
Carlos A. Gaitán a, Vivian Roxana González-Castillo a,
Gerber Daniel Guzmán-Flores a, Pablo Andrés González-Xiloj a, Usi’j Tz’ununja’ Bá a, Rony A. García-Anleu b, Manolo José García a, *
a Universidad de San Carlos de Guatemala, Facultad de Ciencias Químicas y Farmacia, Centro de Estudios Conservacionistas, Avenida Reforma 0-63 zona 10, Guatemala ciudad, Guatemala
b Wildlife Conservation Society – Guatemala, Avenida 15 de Marzo, casa Núm. 3, Flores, Petén, Guatemala
*Corresponding author: garcia.manolo@usac.edu.gt (M.J. García)
Received: 10 June 2021; accepted: 29 September 2021
Abstract
In this study we determined the resident status of individual jaguars (Panthera onca) in a 7-year timeframe, in order to provide insight into the life history of this species as an input for abundance estimations and the management of the Maya Forest shared by Guatemala, Mexico, and Belize. We gathered photographic jaguar records from studies conducted in 2 Core Zones of the Maya Biosphere Reserve from 2013 to 2019. Individual jaguars were identified by their unique coat pattern, and were considered as residents when recorded for 3 or more consecutive years. Also, range sharing and interactions were quantified. Both female and male resident individuals were recorded. We suggest the presence of a resident population with a tendency for spatial or temporal range exclusion within individuals from the same sex and a fast male’s turnover, and a demographically mixed group of transients. These circumstances appoint the relevance to determine the resident status of individuals for accurate abundance estimations and population trends, and also in maintaining the integrity of the transboundary continuous forest cover, as connectivity appears to have an important role for maintaining the fluctuating jaguar population dynamics.
Keywords: Camera-trap; Maya Biosphere Reserve; Natural history; Population ecology
© 2022 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Jaguares (Panthera onca) residentes en el corazón de la selva Maya en Guatemala
Resumen
En este estudio se determinó el estatus de residencia para individuos de jaguar (Panthera onca) registrados en 7 años, con el fin de brindar aproximaciones a la historia natural de esta especie, como un insumo para el manejo de la selva Maya compartida por Guatemala, México y Belice. Se compilaron registros fotográficos de jaguar de estudios implementados en 2 zonas núcleo de la Reserva de la Biosfera Maya del 2013 al 2019. Los individuos se identificaron por el patrón único en su pelaje y fueron considerados como residentes cuando se registraron por 3 o más años consecutivos. Se cuantificó el uso compartido de territorios e interacciones entre individuos. Los datos sugieren la presencia de una población de jaguares residentes con una tendencia a la exclusión espacial o temporal de territorios entre individuos del mismo sexo y una tasa rápida de recambio para los machos, y un grupo demográfico mixto de transeúntes. Estas circunstancias son relevantes al determinar el estatus de residencia en estimaciones de abundancia y tendencias poblacionales más precisas, y también para mantener la integridad de la cobertura forestal continua transfronteriza, ya que desempeña un papel importante en esta dinámica poblacional de jaguares.
Palabras clave: Cámaras-trampa; Reserva de la Biosfera Maya; Historia natural; Ecología de poblaciones
© 2022 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
Land use patterns in Mesoamerica had delimited transboundary large patches of continuous tropical forest, which are considered current strongholds for populations of endangered large and medium-sized mammal species with accelerated population declines, such as jaguar Panthera onca, White-lipped Peccary Tayassu pecari and Baird’s Tapir Tapirella bairdii (Jędrzejewski et al., 2018; Quigley et al., 2017; Schank et al., 2017; Thornton et al., 2020). The Maya Forest, or ‘’Selva Maya’’ extends through southeast Mexico, northern Guatemala and northwestern Belize, with an approximate extension of 2.1 million ha. It is considered as 1 of the most important Neotropical strongholds for biodiversity conservation, where core protected areas and multiple-use zones, including community-based forestry concessions, have played an important role in the maintenance of the forest cover (Bray et al., 2008). In Guatemala, the Maya Biosphere Reserve (MBR), with 11,600 km2 in area, contains the largest portion of this biological hotspot in the country and thus is relevant for the conservation of this regional stronghold.
Due to its ecological and cultural significance, the jaguar was selected as an indicator species to measure the conservation and management effectiveness of the MBR. Therefore, it is fundamental to collect field data through surveys and monitoring programs to periodically assess the conservation status of this species in the reserve. There have been several single-season surveys to estimate abundance and density for jaguars in the MBR, which have provided the data to assess the conservation status for the species in the reserve and some of its management units (Tobler & Powell, 2013). In order to have a better interpretation of abundance estimates, as well as assessing population viability, it is necessary to monitor life history parameters, as they provide information on individuals and population structure (Harmsen et al., 2017).
Resident jaguars have been identified with the use of camera-traps data on the basis of distinctive coat patterns of each individual providing data on life history parameters (Gaitán et al., 2021; García-Anleu et al., 2015; Harmsen et al., 2009, 2017; Olson, Matzinger et al., 2019; Olson, Saborío et al., 2019). Some of these studies had highlighted the importance of border areas for the conservation of this species through the movements of individuals between populations in different countries (García-Anleu et al., 2020; McCain & Childs, 2008; Mena et al., 2020). Life history parameters for jaguars have been estimated from the northern distribution range in Mexico and northern Central America in Belize (Gutiérrez-González et al., 2015; Harmsen et al., 2017). As a relevant conservation area for jaguars, is necessary to increase the knowledge regarding these parameters at the MBR. For this study, we gathered available jaguar records from data collected by CECON-USAC and partners from 2013 to 2019 with camera-traps deployed at Core Zones of the reserve, identifying individuals, in order to provide insights of life parameters from resident jaguars from an area with minimum human influence, but also as an input for future estimates and conservation assessments at the MBR and the Maya Forest.
Materials and methods
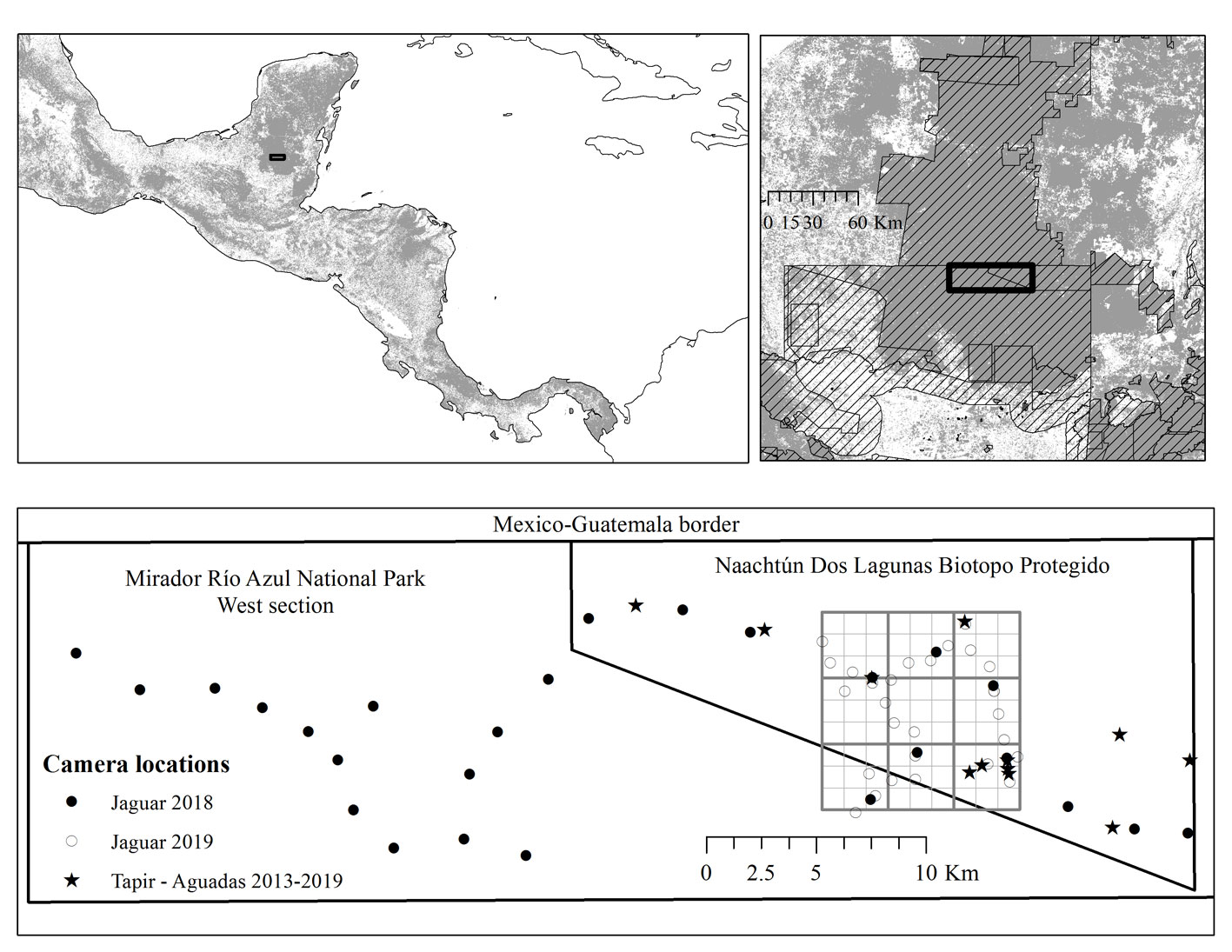
This study was conducted in the Naachtún Dos Lagunas Protected Biotopo (NDLPB; ~ 307 km2), and the west side of the Mirador Río Azul National Park (MRANP; ~ 585 km2), 2 neighbor Core Zones of the MBR, denominated on their joint Master Plan, as the ‘’heart of the Maya Forest’’ due to their location within 1 of the largest forest patches, so it is a well preserved environment with minimal impacts from anthropogenic activities (Fig. 1). Although these areas show high ecological integrity, there is low availability of superficial freshwater during the dry season, which is only available in isolated intermittent water ponds, locally known as ‘’aguadas’’. As main freshwater sources during the dry season, ‘’aguadas’’ are relevant landscape features that influence the temporal and spatial use of jaguars and prey species (Gaitán et al., 2021; Sánchez-Pinzón et al., 2020).
We compiled jaguar photographic records from studies implemented by the San Carlos University and partners in the study area. Our compilation included camera-trap data from 2013 to 2019, collected following 4 different study designs varying in the number of stations, location, camera-trap model, and days activated (Table 1). The study designs were: 1) vertebrate visits to ‘’aguadas’’ during the dry season (March-May) and rainy season (May-September) 2013-2019 following González (2015); 2) Baird’s tapir monitoring, using a 36 km2 grid, some of these locations are the monitored ‘’aguadas’’ adding a camera-trap to the site (García et al., 2019); 3) jaguar survey 2018, with a density of 3 camera-traps per 81 km2, and distance between camera-traps from 1.86 to 4 km; and 4) jaguar survey 2019, with a density of 3 camera-traps per 9 km2 following Chávez et al. (2013) in order to estimate abundance and density. The metadata for each record included the habitat type at each location.
Individuals were considered as residents, when they were detected for 3 or more consecutive years (Harmsen et al., 2017). For jaguar ranging and overlap, we evidenced range exclusivity using location-month as a unit of analysis, which represented pooled data for a single camera-trap location for 1 calendar month, using a dataset including locations with at least 2 individuals as Harmsen et al. (2009). For intraspecific interactions between jaguars we followed Harmsen et al. (2009), as the main unit of analysis was the number of days between consecutive captures per camera-trap location, accounting for ‘’same female’’, ‘’female-female’’, ‘’same male’’ and ‘’male-male’’. Student’s t-tests were used for comparisons with all data log10-transformed.
Table 1
Projects implemented by CECON-USAC and partners in the Naachtún Dos Lagunas Protected Biotopo and Mirador Río Azul National Park from 2013 to 2019.
| Project | Partner institutions | No. camera-trap stations or ‘’aguadas’’ | No. camera-trap per station or ‘’aguada’’ | Year surveyed |
| Vivian González’s B.Sc. Thesis | WCS-Guatemala | 4 | 3 | 2013 |
| Visits of vertebrates to ‘’aguadas’’ monitoring program | – | 2-5 | 1-3 | 2014 – 2019 |
| Baird’s tapir monitoring in the Maya Biosphere Reserve | Fundación Defensores de la Naturaleza | 8 | 1 | 2016 – 2019 |
| Jaguars in the heart of the Maya Forest: phases I and II | CONAP and FUNDAECO | 27 | 1-2 | 2018 and 2019 |
Results
We gathered 1,626 pictures and 148 videos including jaguars. We identified 36 individuals (9 females, 22 males and 5 undetermined) with 26 individuals photographed on both sides, and 10 individuals only on the right side. Additionally, 6 left sides were recorded (2 females, 2 males and 2 undetermined). From the identified individuals, 7 (2 females, 4 males and 1 undetermined) were only recorded on the west side of the MRANP, 3 males were recorded on both areas and the remaining on the NDLPB.
Five resident individuals (2 females and 3 males) were recorded at the NDLPB, being a small proportion of the recorded individuals (Table 2). Resident females were recorded for 4 and 5 consecutive years, and male individuals were recorded for 3 years. For 2019 in a 90-days period, from 137 records with identified individuals, 13 were from residents (6 in ‘’aguadas’’ and 7 in roads) and 123 for non-residents (6 in ‘’aguadas’’ and 117 in roads).

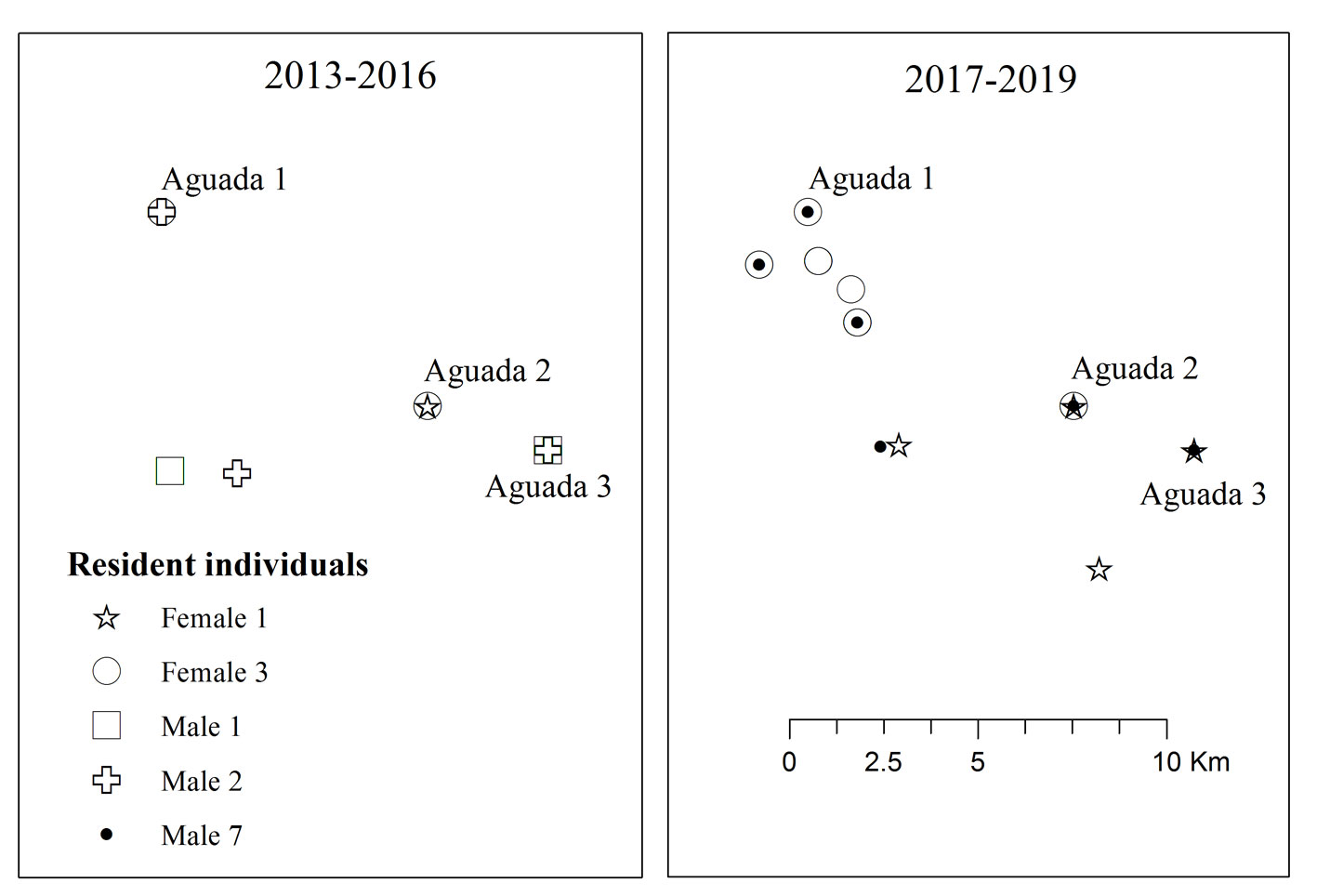
Up to 3 females were recorded in the same location in the same month, but locations were more commonly occupied by a single female (Fig. 2). For males, we recorded up to 4 individuals, with only 9% of location-month units containing a single male individual, showing a tendency of range sharing among conspecific males.
For resident females, we recorded up to 2 individuals (only 1 record) in the same location-month. Resident males were recorded only in single occupancy, showing a tendency of range exclusivity among other resident males and range sharing with resident females (Fig. 2).
Resident females (F1 and F3; Fig. 3) have just 1 location of overlap and they were recorded in the same or near locations over the years. Male jaguars (M1 and M2) had spatial overlap with resident females and within them, and in subsequent years, Male 7 (M7) with resident females (Fig. 3).
We did not find significant differences in the number of consecutive pairs of recaptures of the same individual and different individuals by location per year (X = 2.11 and X = 3.06, respectively; t =1.29, p > 0.05 on Log10-transformed numbers of pairs, 46 locations/year), neither in the time interval between pairs of recaptures (X = 10.13 days from 97 pairs and X = 11.13 days from 141 pairs, t = 0.75, p > 0.05 on Log10-transformed numbers of pairs).
We did not find significant differences in the number of pairs (X = 2.11 and X = 3.06, respectively; t = 0.28, p > 0.05 on Log10-transformed numbers of pairs, 37 locations/year) neither in the time interval between pairs of recaptures for resident-nonresident; and nonresident-nonresident (X = 12.43 days from 53 pairs and X = 10.48 days from 84 pairs, t = -0.05, p > 0.05 on Log10-transformed numbers of pairs).
Discussion
The record of 2 resident females for ≥ 4 consecutive years matches previous reports of resident females at Tikal National Park within the MBR, suggesting that established ranges are maintained for several years (García-Anleu et al., 2015; Gaitán et al., 2022). Considering the low and sporadic detection of females reported by Harmsen et al. (2017), there is a high probability that the resident female (F1) remained in the area for the whole study timeframe, but was not detected in 2014 and 2015, the years with smaller sampling effort (Table 1). Then, F1 would have been at least 9 years-old in 2019 with a stable range for at least 7 years, since she already was an adult when first recorded in 2013. Long-living individuals with stable ranges may be indicative of good quality habitats, which match with our study area (Olson, Saborío et al., 2019).
Table 2
Summary of individuals of Panthera onca recorded > 1 year from 2013 to 2019 in the Naachtún Dos Lagunas Protected Biotopo.
| Jaguar ID | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | Status |
| Female 1 | X | X | X | X | X | Resident | ||
| Male 1 | X | X | X | Resident | ||||
| Male 2 | X | X | X | Resident | ||||
| Female 2 | X | X | X | 2 years | ||||
| Female 3 | X | X | X | X | X | Resident | ||
| Male 3 | X | X | 2 years | |||||
| Male 4 | X | X | 2 years | |||||
| Male 5 | X | X | 2 years | |||||
| Female 4 | X | X | 2 years | |||||
| Male 6 | X | X | 2 years | |||||
| Male 7 | X | X | X | Resident | ||||
| Male 8 | X | X | 2 years | |||||
| Male 9 | X | X | 2 years | |||||
| Male 10 | X | X | 2 years | |||||
| Male 11 | X | X | 2 years | |||||
| Female 5 | X | X | 2 years | |||||
| Female 6 | X | X | 2 years |
The records of resident male jaguars in this study are the first ones for the MBR and Guatemala, evidencing a certain degree of range fidelity for both resident females and males in the Maya Forest. When compared to reports of long-living males in other countries and life history parameters from a population in Belize, male jaguars in the MBR apparently have a fast turnover rate at a specific site, as they were only recorded for 3 consecutive years in our study and there were no recaptures for males by García-Anleu et al. (2015) in a 4-years interval (Harmsen et al., 2017; Olson, Matzinger et al., 2019; Olson, Saborío et al., 2019). Based on reports of transboundary movements and estimated contemporary gene flow of jaguars in the Maya Forest (de la Torre & Rivero, 2019; de la Torre et al., 2017a; García-Anleu et al., 2020; Wultsch et al., 2016), we assume that high mobility is the main source of male individual’s turnover, as the survival probability of males has been reported to be higher than female’s (Harmsen et al., 2017).
The high connectivity and mobility in the Maya Forest would be expected to promote nomadic behaviours for non-resident individuals, such as those individuals that move to another area and return, as this behavior has been recorded for adult males in Guatemala and Belize (García-Anleu et al., 2020; Harmsen et al., 2017). In order to determine local residents, variograms from observed individuals’ track movements would be more reliable than time spent on a specific area. However, due to the lack of this kind of data, we suggest using the ≥ 3 consecutive years parameter for local residents instead of the > 2 years interval employed for large felids, as there have been recorded individuals remaining in a site for > 1 year without establishing a range (Barlow et al., 2009; de Azevedo & Murray, 2007; Morato et al., 2016).
The 3 now-reported resident males’ ranges overlap with the range of the 2 resident females. This matches with previously reported studies, whereas males show larger ranges in order to include several female’s ranges to increase the chance of producing offspring (Cruz et al., 2021; de la Torre & Rivero, 2019; Morato et al., 2016). Therefore, male residents may compete with other resident males for range overlapping with resident females having stable ranges, with the necessary resources to produce viable offspring. Our data shows that M1 and M2 were replaced by M7, while resident females (F1 and F3) remained in the territory. The competition within males can be another driver for the male jaguar’s fast turnover recorded in the study area.
Herein, we suggest the presence of a layered population at local scale, composed of a layer of resident individuals and a fluctuating layer of transients from various demographic groups, as proposed by Harmsen et al. (2017) for the Cockscomb Basin Wildlife Sanctuary (CBWS) in Belize. This complex and dynamic structure of jaguar populations emphasize the importance for monitoring programs to identifying the resident status of individuals, in order to improve accuracy and precision of abundance and density estimates, and thus inferences on population status and long-term viability of populations (Barlow et al., 2009; Gutiérrez-González et al., 2015).
Regarding our design and sampling efforts, the area covered, and considering data from studies from neighbor areas such as Harmsen et al. (2017, 2020), we are cautious in drawing conclusions with respect to our estimates of transient individuals when taking into account the considerations of Tobler and Powell (2013) by having sampling areas large enough according to the biology and ecology of jaguars. Resident individuals with ranges larger than our sampling grid would be recorded as transients.
The conservation implications of considering a fluctuating population of jaguars in the Maya Forest appoints the importance of maintaining the integrity of the continuous forest cover within these 3 joining countries. This circumstance includes avoiding the construction of new roads or any other infrastructure facilitating accessibility in core areas of the Maya Forest in Guatemala, Mexico and Belize, as reduces the potential for sustaining populations of large carnivores (Espinosa et al., 2018; Hernández-Pérez et al., 2020).
The importance of transboundary areas for the conservation of jaguars in the United States and Mexico border, the Maya Forest, and the Amazon, has already been highlighted (de la Torre et al., 2017a, b; García-Anleu et al., 2020; McCain & Childs, 2008; Mena et al., 2020). Consequently, we also consider that transboundary collaboration between research teams from different countries is needed to understand the population dynamics in large habitat areas such as the Maya Forest. The integration of data from different research teams or projects may have the advantage of greater spatial or temporal coverage, but the limitation that this data usually comes from different study designs. A regional homologation of data collection protocols and the use of new techniques that allow the use of data from different sources may be useful tools as they can reduce these limitations (Ngoprasert et al., 2019).
Acknowledgments
This study was funded by Dirección General de Investigación from San Carlos University, projects implemented in 2014, 2016, 2017, 2018 and 2020 (projects 4.8.63.3.57, 4.8.63.4.04, 4.8.63.2.03, 4.8.63.2.35 and B29-2020), Fundaeco-Global Conservation (projects Jaguars in the heart of the Maya Forest: phases I and II) and Fundación Defensores de la Naturaleza-Tapir Specialists Group-Foundation Segré (Baird’s Tapir Guatemala Program-World Tapir Conservation Program and Latin-American Tapir Conservation Program), Consejo Nacional de Ciencia y Tecnología – Secretaría Nacional de Ciencia y Tecnología (Project FODECYT 26-2011) and Wildlife Conservation Society – Guatemala. For their help in the field, park rangers, technicians and students from USAC/CECON, CONAP, IDAEH, and FUNDAECO: Jerson Olivares, Walter García, Elvis Castellanos, Francisco Asturias, Basilio Silvestre, Arturo Palacios, Byron Cruz, Caín Olivares, Carlos Tzul, Darwin Castillo, Elvis Solís, Erwin Mayen, Fredy Bedoya, Isidro Meléndrez, Jaime Gutiérrez, Jaury Morales, José L. Rodas, Juan Najarro, Marcial Hernández, Mario Chun, Marvin García, Marvin Ochaeta, Marvin Tobar, Miguel Hernández, Renan Soto, Saúl Castillo, Víctor Hernández, Luis F. Rodas, Juan J. Romero, Marvin Rosales, Andrea Aguilera, Lourdes Núñez, Gabriela Cajbon, Pablo Lee, Santiago Zetina, Valeria Barrera, and Mynor A. Sandoval. To César E. Fuentes-Montejo as reviewer and Julianna M. Ritson for her help with the grammar structure of this manuscript.
References
Barlow, A. C. D., McDougal, C., Smith, J. L. D., Gurung, B., Bhatta, S. R., Kumal, S. et al. (2009). Temporal variation in tiger (Panthera tigris) populations and its implications for monitoring. Journal of Mammalogy, 90, 472–478. https://doi.org/10.1644/07-MAMM-A-415.1
Bray, D., Duran, E., Ramos, V., May, J., & Velasquez, A. (2008). Tropical deforestation, community forests, and protected areas in the Maya Forest. Ecology and Society, 13, 56. https://doi.org/10.5751/ES-02593-130256
Chávez, C., de la Torre, A., Bárcenas, H., Medellín, R. A., Zarza, H., & Ceballos, G. (2013). Manual de fototrampeo para estudio de fauna silvestre. El jaguar en México como estudio de caso. Alianza WWF-Telcel, Universidad Nacional Autónoma de México. https://biblioteca.semarnat.gob.mx/janium/Documentos/Ciga/Libros2013/CD002776.pdf
Cruz, C., Zarza, H., Vidal-Mateo, J., Urios, V., & Ceballos, G. (2021). Top predator ecology and conservation: Lesson from jaguars in southeastern Mexico. Conservation Science and Practice, 3, e328. https://doi.org/10.1111/csp2.328
de Azevedo, F. C. C., & Murray, D. L. (2007). Spatial organization and food habits of jaguars (Panthera onca) in a floodplain forest. Biological Conservation, 137, 391–402. https://doi.org/10.1016/j.biocon.2007.02.022
de la Torre, J. A., Núñez, J. M., & Medellín, R. A. (2017a). Habitat availability and connectivity for jaguars (Panthera onca) in the southern Mayan Forest: Conservation priorities for a fragmented landscape. Biological Conservation, 206, 270-282. https://doi.org/10.1016/j.biocon.2016.11.034
de la Torre, J. A., Núñez, J. M., &, Medellín, R. A. (2017b). Spatial requirements of jaguars and pumas in southern Mexico. Mammalian Biology, 84, 52–60. https://doi.org/10.1016/j.mambio.2017.01.006
de la Torre, J. A., & Rivero, M. (2019). Insights of the movements of the jaguar in the tropical forests of southern Mexico. In R. Reyna-Hurtado, & C. Chapman (Eds.), Movement ecology of Neotropical forest mammals (pp. 217–241). Springer, Cham. https://doi.org/10.1007/978-3-030-03463-4_13
Espinosa, S., Celis, G., & Branch, L. C. (2018). When roads appear jaguars decline: Increased access to an Amazonian wilderness area reduces potential for jaguar conservation. Plos One, 13, e0189740. https://doi.org/10.1371/journal.pone.0189740
Gaitán, C. A., García, M. J., Sandoval-Lemus, M. A., González-Castillo, V. R., Guzmán-Flores, G. D., & Pineda, C. M. (2022). New queen? Evidence of a long-living Jaguar Panthera onca (Mammalia: Carnivora: Felidae) in Tikal National Park, Guatemala. Journal of Threatened Taxa, 14, 21388–21395. https://doi.org/10.11609/jott.5813.14.7.21388-21395
Gaitán, C. A., González-Castillo, V. R., Guzmán-Flores, G. D., Aguilera, A. L., & García, M. J. (2021). Visitation patterns of jaguars Panthera onca (Carnivora: Felidae) to isolated water ponds in a tropical forest landscape. Therya, 12, 44-55. https://doi.org/10.12933/therya-20-915 ISSN 2007-3364
García, M. J., Leonardo, R. S., González-Castillo, V. R., Guzmán-Flores, G. D., Jurado, N., Sandoval, M. A. et al. (2019). Primera aproximación al uso de la ocupación del tapir (Tapirella bairdii Gill, 1865) como indicador de la integridad ecológica en la Reserva de la Biosfera Maya, Guatemala. Ciencia, Tecnología y Salud, 6, 120–131. http://dx.doi.org/10.36829/63CTS.v6i2.780
García-Anleu, R., Kelly, M. J., Meerman, J., Nipko, R. B., Holcombe, B., McPhail, D. et al. (2020). Short-tail jaguar: the need for transboundary collaboration across the Maya Forest. Cat News, 71, 38-40.
García-Anleu, R., Ponce-Santizo, G., McNab, R. B., Polisar, J., Noss, A., Moreira, J., & Ruano, G. (2015). The queen of Tikal and her suitors. Cat News, 62, 42–43.
González, V. R. (2015). Vertebrados medianos y mayores asociados a las aguadas del biotopo protegido Naachtún – Dos Lagunas, Petén. Universidad de San Carlos de Guatemala, Guatemala. http://biblioteca.usac.edu.gt/tesis/06/06_3844.pdf
Gutiérrez-González, C. E., Gómez-Ramírez, M. A., López-González, C. A., & Doherty, P. F. Jr. (2015). Are private reserves effective for jaguar conservation? Plos One, 10, e0137541. https://doi.org/10.1371/journal.pone.0137541
Harmsen, B. J., Foster, R. J., & Quigley, H. (2020). Spatially explicit capture recapture density estimates: Robustness, accuracy and precision in a long-term study of jaguars (Panthera onca). Plos One, 15, e0227468. https://doi.org/10.1371/journal.pone.0227468
Harmsen, B. J., Foster, R. J., Silver, S. C., Ostro, L. E., & Doncaster, C. P. (2009). Spatial and temporal interactions of sympatric jaguars (Panthera onca) and pumas (Puma concolor) in a Neotropical forest. Journal of Mammalogy, 90, 612-620. https://doi.org/10.1644/08-MAMM-A-140R.1
Harmsen B. J., Foster, R. J., Sánchez, E., Gutiérrez-González, C. E., Silver, S. C., Ostro, L. E. T., Kelly, M. J., Kay, E., & Quigley, H. (2017). Long term monitoring of jaguars in the Cockscomb Basin Wildlife Sanctuary, Belize: Implications for camera trap studies of carnivores. Plos One, 12, e0179505. https://doi.org/10.1371/journal.pone.0179505
Hernández-Pérez, E. L., López, J., Friedeberg, D., Contreras-Moreno, F. M. & Hidalgo-Mihart, M. G. (2020). Jaguars and roads in the Calakmul region, evidence of road crossing and collisions with vehicles. Western North American Naturalist, 80, 556–562. https://scholarsarchive.byu.edu/wnan/vol80/iss4/16
Jędrzejewski, W., Robinson, H. S., Abarca, M., Zeller, K. A., Velasquez, G., Paemelaere, E. et al. (2018). Estimating large carnivore populations at global scale based on spatial predictions of density and distribution – Application to the jaguar (Panthera onca). Plos One, 13, e0194719. https://doi.org/10.1371/journal.pone.0194719
McCain, E., & Childs, J. (2008). Evidence of resident jaguars (Panthera onca) in the Southwestern United States and the implications for conservation. Journal of Mammalogy, 89, 1-10. https://doi.org/10.1644/07-MAMM-F-268.1
Mena, J. L., Yagui, H., Tejeda, V., Cabrera, J., Pacheco-Esquivel, J., Rivero, J., & Pastor, P. (2020). Abundance of jaguars and occupancy of medium- and large-sized vertebrates in a transboundary conservation landscape in the northwestern Amazon. Global Ecology and Conservation, 23, 2020, e01079. https://doi.org/10.1016/j.gecco.2020.e01079
Morato, R. G., Stabach, J. A., Fleming, C. H., Calabrese, J. M., De Paula, R. C., Ferraz K. M. P. M. et al. (2016). Space use and movement of a Neotropical top predator: The endangered Jaguar. Plos One, 11, e0168176. https://doi.org/10.1371/journal.pone.0168176
Ngoprasert, D., Gale, G. A., & Tyre, A. J. (2019). Abundance estimation from multiple data types for group-living animals: An example using dhole (Cuon alpinus). Global Ecology and Conservation, 20, e00792. https://doi.org/10.1016/j.gecco.2019.e00792
Olson, E. R., Matzinger, P. J., Saborío, G., & Carazo-Salazar, J. (2019). Macho uno: a sign of hope for the jaguars of Corcovado National Park, Costa Rica. Cat News, 69, 4-6.
Olson, E. R., Saborío, G., & Carazo-Salazar, J. (2019). Age of the jaguar: A novel approach to evaluating the life-span of a rare carnivore. Cat News, 70, 36-38.
Quigley, H., Foster, R., Petracca, L., Payan, E., Salom, R., & Harmsen, B. (2017). Panthera onca (errata version published in 2018). The IUCN Red List of Threatened Species 2017: e.T15953A123791436. http://dx.doi.org/10.2305/IUCN.UK.2017-3.RLTS.T15953A50658693.en.
Sánchez-Pinzón, K., Reyna-Hurtado, R., Naranjo, E. J., & Keuroghlian, A. (2020). Peccaries and their relationship with water availability and their predators in Calakmul, México. Therya, 11, 213–220. https://doi.org/10.12933/therya-20-878
Schank, C. J., Cove, M. V., Kelly, M. J., Mendoza, E., O’Farrill, G., Reyna-Hurtado, R. et al. (2017). Using a novel model approach to assess the distribution and conservation status of the endangered Baird’s tapir. Diversity and Distributions, 23, 1459–1471. https://doi.org/10.1111/ddi.12631
Thornton, D., Reyna, R., Perera-Romero, L., Radachowsky, J., Hidalgo-Mihart, M. G., García-Anleu, R. et al. (2020). Precipitous decline of white-lipped peccary populations in Mesoamerica. Biological Conservation, 242, 108410. https://doi.org/10.1016/j.biocon.2020.108410
Tobler, M. W., & Powell, G. V. N. (2013). Estimating jaguar densities with camera traps: Problems with current designs and recommendations for future studies. Biological Conservation, 159, 109–118. https://doi.org/10.1016/j.biocon.2012.12.009
Wultsch, C., Caragiulo, A., Dias-Freedman, I., Quigley, H., Rabinowitz, S., & Amato, G. (2016). Genetic diversity and population structure of mesoamerican jaguars (Panthera onca): Implications for conservation and management. Plos One, 11, e0162377. https://doi.org/10.1371/journal.pone.0162377


