Ecología alimentaria del tepezcuintle (Cuniculus paca) en áreas conservadas y transformadas de la Selva Lacandona, Chiapas, México
Yuriana Martínez-Ceceñas a, *, Eduardo J. Naranjoa, Yann Hénaut b y Arturo Carrillo-Reyes c
a El Colegio de la Frontera Sur, Carretera Panamericana y Periférico Sur s/n, 29290 San Cristóbal de Las Casas, Chiapas, México
b El Colegio de la Frontera Sur, Avenida Centenario km 5.5, 424, 77014 Chetumal, Quintana Roo, México
c Universidad de Ciencias y Artes de Chiapas, Libramiento Norte Poniente 47, Caleras Maciel, 29000 Tuxtla Gutiérrez, Chiapas, México
*Autor para correspondencia: limbizkit_07_yr@hotmail.com (Y. Martínez-Ceceñas)
Resumen
Conocer el efecto de la fragmentación del hábitat y las perturbaciones antrópicas es primordial para comprender los procesos de adaptación de las especies y su persistencia en los ecosistemas. Una especie adaptable a ambientes transformados es el tepezcuintle, Cuniculus paca. En este trabajo se evaluó la actividad de forrajeo, la composición y las variaciones en la dieta del tepezcuintle en 2 sitios: uno conservado y otro transformado en la Selva Lacandona, Chiapas, México. Se caracterizaron y monitorearon por fototrampeo 57 sitios de alimentación (“comederos”), en 31 de los cuales se confirmó el consumo de frutos. Comparando los sitios se encontraron diferencias significativas en el estado y cantidad de frutos y la cobertura del dosel. La dieta del tepezcuintle incluyó frutos de 20 especies de árboles, donde Ceiba pentandra y Castilla elastica fueron nuevos registros para la especie. Se encontró una semejanza del 40% en la composición de especies arbóreas consumidas entre sitios, por lo que el tepezcuintle diversifica su dieta aprovechando frutos de especies cultivadas.
Palabras clave:
Dieta; Frugívoro; Fragmentación; Tepezcuintle
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Foraging ecology of lowland paca (Cuniculus paca) in preserved and transformed areas of the Lacandon rainforest, Chiapas, Mexico
Abstract
Recognizing the effect of habitat fragmentation and anthropic disturbances is crucial to understand the processes of species adaptation and their persistence in ecosystems. One species adaptable to transformed environments is the paca, Cuniculus paca. We assessed the foraging activity, composition and variations in the paca diet in pristine and transformed sites in the Lacandon Forest, Chiapas, Mexico. Fifty-seven feeding sites (“feeders”) were characterized and monitored through camera-trapping. Fruit consumption by pacas was confirmed in 31 of those sites. A significant difference in fruit condition and quantity and canopy cover, were found when comparing sites. The paca diet included fruits of 20 tree species, where Ceiba pentandra and Castilla elastica were new records for the species. A 40% similarity was found in the composition of tree species consumed between sites, so that the paca diversifies its diet by taking advantage of cultivated species fruits.
Keywords:
Diet; Frugivorous; Fragmentation; Lowland paca
Introducción
La deforestación, la fragmentación y en general los cambios en la vegetación a causa de las actividades humanas han influido en los procesos ecológicos y en las interacciones entre especies de flora y fauna silvestres como la competencia, depredación, herbívoría y frugivoría (Rodríguez-Rojas, 2005). Aunado a ello, el cambio de uso del suelo para la producción agropecuaria y la caza indiscriminada de especies han producido cambios importantes en las poblaciones de mamíferos y otras especies de fauna silvestre (Camargo-Sanabria y Mendoza, 2016; Naranjo, 2000).
Una de las especies que se ha mencionado que puede adaptarse a los ambientes transformados es el tepezcuintle, Cuniculus paca. Este roedor juega un papel importante en la dinámica de los bosques tropicales, principalmente como consumidor y dispersor de semillas (Bonilla-Morales et al., 2013; Cuarón, 1985). Por ello, si la abundancia poblacional de este roedor disminuyera, probablemente también disminuya la diversidad y supervivencia de muchas especies vegetales en el neotrópico (Cuarón, 1985; Dirzo y Miranda, 1991). Además, el tepezcuintle es un eslabón importante en la cadena alimenticia al formar parte de la dieta de varios carnívoros (Eisenberg y Redford, 1999; Rodríguez-Rojas, 2005).
El tepezcuintle es un consumidor oportunista y su dieta cambia a lo largo de su área de distribución y de acuerdo a la variación estacional y producción de frutos (Dubost y Henry, 2006; Gallina, 1981). Debido a que estos animales no necesariamente consumen frutos en proporción a su disponibilidad en el bosque, es difícil establecer una relación directa entre la disponibilidad y el uso de recursos alimentarios. El conocimiento de la dieta y hábitos alimenticios del tepezcuintle se ha logrado en gran parte por observaciones realizadas en cautiverio (Beck-King et al., 1999; Smythe, 1997). Sin embargo, en los estudios realizados en vida silvestre se han logrado registrar 75 especies de frutos correspondientes a 63 familias que conforman la dieta del tepezcuintle (Beck-King et al., 1999; Figueroa-de León et al., 2017; Gallina, 1981; Muñoz et al., 2002; Zucaratto et al., 2010). En estos estudios, algunas de las especies que se han reportado con mayor frecuencia en la dieta del tepezcuintle son el jobo Spondias mombin, el sonzapote Lycania platypus, aguacate Persea americana, plátano Musa paradisiaca y cacao Theobroma cacao.
Para este roedor, al igual que para muchos mamíferos tropicales, todavía no es posible responder sobre algunos aspectos de su historia natural (i.e., hábitos de alimentación, conducta, cuidado parental), ya que por lo general los estudios realizados en campo resultan parcializados e incompletos debido a que es un animal elusivo de hábitos nocturnos (Beck-King et al., 1999). Los objetivos de esta investigación son: 1) describir los hábitos de alimentación del tepezcuintle; 2) estimar la riqueza de especies vegetales que componen su dieta, y 3) comparar la composición de la dieta y la frecuencia de consumo del tepezcuintle en un sitio conservado y un sitio perturbado en la Selva Lacandona.
Materiales y métodos
El área de estudio se ubicó en la sección sureste de la Reserva de la Biosfera Montes Azules (REBIMA) y en el ejido aledaño Reforma Agraria, en la Selva Lacandona, Chiapas, México (fig. 1). En el ejido se desarrollan el ecoturismo, en una reserva comunitaria de 1,463 ha de selva alta y mediana perennifolia; la agricultura, con 172 ha de cultivos anuales entre los que destacan el maíz y el frijol; y la ganadería, con 768 ha de potreros (García y González, 2015). El ejido se ubica dentro de la cuenca del río Lacantún, el cual representa una barrera natural que lo divide de la REBIMA; lo surcan de oeste a este 2 arroyos principales: el Venado hacia el norte y el Remolino hacia el sur del ejido (García y González, 2015). La REBIMA cuenta con una superficie de 331,200 ha que corresponde al 34.6% de la superficie de la Selva Lacandona (Semarnat, 2000). Tiene un gradiente altitudinal que va desde los 200 m en el río Lacantún hasta los 1,500 m en la región norte de la reserva. En cuanto a vertebrados, se han registrado 112 especies de peces, 54 de reptiles, 23 de anfibios, 341 aves y 112 de mamíferos (Semarnat, 2000). En la REBIMA se encuentran representados todos los órdenes de mamíferos terrestres y 24.8% del total de mamíferos de México. De las 112 especies presentes, 17 son endémicas y algunas otras como el jaguar Panthera onca, ocelote Leopardus pardalis, el tapir Tapirus bairdii, el pecarí de collar Pecari tajacu y el pecarí de labios blancos Tayassu pecari se encuentran dentro de alguna categoría de protección (Semarnat, 2010).
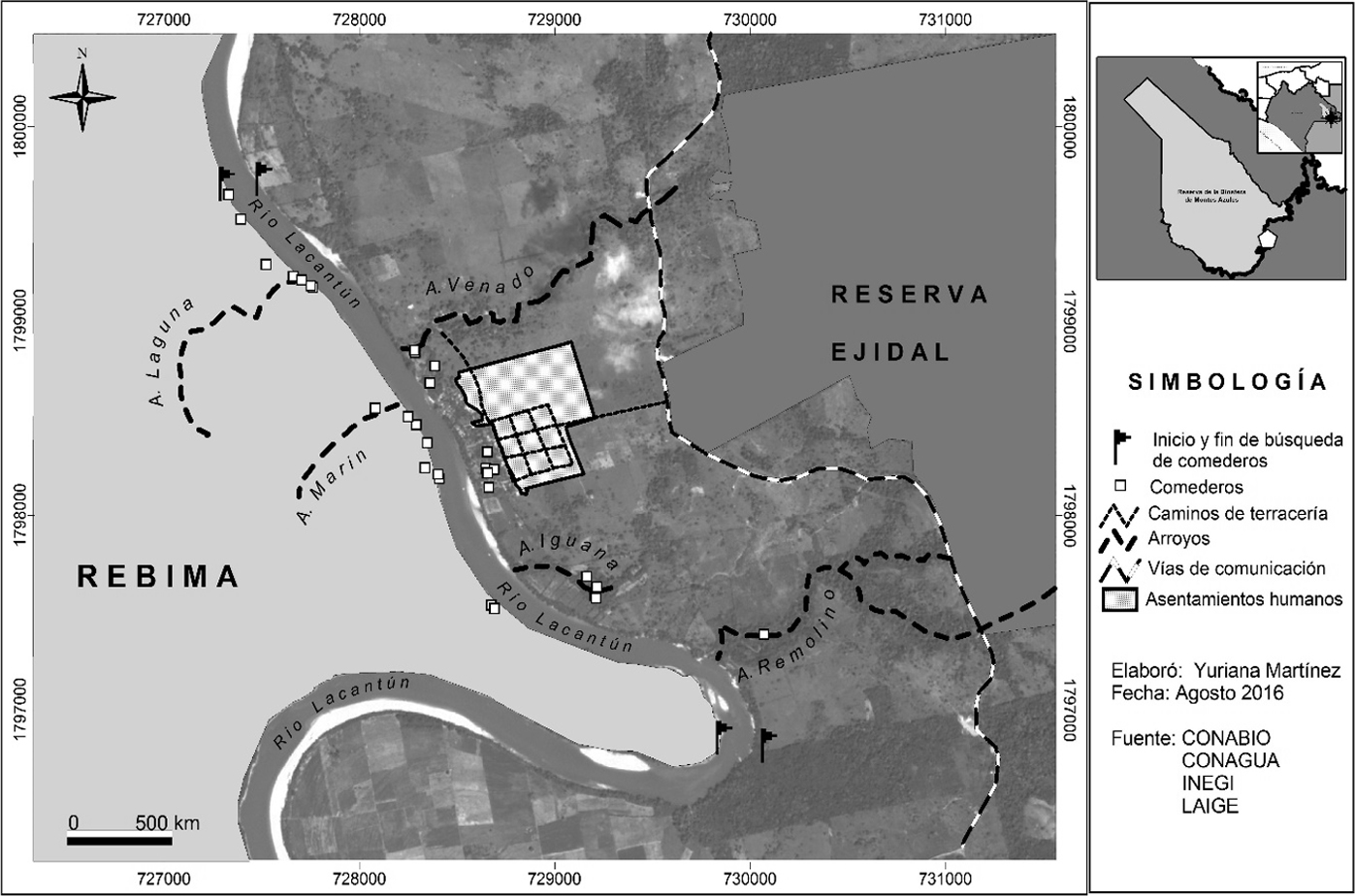
Entre enero y junio de 2016 se llevaron a cabo 6 visitas al área de estudio con intervalos de alrededor de 28 días. En la REBIMA se realizaron solamente 5 visitas debido a las condiciones ambientales adversas como la creciente del río Lacantún y limitaciones de transporte para el acceso a la misma. En cada área se estableció una franja de ancho fijo de 50 × 3,000 m a lo largo del río Lacantún, lo que corresponde a la longitud territorial aproximada del ejido Reforma Agraria.
En febrero, abril y junio de 2016 se realizó una búsqueda intensiva de comederos potencialmente utilizables por los tepezcuintles a lo largo de la franja previamente establecida, con la finalidad de tomar en cuenta aquellas especies vegetales con distinta fenología. Para fines de este estudio se consideró como “comedero” a aquel sitio donde se encontraban frutos transportados, amontonados y que presentaban mordidas recientes del tepezcuintle (marcas de incisivos ≥ 4 mm), así como el área bajo la copa de los árboles cuyos frutos pudieran ser consumidos por el tepezcuintle, apoyándose en comentarios de la gente local. Se monitorearon un total de 57 comederos potencialmente utilizables por el tepezcuintle, 23 en Reforma Agraria y 34 en la REBIMA, correspondientes a 24 especies distintas de árboles (12 en Reforma Agraria y 19 en la REBIMA). Así mismo, se encontraron frutos “solitarios” que presentaban las marcas de incisivos ≥ 4 mm, los cuales no se consideraron como comederos y por tanto no se monitorearon con fototrampeo, pero se registraron para estimar la riqueza de especies que conforman la dieta del tepezcuintle. A este tipo de registros se les denominó “frutos solitarios con marcas de incisivos” (FSMI).
Una vez localizados los comederos, se caracterizaron a través de atributos como: tipo de comedero (bajo enredaderas, bajo raíces de árboles en pie, bajo el árbol que tira el fruto y bajo construcciones humanas), especie, estado de maduración y cantidad de frutos, cobertura del dosel, distancias al árbol que está fructificando, al agua y a actividades humanas (caminos, potreros, asentamientos humanos; tabla 1). En cada comedero se establecieron de 1-2 cámaras trampa Cuddeback Black Flash Modelo E3, que se programaron para estar activas las 24 h del día capturando videos de 30 seg de duración con intervalos de 15 seg entre capturas. Estas cámaras se revisaron cada 28 días aproximadamente y su permanencia en el comedero dependió de la actividad de los tepezcuintles en el mismo, dejando las cámaras activas mientras el tepezcuintle visitara el comedero. Las especies arbóreas que resultaron ser compartidas en ambos sitios (mamey Pouteria sapota, ramón Brosimum alicastrum y corozo Attalea butyracea), se tomaron en cuenta para describir los hábitos alimentarios del tepezcuintle.
A partir de los datos obtenidos de las especies que consume el tepezcuintle (tabla 1) y tomando en cuenta los FSMI, se estimó la riqueza de la dieta para cada sitio de estudio durante todo el periodo de muestreo. Posteriormente se obtuvo el coeficiente de similitud de Sorensen (Magurran, 1988), para determinar el grado de semejanza entre las comunidades de árboles cuyos frutos se encontraron en ambos sitios de muestreo. También se realizó la prueba de Suma de rangos de Wilcoxon con corrección de continuidad (Crowley, 2005) para comparar las frecuencias de consumo de cada especie en cada comedero, con fototrampeo y los FSMI, para cada sitio y mes. Para aquellas especies compartidas entre sitios y registradas mediante fototrampeo, se estimó la frecuencia de consumo para cada especie dividiendo el número de videos efectivos de una especie entre el número de noches cámara. Donde las noches cámara se consideraron como la sumatoria de días que permaneció activa la cámara en cada comedero de cada especie y un video efectivo fue aquel donde se registró el consumo del fruto de la especie vegetal por parte del tepezcuintle. Los análisis para determinar la dieta y frecuencia de consumo de frutos por parte del tepezcuintle se llevaron a cabo utilizando la plataforma R Studio (R Studio Team, 2016).
Tabla 1
Atributos estimados en comederos de Cuniculus paca el ejido Reforma Agraria y la Reserva de la Biosfera de Montes Azules en el periodo enero-junio 2016.
|
Variables |
Unidades |
Descripción |
|
Especie registrada |
– |
Corresponde a la especie vegetal del fruto que se encuentra en el comedero |
|
Estado de fruto |
Categorías 1. Inmaduro 2. Maduro 3. Podrido |
1) Inmaduro, cuando el fruto aún no está en su punto óptimo para ser consumido. |
|
Cantidad de fruto |
Categorías 1.Escaso 2.Medio 3.Abundante |
Se registró al colocar la cámara-trampa en el comedero: 1) Escaso, cuando apenas se encuentra un par de frutos en el sitio y han sido manipulados por el tepezcuintle; |
|
Características del comedero |
Categorías |
|
|
Tipo de comedero |
1. Bajo enredaderas 2. Bajo raíces de árboles en pie 3. Bajo construcciones humanas 3. Bajo construcciones humanas |
1). Bajo enredaderas, los cuales estaban protegidos con lianas y bejucos y eran de difícil acceso incluso para colocar la cámara-trampa; 2) Bajo raíces de árboles en pie, las raíces son numerosas y le brindan protección a este roedor permitiéndole comer entre estas estructuras; 3) Bajo construcciones humanas, el comedero fue establecido bajo estructuras de edificaciones humanas, por ejemplo bajo estufas de leña; y 4) Bajo el árbol que tira frutos, este tipo de comederos se refiere a el área bajo la copa del árbol y cuyos frutos son consumidos en el mismo sitio donde caen. |
|
Cobertura del dosel |
% de cobertura |
Se tomaron cuatro medidas a partir del centro del comedero a 3m hacia los cuatro extremos del mismo utilizando un densitómetro. De dichas medidas se sacó un promedio y se multiplicó por 1.04. El resultado se le restó a 100 para sacar un porcentaje de cobertura (Byrne y Chamberlain, 2012). |
|
Distancia al árbol tirando |
m |
Se estimó la distancia en línea recta al árbol que estaba tirando el fruto |
|
Distancia al agua |
m |
Se estimó la distancia perpendicular al cuerpo de agua más cercano al comedero, |
|
Distancia a actividades humanas |
m |
Se estimó la distancia perpendicular a actividades humanas (potreros, parcelas, asentamientos humanos o caminos). |
Resultados
De los 57 comederos monitoreados, en 16 se registró el consumo de frutos por tepezcuintles en el ejido Reforma Agraria de 11 especies arbóreas. En la REBIMA se registró el consumo de frutos en 15 comederos de 7 especies (tabla 2). El tipo de comedero que más se registró fue el denominado “bajo el árbol que tira el fruto”, donde se encontraron frutos en su mayoría en estado maduro. La especie que presentó mayor número de comederos fue el mamey (n = 11), seguido del amate, ramón y sonzapote con 6, 4 y 4 comederos, respectivamente. Al realizar las comparaciones entre el sitio conservado y perturbado, solo se encontraron diferencias significativas en las variables estado del fruto (X2 = 3.84; p = 0.049), la cantidad de fruto presentes en el comedero (X2 = 8.69; p = 0.012) y cobertura del dosel (W = 43.5; p = 0.0026).
Tabla 2
Comederos monitoreados donde se registró el consumo de frutos por parte del tepezcuintle (Cuniculus paca) en el periodo enero-junio en la Selva Lacandona, Chiapas.
|
Familia |
Nombre común |
Nombre científico |
Ejido Reforma Agraria |
REBIMA |
||
|
CE |
CM |
CE |
CM |
|||
|
Anacardiaceae |
Mango |
Mangifera indica L. |
1 |
1 |
0 |
0 |
|
Anonaceae |
Anona Cultivada |
Af. Anonna reticulata L. |
2 |
2 |
0 |
0 |
|
Anona Silvestre |
Annona scleroderma Saff. |
0 |
0 |
1 |
1 |
|
|
Anonillo |
Cymbopetalum penduliflorum (Dunal) Baill. |
0 |
0 |
2 |
3 |
|
|
Arecaceae |
Corozo |
Attalea butyracea (Mutis ex L.f.) Wess.Boer |
1 |
1 |
1 |
2 |
|
Chrysobalanaceae |
Sunzapote |
Licania platypus (Hemsl.) Fritsch |
1 |
1 |
0 |
3 |
|
Fabaceae |
Frijolillo |
Cojoba arborea L. Britton & Rose |
0 |
0 |
2 |
2 |
|
Sapotaceae |
Mamey |
Pouteria sapota (Jacq.) H. E. Moore y Stearn |
3 |
4 |
6 |
7 |
|
Lauraceae |
Aguacate |
Persea americana Mill. |
2 |
2 |
0 |
0 |
|
Malvaceae |
Molinillo |
Quararibea funebris (La Llave) Vischer |
0 |
0 |
1 |
2 |
|
Moraceae |
Amate grande |
Ficus insipida Wild. |
1 |
5 |
0 |
1 |
|
Hule |
Castilla elastica Cerv. |
1 |
1 |
0 |
0 |
|
|
Ramón |
Brosimum alicastrumSw. |
1 |
1 |
2 |
3 |
|
|
Musaceae |
Plátano Majunche |
var.Musa paradisiaca L. |
2 |
2 |
0 |
0 |
|
Plátano morado |
var. Musa paradisiaca L. |
1 |
1 |
0 |
0 |
|
|
Total |
16 |
21 |
15 |
24 |
CE = comederos efectivos; CM = comederos monitoreados
La dieta del tepezcuintle registrada en el área de estudio incluyó frutos de 20 especies de árboles (tabla 3), de las cuales solo 6 están presentes en la REBIMA, 9 solo en Reforma Agraria y 5 en ambos sitios. Se encontró que el 40% de las especies arbóreas consumidas por el tepezcuintle son las mismas en ambos sitios. El consumo de frutos de hule y ceiba son nuevos registros para la dieta del tepezcuintle en vida silvestre. De las 20 especies registradas, 4 de ellas (plátano manzanita, M. paradisiaca var. Musa sapientum, ceiba, guatope cuajinicuil, Inga inicuil; calabaza, Cucurbita sp. y jobo verde Spondias radlkoferi) fueron identificadas solo por las marcas de incisivos del tepezcuintle (fig. 2).

La frecuencia de consumo de frutos que se registró mediante los videos obtenidos por fototrampeo, añadiendo los FSMI solo mostraron diferencias significativas entre sitios en febrero y marzo con W = 16.5; p = 0.03 y W = 37.5; p = 0.02, respectivamente (tabla 4). En cuanto a las especies arbóreas presentes en ambos sitios, la frecuencia de consumo fue mayor en los comederos de corozo (0.915 videos/noches cámara) respecto a los de mamey y ramón con 0.637 y 0.175 videos/noches cámara, respectivamente (tabla 5).
El tepezcuintle visitó los comederos principalmente durante la noche aunque ocasionalmente presentó actividad al amanecer y atardecer, su intervalo de actividad en los comederos oscila de 19:00 a 7:00 h. Además, se registró la visita de otras especies a los comederos del tepezcuintle tales como: el tlacuache común (Didelphis sp.), tlacuache cuatro ojos (Philander opossum), guaqueques (Dasyprocta sp.), mapaches (Procyon lotor), pecarí de collar, tapir y armadillos (Dasypus novemcinctus). De estos animales, solo al tlacuache cuatro ojos y al mapache no se les observó consumiendo frutos. Además, se registró el paso de algunos depredadores como el jaguar y el puma.
Tabla 3
Lista de especies arbóreas que conforman la dieta del tepezcuintle (Cuniculus paca) en la Selva Lacandona, Chiapas en el periodo de enero- junio de 2016.
|
Familia |
Nombre común |
Nombre científico |
Tipo de fruto |
PFC |
MFC |
Tipo de registro |
Ejido R. Agraria |
REBIMA |
|
Anacardiaceae |
Mango |
Mangifera indica L. |
Drupa |
C, P |
May, Jun |
F, MI |
1 |
0 |
|
Jobo verde |
Spondias radlkoferi Donn. Sm. |
Drupa |
C,P |
Ene, Feb |
MI |
0 |
1 |
|
|
Anonaceae |
Anona Cultivada |
Af. Anonna reticulata L. |
Sincarpico |
C, P |
Ene, Feb |
F, MI |
1 |
0 |
|
Anona Silvestre |
Annona scleroderma Saff. |
Sincarpico |
C, P |
Ene, Feb |
F, MI |
0 |
1 |
|
|
Anonillo |
Cymbopetalum penduliflorum (Dunal) Baill. |
Foliculos agrupados |
C, P |
Ene, Feb |
F |
0 |
1 |
|
|
Arecaceae |
Corozo |
Attalea rostrata Oerst. |
Nueces |
C,P |
May, Jun, Jul |
F |
1 |
1 |
|
Bombacaceae |
Ceiba |
Ceiba pentandra (L.) Gaertn. |
Capsulas obovoides |
C,P,S |
Feb |
MI |
0 |
1 |
|
Chrysobalanaceae |
Sunzapote |
Licania platypus (Hemsl.) Fritsch |
Drupa monospermica |
C, P,CO* |
Abr |
F, MI |
1 |
1 |
|
Cucurbitaceae |
Calabaza |
Cucurbita sp. |
Baya |
C, P |
May |
MI |
1 |
0 |
|
Fabaceae |
Frijolillo |
Cojoba arborea L. Britton & Rose |
Vaina |
C, P, S |
Ene, Feb |
F, MI |
0 |
1 |
|
Guatope guajinicuil |
Inga inicuil Schltdl. & Cham. ex G. Don |
Vaina |
C,P,S |
May, Jun |
MI |
1 |
0 |
|
|
Sapotaceae |
Mamey |
Pouteria sapota (Jacq.) H. E. Moore y Stearn |
Baya |
C, P, E, CO* |
Ene, Feb |
F, MI |
1 |
1 |
|
Lauraceae |
Aguacate |
Persea americana Mill. |
Drupa |
C,P,S, CO* |
May, Jun |
F, MI |
1 |
0 |
|
Malvaceae |
Molinillo |
Quararibea funebris (La Llave) Vischer |
Nuez |
/ |
Ene, Feb |
F |
0 |
1 |
|
Moraceae |
Amate grande |
Ficus insipida Wild. |
Siconos solitarios |
C,P,S |
Enero |
MI |
1 |
1 |
|
Hule** |
Castilla elastica Cerv. |
Drupas agregadas |
C,P,S |
May, Jun |
F |
0 |
0 |
|
|
Ramón |
Brosimum alicastrum Sw. |
Baya |
C,P,S |
Abr, May |
F, MI |
1 |
1 |
|
|
Musaceae |
Plátano manzanita |
Musa paradisiaca var. Musa sapientum, Lin. |
Baya |
C,P,S |
Enero |
MI |
1 |
0 |
|
Plátano Majunche |
var.Musa paradisiaca L. |
Baya |
C,P,S |
Ene, May, Jun |
F, MI |
1 |
0 |
|
|
Plátano morado |
var. Musa paradisiaca L. |
Baya |
C,P,S |
May, Jun |
F, MI |
1 |
0 |
*Observación personal;**nueva especie reportada para la dieta de C. paca; E = endospermo; P = Pulpa; C = cáscara; S = semilla; CO = cotiledones; F = fototrampeo; MI = marcas de incisivos > 4mm; PFC = parte del fruto consumido; MFC = mes de fructificación a partir de observaciones en campo.
Tabla 4
Frecuencia de consumo por mes para cada sitio obtenida a partir de registros de fototrampeo y FSMI.
|
Fototrampeo |
Fototrampeo y FSMI |
|||||||
|
Variable |
Ejido |
REBIMA |
Prueba estadística |
Ejido |
REBIMA |
Prueba estadística |
||
|
n = 11 |
n = 7 |
Wilcoxon |
n = 14 |
n = 11 |
Wilcoxon |
|||
|
W |
p < 0.05 |
W |
p < 0.05 |
|||||
|
Febrero |
0.36 |
1.14 |
16.5 |
0.0303 |
1.07 |
1.54 |
51.5 |
0.149 |
|
Marzo |
0.27 |
1 |
22.5 |
0.0918 |
0.78 |
1.72 |
37.5 |
0.0234 |
|
Abril |
0.27 |
0.28 |
35.5 |
0.755 |
0.92 |
1.36 |
58.5 |
0.283 |
|
Mayo |
0.54 |
1 |
42.5 |
0.722 |
1.92 |
1.81 |
94 |
0.3431 |
|
Junio |
– |
– |
– |
– |
1.07 |
0.63 |
94.5 |
0.3059 |
|
Total de registros |
2.18 |
3.42 |
29 |
0.3958 |
3.28 |
4.81 |
47 |
0.9997 |
Tabla 5
Frecuencia de consumo de frutos de las especies compartidas por parte del tepezcuintle (Cuniculus paca) en la Selva Lacandona, Chiapas en el periodo enero-junio 2016.
|
Núm. de videos efectivos |
Núm. noches cámara |
Frecuencia de consumo |
|||||
|
Fruto |
Ejido |
REBIMA |
Ejido |
REBIMA |
Ejido |
REBIMA |
Total |
|
Mamey |
328 |
155 |
238 |
520 |
1.378 |
0.298 |
0.637 |
|
Ramón |
19 |
5 |
36 |
101 |
0.527 |
0.049 |
0.175 |
|
Corozo |
72 |
32 |
37 |
81 |
2.054 |
0.395 |
0.915 |
Discusión
En estudios previos con tepezcuintles se ha mencionado que éstos trasladan los alimentos del lugar donde los encuentran a sitios más obscuros, donde la vegetación es densa y baja ya que le ofrece protección ante depredadores (Guzmán-Aguirre, 2008; Muñoz et al., 2002). Sin embargo, en este estudio se registraron comederos con vegetación poco densa a baja altura, pero cercanos a cuerpos de agua perennes que podrían significar una ruta de escape en caso de amenaza (Aquino et al., 2012; Contreras-Díaz y Santos-Moreno, 2009).
En cuanto al estado de maduración y la cantidad de frutos en los comederos, se encontraron diferencias entre la REBIMA y el ejido Reforma Agraria, lo que pudo ser consecuencia de que en el ejido el tepezcuintle aprovechó frutos aún inmaduros ante su escasez y debido a su condición de oportunista (Figueroa-de León et al., 2017). En cambio, la abundancia de árboles frutales en la REBIMA proveería de mayor volumen de alimento al tepezcuintle. También se detectó una diferencia significativa en la cobertura del dosel entre sitios. Los tepezcuintles pueden sobrevivir en pequeños fragmentos de bosque que quedan entre grandes extensiones de áreas agrícolas o en acahuales (Gallina et al., 2012; Smythe, 1991). En estos sitios pueden encontrar alimento para sobrevivir, pero en el área de estudio también se observó que estos roedores pueden desplazarse grandes distancias desde los fragmentos de bosque para buscar alimento, tal como lo sugirieron Zucaratto et al. (2010).
En cuanto a la dieta, en el presente trabajo se registraron 20 especies distintas de frutos consumidos por el tepezcuintle, de las cuales 2 son nuevos registros para este roedor. Se ha mencionado que las poblaciones de tepezcuintle pueden verse favorecidas por la flora inducida y manejada por los humanos al aprovechar frutos de especies como el mango, el aguacate, el maíz, entre otros (Gallina, 1981; Zucaratto et al., 2010). En este estudio se confirmó lo anterior, reportándose el consumo de 8 especies introducidas por el ser humano en el área de estudio.
Respecto a la frecuencia de consumo, se observaron diferencias significativas entre sitios en febrero y marzo, posiblemente como consecuencia de una escasa producción de frutos, por lo que la disponibilidad y distribución del alimento disminuye o se concentra en pequeñas áreas obligando al tepezcuintle a visitar repetidamente estos comederos (Gallina, 1981; Guzmán-Aguirre, 2008). De las especies compartidas entre sitios, el corozo fue consumido con mayor frecuencia por estos roedores, posiblemente porque en el ejido son muy escasos los depredadores naturales, los cuales restringirían la actividad de forrajeo del tepezcuintle (Michalski y Norris, 2011). Otro factor importante a considerar fue la cantidad y estado de maduración de los frutos, ya que algunas especies como el amate, que fructifica todo el año, y el corozo producen gran cantidad de frutos. Además, las semillas del corozo se conservan por mucho tiempo y pueden ser consumidas en periodos de mayor escasez (Alvarado-Hernández, 2010).
El tepezcuintle aprovechó varias partes de los frutos en distintas estaciones del año. En la temporada de fructificación del mamey consumió tanto la cáscara como la pulpa, contrario a lo que citaron Guzmán-Aguirre (2008) y Muñoz et al. (2002), quienes notaron que el tepezcuintle retira la cáscara de los frutos para posteriormente consumirlos. Así mismo, los tepezcuintles almacenaron algunas semillas de mamey, las cuales consumieron cuando la producción de frutos disminuyó. Además, en este estudio se observó que al germinar las semillas y las plántulas aún tenían adheridos los cotiledones, que también fueron aprovechados.
En conclusión, en la Selva Lacandona el tepezcuintle diversifica su dieta aprovechando frutos de especies manejadas, los cuales pueden encontrarse en distintos estados de maduración. Las semillas de algunos de estos frutos pueden ser aprovechadas en la misma temporada o una posterior. Además, estos roedores pueden permanecer en sitios fragmentados o desplazarse a otros para buscar alimento. La información de este estudio podría ayudar a desarrollar un plan de manejo y conservación de la especie in situ. Por ello, con base en los resultados de este estudio, sugerimos que se considere la fenología de las especies vegetales que son consumidas por el tepezcuintle, lo que le permitiría contar con alimento en cualquier época del año. Así mismo, es importante prestar atención a la presencia de competidores y depredadores potenciales que pudieran influir en la actividad de forrajeo de este roedor. Por último, además del comportamiento de forrajeo, en el plan de manejo se deberá considerar información sobre la disponibilidad y uso de hábitat (disponibilidad y características del refugio y alimento; Dubost y Henry, 2006; Figueroa-de León et al., 2016, 2017; Gallina 1981) y el comportamiento reproductivo en vida silvestre (tasa reproductiva, recursos necesarios para el éxito reproductivo de la especie).
Agradecimientos
Gracias al Consejo Nacional de Ciencia y Tecnología por la beca otorgada al primer autor de este manuscrito. A los guías de campo del ejido Reforma Agraria: Benjamín Hernández, David Hernández, Sergio Chan y Celedonio Chan. Al Herbario de El Colegio de la Frontera Sur, principalmente a Miguel Martínez Icó por la identificación de las especies arbóreas. Gracias especialmente a Avril Figueroa de León, Karen Gabriela Martínez Ochoa y Henry Castañeda por su apoyo en diversas formas.
Referencias
Alvarado-Hernández, A. (2010). Caracterización florística de los hábitats utilizados por el tepezcuintle, Cuniculus paca (Linneo, 1766; Rodentia: Cuniculidae) en el piso basal del parque nacional Carara, Costa Rica (Tesis). Escuela de Ingeniería Forestal, Instituto Tecnológico de Costa Rica, Cartago, Costa Rica.
Aquino, R., Meléndez, G., Pezo, E. y Gil, D. (2012). Tipos y formas de ambientes de dormir de majás (Cuniculus paca) en la cuenca alta del río Itaya. Revista Peruana de Biología, 19, 27–34.
Beck-King, H., Helversen, O. y Beck-King, R. (1999). Home range, population density, and food resources of Agouti paca (Rodentia: Agoutidae) in Costa Rica: a study using alternative methods. Biotropica, 31, 675–685.
Bonilla-Morales, M., Rodríguez Pulido, J. y Murillo Pacheco, R. (2013). Biología de la lapa (Cuniculus paca Brisson): una perspectiva para la zoocría. Revista CES Medicina Veterinaria y Zootecnia, 8, 83–96.
Byrne, M. y Chamberlain, M. (2012). Using first-passage time to link behaviour and habitat in foraging paths of a terrestrial predator, the racoon. Animal Behaviour, 84, 593–601.
Camargo-Sanabria, A. y Mendoza, E. (2016). Interactions between terrestrial mammals and the fruits of two neotropical rainforest tree species. Acta Oecologica, 73, 45–52.
Contreras-Díaz, R. y Santos-Moreno, A. (2009). Identificación individual de tepezcuintle (Cuniculus paca) mediante el uso de huellas. Revista Mexicana de Mastozoología, 13, 34–45.
Crowley, M. (2005). Statistics, an introduction using R. Chichester, West Sussex, England: Wiley.
Cuarón, A. D. (1985). Crianza del tepezcuintle Agouti paca y los guaqueques Dasyprocta en Chiapas, México. En Libros INE (Eds.), Memorias Primer Simposium Internacional de Fauna Silvestre (pp. 811–822). México D.F.: Instituto Nacional de Biodiversidad, INBio.
Dirzo, R. y Miranda, A. (1991). Altered patterns of hervibory and diversity in the forest understory: a case study of the possible consequences of contemporary defaunation. En P. Price, T. Lewinsohn, G. Fernandes y W. Benson (Eds.), Plant-animal interactions: evolutionary ecology in tropical and temperate regions (pp. 273–287). New York: Wiley and Sons Publications.
Dubost, G. y Henry, O. (2006). Comparison of diets of the acouchy, agouti and paca, the three largest terrestrial rodents of French Guianan Forests. Journal of Tropical Ecology, 22, 641–651.
Eisenberg, F. y Redford, K. (1999). Mammals of the Neotropics. The Central Neotropics. Ecuador, Perú, Bolivia, Brazil. Chicago: University of Chicago Press.
Figueroa-de León, A., Naranjo, E., Perales, H., Santos-Moreno, A. y Lorenzo, C. (2016). Cavity occupancy by lowland paca (Cuniculus paca) in the Lacandon Rainforest, Chiapas, Mexico. Tropical Conservation Science, 9, 246–263.
Figueroa-de León, A., Naranjo, E., Perales, H., Santos-Moreno, A. y Lorenzo, C. (2017). Abundance, density and habitat use of lowland paca (Cuniculus paca, Rodentia: Cuniculidae) in the Lacandon Rainforest, Chiapas, Mexico. Therya, 8, 199–208.
Gallina, S. (1981). Contribución al conocimiento de los hábitos alimenticios del tepezcuintle (Agouti paca Lin.) en Lacanjá-Chansayab, Chiapas. En P. Reyes-Castillo (Eds.), Estudios ecológicos en el trópico mexicano (pp. 58–67). México D.F.: Instituto de Ecología, A.C.
Gallina, S., Pérez-Torres, J. y Guzmán-Aguirre, C. (2012). Use of the paca Cuniculus paca (Rodentia: Agoutidae) in the Sierra de Tabasco State Park, México. Revista de Biología Tropical, 60, 1345–1355.
García, A. B. y González, O. (2015). Uso y conservación del territorio. En J. Cruz-Morales (Eds.), Una entrada a la selva: ejido Reforma Agraria, Marqués de Comillas, Chiapas (pp. 4–5). México D.F.: Universidad Autónoma Chapingo.
Guzmán-Aguirre, C. (2008). Uso, preferencia de hábitat y aprovechamiento del tepezcuintle, Cuniculus paca (Linneo, 1766) en el Parque Estatal de la Sierra de Tabasco, México (Tesis de maestría). Instituto de Ecología, A.C., Xalapa, Veracruz, México.
Magurran, A. (1988). A variety of diversities. En A. Magurran (Eds.), Ecological diversity and its measurement (pp. 81–100). Princeton, New Jersey: Princeton University Press.
Michalski, F. y Norris, D. (2011). Activity pattern of Cuniculus paca (Rodentia: Cuniculidae) in relation to lunar illumination and other abiotic variables in the southern Brazilian Amazon. Zoología, 28, 701–708.
Muñoz, J., Betancur, O. y Duque, M. (2002). Patrones de hábitat y de actividad nocturna de Agouti paca en el Parque Nacional Natural Utría (Chocó, Colombia). Actualidades Biológicas, 24, 75–85.
Naranjo, E. J. (2000). Estimaciones de abundancia y densidad en poblaciones de fauna silvestre tropical. En E. Cabrera, C. Mercolli y R. Resquin (Eds.), Manejo de fauna silvestre en Amazonia y Latinoamérica. (pp. 37–46). University of Florida.
R Studio Core Team. (2016). A languaje and environment for statistical computing. Boston: R Fundation for Statistical Computing. https://www.R-project.org/
Rodríguez-Rojas, C. (2005). Abundancia relativa de mamíferos en dos tipos de cobertura vegetal en la margen nororiental del santuario de flora y fauna Otún Quimbaya, Risaralda (Tesis). Pontificia Universidad Javeriana, Bogotá, Colombia.
Semarnat (Secretaría del Medio Ambiente y Recursos Naturales). (2000). Programa de manejo Reserva de la Biosfera de Montes Azules México. México D.F.: Instituto Nacional de Ecología, Semarnap.
Semarnat (Secretaría del Medio Ambiente y Recursos Naturales). (2010). Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental – Especies nativas de México de flora y fauna silvestres – Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio – Lista de especies en riesgo. Diario Oficial de la Federación. 30 de diciembre de 2010, Segunda Sección, México.
Smythe, N. (1991). Dasyprocta punctata y Agouti paca (Guatusa, Cherenga, Agouti, Tepeizcuinte, Paca). En D. Janzen (Eds.), Historia natural de Costa Rica (pp. 477–499). San José, Costa Rica: Editorial de la Universidad de Costa Rica.
Smythe, N. (1997). Hacia la domesticación de la paca (Agouti = Cuniculus paca) y perspectivas. En J. Robinson y J. Rabinovich (Eds.), Uso y conservación de la vida silvestre neotropical (pp. 245–260). México D.F.: Fondo de Cultura Económica.
Zucaratto, R., Carrara, R. y Siqueira, B. (2010). Dieta da paca (Cuniculus paca) usando métodos indiretos numa área de cultura agrícola na Floresta Atlántica brasileira. Biotemas, 23, 235–239.


