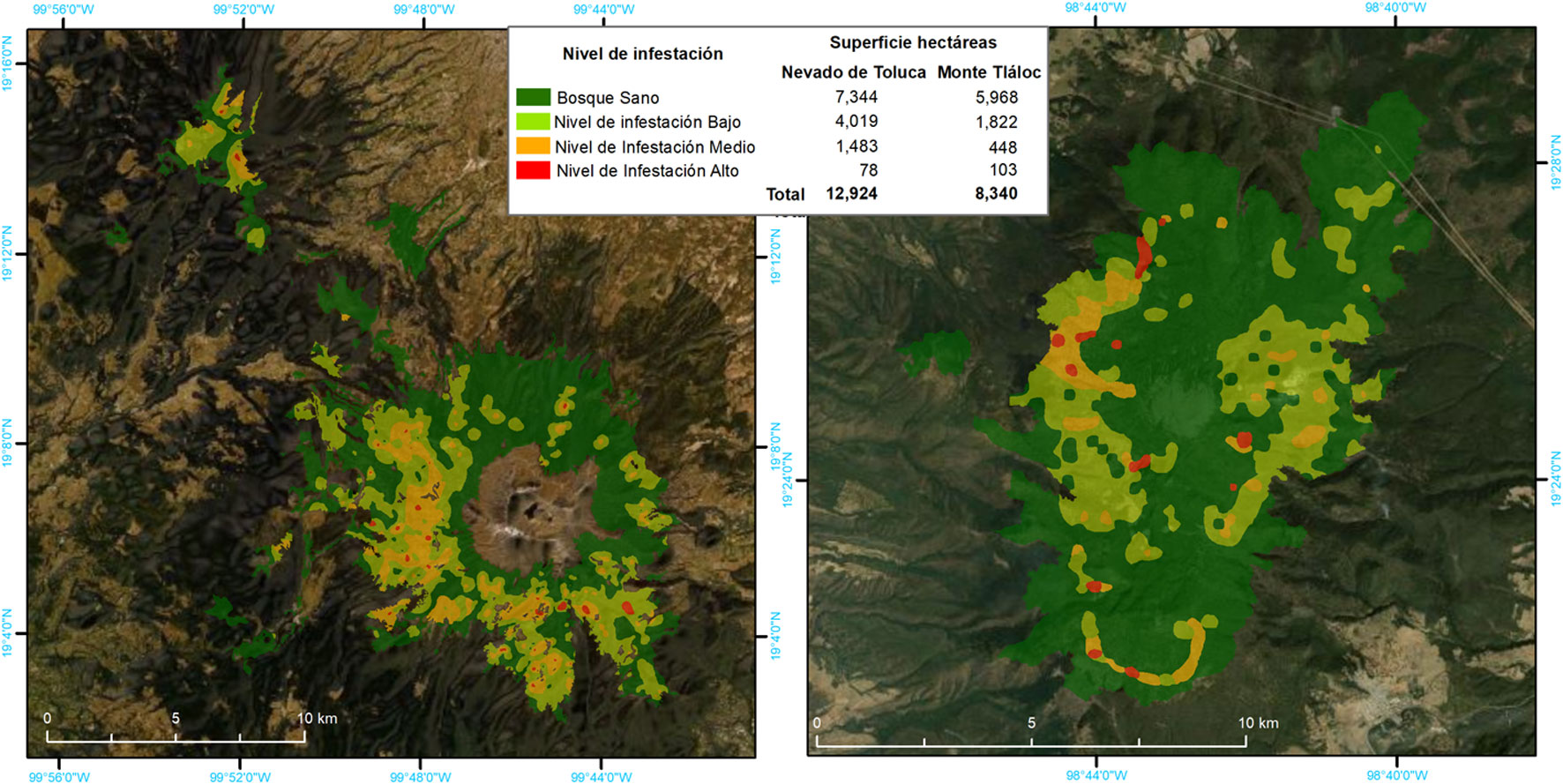
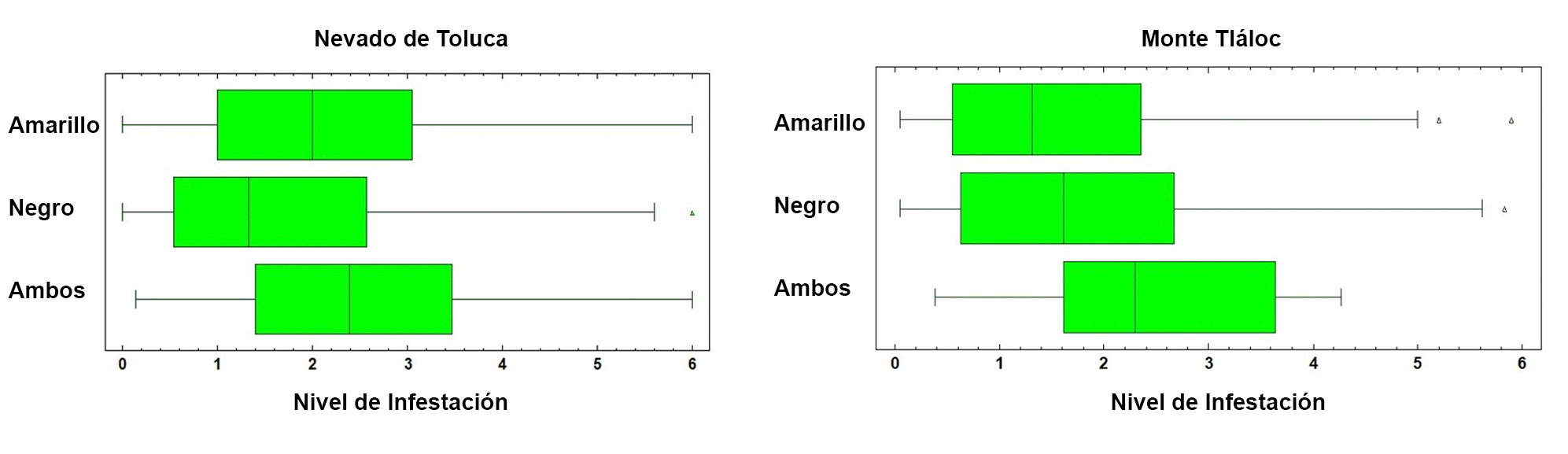
La distribución espacial de Arceuthobium globosum y A. vaginatum en el bosque de Pinus hartwegii de ambas montañas obedece a ciertos patrones: primero, 70% de los SM con muérdago se encuentran entre 3,600 y 3,800 m snm. (figs. 2, 3); segundo, las mayores infestaciones se concentran en las laderas oeste. La prueba de regresión para determinar si las actividades antrópicas tienen algún efecto sobre la presencia de muérdago no fue estadísticamente significativa para el APFFNT ni para MT (APFFNT: r2 = 0.07%, p = 0.3; MT: r = 0.0%, p = 0.9). En el APFFNT, 43% de la superficie (5,576 ha) presenta algún nivel de infestación de muérdagos enanos, mientras que en MT asciende a 28% (2,373 ha) (fig. 4). La prueba Anova y la prueba de diferencias mínimas significativas de Fisher (LSD) mostraron diferencias significativas para el nivel de infestación, indicando mayor afectación para infecciones simultáneas (fig. 5).
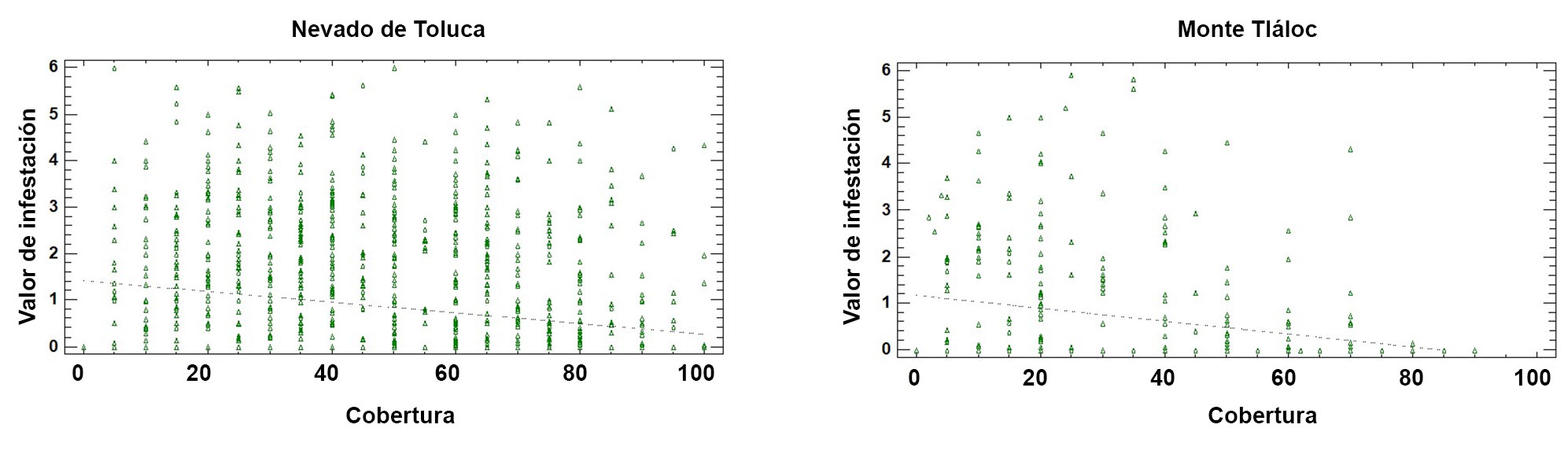
La prueba de regresión entre el valor de infestación del muérdago enano y el porcentaje de cobertura de copa para cada sitio de estudio (APFFNT: r2 = 5.2%, p = 0.000; MT: r = 5.8%, p = 0.000), mostró que para MT los bosques con mayor cobertura forestal presentan menores niveles de infestación (fig. 6).
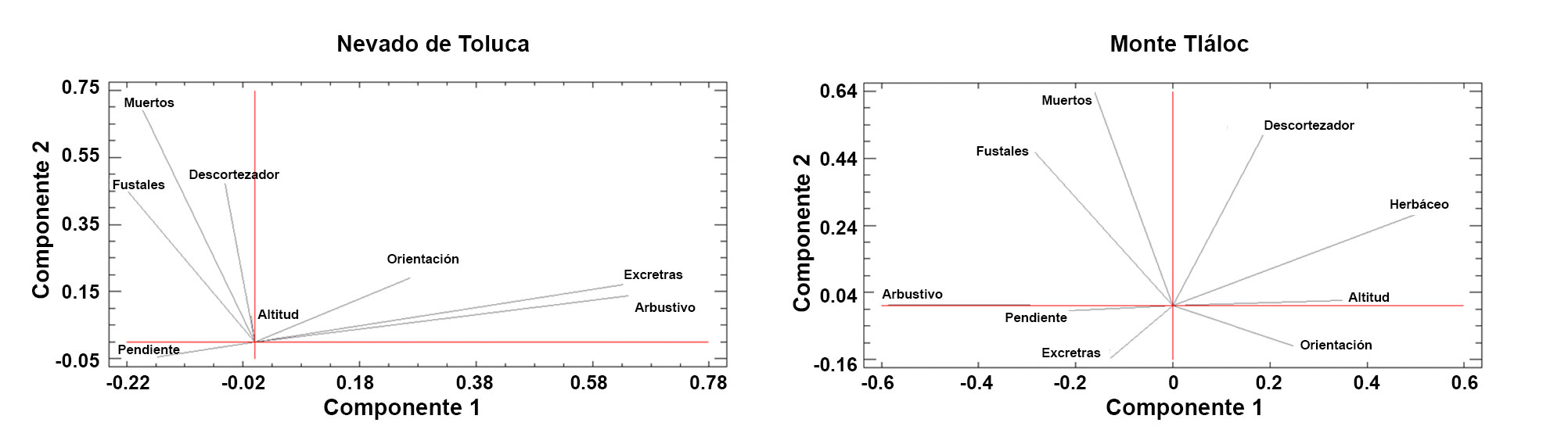
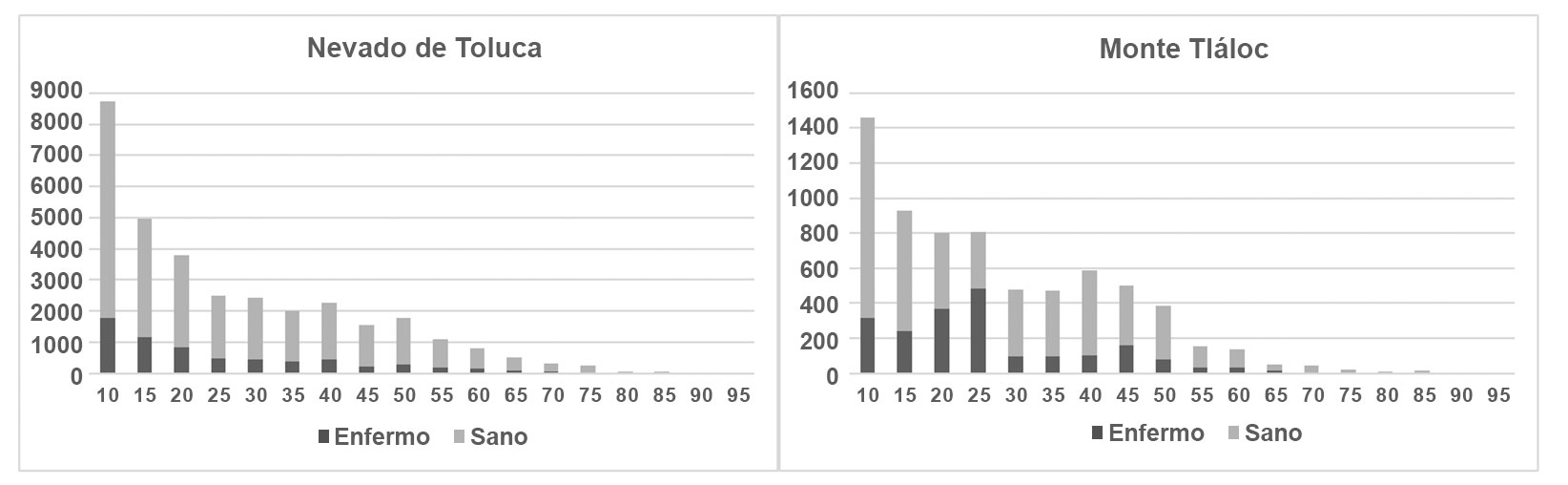
El análisis de componentes principales demostró que las variables bióticas tienen una mayor relación con la presencia de muérdago enano en los bosques de P. hartwegii; por otro lado, las actividades antrópicas resultaron no tener incidencia directa con la presencia de muérdago. En el APFFNT, 4 factores explicaron 69% de la variabilidad de los datos, las variables que más contribuyeron al modelo fueron la altitud, orientación, presencia de descortezador y fustales; cada una de estas variables contribuyó con 25.1%, 16.6%, 14.5% y 12.7% de varianza, respectivamente; en el MT, 4 factores explicaron 58% de la variabilidad del modelo, la altitud, el estrato herbáceo, la presencia de descortezador y los fustales, cada uno contribuyó con 18.6%, 14.5%, 13% y 12% de varianza al modelo, respectivamente. La presencia de descortezador y la altitud tuvieron peso sobre la variable muérdago en las 2 áreas de estudio (fig. 7). Si bien el muérdago enano parasita árboles de todas las categorías diamétricas (fig.8), su abundancia es mayor entre los 10 y 25 cm de DN. Los porcentajes de individuos infestados son de 20% para el APFFNT y de 18% para MT, pero en el caso del renuevo (plántulas, brinzales, latizales y juveniles), dicho porcentaje apenas alcanza 4% en ambas montañas, niveles dentro de los límites naturales de parasitismo (Mathiasen, 1998).
Discusión
De las 2 especies de muérdago identificadas Arceuthobium vaginatum y A. globosum, la primera se considera la más agresiva dado que afecta alrededor de 13 hospederos entre ellos P. hartwegii, además de tener mayor presencia de brotes en el fuste de los árboles (Mathiasen et al., 2004; Queijeiro-Bolaños et al., 2011).
Los resultados de las pruebas estadísticas muestran un patrón de correlación entre la altitud y la presencia de muérdago, no así para el nivel de infestación, lo que concuerda con Queijeiro-Bolaños et al. (2013), quienes mencionan que a mayor altitud existe menor presencia de la planta parásita. El trabajo de campo corrobora que la presencia de muérdago sobre P. hartwegii se limita a 4,000 m snm, altitud en la cual el bosque de P. hartwegii parece encontrar el hábitat ideal (Endara et al., 2013). Otro factor que puede influir en la presencia de muérdago es la densidad del bosque, a mayores altitudes la densidad (árboles por hectárea) es baja, lo cual dificulta la dispersión de semillas de Arceuthobium (Hawksworth, 1983), al respecto, se ha reportado que las semillas de estos organismos se desarrollan mejor bajo un dosel forestal denso, estos resultados son similares a lo que reporta Klutsch et al. (2014), quienes encontraron una relación directa entre la densidad forestal y la presencia de muérdago enano y descortezadores (Brunn et al., 2006; Hawksworth, 1983; Queijeiro-Bolaños et al., 2013; Shaw y Weiss, 2000). Esto, a su vez, sugiere una relación directa con la orientación, argumentando que las horas luz, el estrés hídrico y el incremento de la temperatura en las zonas de montaña, pueden afectar a los hospederos y favorecer la presencia y establecimiento del muérdago. A diferencia del APFFNT, en el MT, los bosques de P. hartwegii presentan una menor densidad y los árboles están más separados entre sí, lo cual reduce la capacidad de dispersión de la planta parásita y se refleja en un porcentaje menor de superficies infestadas.
Los mayores niveles de infestación se registraron en individuos con ataques simultáneos de ambas especies de muérdago. Al respecto, se sabe que existe especificidad por ciertos hospederos dentro del género Arceuthobium y que, aunque A. globosum y A. vaginatum suelen encontrarse atacando un mismo hospedero, son factores estocásticos los que determinan la abundancia de éstos, tales como incendios o tala (González-Elizondo et al., 2019; Mathiasen, 2019; Mathiasen et al., 2004; Queijeiro-Bolaños et al., 2011). Por otra parte, es muy probable que estos ataques ocurran en los árboles con mejor constitución fisiológica, y por ende, altos niveles de polimorfismo genético, dados los altos requerimientos de agua y nutrientes por parte de los muérdagos (Bickford et al., 2005; Heredia-Bobadilla et al., 2018).
Respecto a los factores que pueden determinar la presencia del muérdago enano en el bosque, el análisis de componentes principales sugiere que las infecciones por muérdagos predisponen el ataque por descortezadores (Ferrenberg, 2020; Mathiasen, 2019), lo que coincide con Cibrián-Tovar et al. (1995), quienes reportan la pérdida de vigor y reducción en el crecimiento de árboles parasitados, lo cual facilita la infestación por descortezadores. La interacción de descortezadores y muérdago genera más probabilidades de mortalidad (Klutsch et al., 2014), lo cual provoca que el dosel forestal se reduzca facilitando la dispersión de las semillas del muérdago.
La infestación por muérdagos enanos en individuos de menos de 1 m de altura es muy rara, debido que los árboles de estas tallas no son un blanco fácil para las semillas del parásito, además de que agentes externos, como las lluvias, remueven las semillas de estos hospederos (Mathiasen, 1998). Tal como los resultados de este estudio lo corroboran en ambas montañas (4% de infestación), los individuos adultos y seniles enfermos que ocupan el dosel dominante del bosque favorecen la dispersión de semillas, infestando estratos inferiores y la regeneración natural.
Los ataques en la regeneración ocurren cuando las infestaciones en árboles maduros son muy fuertes (Skay et al., 2021). Casos similares se han reportado en Picea engelmani, P. pungens y Pseudotsuga menziesii, afectados por A. microcarpum y A. douglasii, respectivamente (Mathiasen, 1986).
La presencia de muérdagos enanos suele estar relacionada con disturbios de tipo antrópico, tales como el cambio de uso del suelo y los asentamientos humanos (Challenger, 1998; Sánchez et al., 2003). La quema y el pastoreo son fenómenos frecuentes en el APFFNT y en MT, que inciden en el deterioro del bosque de pino (Salinas et al., 2010; Villers et al., 1998), en este caso, no se encontró una correlación estadística significativa de las variables antrópicas con la presencia de muérdago.
Los disturbios de tipo antrópico pueden contribuir en la pérdida de vigor de los hospederos (Ferrenberg, 2020; Fonseca-González et al., 2014; Queijeiro-Bolaños et al., 2013; Rodríguez-Trejo, 2002), predisponiendo al arbolado al ataque de parásitos; aunado a ésto, algunos estudios sugieren que el cambio climático global puede estar influyendo directamente en el desarrollo, ciclos de reproducción, fenología y distribución espacial de los patógenos (Harrington et al., 2001; Sáenz-Romero et al., 2020).
El 37% de los bosques de Pinus hartwegii del APFFNT y el MT están infestados por Arceuthobium globosum y A. vaginatum, distribuidos entre los 3,600 y 3,800 m snm, afectando a todas las categorías de DN, aunque con mayor intensidad a individuos entre 10 a 25 DN. Las mayores infestaciones se concentran en la ladera oeste de ambos edificios volcánicos, por lo que, el complemento de esta evidencia con datos bioclimáticos, así como afectaciones por estrés hídrico y horas luz sobre las diferentes laderas pueden corroborar este supuesto. La presencia de descortezadores del género Dendroctonus ssp. es una variable directamente relacionada con los niveles de infestación de muérdago. Esto implica que, los árboles infestados por muérdagos son más vulnerables al ataque de descortezadores.
Agradecimientos
Agradecemos el apoyo financiero del proyecto Conafor-Conacyt A-S-130105 denominado “Identificación, evaluación y manejo integrado de plantas parásitas en cuatro regiones de México: Noroeste (Durango), Centro-Occidente (Michoacán y Jalisco), Oriente (Puebla y Veracruz), Centro-Sur (Estado de México y Tlaxcala)”. A las localidades de Raíces, Santa María del Monte, Calimaya, Tlacotepec, Agua Blanca, San Francisco Putla, Palo Seco, La Comunidad, Amanalco y El Capulín en el Nevado de Toluca; a las comunidades San Pablo Ixayoc, San Miguel Tlaixpan, Tequexquináhuac, Santa Catarina, y San Juan Cuauhtémoc en Monte Tláloc.
Referencias
Alfaro-Ramírez, F. U., Arredondo-Moreno, J. T., Pérez-Suárez, M. y Endara-Agramont, A. R. (2017). Pinus hartwegii Lindl. treeline ecotone: structure and altitudinal limits at Nevado de Toluca, Mexico. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 23, 261–273. https://doi.org/10.5154/r.rchscfa.2016.10.055
Astudillo-Sánchez, C. C., Fowler, M. S., Villanueva-Díaz, J., Endara-Agramont, A. R. y Soria-Díaz, L. (2019). Recruitment and facilitation in Pinus hartwegii, a Mexican alpine treeline ecotone, with potential responses to climate warming. Trees, 33, 1087–1100. https://doi.org/10.1007/s00468-019-01844-3
Bickford, C. P., Kolb, T. E. y Geils, B. W. (2005). Host physiological condition regulates parasitic plant performance: Arceuthobium vaginatum subsp. cryptopodum on Pinus ponderosa. Ecophysiology, 146, 179–189. https://doi.org/10.1007/s00442-005-0215-0
Billings, R. F. y Espino, V. (2005). El gorgojo descortezador del pino (Dendroctonus frontalis) en Centroamérica: cómo reconocer, prevenir y controlar plagas. Texas, USA: Texas Forest Service Publication.
Bolaños-González, Y., Bolaños-González, M. A., Paz, P. F. y Ponce-Pulido, J. I. (2017). Estimación de carbono almacenado en bosques de oyamel y ciprés en Texcoco, Estado de México. Terra Latinoamericana, 35, 73–86. https://doi.org/10.28940/terra.v35i1.243
Brunn, H. H., Moen, J., Virtanen, R., Grytnes, J. A., Oksanen, L. y Angerbjörn, A. (2006). Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities. Journal of Vegetation Science, 17, 37–46. https://doi.org/10.1111/j.1654-1103.2006.tb02421.x
Challenger, A. (1998). Utilización y conservación de los ecosistemas terrestres de México. Pasado, presente y futuro. México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/Instituto de Biología, Universidad Nacional Autónoma de México/Agrupación Sierra Madre A.C.
Challenger, A. y Soberón, J. (2008). Los ecosistemas terrestres. En Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio) (Eds.), Capital natural de México, Vol. I. Conocimiento actual de la biodiversidad (pp.87–108). México D.F.: Conabio.
Childs, C. (2004). Interpolating surfaces in ArcGIS spatial analyst. ArcUser, July-September, 3235, 32–35.
Cibrián-Tovar, D., Méndez-Montiel, J. T., Campos-Bolaños, R., Tates III, H. O. y Flores-Lara, J. E. (1995). Insectos forestales de México. México D.F.: Universidad Autónoma Chapingo.
Comisión Nacional de Áreas Naturales Protegidas (2016). Programa de manejo del Área de Protección de Flora y Fauna Nevado de Toluca. México D.F.: Secretaría del Medio Ambiente y Recursos Naturales.
Dauber, E. (1995). Guía práctica y teórica para el diseño de un inventario forestal de reconocimiento. Santa Cruz, Bolivia: BOLFOR.
Declercq, F. A. N. (1996). Interpolation methods for scattered sample data: accuracy, spatial patterns, processing time. Cartography and Geographic Information Systems, 23, 128–144. https://doi.org/10.1559/152304096782438882
Endara, A. A. R., Calderón-Contreras, R., Nava-Bernal, G. y Franco, S. (2013). Analysis of fragmentation processes in high-mountain forests of the Centre of Mexico. American Journal of Plant Sciences, 4, 697–704. http://dx.doi.org/10.4236/ajps.2013.43A088
Endara, A. A. R., Franco, S., Bernal, G. N., Hernández, J. I. V. y Fredericksen, T. S. (2012). Effect of human disturbance on the structure and regeneration of forests in the Nevado de Toluca National Park, Mexico. Journal of Forestry Research, 23, 39–44. https://doi.org/10.1007/s11676-012-0226-8
Endara, A. R. R. y Herrera, T. F. (2017). Deterioro y conservación de los bosques del Nevado de Toluca y el rol de los actores locales. Ciencia Ergo Sum, 23, 247–254.
Ferrenberg, S. (2020). Dwarf mistletoe infection interacts with tree growth rate to produce opposing direct and indirect effects on resin duct defenses in lodgepole pine. Forests, 11, 222. https://doi.org/10.3390/f11020222
Fonseca-González, J., Santos-Posadas, H. M., Rodríguez-Ortega, A. y Rodríguez-Laguna, R. (2014). Efecto del daño por fuego y descortezadores sobre la mortalidad de Pinus patula Schl et Cham en Hidalgo, México. Agrociencia, 48, 103–113.
González-Elizondo, M., Flores-Villegas, M. Y., Álvarez-Zagoya, R., González-Elizondo, M. S., Márquez-Linares, M. A., Quiñones-Barraza, S. et al. (2019). Effects of Mexican dwarf mistletoe (Arceuthobium vaginatum subesp. vaginatum) on the growth of Pinus cooperi in Durango, México, a case study. Forest Pathology, 49, e12473. https://doi.org/10.1111/efp.12473
Geils, B. W., Cibrian-Tovar, J. y Moody, B. (2002). Mistletoes of North American conifers. Gen. Tech. Rep. RMRS-GTR-98. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Harrington, R., Fleming, R. A. y Woiwod, I. P. (2001). Climate change impacts on insect management and conservation in temperate regions: Can they be predicted? Agricultural and Forest Entomology, 3, 233–240. https://doi.org/10.1046/j.1461-9555.2001.00120.x
Hawksworth, F. G. (1983). Mistletoes as forest parasites. En M. Calder y P. Bernhardt (Eds.), The biology of mistletoes (pp. 317–333). Nueva York: Academic Press.
Heredia-Bobadilla, R. L., Gutiérrez-González, G., Arzate-Fernández, A. M. y Franco-Maass, S. (2018). Genetic variability of mountain pine (Pinus hartwegii Lindl.) in the Protection of Flora and Fauna Area Nevado de Toluca. En A. E. Mohamed (Ed.), Genetic diversity in plant species characterization and conservation (pp. 71–85). Londres: InTech Open.
Heredia-Bobadilla, R. L. y Sunny, A. (2021). Análisis de la categoría de riesgo de los ajolotes de arroyos de alta montaña (Caudata: Ambystoma). Acta Zoológica Mexicana (N.S.), 37, 1–19. https://doi.org/10.21829/azm.2021.3712315
IBM Corp. (2019). IBM SPSS Statistics for Windows, Version 26.0. Armonk, NY: IBM Corp.
Iwaniszewski, S. (1994). Archaeology and archaeoastronomy of Mount Tláloc, México: a reconsideration. Society for American Archaeology, 5, 158–176. https://doi.org/10.2307/971561
Klutsch, J. G., Beam, R. D., Jacobi, W. R. y Negrón, J. F. (2014). Bark beetles and dwarf mistletoe interact to alter downed woody material, canopy structure, and stand characteristics in northern Colorado ponderosa pine. Forest Ecology and Management, 315, 63–71. https://doi.org/10.1016/j.foreco.2013.12.024
Lam, N. S. N. (1983). Spatial interpolation methods: a review. The American Cartographer, 10, 129–150. https://doi.org/10.1559/152304083783914958
Luna-Gil, A. A. (2021). Caracterización del hábitat de Romerolagus diazi en el bosque de alta montaña del monte Tláloc, Sierra Nevada (Tesis de maestría). ICAR, Universidad Autónoma del Estado de México. México.
Martínez-Salvador, M., Madrigal-Huendo, S., Vázquez-Collazo, I., Velasco-Bautista, E., Morales-Nieto, C. R. y Villareal-Guerrero, F. (2015). Effect of Arceuthobium vaginatum (Willd.) Presl. subsp. vaginatum in Pinus hartwegii Lindl. in Colima State. Revista Mexicana de Ciencias Forestales, 6, 44–55. https://doi.org/10.29298/rmcf.v6i29.215
Mathiasen, R. L. (1986). Infection of young Douglas-firs and spruces by dwarf mistletoes in the southwest. The Great Basin Naturalist, 46, 528–534.
Mathiasen, R. L. (1998). Infection of young western larch by larch dwarf mistletoe in northern Idaho and western Montana. Western Journal of Applied Forestry, 13, 41–46. https://doi.org/10.1093/wjaf/13.2.41
Mathiasen, R. L. (2019) Susceptibility of red fir and white fir to fir dwarf mistletoe (Arceuthobium abietiunum) in California. Forest Pathology, 2019, e12516. https://doi.org/10.1111/efp.12516
Mathiasen, R. L., Flores, H. M. y Cadio, L. (2004). First report of Arceuthobium vaginatum subsp. vaginatum on Pinus pseudostrobus. Plant Disease, 8, 1046–1046. https://doi.org/10.1094/PDIS.2004.88.9.1046B
Mayer, H. y Ott, E. (1991). Gebirgswaldabau-Schutzwaldpflege: ein waldbauulicher Beitrag zur Landschaftsokologie und zum Umweltschutz (Silviculture in Mountain Forest-Management of protection forest: a silvicultural contribution to landscape ecology and environmental protection). Stuttgart: Gustav Fischer.
Montero-García, I. A. (2004). Atlas arqueológico de la alta montaña mexicana. México D.F.: Semarnat.
Perry, J. P. (1991). The pines of Mexico and Central America. Portland, Oregon: Timber Press.
Pirotti, F. y Tarolli, P. (2010). Suitability of LiDAR point density and derived landform curvature maps for channel network extraction. Hydrological Processes, 24, 1187–1197. https://doi.org/10.1002/hyp.7582
Queijeiro-Bolaños, M. E. y Cano-Santana, Z. (2016). Growth of Hartweg’s pine (Pinus hartwegii) parasitized by two dwarf mistletoe species (Arceuthobium spp.) Botanical Sciences, 94, 51–62. https://doi.org/10.17129/botsci.218
Queijeiro-Bolaños, M. E., Cano-Santana, Z. y Castellanos-Vargas, I. (2011). Distribución diferencial de 2 especies de muérdago enano sobre Pinus hartwegii en el área natural protegida “Zoquiapan y Anexas”, Estado de México. Acta Botanica Mexicana, 96, 49–57. https://doi.org/10.21829/abm96.2011.258
Queijeiro-Bolaños, M. E., Cano-Santana, Z. y Castellanos-Vargas, I. (2013). Does disturbance determine the prevalence of dwarf mistletoe (Arceuthobium, Santalales: Viscaceae) in Central Mexico?. Revista Chilena de Historia Natural, 86, 181–190. http://dx.doi.org/10.4067/S0716-078X2013000200007
Ramírez, J. F. y Porcayo, E. (2009). Estudio de la distribución espacial del muérdago enano (Arceuthobium sp.) en el Nevado de Toluca, México utilizando el método del SADIE. Madera y Bosques, 15, 93–112. https://doi.org/10.21829/myb.2009.1521193
Ramírez, J. F. y Porcayo, E. (2010). Estudio comparativo de la distribución espacial del muérdago enano (Arceuthobium sp.) en la ladera norte del Parque Nacional Nevado de Toluca, México. Bosque (Valdivia), 31, 28–38. https://doi.org/10.4067/S0717-92002010000100004
Rodríguez-Trejo, D. A. (2002). Ecología del fuego en el ecosistema de Pinus hartwegii Lindl. Revista Chapingo Serie Ciencias Forestales y Ambiente, 7, 145–151.
Rojas-García, F., Fredericksen, T. S., Vázquez-Lozada, S. y Endara-Agramont, A. R. (2019). Impact of timber harvesting on carbon storage in montane forests of central Mexico. New Forests, 50, 1043–1061. https://doi.org/10.1007/s11056-019-09714-z
Sáenz-Romero, C., Mendoza-Maya, E., Gómez-Pineda, E., Blanco-García, A., Endara-Agramont, A. R., Lindig-Cisneros, R. et al. (2020). Recent evidence of Mexican temperate forest decline and the need for ex situ conservation, assisted migration, and translocation of species ensembles as adaptive management to face projected climatic change impacts in a megadiverse country. Canadian Journal of Forest Research, 50, 843–854. https://doi.org/10.1139/cjfr-2019-0329
Salinas-Moreno, Y., Ager, A., Vargas, C. F., Hayes, J. L. y Zúñiga, G. (2010). Determining the vulnerability of Mexican pine forests to bark beetles of the genus Dendroctonus Erichson (Coleoptera: Curculionidae: Scolytinae). Forest Ecology and Management, 260, 52–61. https://doi.org/10.1016/j.foreco.2010.03.029
Sánchez, O., Vega, E., Peters, E. y Monroy, V. O. (2003). Conservación de los ecosistemas templados de montaña en México. México D.F.: Instituto Nacional de Ecología.
Sánchez-González, A. (2008). Una visión actual de la diversidad y distribución de los pinos de México. Madera y Bosques, 14, 107–120. https://doi.org/10.21829/myb.2008.1411222
Shaw, D. C. y Weiss, S. B. (2000). Canopy light and distribution of hemlock dwarf mistletoe (Arceuthobium tsugense [Rosendahl] G.N. Jones subesp. tsugense) aerial shootsin an old-growth Douglass fir/Western hemlock forest. Northwest Science, 74, 306–315.
Sibson, R. (1982). A brief description of natural neighbor interpolation. En John Wiley y Sons (Eds.), Interpolating multivariate data (pp. 21–36). Nueva York.
Skay, R., Windmuller-Campione, M. A., Russel, M. B. y Reuling, M. F. (2021). Influence of eastern spruce dwarf mistletoe on stand structure and composition in northern Minnesota. Forest Ecology and Management, 481, 118712. https://doi.org/10.1016/j.foreco.2020.118712
StatPoint Technologies. INC. version, 16, 17. (2009). Statgraphics Centurion, X. V. I.
Varo-Rodríguez, R., Avila-Akerberg, V. y Gheno, J. (2016). Caracterización de la infestación por muérdago enano (Arceuthobium globosum subesp. grandicaule) en el bosque de Pinus hartwegii de la cara sur del Pico de Orizaba. Teoría y Praxis, 19, 11–31. https://doi.org/10.14483/2256201x.17163
Villers, R. L., García del Valle, L. y López-Blanco, J. (1998). Evaluación de los bosques templados en México: una aplicación en el parque nacional Nevado de Toluca. Investigaciones Geográficas, 1, 7–21. https://doi.org/10.14350/rig.59058


