
Thomas H. Atkinson *
University of Texas at Austin, Department of Integrative Biology, University of Texas Insect Collection, 3001 Lake Austin Boulevard, Suite 1.314, Austin, Texas, 78703 USA
*Corresponding author: thatkinson.austin@gmail.com (T.H. Atkinson)
Received: 29 July 2022; accepted: 07 December 2022
http://zoobank.org/urn:lsid:zoobank.org:pub:AAE503EC-42C9-4A7E-AE94-17675C2A91B3
Abstract
The genus Phloeocleptus Wood includes 8 species of phloeophagous bark beetles breeding in species of Persea (Lauraceae) and that are restricted to Mexico. Phloeocletptus lorenzfischeri Atkinson is described as new. Phloeocleptus cristatus Wood is a synonym of P. atkinsoni Wood. The male of P. ardis Wood is described. All species are illustrated and redescribed and a new generic key is included. Phloeocleptus punctatus Wood and Hylocurus tresmariae Schedl are transferred to Pseudothysanoes Blackman. Phloeocleptus parvus Wood is transferred to Hylocurus Eichhoff.
Keywords: Phloeophagous; Lauraceae; Persea; Pseudothysanoes
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Revisión del género Phloeocleptus (Coleoptera: Curculionidae: Scolytinae)
Resumen
El género Phloeocleptus Wood incluye 8 especies de descortezadores floeófagos los cuales se reproducen en especies de Persea (Lauraceae), las cuales están restringidos a México. Phloeocleptus lorenzfischeri Atkinson se describe como especie nueva. Phloeocleptus cristatus Wood es sinónimo de P. atkinsoni Wood. Se describe el macho de P. ardis Wood. Todas especies se illustran, se redescriben y se incluye una clave nueva para el género. Phloeocleptus punctatus Wood y Hylocurus tresmariae Schedl se transfieren a Pseudothysanoes Blackman. Phloeocleptus parvus Wood se transfiere a Hylocurus Eichhoff.
Palabras clave: Floeófago; Lauraceae; Persea; Pseudothysanoes
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
As treated here, the genus Phloeocleptus Wood, 1956 consists of 8 species of small phloem-feeding bark beetles in the tribe Micracidini, all of which have only been found in Mexico. All species are known to breed in hosts in the genus Persea (Lauraceae), including domesticated avocados. These species are seldom collected by general collectors or in traps, though they are easy to find in broken or shaded-out branches of their hosts. Several of these species are still known only from the original type series. Since their descriptions additional records have been published by Atkinson and Equihua (1985), Atkinson et al. (1986), Noguera and Atkinson (1990) and Lázaro et al. (2020).
Wood (1982) included 11 species in the genus, including 1 from Costa Rica. The discovery of a new species from Oaxaca and new collections prompted a revision of the known species and a study of the generic boundaries, including revision of all type material. As a result, the synonymy of P. cristatus and P. atkinsoni Wood is proposed. Two of the species included by Wood, Hylocurus tresmariae Schedl and Phloeocleptus punctatus Wood, are transferred to the genus Pseudothysanoes Blackman. Phloeocleptus parvus Wood is transferred to Hylocurus Eichhoff.
Materials and methods
For this study all type material and all specimens from published records were revised. Specimens are deposited in the following collections: CAS California Academy of Sciences, San Francisco, California, USA; CEAM Coleción Entomológica, Centro de Fitosanidad, Colegio de Postgraduados, Montecillo, Estado de México, Mexico; CNIN Colección Nacional de Insectos, Instituto de Biología, Universidad Nacional Autónoma de México, Cd. México, Mexico; NMW Naturhistorische Museum Wien, Vienna, Austria; SEMC Snow Entomological Museum, University of Kansas, Lawrence, Kansas, USA; TAMU Entomology Department, Texas A&M University, College Station, Texas, USA; USNM National Museum of Natural History, Smithsonian Institution, Washington D.C., USA; UTIC University of Texas Insect Collection, University of Texas at Austin, Austin, Texas, USA.
Photographs by the author were taken with a Canon EOS camera with a Canon MP-E 65 mm 1-5X macro lens added to an additional 68 mm of extension tubes mounted on a Stack Shot rail (Cognisys, Inc, Traverse City, MI). Pictures were taken with 3 portable speed flashes and montaged with Zerene Stacker (Zerene Systems, LLC, Richland, WA). The system was described by Wild (2015). Other photos by Sarah Smith were taken with a similar system by Visionary Digital Passport II system (Dun Inc., Palmyra, VA). Montage images were assembled using Helicon Focus Mac Pro 6.7.1 (Helicon Soft, Kharkiv, Ukraine).
All measurements presented here are original, and not taken from literature. Many were taken by the author with an optical micrometer and an Olympus SZ40 steromicroscope. Others were taken directly from photographs taken at known magnifications using Photoshop’s measurement tools (Adobe Inc., San Jose, CA). The length of the pronotum (which completely covers the head in dorsal view) and the length of the elytra (from base to apex) were measured separately and the combined length is the sum of these 2. This is necessary because in many specimens the body is bent or extended between the pronotum and the rest of the body and a single measurement is not accurate. Body width is the maximum width taken either at the base of the pronotum or the base of the elytra, according to the mounting and condition of specimens.
Descriptions
Genus Phloeocleptus Wood 1956
Type species: Phloeocleptus caudatus Wood 1956, original designation
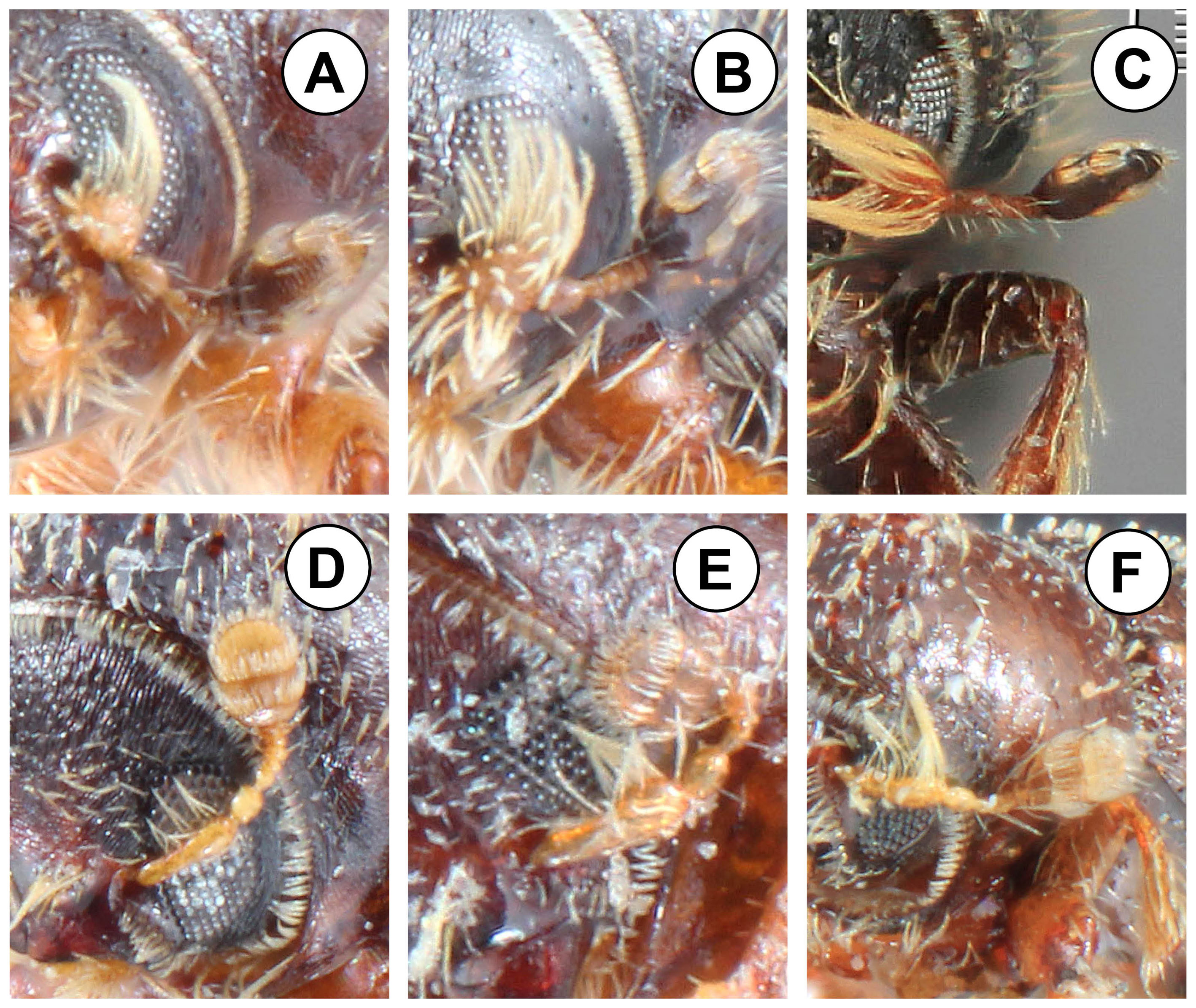
Characters. Wood (1956, 1982, 1986) separated Phloeocleptus from other genera in the Micracidini by the combination of an acuminate apex of the declivity and the arrangement of the sutures on the antennal club. The antennal club is slender and oval, with no externally marked sutures. Two sutures are indicated laterally by tufts of hairs that do not extend across the face of the club (Fig. 1A-C). In the related genus, Pseudothysanoes, the 2 sutures are complete across the face of the club and marked with setae throughout (Fig. 1D-F). The posterior face of the antenna club is similar to the anterior face. The antennal scape is short, much shorter than the funicle or club, and is rounded or widened distally, somewhat triangular. There are long, fimbriate setae on the scape. As is the case in most Micracidini, there are 6 segments in the funicle. The frons is convex and variously punctured or reticulated. Frontal vestiture is short, usually sparse, and much shorter than the setae on the antennal scape. In most species the central part of the frons is impunctate and less setose in a longitidinal band.
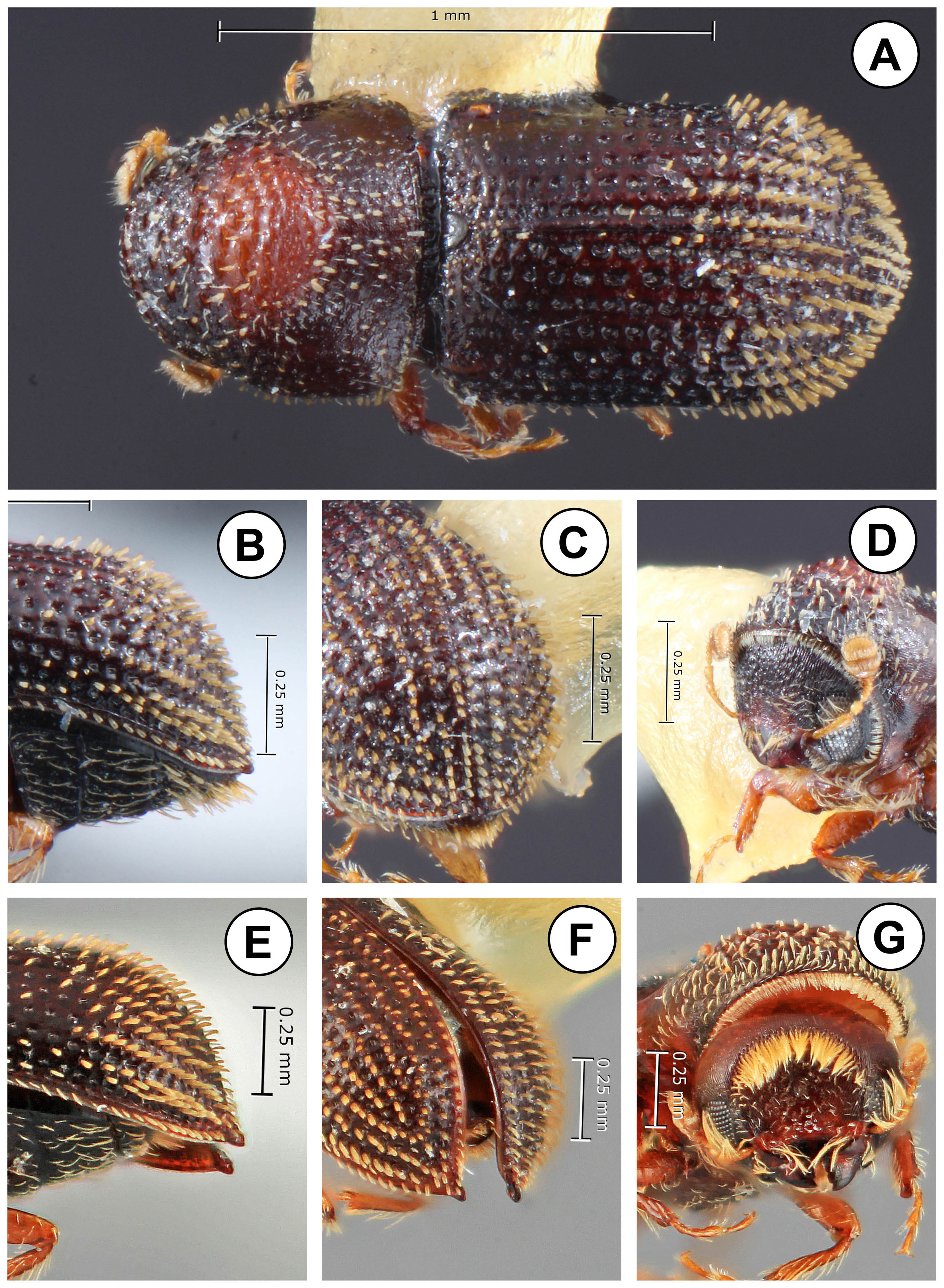
The anterior margin of the pronotum is slightly elevated in most species, with 4 or more moderately sized marginal teeth, though these are absent in a few species. The anterior slope has large, widely spaced rugosities. The pronotal summit is clearly defined, abruptly elevated, at or near the middle, with the rugosities smaller, abundant and confused. The pronotal disc typically has abundant, semi-recumbent, setae, directed anteriorad. The protibia is slender, with an undivided, curved mucro apically.
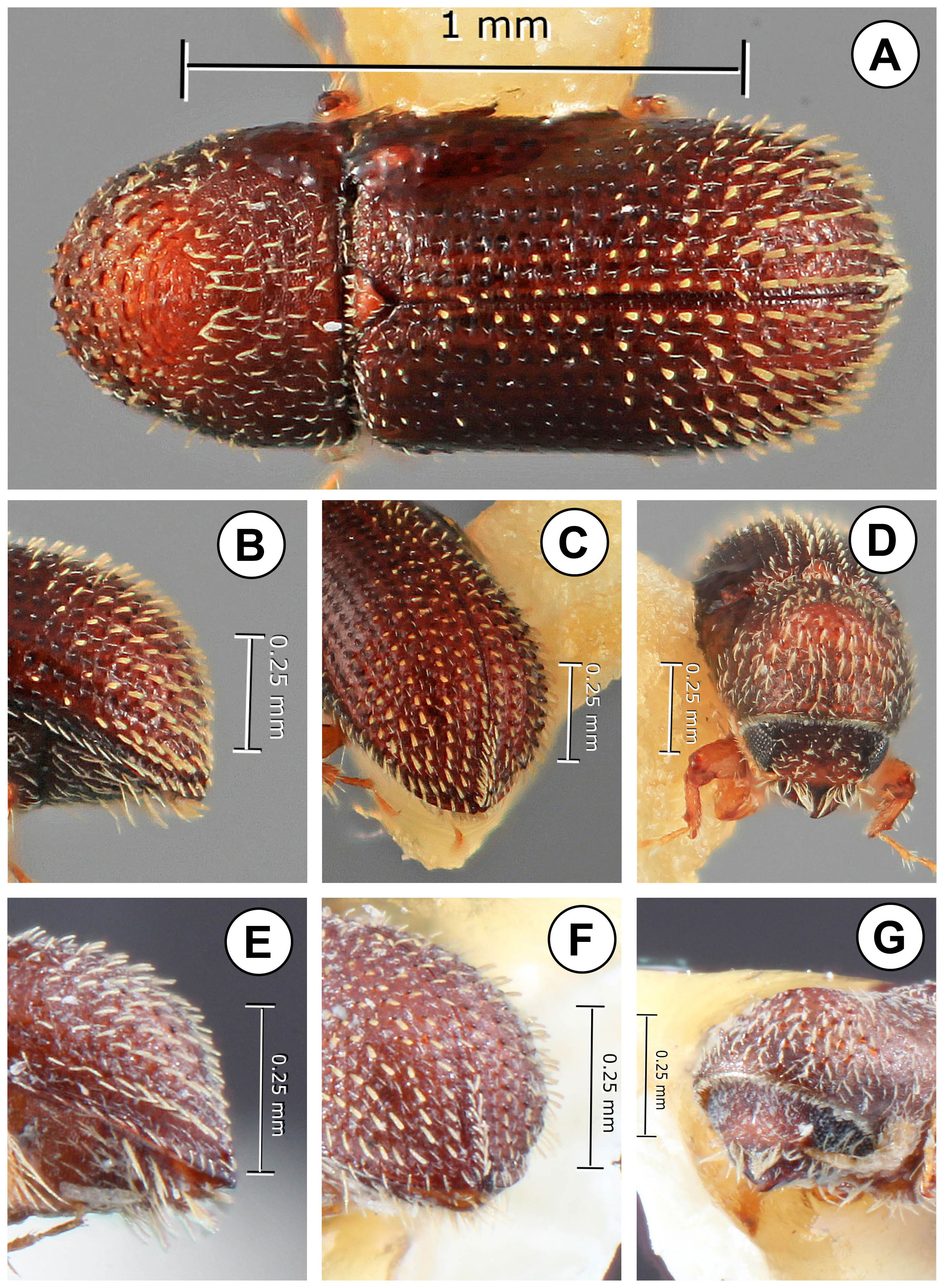
The elytral disc occupies 3/4 of the elytral length. Strial punctures are uniseriate and prominent (Fig. 2A-D), ranging from moderately to deeply impressed. Interstrial setae are uniseriate, and flattened in some species, generally longer towards the declivity. Recumbent strial setae are present, and in 1 species nearly as long as the strial setae.
In all species the declivital apex is sharply acuminate in dorsal view (Fig. 3A-F). There is a relatively abrupt change in the slope and appearance between the elytral disc and the declivity. Towards the base of the declivity interstrial granules or denticles are present on all interstriae. All species have visible granules at the base of the declivity, some quite large and modified. In some species these form a complete circumdeclivital ring, similar to those in some species of Micracis LeConte and Hylocurus Eichhoff (Fig. 3C-F). The posterolateral portion of interstria 9 is elevated, weakly or strongly serrate, and forms an upcurved crest, in line with granules at the base of the declivity on other interstriae.
The American species in the tribe Micracidini comprise over 300 species in 7 genera (Wood, 1986). Genera are primarily separated on the basis of characters of the antennal club, declivity and protibiae. Other characters do not consistently separate genera, as they do not occur in all species of a genus, and similar characters, such as a circumdeclivital ring of interstrial asperities occur in several genera. While species of Phloeocleptus include several characters that make them recognizable with respect to other related genera, the only character that is absolutely unique is the structure of the antennal club. Wood (1982) included Phloeocleptus punctatus Wood, 1980 and Hylocurus tresmariae Schedl, 1956 in the genus, apparently because both have a somewhat acuminate elytral apex. This elytral apex is a key character used in his generic keys (1982, 1986) to separate Phloeocleptus from Pseudothysanoes Blackman. This is not strictly consistent as he included a species with an acuminate elytral apex, Pseudothysanoes mucronatus Wood, 1980 in that genus. The antennae of Pseudothysanoes punctatus, P. tresmariae and P. huachucae Blackman, 1943 (the latter included as an undisputed member of the genus Pseudothysanoes) are shown in figure 1 (D-F). In all of these the antennal club is more oval in shape, rounded apically, with 2 external sutures marked with complete rows of setae. In addition, the antennal scape is slender and elongated, almost as long as the funicle. For this reason, P. tresmariae and P. punctatus are transferred from Phloeocleptus to Pseudothysanoes. Likewise, a closer examination of the unique type of Phloeocleptus parvus Wood, 1969 shows that it fits better in the genus Hylocurus and is transferred to that genus. The antennal scape is more elongate than is typical for Phloeocleptus and the club is rounded with weakly recurved sutures. The pronotal summit is not elevated. Finally, the posterior face of the protibial is rugose, a character of Hylocurus. As a convenience to students of the group, these 3 species that are removed from Phloeocleptus are illustrated and redescribed at the end of the paper.
Sexual dimorphism. In females, the antennal scape is wider distally, more triangular, and with more abundant setae compared to males in which the scape is more rounded and with fewer setae (Fig. 1A, B). This is the only easily observed external character for determining the sex of individuals of all species. Unfortunately, this character is variable among species so it is not possible to determine the sex of a single individual by this character alone. In most species the declivital armature is larger in males, in some cases markedly so, while in other species the declivital differences between the sexes are subtle and of little use in determining sex (Figs. 4-12).
Biology. All of the species have been exclusively collected in branches of species of Persea (Lauraceae), including commercial and criollo varieties of avocado, Persea americana Mill. While some species have been collected in commercially grown varieties, my personal experience is that they are more likely to be found in criollo varieties which are closer to the “wild type”. Beetles breed in broken, cut, or shaded out branches on living trees. There is no indication that any of these species are capable of causing damage to healthy trees and most avocado growers are unaware of them. Phloeocleptus caudatus Wood has also been collected from Persea schiedeana Nees. Collecting notes by S.L. Wood from Puebla suggest that some of his collections might also have been from this host (Wood, 1982). All collections have been made at elevations between 800-2,000 m, in pine-oak forest, mesic montane forest, or in agricultural or patio settings.
Males make a short entrance tunnel (1-2 mm long) in the phloem under the bark, perpendicular to the wood fiber. From this entry tunnel, he begins 2 short extensions parallel to the grain in opposite directions. Typically, 2 females join the founding male, each of which extends 1 of the short branches, again, parallel to the grain. Tunnels generally lightly score the underlying sapwood, depending on the thickness of the phloem. Unlike most phloem-feeding Scolytinae, females fill the tunnels behind them with frass, not keeping them open. At least initially, the founding male will remain in the tunnel entrance, even though he is not in direct contact with either female. Females place eggs directly in the oviposition tunnel, separated from each other by packed frass, not into niches cut into the tunnel walls as is most common in phloem-feeding Scolytinae. Because eggs are left in the tunnels, subsequent larval tunnels can go off in either direction, giving an irregular appearance to the pattern of adult and larval mines. At low population levels many gallery systems are elongated in both directions from the initial starting point. At higher population densities it is common for other males to bore in and intercept females in their oviposition tunnels. At this point, 1 or more additional females may join him. The result can be a confusing pattern that does not conform to the simple case.
Phylogeny. Species in the tribe Micracidini are primarily found in the Afrotropical and Neotropical regions where the group has undergone extensive diversifications. Jordal and Kaidel (2017) examined the phylogeny of the tribe at the generic level. All American groups were monophyletic as were African groups, suggesting a single dispersal event to the Americas from a possible African origin. Their study suggest that the age of the tribe is 89.8 mya, with the age of the American clade as 74.8 mya. In their phylogeny, the genus Phloeocleptus was sister to Cactopinus Blackman. Like Phloeocleptus, Cactopinus is restricted to Mexico and adjacent arid regions of the southwestern United States.
Phloeocleptus ardis Wood 1981 (Fig. 4)
Phloeocleptus ardis Wood 1981: 124
Comments. Contrary to Wood’s description, the holotype is a female. All 8 specimens in the original type series were females. Later other specimens from the same series as the types were sent to Wood that included males but his description was never modified and these specimens were not formally designated as types. Wood’s confusion about the sex of the holotype is due to the fact that the declivity of the females resembles that of males of other species and because no males were present in the specimens he had at hand for his original description. The male is described here for the first time.
Diagnosis. Males of this species have asperities at the base of the declivity that form a complete ring around the declivity, similar to P. caudatus Wood, and P. spicatus Wood. Females differ from those of the other 2 species in that they also have a complete ring of asperities surrounding the declivity, resembling males of other species.

Male. Length 1.44; width 0.61 mm, length/width = 2.36 (n = 1). Antennal scape rounded, fimbriate setae sparse and short, slightly longer than length of scape. Funicle and club typical for the genus. Anterior margin of pronotum slightly elevated, with 4-6 apically curved asperities. Summit prominent slightly anterior to middle; anterior slope with abundant, rounded asperities, separated by distances less than their widths, interspersed with erect setae; asperities on summit smaller, denser and confused. Pronotal disc with large, deep punctures, separated by less than their diameters. Surface granulate, with abundant, anteriorly-directed setae from pronotal base to summit. Striae on elytral disc deep, large, set within a row by 1/2 their diameters, especially near suture, becoming shallower laterally; interstriae much narrower than striae. Interstriae smooth, becoming granulate near the base of the declivity. Interstrial setae erect, apically acute, becoming longer posteriorly. Transition to declivity abrupt, with a large, acute, hooked tooth on each interstria, forming a complete circumdeclivital ring. Interstria 9 elevated, serrate, with acute denticles, forming a posterolateral crest that joins the costal margin before the suture and continues in line with the asperities at the declivital base on other interstriae. Declivital apex strongly acute, face convex, with strial punctures slightly shallower than those on disc, some acute denticles on interstria 3 near the base.
Female. Length 1.6; width 0.64 mm, length/width = 2.5 (n = 1). Similar to male except as noted. Antennal scape distally widened, almost triangular with abundant fimbriate setae almost twice as long as the scape. Interstrial denticles at the base of elytral declivity much shorter and rounded apically on all interstriae. Posterolateral crest of interstria 9 with smaller, less acute denticles.
Distribution. Known only from the type series collected in a moist, pine-oak forest above Monterrey.
Material examined. Mexico: Nuevo León: cerro Chipinque, 1,300 m, 30-I-1980, Persea sp., T.H. Atkinson, S-20, (USNM 8 [holotype, paratypes], TAMU 2,
CEAM 12).
Phloeocleptus atkinsoni Wood 1981 (Fig. 5)
Phloeocleptus atkinsoni Wood 1981:124
Phloeocleptus cristatus Wood 1981: 125, new synonymy

Diagnosis. This species most closely resembles P. obscurus and P. plagiatus. In these species the elytral striae are much less impressed and there is no circumdeclivital ring of large asperities in either sex. In these 3 species differences in the declivities of males and females are slight.

Male. Length 1.43-1.73 mm; width 0.51 mm, length/width = 2.29 (n = 7). Antennal scape rounded, fimbriate setae in a distinct tuft, about twice as long as length of scape. Funicle and club typical for the genus. Anterior margin of pronotum slightly elevated, with 4-6 apically curved asperities. Summit prominent, slightly behind middle; anterior slope with rounded asperities, separated by distances greater than their widths, interspersed with erect setae; asperities on summit smaller, denser and confused. Surface of pronotal disc granulate, with abundant, anteriorly-directed setae from pronotal base to summit; those closest to summit flattened, yellow; those close to base paler and thinner. Striae on elytral disc not impressed, punctures shallow, set within a row slightly more than their diameters. Strial setae recumbent, about as long as diameter of associated puncture. Interstriae slightly wider than striae, smooth, becoming weakly granulate near the base of the declivity. Interstrial setae, erect, apically obtuse, shorter than interstrial width, becoming longer posteriorly. Declivity steep, transition gradual. Declivital interstria 1 slightly elevated, with uniseriate granules and setae similar to those on the disc to apex. Interstria 2 with granules on basal half, continuing nearly to apex on interstria 3. Interstria 9 weakly elevated, with rounded granules, forming a posterolateral crest that joins the costal margin before the suture and continues in line with the asperities at the declivital base on other interstriae. Declivital apex strongly acute, face convex, with strial punctures slightly shallower than those on disc.

Female. Length 1.27-1.73 mm; width 0.491 mm, length/width = 2.31 (n = 6). Antennal club slightly wider distally, triangular, with much more abundant setae that males. Granules on the declivity are slightly smaller than those of the male.
Distribution. This is the most commonly collected species and has been collected in Michoacán, Morelos, Puebla and Oaxaca at elevations from 892 to 2,150 m.
Material examined. Mexico: Michoacán: Nuevo San Juan Parangarícutiro, huerta El Durazno 2, 19.2230° N, 102.1416° W, 2,245 m, VII-2016-VI-2017, bottle trap with ethanol in avocado orchard, M. Lazaro D., (CEAM 1); Uruapan, 1,600 m, 17-II-1980, Persea americana, T.H. Atkinson, S-26, (USNM 5 [holotype, allotype P. cristatus], TAMU 46, CEAM 16); Morelos: Carr. Tetela-Hueyapan, km 6, 2,080 m, 19-V-1984, Persea americana, E. Saucedo C., SM-273, (CEAM 2); Carr. Xochimilco-Oaxtepec, desv. Tlanepantla, 2,070 m, 2-IX-1982, Persea americana, Burgos S., A., SM-43, (UAEM 1); Jumiltepec, 2,250 m, 21-I-1982, Persea americana, T.H. Atkinson, A. Equihua M., S-302, (UAEM 1); Jumiltepec, 2,250 m, 21-I-1982, Persea americana, S-555, (CEAM 7); Tepoztlan, 1,710 m, 20-VI-1980, Persea americana, T.H. Atkinson, S-71, (USNM 5 [holotype, allotype, paratypes P. atkinsoni], CEAM 17); Tepoztlan, 1,710 m, 23-IV-1981, Persea americana, E. Saucedo C., A. Burgos S., S-621, (CEAM 4); Tetela, 2,020 m, 4-V-1985, Persea americana, A. Equihua M., E. Estrada V., S-991, (CEAM 11); Tlalnepantla, 1,670 m, 4-VII-1989, Persea americana, A. Equihua M., (CEAM 6); Oaxaca: Sta. Ma. Xadani, 10 km N, Rancho Dioon, 15.9333° N, 96.0067° W, 22-III-2008, Persea americana, T.H. Atkinson, (UTIC 3); Sta. Ma. Xadani, 10 km N, Rancho Dioon, 15.9926° N, 96.1055° W, 892 m, 24-VI-2009, Persea americana, T.H. Atkinson, THA-876, (TAMU 5); Sta. Catarina Lachatao, 17.2693° N, 96.4729° W, 2,110 m, 25-I-2013, Persea americana, T.H. Atkinson, THA-982, (UTIC 16); Puebla: Atlixco, 1,830 m, 19-I-1983, Persea americana, T.H. Atkinson, S-847, (CEAM 6, TAMU 2); Tiaguismanalco, 2,150 m, 19-I-1983, Persea americana, T.H. Atkinson, S-848, (CEAM 7, TAMU 1).
Comments. Wood (1981) described P. atkinsoni and P.cristatus from Michoacán and Morelos, respectively, in the same publication. The differences cited in the descriptions are minute. The type series of both, as well as numerous other specimens collected since 1981 were examined. There is no basis for recognizing 2 separate species.
Phloeocleptus caudatus Wood 1956 (Fig. 6)
Phloeocleptus caudatus Wood 1956:147
Diagnosis. Males of this species resemble those of P. ardis and P. spicatus because of the elevated circumdeclivital ring on the elytra. Females of P. caudatus lack this elevated ring.
Male. Length 1.36-2.01 mm; width 0.58 mm, length/width = 2.49 (n = 2). Antennal scape triangular, fimbriate setae sparse and short, slightly longer than length of scape. Funicle and club typical for the genus. Anterior margin of pronotum slightly elevated, with 4-6 apically curved asperities. Summit prominent, slightly posterior to middle; anterior slope with rounded asperities, separated by distances equal to their width, interspersed with erect setae; asperities on summit smaller, denser and confused. Pronotal disc punctate and granulate, with abundant, anteriorly-directed setae from pronotal base to summit. Striae on elytral disc deep, large, set within a row by 1/4 their diameters, especially near suture, becoming shallower laterally; interstriae much narrower than striae, almost obsolete. Interstriae smooth, becoming granulate near the base of the declivity, with 3-4 prominent denticles anterior to base of elytra. Interstrial setae erect, fine, becoming longer posteriorly. Transition to declivity abrupt, with a large, rounded elevation on each interstria, occupying its full width, forming a complete circumdeclivital ring. Interstria 9 elevated, with closely spaced, distinct, rounded granules, forming a posterolateral crest that joins the costal margin before the suture and continues in line with the asperities at the declivital base on other interstriae. Declivital apex strongly acute, face flattened, with strial punctures shallower than those on disc. Declivital interstria 1 elevated, with denticles along its entire length, becoming larger towards apex. Denticles irregularly spaced on other interstriae, especially interstria 3.
Female. Length 2.04 mm; width 0.71 mm, length/width = 2.87 (n = 1). Similar to male except as noted. Antennal scape distally widened, almost triangular with abundant fimbriate setae almost twice as long as the scape. Strial punctures on elytral disc smaller, separated by about 1/2 their width; interstriae almost as wide as striae. Transition between elytral disc and declivity much less abrupt, face convex. Interstrial denticles at the base of elytral declivity much shorter and rounded apically on all interstriae, continuing on declivity on all interstriae. Posterolateral crest of interstria 9 with smaller, less acute denticles.
Distribution. Humid forests on the Sierra de Puebla and area surrounding Xalapa.
Material examined. Mexico: Puebla: Apulco, 1,600 m, 6-V-1981, Persea americana, T.H. Atkinson, A. Equihua M., S-228, (USNM 10, CEAM 12, TAMU 3); Teziutlan, 8 km NE, 1,600 m, 27-VI-1953, S.L. Wood, (SEMC 3 [holotype, allotype], USNM 10 [paratypes]); Veracruz: Banderilla, 17-II-1984, Persea americana, F.A. Noguera M., FANM-132, (CEAM 3); Coacoatzintla, 18-II-1984, Persea americana, F.A. Noguera M., FANM-136, (CEAM 1); Naolinco, 28-I-1984, Persea americana, F.A. Noguera M., FANM-118, (CEAM 4); Olite, 23-I-1984, Persea americana, F.A. Noguera M., FANM-116, (CEAM 8); San Andrés, 5-XI-1983, Persea americana, F.A. Noguera M., FANM-87, (CEAM 2); El Castillo, 19.5770 N, 96.8418 W, 1-VI-2019, branch Persea scheideana, T.H. Atkinson, THA-1259 (UTIC 2); Xalapa, 18-XII-1983, Persea americana, F.A. Noguera M., FANM-111, (CEAM 3); Xalapa, 23-X-1983, F.A. Noguera M., FANM-78, (CEAM 2).
Phloeocleptus lorenzfischeri Atkinson, new species (Fig. 7)
http://zoobank.org/urn:lsid:zoobank.org:act:7EE04F8D-
68C8-40FC-9DF2-BCB0D5A13EB4
Diagnosis. This is the most distinctive species in the genus because of its strongly developed declivital spines in both sexes. Like P. ardis, there is a circumdeclivital ring of projections in both sexes, but these are much longer and acute.

Male. Length 1.65-1.87 mm; width 0.63 mm, length/width = 2.66 (n = 6). Antennal scape rounded, fimbriate setae sparse and short, slightly longer than length of scape. Funicle and club typical for the genus. Anterior margin of pronotum slightly elevated, with 4-6 apically curved asperities. Summit prominent, slightly posterior to middle; anterior slope with rounded asperities, separated by distances equal to their width, interspersed with erect setae; asperities on summit smaller, denser and confused. Pronotal disc granulate, reticulate, with abundant, anteriorly-directed setae from pronotal base to summit; longest and densest near summit. Striae on elytral disc shallow, set within a row by greater than their diameters, larger and deeper posteriorly; interstriae about twice as wide striae. Interstriae smooth, becoming weakly granulate near the base of the declivity. Interstrial setae erect, becoming longer posteriorly; those on interstriae 1-3 short, length less than width of interstriae, yellow; longer and paler laterally. Transition to declivity abrupt, with a large, backwards pointing, acute spine on each interstria, forming a complete circumdeclivital ring. Spines on 2nd interstria 3 times as long as any other, displaced posteriorly from others; Interstria 9 elevated, almost smooth, forming a posterolateral crest that joins the costal margin before the suture and continues in line with the asperities at the declivital base on other interstriae. Declivital apex strongly acute, face flattened, with strial punctures shallower than those on disc, vestiture shorter and sparser. Declivital interstria 1 not elevated at base, with denticles along its apical 2/3 and forming an elevated costa at apex. Some denticles in the middle part of interstria 3.

Female. Length 1.63-1.80 mm; width 0.55 mm, length/width = 2.56 (n = 7). Similar to male except as noted. Antennal scape distally widened, with abundant fimbriate setae slightly longer than the scape. The declivital armature is similar to that of the male.
Distribution. Known only from the type locality in humid forest in the Sierra Sur de Oaxaca.
Type material. Holotype, male. Mexico: Oaxaca: Sta. Ma. Xadani, 10 km N, Rancho Dioon, 15.9333° N, 96.0067° W, 892 m, 24-VI-2009, Persea americana, T.H. Atkinson, THA-876, (CNIN), Allotype, female, same data (CNIN). Paratypes, same data (CNIN 1, USNM 3); Sta. Ma. Xadani, 10 km N, Rancho Dioon, 15.9333° N, 96.0067° W, 22-III-2008, Persea americana, T.H. Atkinson, (UTIC 4).
Etymology. This species is named in honor of Lorenz Fischer, a young naturalist.
Phloeocleptus nanulus Wood 1969 (Fig. 8)
Phloeocleptus nanulus Wood 1969:45
Diagnosis. This species resembles P. obscurus in that it has abundant, long setae on the declivity. It is distinguished by the larger interstrial teeth at the base of the declivity in males.
Male. Length 1.57 mm; width 0.63 mm, length/width = 2.49 (n = 1). Antennal scape rounded, setae sparse, shorter than length of scape. Funicle and club typical for the genus. Anterior margin of pronotum slightly elevated, with 2-4 apically curved asperities. Summit prominent, elevated, before middle; anterior slope with rounded asperities, separated by distances subequal to their widths, interspersed with erect setae; asperities on summit smaller, denser and confused. Surface of pronotal disc reticulate, with abundant, anteriorly-directed setae from pronotal base to summit; those closest to summit denser, longer; those close to base noticeably shorter and less dense. Striae on elytral disc not impressed, punctures shallow, set within a row about their diameters. Strial setae recumbent, shorter than diameter of associated puncture. Interstriae wider than striae, smooth, becoming weakly granulate near the base of the declivity. Interstrial setae, erect, apically obtuse, about equal to interstrial width at base, becoming much longer posteriorly and on declivital face. Declivity steep, transition gradual. Interstrial granules at base of declivity rounded on interstriae 1-3, larger and more acute laterally. Declivital interstria 1 not elevated, with uniseriate granules to apex. Interstria 2 and 3 with granules only on basal half. Interstria 9 strongly elevated, serrate, forming a posterolateral crest that joins the costal margin before the suture and continues in line with the asperities at the declivital base on other interstriae. Declivital apex strongly acute, face convex, with strial punctures slightly shallower than those on disc. Interstrial vestiture abundant, longer than on disc.
Female. Length 1.45-1.46 mm; width 0.56 mm, length/width = 2.60 (n = 2). Antennal club wider distally, triangular, with much more abundant setae than males. Declivity with granules smaller, less acute than those of male.
Distribution. Known only from the type locality.
Material examined. Mexico: Nayarit: Tepic, 7 km W, 1,000 m, 13-VII-1965, S.L. Wood, SLW-240, (USNM 3 [holotype, allotype, paratype]).
Comments. The type series was taken from branches of the same unknown tree as the type series of P. plagiatus. Given that the latter species is known from species of Persea, it is assumed that this is also the case for P. nanulus.
Phloeocleptus obscurus Wood 1956:147 (Fig. 9)
Phloeocleptus obscurus Wood 1956:147
Diagnosis. This species most closely resembles P. atkinsoni and P. plagiatus. In these species the elytral striae are much less impressed and there is no circumdeclivital ring of large asperities in either sex. In these 3 species differences in the declivities of males and females are slight. This species is distinguished by the long, spatulate setae on the declivity.
Male. Length 1.01-1.05 mm; width 0.41 mm, length/width = 2.26 (n = 2). Antennal scape rounded, fimbriate setae sparse, about as long as length of scape. Funicle and club typical for the genus. Anterior margin of pronotum slightly elevated, with 4-6 apically curved asperities. Summit prominent, at middle; anterior slope with rounded asperities, separated by distances greater than their widths, interspersed with erect setae; asperities on summit smaller, denser and confused. Surface of pronotal disc shining, with abundant, anteriorly-directed setae from pronotal base to summit; those closest to summit longer; those close to base noticeably shorter. Striae on elytral disc not impressed, punctures shallow, set within a row slightly more than their diameters. Strial setae recumbent, shorter than diameter of associated puncture. Interstriae wider than striae, smooth, becoming weakly granulate near the base of the declivity. Interstrial setae, erect, apically obtuse, about equal to interstrial width, becoming longer posteriorly. Declivity steep, transition gradual. Declivital interstria 1 not elevated, with uniseriate to apex. Interstria 2 without granules on declivital face, uniseriate to apex on interstria 3. Interstria 9 weakly elevated, with rounded granules, forming a posterolateral crest that joins the costal margin before the suture and continues in line with the asperities at the declivital base on other interstriae. Declivital apex strongly acute, face convex, with strial punctures slightly shallower than those on disc, interstrial vestiture abundant, longer than on disc.

Female. Length 1.18 mm; width 0.40 mm, length/width = 2.95 (n = 1). Antennal club wider distally, triangular, with much more abundant setae than males. Declivity similar to that of the male.
Distribution. Known only from the type locality in humid forest in San Luís Potosí.

Material examined. Mexico: San Luís Potosí: El Salto, 19-VI-1953, Persea americana, S.L. Wood, (SEMC 3 [holotype, allotype, paratype], USNM 11 [paratypes]).
Phloeocleptus plagiatus Wood 1969 (Fig. 10)
Phloeocleptus plagiatus Wood 1969:45
Diagnosis. This species most closely resembles P. atkinsoni and P. obscurus but is larger and more elongate proportionally. In these species the elytral striae are much less impressed and there is no circumdeclivital ring of large asperities in either sex. In these 3 species differences in the declivities of males and females are slight.

Male. Length 1.65-2.13 mm; width 0.65 mm, length/width = 2.66 (n = 7). Antennal scape rounded, fimbriate setae sparse, slightly longer than length of scape. Funicle and club typical for the genus. Anterior margin of pronotum not elevated or armed. Summit prominent, at middle; anterior slope with abundant, rounded asperities, almost contiguous, interspersed with erect setae; asperities on summit smaller, denser and confused. Surface of pronotal disc granulate, reticulate, with abundant, pale, fimbriate, anteriorly-directed setae from pronotal base to summit. Striae on elytral disc weakly impressed, punctures shallow, set within a row slightly less than their diameters. Strial setae recumbent, prominent, about twice as long as diameter of associated puncture. Interstriae slightly wider than striae, smooth, becoming weakly granulate near the base of the declivity. Interstrial setae pale, erect, apically obtuse, shorter than interstrial width, becoming longer posteriorly. Declivity steep, transition gradual. Strial and interstrial vestiture on elytra similar to that on disc, surface noticeably reticulate, dull. Declivital interstria 1 slightly elevated, with uniseriate granules and setae similar to those on the disc to apex. Interstria 2 with granules only near base, continuing nearly to apex on interstria 3. Interstria 9 weakly elevated, with rounded granules, forming a posterolateral crest that does not join the costal margin before the suture and continues in line with the asperities at the declivital base on other interstriae. Declivital apex strongly acute, face convex, with strial punctures slightly shallower than those on disc.
Female. Length 1.73-2.13 mm; width 0.70 mm, length/width = 2.60 (n = 7). Antennal club slightly wider distally, triangular, with much more abundant setae that males. Declivity similar to that of the male.
Distribution. Nayarit and Michoacán.
Material examined: Mexico: Michoacán: Nuevo San Juan Parangarícutiro, huerta El Durazno 2, 19.2400° N, 101.5456° W, 2,245 m, VII-2016-VI-2017, bottle trap with ethanol in avocado orchard, M. Lazaro D., (CEAM 1); San Ángel Zurumucapio, 30-V-2018, Persea americana, (UTIC 2); Uruapan, 1,600 m, 19-II-1980, Persea americana, T.H. Atkinson, S-27, (CEAM 7, UTIC 4); Uruapan, 1,600 m, 1-XI-1980, Persea americana, T.H. Atkinson, S-148, (CEAM 36, TAMU 180); Uruapan, Huerta El Colegio, 19.2119° N, 102.0334° W, 29-V-2018, Persea americana, (UTIC 12); Uruápan, huerta La Piedra China, 1,564 m, VII-2016-VI-2017, bottle trap with ethanol in avocado orchard, M. Lazaro D., (CEAM 1); Ziracuaretiro, Huerta La Ziranda, 1,304 m, VII-2016-VI-2017, bottle trap with ethanol in avocado orchard, M. Lazaro D., (CEAM 1); Nayarit: Tepic, 7 km W, 1000 m, 13-VII-1965, S.L. Wood, SLW-240, (USNM 35 [holotype, allotype, paratypes]).
Comments. The type series was taken from branches of an unknown tree, but all other host records are from Persea americana.
Phloeocleptus spicatus Wood 1981 (Fig. 11)
Phloeocleptus spicatus Wood 1981:125
Diagnosis. The males of this species resemble those of P. ardis and P. caudatus in having an elevated, complete circumdeclivital ring of teeth or asperities. As is the case with females of P. caudatus, the females of this species lack the elevated declivital ring. In both sexes the anterior margin of the pronotum is unarmed.
Male. Length 1.07 mm; width 0.41 mm, length/width = 2.48 (n = 1). Antennal scape wider distally, fimbriate setae sparse and short, slightly longer than length of scape. Funicle and club typical for the genus. Anterior margin of pronotum slightly elevated, unarmed. Summit prominent, posterior to middle; anterior slope with few, rounded asperities, separated by distances greater than their widths, largest near summit, interspersed with erect setae. Asperities on summit smaller, denser and confused. Pronotal disc with coarsely granulate. Surface granulate, with abundant, anteriorly-directed setae from pronotal base to summit; those nearest summit yellow, flattened; those nearer basal margin longer, hair-like, and paler in color. Striae on elytral disc prominent, set within a row by approximately their diameters, especially near suture, becoming shallower laterally; interstriae twice as wide as striae. Interstrial setae on first 3 interstriae flattened, yellow, length about width of interstria; striae on lateral interstriae longer, paler in color. Interstriae smooth, becoming granulate near the base of the declivity. Transition to declivity abrupt, with a large tooth or asperity on each interstria, forming a complete circumdeclivital ring; those at base of declivity acute, more rounded towards apex. Tooth on interstria 3 longest, noticeably displaced posteriorly, out of line with its neighbors. Interstria 9 elevated, weakly serrate, with small denticles, forming a posterolateral crest that joins the costal margin before the suture and continues in line with the asperities at the declivital base on other interstriae. Declivital apex strongly acute, face convex, with strial punctures slightly shallower than those on the disc. Declivital interstria 1 elevated, with small, poorly defined granules. Some small granules on the base of interstria 3.
Female. Length 1.01 mm; width 0.371 mm, length/width = 2.73 (n = 1). Similar to male except as noted. Antennal scape distally widened, almost triangular with abundant fimbriate setae more than twice as long as the scape. Elytral interstriae smooth basally, becoming granulate towards declivity. Transition between elytral disc and declivity much less abrupt, face convex. Interstrial granules at the base of elytral declivity short and rounded apically on all interstriae, continuing on declivity on all interstriae. Interstria 1 weakly elevated, with granules to apex. Posterolateral crest of interstria 9 weakly elevated.
Distribution. Known only from the type series collected in a moist, pine-oak forest above Monterrey.
Specimens examined. Mexico: Nuevo León: Cerro Chipinque, 1,300 m, 30-I-1980, Persea sp., T.H. Atkinson, S-20, (USNM 3 [holotype, allotype, paratype], CEAM 1)

Key to species. Because of the marked sexual dimorphism of some species, males and females are keyed out separately for some species. If sex is not specified, the couplet works for both.
1. Interstriae at base of declivity with enlarged rounded or acute elevations, as wide as interstria, forming a complete circumdeclivital ring (Fig. 3C-F) ……………………………………………………………………………………………… 2
1ʹ. Interstriae with granules at base of declivity smaller, usually rounded, not forming a circumdeclivital ring
(Fig. 3A, B) ……………………………………………………………………………………………… 6
2.(1) All asperities at base of declivity spinose (Fig. 4C) ……………………………………………………………………………………………… 3
2ʹ. Asperities at base of declivity mostly rounded (Fig. 6C) ……………………………………………………………………………………………… 4
3.(2) Spines at base of declivity strongly acute, projecting (Fig. 7) ……………………………………………………………………………………………… P. lorenzfischeri Atkinson
3ʹ. Spines at base of declivity much shorter, hooked (Fig. 4A-C) ……………………………………………………………………………………………… P. ardis Wood, male
4.(2ʹ) Asperities at base of declivital interstriae 1-2 acute, others rounded (Fig. 11A-C) ……………………………………………………………………………………………… P. spicatus Wood, male
4ʹ. All asperities at base of declivity rounded……………………………………………………………………………………………… 5
5.(4ʹ) Asperities at base of declivity strongly elevated (Fig. 6A-C)……………………………………………………………………………………………… P. caudatus Wood, male
5ʹ. Most asperities at base of declivity reduced, short (Fig. 4E-F) ………………………………………………………………………………………………P. ardis Wood, female
6.(1ʹ) Strial punctures large, deep on elytral disc, interstriae much narrower than striae (Fig. 4A, 11A)……………………………………………………………………………………………… 7
6ʹ. Strial punctures shallow on disc, interstriae wider than striae (Fig. 5A)……………………………………………………………………………………………… 8
7.(6) Declivital interstria 1 elevated apically; all setae on disc and declivity hair-like (Fig. 6E, F) ……………………………………………………………………………………………… P. caudatus Wood, female
7ʹ. Declivital interstria 1 not elevated apically; most setae on disc and some on declivity blunt, flattened (Fig. 11E, F) ………………………………………………………………………………………………P. spicatus Wood, female
8.(6ʹ) Setae on declivity at least twice as long as on disc (Fig. 9A, E, F) ……………………………………………………………………………………………… 9
8ʹ. Setae on declivity similar in length to those on disc, only slightly longer (Fig. 5E, F) ……………………………………………………………………………………………… 10
9.(8) Declivital interstria 1 not elevated, declivital setae blunt. (Fig. 8) ……………………………………………………………………………………………… P. nanulus Wood
9ʹ. Declivital interstria 1 strongly elevated apically, declivital setae acute (Fig. 9) ……………………………………………………………………………………………… P. obscurus Wood
10.(8ʹ) Strial setae on disc prominent, longer than diameter of associated punctures; declivital interstrial setae shorter than interstrial width (Fig. 10) ……………………………………………………………………………………………… P. plagiatus Wood
10ʹ. Strial setae on disc inconspicuous, much shorter than diameter of associated punctures, declivital interstrial setate longer than interstrial width (Fig. 5) ……………………………………………………………………………………………… P. atkinsoni Wood
Species removed from Phloeocleptus
Hylocurus parvus (Wood 1969) (Fig. 12)
Phloeocleptus parvus Wood 1969:46
Hylocurus parvus (Wood) 1969:46, new combination.
Diagnosis. The antennal scape is elongate and the club is rounded, with recurved sutures and is not consistent with that of species of Phloeocleptus. The posterior face of the protibia is rugose. Because of these characters this species is transferred to Hylocurus. The unique type specimen is in poor condition and the frons is mostly hidden. Given the variation in expression of sexual dimorphism in Hylocurus, it is not possible to tell whether this is a male or a female. It would reach couplet 8 in Wood’s 1982 key, but would go no further because of the hidden frons. It most closely resembles H. dilutus Wood.
Male (?). Length 1.49 mm; width 0.56 mm, length/width = 2.66 (n = 1). Frons on type specimen mostly hidden. Antennal scape elongate, almost as long as funicle, with a few short hairs. Club rounded, with 2 weakly procurved sutures on the anterior face, the displaced apically on the posterior face (difficult to see in specimen). Posterior face of protibial (Fig. 12C) with clearly visible asperities. Anterior margin of pronotum not elevated or armed. Summit before middle, not elevated. Anterior slope with abundant, closely spaced, curved asperities, these smaller and confused at summit. Pronotal disc shallowly punctate, punctures separated by distance greater than their diameters, surface reticulate. Setae apparently abraded. Striae on elytral disc not impressed, punctures larger, shallow, separated within rows by slightly shorter than their diameters. Interstriae twice as wide as striae, surface smooth, shining. Declivity steep, transition gradual, strial punctures larger, closer than on disc. Interstria 1 weakly elevated with small granules. Interstria 3 with 2 acute denticles in middle. Some small denticles at base of more lateral interstriae. Interstria 9 elevated, weakly serrate, not reaching costal margin. Apex strongly acute. Declivity with erect setae on most interstriae, their length slightly greater than the width of discal interstriae.
Distribution. Known only from the type locality.
Material examined. Mexico: Nayarit: Los Corchos, 10 m, 10-VII-1965, S.L. Wood, SLW-206, (USNM 1 [holotype]).
Comments. Known only from the unique holotype.
Pseudothysanoes punctatus (Wood 1980) (Fig. 13)
Phloeocleptus punctatus Wood 1980:355
Pseudothysanoes punctatus (Wood), new combination.
Diagnosis. Other than the weakly acuminate declivital apex, this is a typical species of Pseudothysanoes. The antennal scape is elongate and the antennal club has complete, horizontal sutures, unlike the case in Phloeocleptus. This species would come out to couplet 23 in Wood’s 1982 key to Pseudothysanoes, along with P. mucronatus Wood (Fig. 14) and P. tresmariae.
Male. Length 1.57 mm; width 0.62 mm, length/width = 2.53 (n = 1). Frons flattened, punctate, becoming strigose near vertex, vestiture of sparse setae below level of eyes. Antennal scape slender, about as long as funicle. Club rounded, with 2 clearly marked transverse sutures marked by rows of setae. Anterior margin of pronotum raised, armed by 6-8 denticles. Summit well defined, elevated, in middle. Anterior slope of pronotum with a few wide asperities, becoming smaller, denser and confused on summit. Pronotal disc shallowly punctate, reticulate. Some short, anterior-directed, scale-like setae from base to summit. Elytral disc with deep strial puncture, separated by less than their diameters. Interstriae about as wide as striae, surface smooth, becoming weakly granulate towards base of declivity. Interstrial setae absent on disc (possibly abraded), flattened, apically truncate, beginning near base of declivity and continuing on face of declivity. Declivity evenly convex, striae clearly, interstriae granulate to apex with associated short setae, not noticeably longer than on disc. Interstrial setae confused on interstria 9. Declivital apex with a short mucro.

Female. Length 2.13 mm; width 0.75 mm, length/width = 2.84 (n = 1). Frons deeply concave, from epistoma to vertex; surface deeply punctate with very short setae, a long fringe of fimbriate setae distributed along upper margin of concavity, between the eyes. Antennal scape slightly widened distally, covered with long setae, length about about half the length of the scape. Club similar to that of male. Elytra similar to those of male, interstrial granules absent from declivity.
Distribution. Known only from the type locality.
Material examined. Costa Rica: Guanacaste: Parque Nal. Santa Rosa, 15-XII-1979, G.C. Stevens, (USNM 5 [holotype, allotype, paratypes]).
Pseudothysanoes tresmariae (Schedl 1956) (Fig. 15)
Hylocurus tresmariae Schedl 1956:32
Phloeocleptus tresmariae (Schedl) 1956, combination Wood 1982: 570
Pseudothysanoes tresmariae (Schedl), new combination
Diagnosis. Other than the weakly acuminate declivital apex, this is a typical species of Pseudothysanoes. The antennal scape is elongate and the antennal club has complete, horizontal sutures, unlike the case in Phloeocleptus. This species would come out to couplet 23 in Wood’s 1982 key to Pseudothysanoes, along with P. mucronatus Wood (Fig. 13) and P. punctatus.
Male. Length 1.22-1.41 mm; width 0.49 mm, length/width = 2.52 (n = 2). Frons flattened, with a shallow impression above the epistoma between the eyes; surface sparsely, shallowly punctures, shining. Antennal scape slender, length slightly shorter than funicle, with long fimbriate setae, these about as long as scape. Club rounded, with 2 transverse sutures marked by rows of setae. Anterior margin of pronotum not elevated, armed with 4-6 small denticles. Pronotal summit well defined, elevated, in middle. Anterior slope with widely spaced asperities, becoming smaller, denser and confused at summit. Pronotal disc very shallowly, sparsely punctured, surface shining, with short, anterior facing, slender setae from base to summit. Elytral disc with shallow strial punctures, these separated by less than their diameters. Striae greater than twice as wide as striae. Interstrial vestiture of short, pale, flattened, blunt setae, their length about half the width of the interstriae, becoming longer towards declivity. Declivity evenly convex, striae impressed, their punctures deeper than on disc. All interstriae weakly granulate, with scale-like setae similar to those on disc. Elytral apex distinctly acuminate.

Female. Length 1.41 mm; width 0.55 mm, length/width = 2.56 (n = 1). Frons shallowly concave from epistoma to above upper level of eyes, upper margin indistinct. Surface of concavity impunctate, shining; short, slender, pale setae near upper margin of concavity, not forming a distinct fringe. Antennal scape with more abundant setae than that of male. Pronotum and elytra similar to male, but declivital striae not impressed, granules smaller.
Distribution. Known only from the Tres Marías Islands, off the coast of Nayarit.
Material examined. Mexico: Nayarit: Islas Tres Marías, María Madre, 15-V-1925, H.H. Keifer, (CAS 2 [holotype, paratype], NMW 2 [male paratype, female paratype]).
Comments. Other than the weakly acuminate declivital apex, this is a typical species of Pseudothysanoes. The male holotype and a male paratype are in the California Academy of Sciences (treated as female by Wood [1982]). A male and a female paratype are in Schedl’s collection in Vienna and are wrongly labeled as holotype and allotype, respectively (Schedl, 1979). Wood (1982) was apparently unaware of the female specimen in Vienna (NMW) and does not include it in his description.
Acknowledgments
To Harald Schillhammer of the Natural History Museum Vienna, for the loan of types. I thank Sarah M. Smith for the use of her photographs.
References
Atkinson, T. H. (2010). New species and records of Cactopinus Schwarz with a key to species (Coleoptera: Curculionidae: Scolytinae), Zookeys, 56, 17–33. https://doi.org/10.3897/zookeys.56.515
Atkinson, T. H., Saucedo, C. E., Martínez, F. E., & Burgos, S. A. (1986). Coleópteros Scolytidae y Platypodidae asociados con las comunidades vegetales de clima templado y frío en el estado de Morelos. Acta Zoológica Mexicana Nueva Serie, 17, 1–58. https://azm.ojs.inecol.mx/index.php/azm/article/view/2048/2161
Atkinson, T. H., & Equihua, M. A. (1985). Notes on biology and distribution of Mexican and Central American Scolytidae (Coleoptera). I. Hylesininae, Scolytinae except Cryphalini and Corthylini. Coleopterists Bulletin, 39, 227–238.
Jordal, B. H., & Kaidel, J. (2017). Phylogenetic analysis of Micracidini bark beetles (Coleoptera: Curculionidae) demonstrates a single trans-Atlantic disjunction and inclusion of Cactopinus in the New World clade. Canadian Entomologist, 149, 8–25. https://doi.org/10.4039/tce.2016.31
Lázaro-Dzul, M. O., Equihua-Martínez, A., Romero-Nápoles, J., González, H. H., Alvarado, D., Macías-Samaño, J. et al. (2020). Diversity of Scolytinae (Coleoptera: Curculionidae) at three sites in the Central Avocado-Producing Region of Michoacán, Mexico. Coleopterists Bulletin, 74, 454–462. https://doi.org/10.1649/0010-065X-74.2.454
Noguera, M. F. A., & Atkinson, T. H. (1990). Biogeography and biology of bark and ambrosia beetles (Coleoptera: Scolytidae and Platypodidae) in a mesic montane forest in Mexico with an annotated checklist of species. Annals of the Entomological Society of America, 83, 453–466. https://doi.org/10.1093/aesa/83.3.453
Schedl, K. E. (1956). Some bark and ambrosia beetles from the Tres Maria Islands, Mexico. 143 Contribution. Pan Pacific Entomologist, 32, 30–32.
Schedl, K. E. (1979). Die Typen der Sammlung Schedl Familie Scolytidae (Coleoptera). Kataloge der Wissenschaftlichen Sammlung des Naturhistorischen Museums in Wien, 3, 1–286.
Wild, A. (2015). Build a world-class insect imaging system for under $6,000. Compound Eye Blog, Scientific American. Available at (accessed July 22, 2022): https://blogs.scientificamerican.com/compound-eye/build-a-world-class-
insect-imaging-system-for-under-6-000/
Wood, S. L. (1956). New species of bark beetles (Coleoptera: Scolytidae), mostly Mexican, Part I. Canadian Entomologist, 88, 141–154. https://doi.org/10.4039/Ent88141-4
Wood, S. L. (1969). New records and species of neotropical bark beetles (Scolytidae: Coleoptera), Part IV. Brigham Young University Science Bulletin, Biology Series, 10, 1–46.
Wood, S. L. (1980). New American bark beetles (Coleoptera: Scolytidae), with two recently introduced species. Great Basin Naturalist, 40, 353–358.
Wood, S. L. (1981). Nomenclatorial changes and new species in Platypodidae and Scolytidae (Coleoptera). Great Basin Naturalist, 41, 121–128.
Wood, S. L. (1982). The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae), a taxonomic monograph. Great Basin Naturalist Memoirs, 6, 1–1356.
Wood, S. L. (1986). A reclassification of the genera of Scolytidae (Coleoptera) Great Basin Naturalist Memoirs, 10, 1–126.


