Dung beetles (Coleoptera: Scarabaeidae, Scarabaeinae) and dung removal in Mexican livestock pastures
Carmen Huerta *, Lucrecia Arellano, Magdalena Cruz
Red de Ecoetología, Instituto de Ecología A.C., Carretera Antigua a Coatepec 351, El Haya, 91070 Xalapa, Veracruz, Mexico
*Corresponding author: carmen.huerta@inecol.mx (C. Huerta)
Abstract
In the present article, a summary and analysis of literature published in Mexico on the removal and burial of dung by dung beetles, a primary function that represents an ecosystem service of great importance for livestock production, is presented. Studies from distinct ecosystems are examined, emphasizing the findings in livestock pastures with active grazing in several different states of Mexico. In addition, the unpublished results of the authors obtained in pastures across an altitudinal gradient in central Veracruz are presented for a dung exposure time of 24 hours. No significant differences were found in species richness among sites. However, differences were found in the dung removal rate; higher values were found at the sites with higher elevation and lower values at sites with lower elevation. The species with the highest biomass included the 3 largest species and small but very abundant species. Nocturnal tunnelers reincorporated the largest quantity of dung over the first 24 hours; in contrast, rollers only removed dung at the lowest areas of the gradient (< 250 m asl). However, much more information is still required regarding the capacity of the different functional groups and dung beetle species for dung burial in the livestock production zones of Mexico.
Keywords:
Tunnelers; Rollers; Dwellers; Waste degradation; Relocation; Ecosystem services
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Escarabajos coprófagos (Coleoptera: Scarabaeidae, Scarabaeinae) y remoción de estiércol en pastizales ganaderos mexicanos
Abstract
Se presenta una síntesis y análisis de lo publicado en México sobre remoción y enterramiento de estiércol por escarabajos estercoleros (Coleoptera: Scarabaeinae), función primaria que se traduce en servicios ecosistémicos importantes para la ganadería. Se analizan estudios realizados en distintos ecosistemas, haciendo énfasis en lo encontrado en potreros con pastoreo activo en México. Incluimos resultados inéditos obtenidos en potreros localizados en un gradiente altitudinal de la zona central de Veracruz, para un tiempo de exposición del estiércol de 24 horas, donde no hubo cambios importantes en la riqueza de especies entre sitios, pero sí en la tasa de remoción de estiércol que mostró una tendencia positiva con la altitud. La mayor biomasa estuvo representada por las 3 especies más grandes y 1 pequeña, pero muy abundante. Los escarabajos cavadores nocturnos reincorporaron la mayor cantidad de estiércol durante las primeras 24 horas. Los rodadores sólo realizaron remoción de estiércol en las zonas más bajas del gradiente (< 250 m snm). Aún hace falta mucha información sobre la capacidad de enterramiento del estiércol que tienen los grupos funcionales y las diversas especies de escarabajos coprófagos en las zonas ganaderas de nuestro país.
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Palabras clave:
Cavadores; Rodadores; Moradores; Degradación de residuos; Relocalización; Servicios ecosistémicos
Introduction
Dung beetles (Coleoptera: Scarabaeidae, Scarabaeinae) differentially contribute to the daily removal, burial, and disintegration of large quantities of dung deposited by animals onto soil. Specifically, dung beetles relocate such organic wastes and use them for food and nesting (Doube, 1990; Halffter & Edmonds, 1982; Halffter & Matthews, 1966; Hanski & Cambefort, 1991; Zunino, 1991). This primary function represents an ecosystem service in both natural ecosystems and agroecosystems (Nichols et al., 2008), as it facilitates the degradation and recycling of nutrients contained in dung, such as nitrogen, phosphorous, potassium, sodium, and magnesium, among others, and also promotes the incorporation of those nutrients into soil (Bang et al., 2005). This function additionally serves to reduce the emission of greenhouse gases produced by dung, such as methane and nitrous oxide (Penttilä et al., 2013; Slade et al., 2016; Yokoyama et al., 1991). For this reason, it is important to study dung removal not only from an ecological point of view but also because of its direct connection with livestock production.
The earliest dung removal studies in Mexico were carried out in 2004. However, as in other parts of the world, the number of studies greatly rose by 2012 with increasing interest in the process of dung removal by Scarabaeinae beetles. In addition, interest in the ecosystem services provided by these insects in natural ecosystems and agroecosystems has continued to increase. Nevertheless, the number of sites and states in Mexico where these studies have been conducted are relatively few (Fig. 1). This study presents a summary and analysis of the material published on dung removal in Mexico, emphasizing those studies carried out in grazing pastures. Some data produced and recorded by the authors at active grazing sites are also presented.
The structure of this summary is as follows: first, a general overview is provided of the process of dung removal, burial, and disintegration by dung beetles (Scarabaeinae), including its importance and highlighting the different variables that, according to the literature, are closely related with this process. Based on literature published in Mexico (Table 1), the relationship among different factors that can directly or indirectly influence the efficiency of dung removal by dung beetles is examined. These factors include a) the richness, abundance, biomass, and composition of species and their relationship with dung removal; b) dung removal rates; c) functional groups and their differential contribution toward dung removal; d) species preference according to origin of dung or season; e) exposure time; f) extrinsic and intrinsic factors, and g) contribution of dung beetles to dung removal. Some findings obtained by the research team of the authors are included (both published and non-published). These are the results of a line of study of dung removal processes that has been established for more than 10 years. The unpublished data were obtained using the same methodology (Arellano, 2016), over the same period in different active grazing pastures of central Veracruz, Mexico, and cover a broad range of conditions (Table 2). Finally, we highlight the economic importance of the ecosystem services provided by dung beetles for livestock producers.
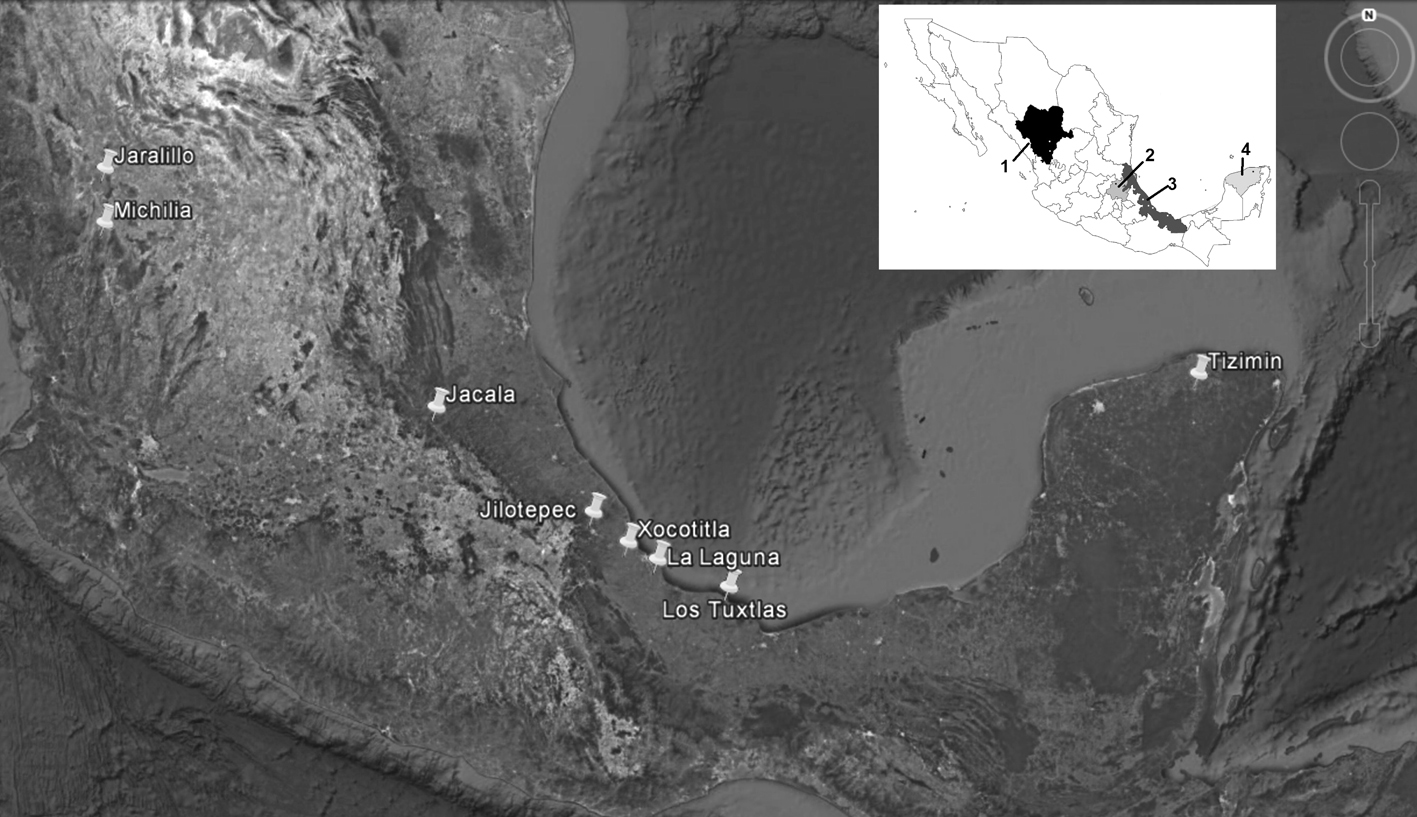
Table 1
List of articles about dung removal by dung beetles (Coleoptera: Scarabaeidae, Scarabaeinae) conducted in Mexico up to June 2017.
|
State |
Location |
Subject themes |
Authors |
|
Durango |
El Jaralillo |
Diversity and abundance of species, dung removal, functional groups, seasonality |
Anduaga, 2004 |
|
Durango |
La Michilía Biosphere Reserve. |
Species abundance, dung removal, seasonality, size, functional groups, reproduction, larval development |
Anduaga & Huerta, 2007; Huerta et al., 2010 |
|
Veracruz |
Los Tuxtlas Biosphere Reserve |
Excreta removal rates, origin of dung (native and exotic), seasonality, richness and biomass of species. |
Amézquita & Favila, 2010. |
|
Veracruz |
La Laguna, Medellín de Bravo |
Dung degradation, seasonality, abundance and diversity of species |
Cruz et al., 2012 |
|
Hidalgo |
Neighbor to Jacala de Ledezma, Sierra Madre Oriental |
Dung removal, functional groups, rollers |
Ortega-Martínez et al., 2014 |
|
Dung removal, functional groups, types of excreta |
Ortega-Martínez et al., 2016 |
||
|
Yucatán |
Tizimín |
Richness and abundance of species, patterns of activity |
Basto- Estrella et al., 2012 |
|
Biomass of species, removal, functional groups |
Basto- Estrella et al., 2016 |
Table 2
Description of the plots studied in central Veracruz. The new information incorporated into this review article was obtained in August 2014, in ranching pastures (1 ha) with active cattle grazing, in 3 different municipalities of central Veracruz: Jilotepec (J), Tlacolulan (T) and Paso de Ovejas (PO). The average distance between sites in Jilotepec and Tlacolulan was 3 km. In the Municipality of Paso de Ovejas, the average distance was a little greater (6.9 km). The period of exposure of the cattle dung was 24 hours, using in each case 8 sampling units per site and following the methodology proposed by Arellano (2016).
|
Municipalities and sites |
Type of system |
Geographic location (UTM) |
Elevation (m asl) |
|
1 – (PO) Xocotitla |
Pasture with the following grasses: llanero (Andropogon gayanus, estrella (Cynodon plectostachyus) and privilegio (Megathyrsus maximus) |
E(764155), N(2125524) |
179 |
|
2 – (PO) Xocotitla |
Native grass pasture |
E(757826), N(2124470) |
266 |
|
3 – (J) La Concepción – Vista Hermosa |
Guinea grass (Megathyrsus maximus) |
E(720158), N(2170119) |
1,032 |
|
4 – (J) La Concepción – Vista Hermosa |
Guinea grass |
E(719447), N(2170481) |
1,062 |
|
5 – (J) La Concepción – Vista Hermosa |
Estrella grass |
E(716864), N(2170488) |
1,231 |
|
6 – (J) Jilotepec – Tlacolulan |
Estrella grass |
E(712871), N(2169683) |
1,628 |
|
7 – (J) Jilotepec – Tlacolulan |
Estrella grass |
E(713241), N(2171663) |
1,654 |
|
8 – (J) Jilotepec – Tlacolulan |
Estrella grass |
E(710369), N(2173747) |
1,861 |
General overview: removal, burial, and disintegration of dung by dung beetles
As stated above, the removal, burial and disintegration of dung is a primary ecological function performed by the Scarabaeinae that translates into an ecosystem service by improving the physical properties of the soil, such as aeration, water infiltration, and reducing compaction through bioturbation and the construction of tunnels (Brown et al., 2010; Huerta et al., 2013; Nichols et al., 2008). Dung beetles also improve the chemical properties of soil by facilitating the degradation, recycling, and incorporation of nutrients contained in dung, such as nitrogen, phosphorus, potassium, sodium, and magnesium, among others (Bang et al., 2005; Bertone et al., 2006).
The accumulation of dung for a prolonged period in the pastures or active grazing sites can have negative economic effects for the livestock producers, who must pay to remove it because of the sanitary problems it can create, as well as to control the pests that develop within it (Corwin, 1997), but also because of the loss of useful pasture for the livestock which its presence implies (Cruz-Rosales, 2011; Forbes & Hodgson, 1985). In addition, the presence of bare soil generates erosion and increases compaction, thus reducing the infiltration of rainwater (Pinheiro-Machado, 2016; Valdez-Zertuche et al., 2015). The producer also pays for the chemical inputs necessary to compensate for the nutrient loss (Aarons et al., 2004; López-Collado et al., 2017; Omaliko, 1981). In this context, the ecosystem services provided by the dung beetles play a vital role.
The Scarabaeinae have 3 functional groups, according to dung removal strategy (Doube, 1990; Doube et al., 1988; Halffter & Edmonds, 1982; Halffter & Matthews, 1966; Zunino, 1991): tunneler species are characterized by the excavation of tunnels below the dung pat, where they store the dung, thus contributing to its removal and to the aeration of the soil, facilitating water infiltration from the surface to the deeper soil layers (Bang et al., 2005; Brown et al., 2010; Nichols et al., 2008). The roller species form balls of dung and disperse these at a distance from where the dung pat was originally deposited. These species thus act as secondary dispersers of the seeds contained within this dung (Amézquita & Favila, 2010; Andresen, 2002; Andresen & Feer, 2005; Estrada & Coates-Estrada, 1991; Huerta et al., 2013; Nichols et al., 2008; Slade et al., 2007). Finally, dweller species are characterized by feeding and nesting within the dung itself, causing its disintegration. While these species do not remove the dung, they do break it down (Scholtz et al., 2009), for which reason all of the dung beetle species are important for dung elimination and the cleansing of livestock pastures (Anduaga, 2004; Anduaga & Huerta, 2007; Doube, 1990; Halffter & Edmonds, 1982; Halffter & Matthews, 1966; Hanski & Cambefort, 1991; Huerta et al., 2013; Kryger, 2009).
It has been observed that the ecosystem services provided by the dung beetles are closely related to different environmental factors, such as seasonality, temperature and humidity (Anduaga & Huerta, 2007; Ortega-Martínez et al., 2016; Slade et al., 2007), as well as the physical characteristics and type of dung (Amézquita & Favila, 2010; Horgan, 2001; Ortega-Martínez et al., 2016) and soil (Menegaz & Medina-Hernández, 2017; Nervo et al., 2014), type of vegetation (Lizardo & Castellanos-Vargas, 2016; Ortega-Martínez et al., 2016), among others. However, the efficiency with which this process is conducted also depends on other factors particular to each species, such as richness and abundance (Braga et al., 2013; Ortega-Martínez et al., 2016; Slade et al., 2007), size (Amézquita & Favila, 2010; Anduaga, 2004; Anduaga & Huerta, 2007; Nervo et al., 2014; Slade et al., 2007), period of activity, behavioral type displayed in terms of the removal of food (Halffter & Edmonds, 1982; Halffter & Matthews, 1966; Hanski & Cambefort, 1991), biomass (Amézquita & Favila; 2010; Basto-Estrella et al., 2016; Braga et al., 2013; Ortega-Martínez et al., 2016; Shahabuddin, 2008), and reproductive status (Andresen, 2003; Carpaneto et al., 2010; Halffter & Edmonds, 1982; Huerta et al., 2010; Jankielsohn et al., 2001; Nervo et al., 2014; Nichols et al., 2008; Ortega-Martínez et al., 2016; Shepherd & Chapman, 1998; Woodward et al., 2005).
Although dung beetles modify agroecosystem soils, the management of production (in this case, livestock production) can, in turn, affect dung beetle assemblies (Alvarado et al., 2018; Lumaret et al., 1992; Williams et al., 2017). Such effects may be manifested in changes in the number and composition of the species, in the relative abundance of the functional groups within the assembly and in the ecological functions of the beetles themselves (Braga et al., 2013).
Factors that affect dung removal function in Mexico
Richness, abundance, biomass and composition of species, and their relationship to dung removal. The quantity of dung removed is closely related to the richness, biomass, and abundance of the assemblies of dung beetles (Braga et al., 2013). However, in general, the literature consulted in this regard shows that richness is not a determinant variable in the assemblies. The determinant variable is in fact the composition of species and their relationships with abundance and biomass at each site and study area.
In Durango State, Mexico, in El Jaralillo, the highest values of abundance, biomass and rates of dung removal were presented during July, September and October. The most important species were Euoniticellus intermedius Reiche, Digitontophagus gazella Fabricius, 2 exotic species and the native species Canthon humectus Say (Anduaga, 2004). In the locality of Minillas, the highest abundance of species were found in September, with the dominant species being those of larger size: Dichotomius colonicus Say, Phanaeus quadridens Say, and Copris sierrensis Matthews. This was reflected directly in the proportion of dung removed by these species (Anduaga & Huerta, 2007).
In livestock production ranches, neighbor to Jacala, Hidalgo State, that feature remnants of temperate forest, Ortega-Martínez et al. (2016) found that beetle species richness was similar in both habitat types and that there was a significant relationship between the biomass of these species and the quantity of dung removed. These authors found differences in the quantity of dung removed by the most abundant species: Deltochilum scabriusculum Bates, D. colonicus, and Phanaeus adonis Harold.
In Los Tuxtlas in southern Veracruz, a study by Amézquita and Favila (2010) conducted in tropical rain forest indicated that biomass and species richness, as well as season, were important factors for the removal of dung. Dung type (native represented by monkey dung, and exotic represented by cow dung) also affected the dung removal rates. Dung beetle biomass and species richness were near 3 times greater in monkey dung than in cow dung.
In Medellin, Veracruz, Cruz et al. (2012) compared the dung removal rate in areas that had free access or that were covered with mesh during the rainy and dry season. The greatest and fastest rate was observed during the rainy season, which coincides with the period of greater abundance of dung beetles in this area, mainly E. intermedius, D. gazella, and Copris lugubris Boheman, which are also present throughout the year (Flota-Bañuelos et al., 2012).
In the pastures we studied in central Veracruz (Municipalities of Jilotepec, Tlacolulan, and Paso de Ovejas), no important differences were observed among the sites: in pastures located between 1,000 and 1,840 m asl, there were 3 to 5 species, and in those found below 300 m asl, there were 5 to 7 species. With respect to abundance, in the pastures below 300 m asl, E. intermedius and D. gazella were the most abundant species, supporting that reported by Anduaga (2004) for El Jaralillo, Durango. In the pastures at between 1,000 and 1,200 m asl, the most abundant species was D. colonicus, which is similar to that reported by Anduaga and Huerta (2007) in the locality of Minillas, Durango. At this same elevation in Central Veracruz, Copris incertus Say was found to be dominant. In zones above 1,600 m asl, the dominant species were Onthophagus incensus Say and Onthophagus cyanellus Bates.
In terms of biomass, 3 species represented the highest biomass in the pastures located at elevations below 300 m asl; however, these were not those of greatest abundance, but rather the species of largest size: D. colonicus, D. scabriusculum, and Phanaeus tridens Laporte. In the pastures located between 1,000 and 1,200 m asl, the 2 most abundant species also represented the greatest biomass. In the pastures located above 1,600 m asl, 2 species represented the greatest biomass: D. colonicus for its size and O. incensus for its abundance. The list of species, and their abundances per site and elevation is presented in table 3. As with the studies analyzed above, there was a direct relationship along the gradient between the abundance, composition and biomass of the species and the quantity of dung removed.
In the municipality of Tizimín, Yucatán State, a total of 17 species were observed, with Onthophagus landolti Harold the most abundant, while Canthon indigaceus chevrolati Harold, was the second most abundant (Basto-Estrella et al., 2012). Due to the high abundances of these 2 species (with more than 20,000 individuals each), they were important to the process of dung removal despite their small size.
It can therefore be concluded that the quantity of dung that can be removed in a given site can be defined according to the species composition and their abundance in each dung pat as well as the dry weight of the dung itself (Braga et al., 2013).
Quantification of the dung removal rate. One of the most utilized variables at international level is the difference between the quantity of dung deposited and the quantity found after a given period, or under given land uses, commonly known as the dung removal rate. In this respect, it was found in El Jaralillo, Durango, that the highest dung removal rate was presented in October, coinciding thus with the highest species richness (Anduaga, 2004). For this reason, in the locality of Minillas in the same state, Anduaga and Huerta (2007) found that September was when the highest quantity of dung was removed (69%), which was related to the high activity of the dominant species recorded at that time. In Jacala, Hidalgo, Ortega-Martínez et al. (2016) found that dung removal was significantly greater in the pastures that in the forest, which was related to the abundance of the most important species in each habitat type. However, these authors do not provide the removal rate as such. In La Laguna, municipality of Medellín, in the central lowland zone of Veracruz, Cruz et al. (2012) observed that after 4 days, 86% of the dung placed in uncovered dung pats had disappeared, with nearly 100% eliminated after a period of 32 days. In central Veracruz, we found that the average dung removal rate (± SE) was 0.171 ± 0.024 in the zone located at more than 1,600 m asl, 0.146 ± 0.015 in pastures located between 1,000 and 1,200 m asl and 0.112 ± 0.01 in those located below 300 m asl. A decreasing trend was observed in the dung removal rate with elevation, although this was not significant, since the dung removal rate did not differ drastically, only varying from 11 to 17% in a period of 24 hours. In Tizimín, Yucatán, Basto-Estrella et al. (2016) indicated that the dung beetles removed 40.1 % of the initial weight of the dung pat and that the removal activity on their part was initially less rapid (33.8%) than at the end of the study period (44%). It can therefore be concluded that in the different studies analyzed, variable removal rates were changing according region, area, season, and exposure time.
Dung removal and functional groups: contribution of the species to dung removal. The composition of the dung beetle functional groups that comprise an assembly, is closely related to the quantity of dung removed by each of these groups (Halffter & Edmonds, 1982; Halffter & Matthews, 1966; Hanski & Cambefort, 1991; Scholtz et al., 2009). Most studies conducted in Mexico have shown that, as in other parts of the world, the tunneler species of greater size are those that remove the highest quantity of food following emergence and during the nesting period (Anduaga & Huerta, 2007; Halffter & Edmonds, 1982; Huerta et al., 2010; Scholtz et al., 2009). However, the removal conducted by the roller species has also been quantified in Mexico. Below, we present the findings of various studies in this regard.
Table 3
List of coprophagous species recorded in pastures with active grazing in central Veracruz. Si = size, G = large, M = medium, S = small. DA = daily activity, Di = diurnal, N = nocturnal, C = crepuscular. RF = removal of food, R = roller, T = tunneler, D = dweller*. Sample coverage was close to 100% in all studied areas.
|
Si |
DA |
RF |
Species |
Sites/ elevation (m asl) |
Totals |
|||||||
|
1/179 |
2/266 |
3/1032 |
4/1062 |
5/1231 |
6/1628 |
7/1654 |
8/1861 |
|||||
|
M |
Di |
R |
Canthon indigaceus chevrolati Harold, 1868 |
8 |
12 |
0 |
0 |
0 |
0 |
0 |
0 |
20 |
|
L |
N |
T |
Copris incertus (Say), 1835 |
0 |
0 |
6 |
8 |
12 |
0 |
0 |
0 |
26 |
|
L |
N |
T |
Copris lugubris Boheman, 1868 |
4 |
0 |
0 |
0 |
0 |
2 |
0 |
0 |
6 |
|
L |
N |
R |
Deltochilium scabriusculum Bates, 1887 |
3 |
5 |
0 |
0 |
0 |
0 |
0 |
0 |
8 |
|
L |
N |
T |
Dichotomius colonicus (Linneo), 1767 |
4 |
1 |
39 |
16 |
1 |
45 |
30 |
0 |
136 |
|
M |
C |
T |
Digitonthophagus gazella (Fabricius), 1787 |
16 |
12 |
1 |
0 |
0 |
0 |
0 |
0 |
29 |
|
S |
Di |
T |
Euoniticellus intermedius Reiche, 1849 |
35 |
22 |
2 |
1 |
1 |
0 |
0 |
0 |
61 |
|
L |
Di |
D |
Eurysternus magnus Castelnau, 1840 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
|
L |
N |
T |
Ontherus mexicanus Harold, 1869 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
34 |
37 |
|
M |
N |
T |
Onthophagus cyanellus Bates, 1887 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
165 |
165 |
|
M |
Di |
T |
Onthophagus incensus (Say), 1835 |
0 |
0 |
3 |
1 |
2 |
637 |
192 |
75 |
910 |
|
L |
Di |
T |
Phanaeus tridens Castelnau, 1840 |
5 |
10 |
0 |
0 |
0 |
0 |
0 |
0 |
15 |
|
M |
Di |
T |
Scatimus ovatus Harold, 1862 |
0 |
0 |
0 |
0 |
0 |
0 |
2 |
0 |
2 |
|
Total number of individuals |
75 |
62 |
51 |
26 |
16 |
687 |
225 |
274 |
1,416 |
|||
|
Total number of species |
7 |
6 |
5 |
4 |
4 |
4 |
4 |
3 |
Of the studies conducted in Durango, in El Jaralillo, Anduaga (2004) found the presence of both tunnelers and rollers, but did not measure removal per functional group, only the total removal. Anduaga and Huerta, (2007) and Huerta et al. (2010), highlighted that, in Minillas for example, while 2 functional groups do exist (tunnelers and rollers), the only one found was that of the tunnelers, of which the species of largest size (D. colonicus, P. quadridens, and C. sierrensis) removed the most dung over the rainy season, when they come out to feed, mature sexually, and reproduce. Of these species, D. colonicus was the large tunneler that removed the highest quantity of dung in September (5 to 135 g of dung per individual or pair, respectively), followed by C. sierrensis, another large tunneler species with around 30 g of dung removed per pair and, finally, P. quadridens, with the same characteristics as those described before, with around 15 g per pair. This study did not analyze the role of the species of smaller size in dung removal, nor that of the dweller species.
In central Mexico, specifically in the Barranca de Meztitlán and close to Jacala, Hidalgo State, one of the most abundant species of the zone was the roller species Canthon humectus hidalgoensis (Bates), which could bury an estimated 820 g of dung per day and per hectare. This figure was determined from determining the weight of the dung balls made by the pairs and by individuals alone, in relation to the abundance present in the season of greatest activity of this species (Ortega-Martínez et al., 2014). In a subsequent study, Ortega-Martínez et al. (2016) found that the greatest individual dung removal rate was represented by the 3 species of greatest size and biomass, but of different habits; D. colonicus is a tunneler, nocturnal and coprophagous, D. scabriusculum is a roller, nocturnal and copro-necrophagous and P. adonis is a tunneler, diurnal and copro-necrophagous in habit.
In the region of Los Tuxtlas, Veracruz, monkey dung attracted beetle species belonging to all relocation strategies: dwellers (3 species of Eurysternus), rollers (5 species of Canthon, and 2 of Deltochilum) and tunnelers (9 species of different genera). In contrast, cattle dung mainly attracted tunnelers (5 species) and species of dweller (Eurysternus mexicanus), although individual of Canthon femoralis, which is commonly associated with monkey dung, was recorded (Amézquita & Favila, 2010). The latter authors propose that these associations are related to the coevolution of these beetle species and the native organisms that produce their food, which in this zone are mainly omnivorous mammals, such as monkeys, rather than herbivorous animals, such as cattle, which were introduced to America over 500 years ago (Halffter & Matthews, 1966; Vieira et al., 2008).
In La Laguna, in the municipality of Medellín, Veracruz, there were 2 functional groups of dung beetles present; the tunnelers and some dwellers, who conducted the dung removal over the 2 months of field experimentation by Cruz et al. (2012).
Of the 13 species in central Veracruz, most were large tunnelers, and only 2 were rollers (C. indigaceus chevrolati and D. scabriusculum). Accordingly, members of the tunneler functional group removed the highest proportion of dung (greater number of species and higher abundance = more removed dung).
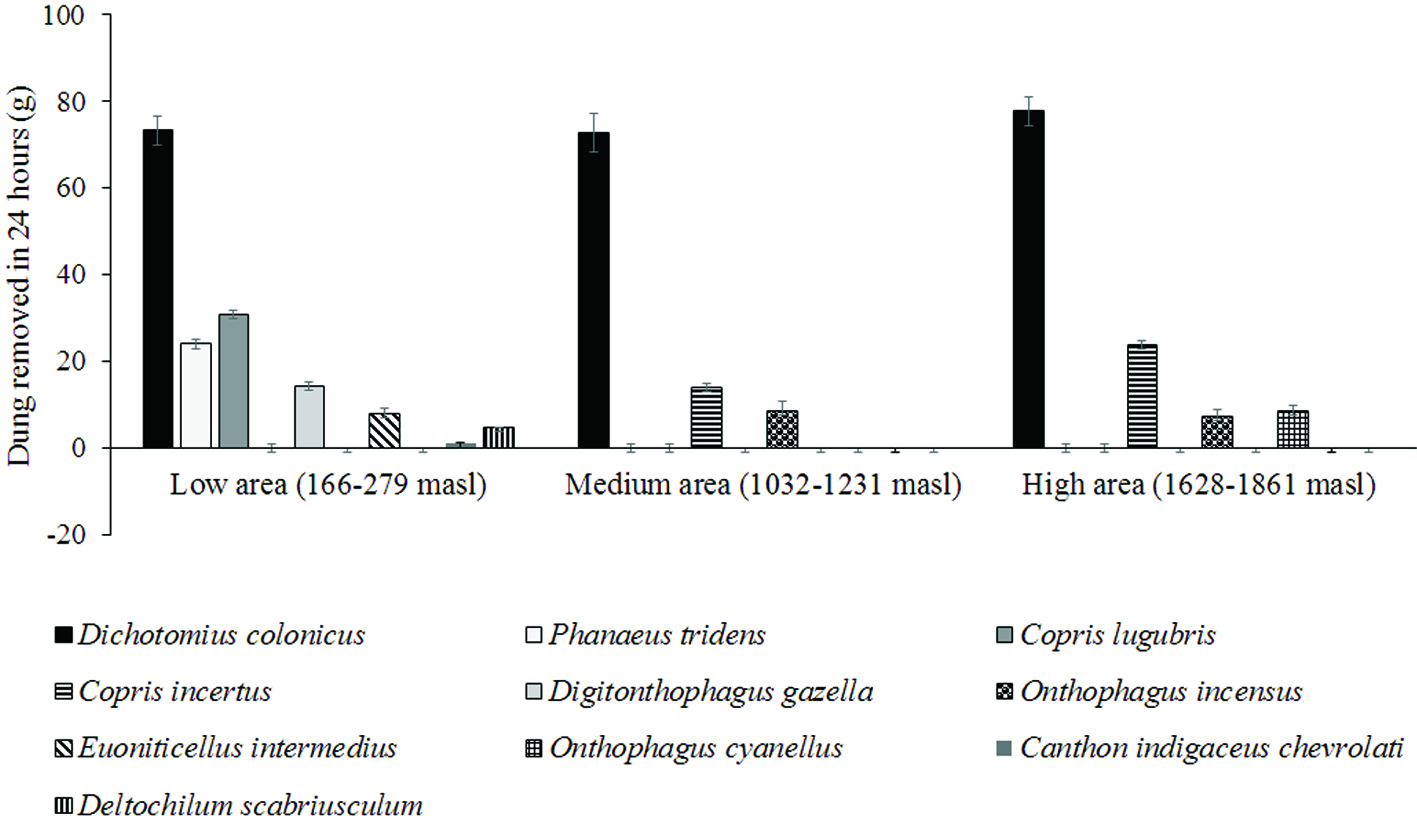
Dung removal by species differed independently of functional group according to biomass and length (Fig. 2). All of the results we describe hereafter correspond to a 24-hour period of exposure of the dung. In the pastures located below 300 m asl, individuals of D. gazella and of E. intermedius bury small quantities of dung, which is related to their small size, and their role in the function of dung removal is compensated with their high abundance in these sites, where individuals of larger and less abundant species, such as D. colonicus, bury more than 5 times the quantity of dung and individuals of P. tridens bury almost double (Fig. 2). In pastures located between 1,000 and 1,200 m asl, D. colonicus, a large tunneler species bury similar quantities of dung than at the lower elevation. C. incertus, a large tunneler species, buries almost half the quantity of dung than it buries in pastures located above 1,600 m asl. The species D. colonicus buries quantities that are a little larger than those it buries at the medium and low elevations. Onthophagus incensus is a medium-sized tunneler species that is very abundant from 1,600 m asl and buries similar quantities between 1,000 and 1,800 m asl; however, it presents its highest values above 1,600 m asl (Fig. 2). Notably, the rollers were only found at pastures below 300 m asl. Canthon indigaceus chevrolati was the roller species with the highest abundance (< 300 m asl); it removed 6.95 ± 0.36 g per day from the dung pads (a similar but slightly lesser amount than E. intermedius and a higher amount than Onthophagus landolti). On the other hand, individuals of Deltochilum scabriusculum (the other roller species) removed 21 ± 0.56 g per day from the dung pads (less than P. tridens and C. lugubris) (Fig. 2). Likewise, the dung balls of C. indigaceus were smaller than those of D. scabriusculum (1 g and 5 g respectively).
In Tizimín, Yucatán, the presence of 2 of the 3 functional groups were reported: tunnelers and rollers. However, the former removed more dung due to presenting more than double the abundance of the latter group (Basto-Estrella et al., 2016).
In summary, the studies conducted in Durango mainly analyzed the role of large tunnelers in the ecosystem function of dung removal, while the roles of smaller species and dwellers were not explored. Of the studies conducted in Hidalgo, only one determined the importance of the different removal strategies (Ortega-Martínez et al., 2016) through considering the entire assembly rather than simply one very abundant species in the area. This latter study concluded that the greatest individual dung removal was performed by the 3 species of greatest size and biomass but of differing habits in terms of both dung removal strategies and daily activity and food preferences.
The studies conducted in Veracruz arrived at different conclusions: in the region of Los Tuxtlas, monkey dung (a native resource in the tropical forest) attracted beetles of all 3 dung removal strategies and, for this reason, the dung removal rates were higher in this dung type than in that of cattle (an exotic resource in the tropical forest), which attracted mainly tunnelers (Amézquita & Favila, 2010). In La Laguna, in the municipality of Medellín, Veracruz, there were 2 functional groups, with the tunnelers being the most important of these, in terms of dung removal (Cruz et al., 2012). This coincides with that found along the studied gradient in the central zone of the state, since the large nocturnal tunnelers were those that dominated the dung removal process, as well as small but very abundant diurnal tunneler species (Onthophagus incensus). The rollers made a very low contribution, since they represented only 15% of the species of the assembly and had very low abundances. Finally, in Yucatán, the small tunnelers removed the greatest quantity of dung due to their high abundance (Basto-Estrella et al., 2016).
Effect of species preference, origin of dung and seasonality on dung removal. The animal origin of the dung and the species on which those animals feed are both very important factors, since there are species that prefer different types of dung, both for feeding and nesting, and such preferences could thus be related to the observed removal rates. Another important factor is seasonality on the part of the dung (Amézquita & Favila, 2010; Anduaga, 2004; Anduaga & Huerta, 2007; Basto-Estrella et al., 2016; Estrada et al., 1993; Halffter & Matthews, 1966; Huerta et al., 2010; Whipple & Hoback, 2012), since most dung beetle species are active during the rainy season.
In Durango, the dung of the pastures of both El Jaralillo and Minillas are mainly bovine in origin and the highest beetle activity is presented in the rainy season. In El Jaralillo, the highest quantity and activity of species was found in September, when the rainy season is well established (Anduaga, 2004) while, in Minillas, most species were active from the end of May until September (Anduaga & Huerta, 2007). For this reason, the greatest removal of dung is found in this season, which is also the reproductive period of the beetles (Huerta et al., 2010). During the rainy season (June to October) neighbor to Jacala in Hidalgo, no differences were found between the quantities of bovine and ovine dung removed (Ortega-Martínez et al., 2016).
In Los Tuxtlas, Veracruz, in tropical rain forest fragments of different sizes, Amézquita and Favila (2010) found that monkey dung removal rates remained more or less similar during the dry and rainy seasons. In contrast, cattle dung removal rates were very much higher during the rainy season. In La Laguna, municipality of Medellín de Bravo, Veracruz, Cruz et al. (2012) also report that the greatest activity of the beetles and removal of bovine dung takes place in the rainy season. In the pastures of central Veracruz, we only used bovine dung and the results we present are from the rainy season, which is when the highest abundances of species are presented in the region studied, coinciding with that found in the pastures of Tizimín, Yucatán, where most bovine dung removal was presented during July. This date corresponds to the middle of the rainy season, which is when the highest species abundance is presented (Basto-Estrella et al., 2016).
The studies analyzed were mostly conducted in the rainy season and it is during this period that the beetles are most active and contribute most to the function of dung removal.
Dung removal and exposure time. One important variable that can influence the quantity of dung removed by beetles is the exposure time of dung, as many species are most attracted to fresh dung (Lee & Wall, 2006). Specifically, the attraction of dung decreases over time because of the effects of desiccation (Cruz et al., 2012). Anduaga (2004) states that, in montane pastures in Durango, a higher proportion of dung pats were exploited or colonized during the first 48 hours following deposition. In Minillas, Anduaga and Huerta (2007) found that removal is also related to the months of the year in which beetles are most active, as more dung was removed in September than in October.
In Los Tuxtlas, Veracruz, Amézquita and Favila (2010) found that the speed of elimination of both monkey and cattle dung was high between 24 and 48 hours; after this period, the consumption of both dung types decreased. Cruz et al. (2012) observed that the greatest reduction in weight of experimental dung pats in La Laguna occurred over the first 4 to 8 days of exposure and subsequently decreased. Greater reduction in dung pat weight occurred during the rainy season, coinciding with the period of highest activity of dung beetles. In central Veracruz, the highest removal was generally observed at 24 hours after deposition, which is when bovine dung is most fresh and attractive to a high percentage of the species present in our study areas. We can therefore conclude that the highest dung removal activity of beetles is presented between 24 and 48 hours of dung exposure, although peaks in activity may occur up to 96 hours after deposition.
Extrinsic and intrinsic factors related to dung removal. Dung removal is one of the most important ecosystem services provided by the dung beetles and there is a series of extrinsic and intrinsic factors related to the removal rates and efficiency of species in terms of this activity. The former, as stated in the introduction, include seasonality, temperature, precipitation, humidity, soil characteristics, availability and type of food and pasture management, among others. In the case of the intrinsic factors, the most important are the period of activity of the individuals (diurnal, nocturnal and crepuscular), their physiological condition, and the sex and behavior type displayed by each (Anduaga & Huerta, 2007; Horgan & Fuentes, 2005; Ortega-Martínez et al., 2016; Slade et al., 2007).
In El Jaralillo, Durango, Anduaga (2004) analyzed the efficiency of dung beetles and found that, even with the abundance and activity of the species found there, it would be insufficient to achieve the total removal of the dung deposited by the cattle in this semiarid zone of marked seasonality. Due to the fact that the soil begins to moisten with the first rains in July, the quantity of species that emerge at the beginning of the rainy season is few and it is not until October, when there has been sufficient rainfall to penetrate deep into the soil layers, that the highest emergence, richness and abundance of beetle species is presented. This undoubtedly has influence in terms of the low dung removal rates found by this author. Moreover, most of the beetle species found are of small (7 to 13 mm) and medium (14 to 17 mm) sizes, while large-sized species (20-25 mm) were very rare. In Minillas, also in Durango and with very similar environmental conditions to those of El Jaralillo, Anduaga and Huerta (2007) found that emergence of the larger sized species (20-24 mm), and those that are univoltine, is presented in mid-July or the beginning of August, such that it is not until September that their greatest activity is presented and thus the highest percentage of dung removed. However, when this study was conducted, only 69% of the dung pats presented activity and removal on the part of the dung beetles, while in October the percentage of dung removed was lower, even though availability of this resource was greater than in September. This was corroborated by Huerta et al. (2010), who observed that the greatest quantity of removal was conducted during the reproductive season of the beetle species of larger size.
In Jacala, Hidalgo, the livestock pastures still feature remnants of the original temperate forest vegetation. The rainy season is presented from June to September and the greatest richness and abundance of dung beetle species is also presented during this period (Ortega-Martínez et al., 2014, 2016). In this zone, 19 species are found according to Ortega-Martínez et al. (2016). Most of the dung removal is conducted by 2 nocturnal and diurnal species, all of which present high biomass and large size. No differences were found between the removal of bovine and ovine dung.
In Los Tuxtlas, Veracruz, Amézquita and Favila (2010) do not include the influence of environmental factors, or of the periods of activity of the species, although it is known that some are nocturnal while others are diurnal.
Unlike the other studies that had short observation periods, in La Laguna, Veracruz, Cruz et al. (2012) exposed the dung pats for 2 months in order to observe the rates of degradation in both dry and rainy seasons. The differences were marked, since in the rainy season, 80% of the dung pat weight was lost within 4 days, while in the dry season, this same weight loss required 2 weeks of exposure. The weight could be maintained with little variation for up to 2 months. In both cases, this degradation was also related to the activity of the beetles present in the zone, represented mainly by E. intermedius and D. gazella. Flota-Bañuelos et al. (2012) also observed that these 2 dung beetle species are prominent among the 15 species recorded in that zone. The authors even mention that E. intermedius was present throughout the year, although in variable abundance. Cruz et al. (2012) found that, during the rainy season, when the dung beetles were present, dung pats exposed to them presented a considerable reduction in weight, since in 4 days they had lost 86% of their initial weight, in 1 month they had lost 98% and in 2 months the dung pats had disappeared completely.
In central Veracruz (municipalities of Jilotepec and Paso de Ovejas), while dung removal data were taken at 24, 48 and 72 h, we only report the results corresponding to 24 hours in this study since this was the period in which we obtained the highest values of removal by the beetles. The majority of the dominant and most efficient species at dung burial were nocturnal, although some were diurnal (31% of the total).
In Tizimín, Yucatán, where at present patches of secondary vegetation of low and medium deciduous tropical forest are found within the livestock pastures, the assembly of dung beetles was formed by 19 species, of which 10 are nocturnal, 8 diurnal and 1 crepuscular (Basto-Estrella et al., 2012). During the rainy season, 40.1% of the initial dung pat weight was removed in 24 hours by the beetles, the removal activity of which was initially slower (33.8%) than it was at the end of the season (44%) (Basto-Estrella et al., 2016).
From this, we can conclude that the extrinsic and intrinsic factors play an important role in the removal of dung and must be taken into account when attempting to make an adequate interpretation of the dung removal efficiency of each species and functional group.
Contribution of dung beetles to removal. Despite the activity, richness and/or abundance of the dung beetles, as well as the functional groups present in many regions of Mexico, it appears that it is insufficient to remove all of the fresh dung deposited daily on grazing lands, for which reason, much of this dung remains unused by the beetles. Recently, a mathematical model was used to estimate that only 65 to 69% of the available dung can be removed (López-Collado et al., 2017). As stated previously, various environmental factors are associated with dung removal. However, the seasonality defines the periods of activity of dung beetles, being more abundant and active during the rainy season, although there are species that can be active throughout the year (Anduaga, 2004; Anduaga & Huerta, 2007; Basto-Estrella et al., 2016; Cruz et al., 2012; Huerta et al., 2010; Ortega Martínez et al., 2014, 2016).
Knowledge of the rate of disintegration of dung has helped to establish the economic value of the ecosystem service provided by the dung beetles to livestock production sector of different zones (Beynon et al., 2015; Fincher, 1981; López-Collado et al., 2017; Losey & Vaughan, 2006). Broadening this type of study to the level of species or functional groups would contribute towards the improved definition of the importance of these to livestock production in Mexico.
Conclusions
In Mexico, large areas of territory originally occupied by tropical forests and other vegetation types have been cleared for livestock production since the past century. At present, livestock production zones occupy an area of 68 million hectares and contain more than 28 million head of cattle. Accordingly, the accumulation of dung in these agroecosystems is considerable.
It is very important to inform livestock producers of the important ecosystem services provided by dung beetles, such as facilitating the degradation of dung deposited daily in pastures in large quantities by livestock, during feeding and nesting periods. To achieve this, a sustained educational effort should be directed toward livestock producers. With such information, producers can become aware of the ecosystem services provided by these insects.
In Mexico, dung beetle fauna is still well represented, but populations of these insects can be affected in different livestock producing regions, such as Veracruz, by traditional livestock methods. Livestock production can have serious consequences for the existence of these beneficial organisms. In the absence of dung beetles, alternatives must be sought to recover the services provided by these organisms to livestock production and pasture agroecosystems, which would lead to a high economic cost for livestock producers, as occurred in Australia in the 1960s.
If this ecosystem service were to be lost, the estimated economic and ecological impacts for pasture agroecosystems would likely be enormous. Laboratory rearing of dung beetles is one potential means of addressing the loss of this ecosystem service. In addition, species may be reintroduced at suitable sites in terms of soil health where prior studies have documented the fauna inhabiting the area, which may imply a high cost.
Acknowledgements
The field studies in the Municipality of Jilotepec formed part of the second stage of the project “Hacia una ganadería sustentable” No. 20035 30893 of the Instituto de Ecología, A.C. We are grateful to the owners that granted permission to work on their properties: Silvino Huesca Libreros, Pedro Méndez Olivares, Victoriano Gómez Galicia, Luis Caraza Stoumen, Servando López Ramírez, Adrián Alberto Gómez Sangabriel, Armando Felipe Gutiérrez, Jorge Melchor and Ignacio Domínguez and family. To Albina Demeza, Teresa Lorenzo, Alfonso Díaz, Maira and Ignacio Domínguez, and Leonel Hernández Cortina, for helping with the fieldwork. Particular thanks go to Patricia Menegaz and Malva I. Medina Hernández, of the Universidad de Santa Catarina, who helped us with field and office work through the predoctoral internship of Patricia in the Instituto de Ecología, A.C. during the second semester of 2014. Keith MacMillan provided the English translation.
References
Aarons, S. R., O’Connor, C. R., & Gourley, C. J. P. (2004). Dung decomposition in temperate dairy pastures I. Changes in soil chemical properties. Australian Journal of Soil Research, 42, 107–114.
Alvarado, F., Escobar, F., Williams, D. R., Arroyo-Rodríguez, V., & Escobar-Hernández, F. (2018). The role of livestock intensification and landscape structure in maintaining tropical biodiversity. Journal of Applied Ecology, 55, 185–194.
Amézquita, S., & Favila, M. E. (2010). Removal rates of native and exotic dung by dung beetles (Scarabaeidae: Scarabaeinae) in a fragmented tropical rain forest. Environmental Entomology, 39, 328–336.
Andresen, E. (2002). Dung beetles in a Central Amazonian rainforest and their ecological role as secondary seed dispersers. Ecological Entomology, 27, 257–270.
Andresen, E. (2003). Effect of Forest fragmentation on Dung Beetle Communities and Functional Consequences for Plant Regeneration. Ecography, 26, 87–97.
Andresen, E., & Feer, F. (2005). The role of dung beetles as secondary seed dispersers and their effect on plant regeneration in tropical rainforests. In P. M. Forget, J. E. Lambert, P. E.Hulme, & Vander Wall, S.B. (Eds.), Seed fate: predation, dispersal and seedling establishment (pp. 331–349). Oxfordshire, UK: CABI International, Wallingford.
Anduaga, S. (2004). Impact of the activity of dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae) inhabiting pasture land in Durango, Mexico. Environmental Entomology, 33, 1306–1312.
Anduaga, S., & Huerta, C. (2007). Importance of dung incorporation activity by three species of coprophagous beetle (Coleoptera: Scarabaeidae: Scarabaeinae) macrofauna in pastureland on “La Michilía” Biosphere Reserve in Durango, Mexico. Environmental Entomology, 36, 555–559.
Arellano, G. L. (2016). A novel method for measuring dung removal by tunneler dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae) in pastures. The Coleopterists Bulletin, 70, 185–188.
Bang, H. S., Lee, J. H., Kwon, O. S., Na, Y. E., Jang, Y. S., & Kim, W. H. (2005). Effects of paracoprid dung beetles (Coleoptera: Scarabaeidae) on the growth of pasture herbage and on the underlying soil. Applied Soil Ecology, 29, 165–171.
Basto-Estrella, G. S., Rodríguez-Vivas, R. I., Delfín-González, H., Navarro-Alberto, J. A., & Reyes-Novelo, E. (2012). Escarabajos estercoleros (Coleoptera: Scarabaeinae) en ranchos de Yucatán, México. Revista Mexicana de Biodiversidad, 83, 380–386.
Basto-Estrella, G. S., Rodríguez-Vivas, R. I., Delfín-González, H., Navarro-Alberto, J. A., Favila, M. E., & Reyes-Novelo, E. (2016). Dung removal by dung beetles (Coleoptera: Scarabaeidae) and macrocyclic lactone use on cattle ranches of Yucatan, Mexico. Revista de Biología Tropical (International Journal of Tropical Biology), 64, 945–954.
Bertone, M. A., Green, J. T., Washburn, S. P., Poore, M. H., & Watson, D. W. (2006). The contribution of tunneling dung beetles to pasture soil nutrition. Online. Forage and Grazinglands, 4, DOI: 10.1094/FG-2006-0711-02-RS.
Beynon, S. A., Wainwright, W. A., & Christie, M. (2015). The application of an ecosystem services framework to estimate the economic value of dung beetles to the UK cattle industry. Ecological Entomology, 40, 124–135.
Braga, R. F., Korasaki, V., Andresen, E., & Louzada, J. (2013). Dung beetle community and functions along a habitat-disturbance gradient in the Amazon: A rapid assessment of ecological functions associated to biodiversity. Plos One, 8, e57786.
Brown, J., Scholtz, C. H., Janeau, J. L., Grellier, S., & Podwojewski, P. (2010). Dung beetles (Coleoptera: Scarabaeidae) can improve soil hydrological properties. Applied Soil Ecology, 46, 9–16.
Carpaneto, G. M., Mazziota, A., & Ieradi, M. (2010). Use of habitat resources by scarab dung beetles in an African savanna. Environmental Entomology, 39, 1756–1764.
Corwin, R. M. (1997). Economics of gastrointestinal parasitism of cattle. Veterinary Parasitology, 72, 451–460.
Cruz-Rosales, M. (2011). Contribución de los escarabajos estercoleros a la productividad ganadera en Veracruz (Tesis Doctoral). Colegio de Posgraduados, Campus Veracruz, México.
Cruz, R. M., Martínez, M. I., López-Collado, J., Vargas-Mendoza, M., González-Hernández, H., & Platas-Rosado, D. E. (2012). Degradación del estiércol vacuno por escarabajos estercoleros en un pastizal tropical de Veracruz, México. Revista Colombiana de Entomología, 38, 148–155.
Doube, B. M. (1990). A functional classification for analysis of the structure of dung beetle assemblages. Ecological Entomology, 15, 371–383.
Doube, B. M., Giller, P. S., & Moola, F. (1988). Dung burial strategies in some South African coprine and onitine dung beetles (Scarabaeidae: Scarabaeinae). Ecological Entomology, 13, 251–261.
Estrada, A., & Coates-Estrada, R. (1991). Howler monkeys (Alouatta palliata), dung beetles (Scarabaeidae) and seed dispersal: Ecological interactions in the tropical rain forest of Los Tuxtlas, Mexico. Journal of Tropical Ecology, 7, 459–474.
Estrada, A., Halffter, G., Coates-Estrada, R., & Merritt, D. A. (1993). Dung beetles attracted mammalian herbivore (Alouatta palliata) and omnivore (Nasua narica) dung in the tropical rain forest of Los Tuxtlas, Mexico. Journal of Tropical Ecology, 9, 45–54.
Fincher, G. T. (1981). The potential value of dung beetles in pasture ecosystems. Journal of Georgia Entomological Society, 16, 316–333.
Flota-Bañuelos, C., López-Collado, J., Vargas-Mendoza, M., Fajersson, P., González-Hernández, H., & Martínez, M. I. (2012). Efecto de la ivermectina en la dinámica espacio-temporal de escarabajos estercoleros en Veracruz, México. Tropical and Subtropical Agroecosystems, 15, 227–239.
Forbes, T. D. A., & Hodgson, J. (1985). The reaction of grazing sheep and cattle to the presence of dung from the same or the other species. Grass and Forage Science, 40, 177–182.
Halffter, G., & Edmonds, W. D. (1982). The nesting behaviour of dung beetles (Coleoptera: Scarabaeidae): an ecological and evolutive approach. México D.F.: Instituto de Ecología, A.C.
Halffter, G., & Matthews, E. G. (1966). The natural history of dung beetles of the subfamily Scarabaeinae. Folia Entomológica Mexicana, 12-14, 1–312.
Hanski, I., & Cambefort, Y. (1991). Dung beetle ecology. Nueva Jersey: Princeton University Press.
Horgan, F. G. (2001). Burial of bovine dung by coprophagous beetles (Coleoptera: Scarabaeidae) from horse and cow grazing sites in El Salvador. European Journal of Soil Biology, 37, 103–111.
Horgan, F. G., & Fuentes, R. C. (2005). Asymmetrical competition between Neotropical dung beetles and its consequences for assemblage structure. Ecological Entomology, 30, 182–193.
Huerta, C., Anduaga, S., López-Portillo, J., & Halffter, G. (2010). Use of food and space by tunneler dung beetles (Coleoptera; Scarabaeinae) during reproduction. Environmental Entomology, 39, 1165–1169
Huerta, C., Martínez, M. I., Montes de Oca, E., Cruz-Rosales, M., & Favila, M. E. (2013). The role of dung beetles in the sustainability of pasture and grasslands. In A. Yañez-Arancibia, R. Dávalos-Sotelo, J. W. Day, & E. Reyes (Eds.), Ecological dimensions for sustainable socio economic development (pp. 441–463). Boston: WITPRESS Southampton.
Jankielsohn, A., Scholtz, C. H., & Louw, S. V. D. M. (2001). Effect of habitat transformation on dung beetle assemblages: A comparison between a South African nature reserve and neighboring farms. Environmental Entomology, 30, 474–483.
Kryger, U. (2009). Conservation of dung beetles, Section E. In C. H. Scholtz, A. L. V. Davis, & U. Kryger. (Eds), Evolutionary biology and conservation of dung beetles (pp. 289–412). Sofia-Moscow: Pensoft Publishers.
Lee, C. M., & Wall, R. (2006). Cow-dung colonization and decomposition following insect exclusion. Bulletin of Entomological Research, 96, 315–322.
Lizardo, V., & Castellanos-Vargas, I. (2016). Dung beetle community response to vegetation type and season in an arid zone of the Mexican Plateau. Southwestern Entomologist, 41, 441–452.
López-Collado, J., Cruz-Rosales, M., Vilaboa-Arroniz, J., Martínez, M. I., & González- Hernández, H. (2017). Contribution of dung beetles to cattle productivity in the tropics: A stochastic-dynamic modeling approach. Agricultural Systems, 155,
78–87.
Losey, J. E., & Vaughan, M. (2006). The economic value of ecological services provided by Insects. Bioscience, 56, 311–323.
Lumaret, J. P., Kadiri, N., & Bertrand, M. (1992). Changes in resources: consequences for the dynamics of dung beetle communities. The Journal of Applied Ecology, 29, 349–356.
Menegaz, F. P., & Medina-Hernández, M. I. (2017). Dung beetles associated with agroecosystems of southern Brazil: relationship with soil properties. Revista Brasileira de Ciência do Solo, 41, e0160248.
Nervo, B., Tocco, C., Caprio, E., Palestrini, C., & Rolando, A. (2014). The effects of body mass on dung removal efficiency in dung beetles. Plos One, 9, e107699.
Nichols, E., Spector, S., Louzada, J., Larsen, T., Amézquita, S., & Favila, M. E. (2008). Ecological functions and ecosystem services provided by Scarabaeinae dung beetles. Biological Conservation, 141, 1461–1474.
Omaliko, C. P. E. (1981). Dung deposition, breakdown and grazing behavior of beef cattle at two seasons in a tropical grassland ecosystem. Journal of Range Management, 34, 360–362.
Ortega-Martínez, I. J., Moreno, C. E., & Escobar, F. (2016). A dirty job: manure removal by dung beetles in both a cattle ranch and laboratory setting. Entomologia Experimentalis et Applicata, 161, 70–78.
Ortega-Martínez, J., Moreno, C. E., Sánchez-Rojas, G., & Barragán, F. (2014). The role of Canthon humectus hidalgoensis (Bates) (Coleoptera: Scarabaeidae) in dung removal from a cattle pasture. The Coleopterists Bulletin, 68, 656–658.
Penttilä, A., Slade, E. M., Simojoki, A., Riutta, T., Minkkinen, K., & Roslin, T. (2013). Quantifying beetle-mediated effects on gas fluxes from dung pats. Plos One, 8, e71454.
Pinheiro-Machado, L. C. (2016). Pastoreo racional Voisin. Tecnología Agroecológica para el Tercer Milenio. Buenos Aires: Editorial Hemisferio Sur.
Scholtz, C. H., Davis, A. L. V., & Kryger, U. (2009). Evolutionary biology and conservation of dung beetles. Sofia-Moscow: Pensoft Publishers.
Shahabuddin, O. (2008). The role of coprophagous beetles on dung decomposition and enhacement soil fertility: effect of body size, species diversity and biomass. Journal Agroland, 15, 51–57.
Shepherd, V. E., & Chapman, C. A. (1998). Dung beetles as secondary seed dispersers: impact on seed predation and germination. Journal of Tropical Ecology, 14, 199–215.
Slade, E. M., Mann, D. J., Villanueva, J. F., & Lewis, O. T. (2007). Experimental evidence for the effects of dung beetle functional group richness and composition on ecosystem function in a tropical forest. Journal of Animal Ecology, 76, 1094–1104.
Slade, E. M., Riutta, T., Roslin, T., & Tuomisto, L. (2016). The role of dung beetles in reducing greenhouse gas emissions from cattle farming. Scientific Reports, 6, 18140.
Valdez-Zertuche, J. I., Badii, M. H., Guille, A., & Acuña-Zepeda, M. S. (2015). Causas e impactos socio-económico y ambientales de la erosión. Daena: International Journal of Good Conscience, 10, 76–87.
Vieira, L., Louzada, J. N. C., & Spector, S. (2008). Effects of degradation and replacement of Southern Brazilian coastal sandy vegetation on the dung beetles (Coleoptera: Scarabaeidae). Biotropica, 40, 719–727.
Whipple, S. D., & Hoback, W. W. (2012). A comparison of dung beetle (Coleoptera: Scarabaeidae) attraction to native and exotic mammal dung. Environmental Entomology, 41, 238–244.
Williams, D. R., Alvarado, F., Green, R. E., Manica, A., Phalan, B., & Balmford, A. (2017). Land-use strategies to balance livestock production, biodiversity conservation and carbon storage in Yucatán, Mexico. Global Change Biology, 23, 5260–5272.
Woodward, G., Ebenman, B., Emmerson, M., Montoya, J. M., Olesen, J. M., Valido, A. et al. (2005). Body size in ecological networks. Trends in Ecology and Evolution, 20, 402–409.
Yokoyama, K., Kai, H., & Tsuchiyama, H. (1991). Paracoprid dung beetles and gaseous loss of nitrogen from cow dung. Soil Biology and Biochemistry, 23, 643–647.
Zunino, M. (1991). Food relocation behavior: a multivalent strategy of Coleoptera. In M. Zunino, X. Bellés, & M. Blas (Eds.), Advances in Coleopterology (pp. 297–314). Barcelona: Asociación Europea de Coleopterología.


