
Silvia Cappello-García a, *, Manuel Antonio García-García a, Ma. Guadalupe Rivas-Acuña a, Santa Dolores Carreño-Ruiz b, Joaquín Cifuentes-Blanco c
a Universidad Juárez Autónoma de Tabasco, División Académica de Ciencias Biológicas, Laboratorio de Micología, Km. 0.5, Carr. Villahermosa-Cárdenas entronque a Bosque de Saloya, 86150 Villahermosa, Tabasco, México
b Universidad Autónoma de Chiapas, Facultad Maya de Estudios Agropecuarios, Carr. Catazajá-Palenque, Km. 4, 29980 Catazajá, Chiapas, México
c Universidad Nacional Autónoma de México, Facultad de Ciencias, Herbario FCME, Ciudad Universitaria, Apartado postal 70-181, 04510 Ciudad de México, México
*Autor para correspondencia: cappellogs@hotmail.com (S. Capello-García)
Recibido: 3 mayo 2022; aceptado: 5 mayo 2023
Resumen
El género Ganoderma se ha estudiado escasamente en el sureste de México. El objetivo del presente estudio fue contribuir al conocimiento de la diversidad de especies de Ganoderma para México y Tabasco. Se analizaron 247 ejemplares de herbario provenientes de 16 municipios del estado. Se observaron las características macro y micromorfológicas de los basidiomas. Las observaciones microscópicas se hicieron en KOH al 5% y la determinación taxonómica se realizó mediante diversas claves especializadas. Se identificaron y describieron 21 especies de Ganoderma, de las cuales 17 corresponden al subgénero Ganoderma y 4 al subgénero Elfvingia. G. carnosum, G. dunense, G. lipsiense, G. multipileum, G. multiplicatum, G. neojaponicum, G. orbiforme, G. philippii, G. sichuanense, G. stipitatum y G. subresinosum son registros nuevos para México. Dichas especies también son reportes nuevos para Tabasco, incluyendo a G. australe, G. oerstedii, G. oregonense, G. sessiliforme, G. tsugae y G. zonatum. Se presenta una clave dicotómica para las especies identificadas. El inventario refleja la diversidad de Ganoderma en México y el trópico tabasqueño.
Palabras clave: Subgénero Elfvingia; Basidioma laqueado; Hongos xilófagos; Basidiosporas equinuladas; Corología
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
New records of the genus Ganoderma (Agaricomycetes, Polyporales) for Mexico and Tabasco
Abstract
The genus Ganoderma has been poorly studied in southeastern Mexico. The aim of this study was to contribute to the knowledge of diversity of species of the Ganoderma for Mexico and Tabasco. Two hundred and forty seven herbarium specimens from 16 municipalities of the state were analyzed. The macro and micromorphological characteristics of the basidiomes were observed. Microscopic observations were made in KOH at 5%, and taxonomic determination was made using various specialized keys. Twenty-one species of Ganoderma were identified and described, of which 17 correspond to the subgenus Ganoderma and 4 to the subgenus Elfvingia. G. carnosum, G. dunense, G. lipsiense, G. multipileum, G. multiplicatum, G. neojaponicum, G. orbiforme, G. philippii, G. sichuanense, G. stipitatum, and
G. subresinosum are new records for Mexico. These species are also new reports for Tabasco, including G. australe, G. oerstedii, G. oregonense, G. sessiliforme, G. tsugae, and G. zonatum. A dichotomous key is presented for the identified species. The inventory reflects the diversity of Ganoderma in Mexico and the Tabasco tropics.
Keywords: Subgenus Elfvingia; Lacquered basidioma; Xylophagous fungi; Echinulate basidiospores; Chorology
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
De acuerdo con Zmitrovich (2018), el género Ganoderma P. Karst., pertenece a la familia Ganodermataceae dentro del orden de los Polyporales y comprende 487 especies (Index Fungorum, 2022). El género se caracteriza por poseer basidiocarpos prominentes, estipitados o sésiles, anuales o perennes, con poros en la superficie himenial de forma y tamaño variable. Las basidiosporas van de elipsoidales a ovoides, con ápices truncados, equinuladas y con pared de doble capa con exosporio delgado y endosporio grueso y coloreado (Kaur et al., 2018; Torres-Torres y Guzmán-Dávalos, 2012). Debido a la plasticidad y complejidad del género, algunas características macro y micromorfológicas resultan determinantes en la taxonomía tradicional de los ejemplares, como el tipo de contexto, elementos cuticulares, tamaño y color de los poros, forma y tamaño de las basidiosporas (Bhosle et al., 2010; Gottlieb y Wright, 1999a, b; Torres-Torres et al., 2012). En la actualidad los estudios filogenéticos han permitido esclarecer la plasticidad morfológica de diversas especies del género Ganoderma (Cao et al., 2012; Hapuarachchi et al., 2019; Jargalmaa et al., 2017; Lima-Júnior et al., 2014; Moncalvo et al., 1995; Tchotet Tchoumi et al., 2018; Thawthong et al., 2017; Wang et al., 2009, 2014; Zhou et al., 2015). No obstante, para el presente estudio se siguió la propuesta taxonómica de Zheng et al. (2009) basada en rasgos morfológicos, donde segregan al género en 2 grupos distintos, las especies lacadas (subgénero Ganoderma del complejo G. lucidum) y las no lacadas (subgénero Elfvingia del complejo G. applanatum). El objetivo de la investigación fue contribuir al conocimiento de la diversidad de especies del género Ganoderma para México y Tabasco.
Materiales y métodos
Se revisaron 247 ejemplares depositados en el Herbario de la División Académica de Ciencias Biológicas (UJAT). Las muestras provienen de 16 municipios del estado de Tabasco (tabla 1). Se siguió la metodología propuesta por Torres-Torres et al. (2012) para analizar cada una de las características macro y micromorfológicas de los especímenes. Para las características macroscópicas se consideró el tamaño del basidioma, la forma y dureza del píleo, el tipo de contexto, con o sin incrustaciones resinosas. Para el color de las estructuras se siguió la guía de Küppers (2002). Para las observaciones microscópicas se hicieron cortes finos con navaja y fueron montados en KOH al 5% y reactivo de Melzer. Se tomó en cuenta la forma, tamaño y ornamentación de las basidiosporas, elementos cuticulares y grosor de los disepimentos (Bhosle et al., 2010; Gottlieb y Wright, 1999a, b; Torres-Torres y Guzmán-Dávalos, 2012). Para medir las estructuras microscópicas se usó el programa AxioVisión 4.6. Para cada ejemplar se midieron 20 basidiosporas seleccionadas al azar para determinar la relación Q (longitud-ancho) según Bas (1969). La determinación de los ejemplares se realizó mediante el uso de claves taxonómicas y literatura especializada, y se verificó con descripciones previas de ejemplares (Bhosle et al., 2010; Gomes-Silva et al., 2011; Gottlieb y Wright 1999a, b; Hapuarachchi et al., 2018; Kaur et al., 2017, 2018; López-Peña et al., 2016; Loyd et al., 2018; Ryvarden, 2000; Torres-Torres et al., 2012, 2015). Se proporciona la descripción completa, la ilustración de las especies y se presenta una clave dicotómica para la identificación de los taxones estudiados en la zona.
Resultados
Se registraron 21 especies de Ganoderma (tabla 2), 17 del subgénero Ganoderma y 4 del subgénero Elfvingia. Ganoderma dunense, G. multipileum, G. neojaponicum, G. sichuanense y G. subresinosum son nuevos registros para el continente americano. Ganoderma carnosum, G. lipsiense, G. multiplicatum, G. orbiforme, G. philippii, G. stipitatum son nuevos registros para México y G. australe, G. oerstedii, G. oregonense, G. sessiliforme, G. tsugae, G. zonatum para Tabasco.
| Tabla 1
Localidades de recolecta de los especímenes en Tabasco. |
||||||
| Localidad | Municipio | Vegetación | Altitud
(msnm) |
Clima | Coordenadas | |
| Ejido la Cuchilla, Rancho San Juan | Balancán | Acahual | 5 | Am: cálido húmedo con abundantes lluvias en verano | 17°81’55” | 91°14’55.7” |
| R/o La discordia. Col. Guadalupe | Vegetación urbana | 15 | 17°49’11” | 91°32’03” | ||
| cabecera Municipal Cárdenas | Cárdenas | Acahual | 30 | 17°59’37” | 93°22’26” | |
| Poblado C-29 | Agroecosistema cacaotal | 18 | 18°02’58” | 93°26’30.8” | ||
| Hacienda Cacao km 10 carretera Cárdenas-Villahermosa | Selva alta perennifolia | 10 | 17°59’05” | 93°20’09” | ||
| Tabasquillo a 7.30 km NE de la carretera interestatal Frontera-Jonuta | Centla | Acahual | 03-05 | 18°22’05” | 92°40’10” | |
| El Faisán | Manglar | 3 | 18°32’50” | 92°33’51” | ||
| Ejido la Sabana | Selva baja caducifolia | 15 | 18°26’49.78” | 92°50’19.13” | ||
| Jardines de la Cd. Deportiva de Villahermosa | Centro | Vegetación urbana | 15 | 17°58’34” | 92°56’30” | |
| Estrellas Buenavista | Vegetación urbana | 7 | 17°57’19” | 93°01’11” | ||
| Carretera Apasco | Orilla de carretera | 4 | 18°02’00” | 92°54’34” | ||
| cabecera municipal | Vegetación urbana | 5 | 17°59’29.43” | 92°56’03.99” | ||
| Parrilla II carretera Villahermosa a Teapa | Vegetación urbana | 7 | 17°87’53” | 92°52’21” | ||
| Parque museo la Venta | Vegetación urbana | 7 | 18°00’05” | 92°56′08” | ||
| ranchería Curahueso | Vegetación urbana | 15 | 17°57’55” | 92°55’23” | ||
| ranchería Anacleto Calle Pino Suárez Col. Tamulté de las Sabanas. | Acahual | 21 | 18°09’31.88” | 92°47’11.52” | ||
| Río Carrizal | Acahual | 5-10 | 18°00’18” | 92°57’22” | ||
| DACBIOL km 0.5 carretera Villahermosa-Cárdenas desviación Bosques de Saloya | Tintal | 5-10 | 17°59’22” | 92°58’25” | ||
| Jardín Botánico Parque Museo de la Venta | Vegetación urbana | 15 | 18°00’03” | 92°56’05” | ||
| Col. Petrolera | Vegetación urbana | 15 | 18°00’25.14” | 92°56’08.70” | ||
| R/a Anacleto Canabal primera sección | Cercos vivos | 11 | 17°58’26.00” | 92°59’44.00” | ||
| Villa Luis Gil Pérez | Acahual | 10 | 17°52’45” | 93°04’18” | ||
| Yumka’ | Acahual de selva alta | 10 | 17°59’59” | 92°48’22” | ||
| ranchería Zapotal 2ª Secc. | Comalcalco | Agroecosistema cacaotal | 5 | 18°18’46” | 93°15’41” | |
| cabecera Municipal Comalcalco | Acahual | 30 | 18°15’40” | 93°13’18” | ||
| ranchería Centro Tular 2ª sec. | Acahual | 5 | 18°20’02” | 93°24’09” | ||
| R/a. Nicolás Bravo 3ra sección | Selva alta perennifolia | 7 | 18°17’39” | 93°07’48” | ||
| Poblado Chichicapa | Selva mediana subperennifolia | 10 | 18°14’59” | 93°10’57” | ||
| ranchería Pichucalco primera sección. | Cunduacán | Acahual | 10 | 18°05’37” | 93°09’53” | |
| Fraccionamiento San Antonio | Acahual | 16 | 18°03’46” | 93°09’57” | ||
| cabecera municipal Cunduacán | Vegetación urbana | 10 | 18°04’20” | 93°10’16” | ||
| Poblado Huimango 2da sección | Vegetación urbana | 11 | 18°06’19” | 93°08’31” | ||
| cabecera Municipal de Emiliano Zapata | Emiliano Zapata | Acahual | 5-10 | 17°47’44” | 91°46’36” | |
| Malpasito | Huimanguillo | Acahual | 160 | Af: cálido húmedo con lluvias todo el año | 17°20’40.8” | 93°35’26.1” |
| Agua Selva | Acahual de selva alta | 189 | 17°20’38” | 93°35’40” | ||
| cabecera Municipal de Huimanguillo | Vegetación urbana | 10-15 | 17°49’57” | 93°23’30” | ||
| ranchería Paredón 1ª secc. | Vegetación urbana | 15 | 17º 45’46” | 93º 22’40” | ||
| Tabla 1. Continúa | ||||||
| Localidad | Municipio | Vegetación | Altitud
(msnm) |
Clima | Coordenadas | |
| Víctor Fernández Manero 2da secc. | Jalapa | Selva mediana subperennifolia | 10 | 17°51’20” | 92°48’06” | |
| Cabecera municipal | Vegetación urbana | 18 | 17°38’15” | 92°50’06” | ||
| Ranchería Chacalapa 1ra secc. | Jalpa de Méndez | Agroecosistema cacaotal | 10 | Am: cálido húmedo con abundantes lluvias en verano | 18°09’06” | 93°05’31” |
| Parque Estatal Agua Blanca | Macuspana | Selva alta perennifolia | 20-100 | Af: cálido húmedo con lluvias todo el año | 17°37’17” | 92°29’05” |
| Apasco | Vegetación urbana | 10 | 17°45´44” | 92°44’00” | ||
| Poblado Límbano Blandín | Acahual | 31 | 17°45’19.0” | 92°24’42.4” | ||
| ranchería San Antonio | Vegetación urbana | 14 | 17°43’06” | 92°55’02” | ||
| ranchería San Carlos | Acahual | 14 | 17°50’05” | 92°31’49” | ||
| Carretera caparroso 1era. Sección | Acahual de selva alta | 26 | 17°36’42” | 92°29’14.7” | ||
| Ranchería Arroyo | Nacajuca | Selva mediana subperennifolia | 8 | Am: cálido húmedo con abundantes lluvias en verano | 18°07’25.6” | 92°59’28.2” |
| Ranchería La Cruz 1ra Sección | Vegetación urbana | 10 | 18°07’05” | 93°00’00” | ||
| Cabecera municipal de Nacajuca | Vegetación urbana | 10 | 18°10’13” | 93°00’54” | ||
| Ranchería Belén | Acahual | 05-10 | 18°12’00” | 92°57’22” | ||
| Bosques de Saloya sobre carretera Nacajuca-Villahermosa | Pastizal | 10 | 18°05’01” | 92°55’01” | ||
| Ranchería Potreritos | Paraíso | Agroecosistema Palmar | 12 | 18°18’42” | 93°16’53” | |
| Ejido las flores | Acahual | 20 | 17°21’50” | 93°35’08” | ||
| Ejido el Madrigal 5a secc. | Tacotalpa | Acahual | 23 | Af: cálido húmedo con lluvias todo el año | 17°35’36” | 92°46’49” |
| Estación Biológica la Florida, Tapijulapa | Acahual de selva alta | 45 | 17°27’48.3’’ | 92°46’12.3’’ | ||
| Ejido Agua Blanca, sierra Tapijulapa | Vegetación urbana | 53 | 17°29’09” | 92°37’18” | ||
| Villa Luz | Selva mediana subperennifolia | 120 | 17°26’32” | 92°45’52” | ||
| Ejido Pomoca | Acahual de selva alta | 40 | 17°29’53” | 92°39’31” | ||
| Ejido Cerro Blanco 5ta Sección | Acahual de selva alta | 106 | 17°26’48.9” | 92°48’44.7” | ||
| Ejido La Pila | Acahual de selva alta | 170 | 17°26’20” | 92°45’20” | ||
| Ejido Santa Rosa 1era. Sección | Acahual | 20 | 17°34’39” | 92°48’02” | ||
| San Manuel, sierra Tapijulapa | Acahual | 128 | 17°29’03” | 92°45’05” | ||
| Ejido Lomas Alegres 3ra Sección San Antonio, sierra Poaná | Acahual de selva alta | 42-54 | 17°35’02” | 92°47’10” | ||
| Estación Poaná-sierra Poaná | Selva mediana subperennifolia | 70 | 17°33’32” | 92°42’40” | ||
| Centro Ecoturístico Yu-Balcah, Ejido Ceibita | Acahual de selva alta | 60 | 17°35’12” | 92°49’25” | ||
| 2 km de la desviación a Oxolotán de la carretera T | Riparia | 40 | 17º 27’07” | 92°47’11” | ||
| Ranchería San Antonio | Teapa | Agroecosistema cacaotal | 40-130 | 17°31’03” | 92°52’02” | |
| Puyacatengo | Acahual | 76 | 17°31’04” | 92°55’31” | ||
| Cabecera Municipal de Teapa | Acahual | 100 | 17°33’55” | 93°57’58” | ||
| Monumento Natural Grutas de Cocona | Selva alta perennifolia | 100-150 | 17°34’40” | 93°56’01” | ||
| Rancho El Tigre en punta de montaña 30 km de la Palma hacia Guatemala sobre el Rio San Pedro | Tenosique | Selva alta perennifolia | 50 | Am: cálido húmedo con abundantes lluvias en verano | 17°35’40” | 91°28’10” |
| Ranchería Santo Tomás Centro Integrador Tenosique | Selva alta perennifolia | 60 | 17°17’15” | 91°20’10” |
Tabla 2
Lista de especies previamente citadas para México y los nuevos registros contemplados en el estudio.
| Especies | Estados de la República Mexicana | Referencia |
| **Ganoderma carnosum | — | — |
| Ganoderma colossus | Chiapas, Quintana Roo, Tabasco, Veracruz, Yucatán | Ramos y Cappello (2009), Torres-Torres et al. (2015) |
| Ganoderma curtisii | Baja California, Estado de México, Guanajuato, Hidalgo, Jalisco, Michoacán, Morelos, Nuevo León, Oaxaca, Sonora, Tabasco, Yucatán | Ayala y Guzmán (1984), Castillo y Guzmán (1970), Guzmán (1983), Guzmán-Dávalos (2005), López-Peña et al. (2016), Nava-Mora y Valenzuela-Garza (1997), Ojeda-López et al. (1986), Ramos y Cappello (2009), Torres-Torres et al. (2015), Torres-Torres y Guzmán-Dávalos (2005), Welden y Guzmán (1978) |
| *Ganoderma dunense | — | — |
| *Ganoderma multipileum | — | — |
| **Ganoderma multiplicatum | — | — |
| *Ganoderma neojaponicum | — | — |
| ***Ganoderma oerstedii
= G. tuberculosum |
Baja California Sur, Chiapas, Durango, Guerrero, Jalisco, Morelos, Oaxaca, Sinaloa, Sonora, Veracruz | Bandala et al. (1993), López-Peña et al. (2016), Torres-Torres et al. (2015) |
| **Ganoderma orbiforme | — | — |
| ***Ganoderma oregonense | Hidalgo, Estado de México, Veracruz | Torres-Torres et al. (2015) |
| Ganoderma resinaceum
= G. aerolatum = G. sessile = G. subincrustatum = G. perturbatum |
Colima, Hidalgo, Jalisco, Nuevo León, Oaxaca, Quintana Roo, Sonora, Tabasco, Veracruz | Guzmán (1972), López-Peña et al. (2019), Ramos y Cappello (2009), Raymundo y Valenzuela (2003), Torres-Torres et al. (2015), Welden y Guzmán (1978) |
| ***Ganoderma sessiliforme | Morelos, Sonora | Guzmán (1972), López-Peña et al. (2016), Murrill (1912), Torres-Torres et al. (2012; 2015) |
| *Ganoderma sichuanense | — | — |
| **Ganoderma stipitatum
=G. parvulum |
— | — |
| *Ganoderma subresinosum | — | — |
| ***Ganoderma tsugae | Durango, Estado de México, Hidalgo | Bandala et al. (1993) |
| Ganoderma applanatum | Guanajuato, Nuevo León, Oaxaca, Sonora, Tabasco | Cappello (2006), Castillo y Guzmán (1970), Guzmán (1972), López-Peña et al. (2016), Ojeda-López (1986) |
| ***Ganoderma australe
= G. tornatum |
Guanajuato, Nayarit, Oaxaca, San Luis Potosí, Tamaulipas, Veracruz | Bandala et al. (1993), Guzmán (1972), Ojeda-López et al. (1986) |
| **Ganoderma lipsiense | — | — |
| **Ganoderma philippii | — | — |
| Ganoderma mexicanum | Estado de México | Torres-Torres et al. (2015) |
| Ganoderma lobatum | Baja California Sur, Colima, Durango, Guanajuato, Hidalgo, Michoacán, Nuevo León, Oaxaca, Puebla, Quintana Roo, San Luis Potosí, Sinaloa, Sonora, Tabasco, Tamaulipas, Veracruz, Zacatecas | Bandala et al. (1993), Guzmán (1972), López-Peña et al. (2016), Ramos y Cappello (2009), Raymundo y Valenzuela (2003) |
| Tabla 2. Continúa | ||
| Especies | Estados de la República Mexicana | Referencia |
| Ganoderma lucidum
= G. nitens |
Baja California Norte, Baja California Sur, Campeche, Chiapas, Chihuahua, Colima, Distrito Federal, Durango, Estado de México, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Morelos, Nuevo León, Oaxaca, Quintana Roo, San Luis Potosí, Sinaloa, Sonora, Tabasco, Tamaulipas, Veracruz, Zacatecas | Guzmán (1972), Ramos y Cappello (2009), Raymundo y Valenzuela (2003), Welden y Guzmán (1978) |
| Ganoderma weberianum | Jalisco, Sonora | López-Peña et al. (2016), Torres-Torres et al. (2015) |
| Ganoderma mexicanum | Estado de México | Torres-Torres et al. (2015) |
| Ganoderma brownii | Baja California Sur, Hidalgo, Nuevo León, Sinaloa, Valle de México, Veracruz | Bandala et al. (1993), Welden y Guzmán (1978) |
| Ganoderma neglectus | Guanajuato | Guzmán (1972) |
| Ganoderma polycromum | Nuevo León | Guzmán (1972) |
| ***Ganoderma zonatum | Baja California Sur, Jalisco, Nayarit, Puebla | Bandala et al. (1993), Guzmán (1972), Torres-Torres et al. (2015) |
(*) Registros nuevos para América y México. (**) Registros nuevos para México. (***) Registro nuevo para Tabasco
Descripciones
Subgénero Ganoderma
Ganoderma carnosum Pat., Bull. Soc. mycol. Fr. 5(2,3): 66 (1889) (fig. 1A)
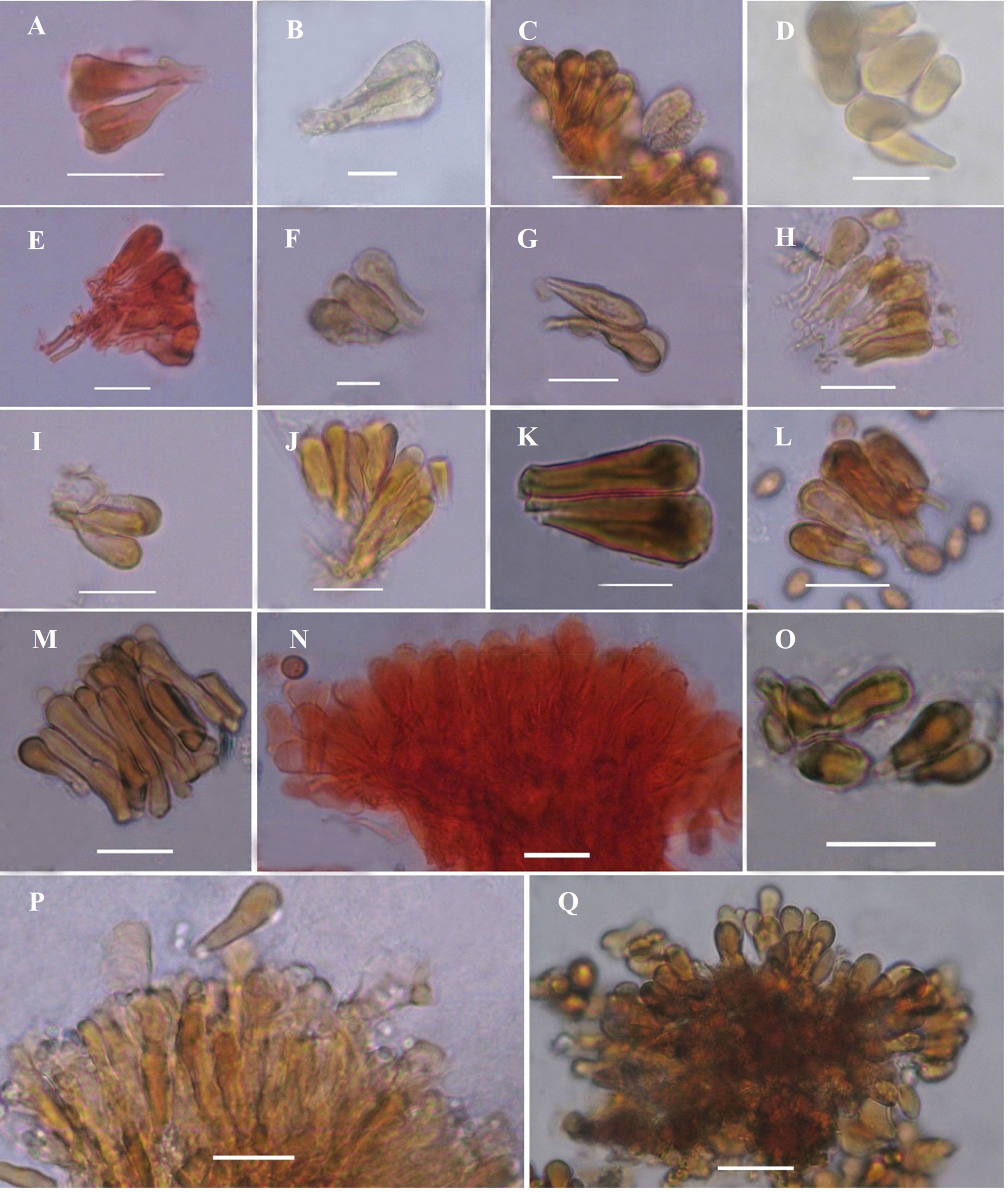
Basidioma sésil. Píleo de 105-135 × 75-90 × 5 mm, reniforme a semicircular, liso, sulcado concéntricamente, laqueado, color marrón oscuro (N90 Y70 M90) a negro, siendo más brillante cuando joven. Margen obtuso, carnoso, estéril, concoloro al píleo, presenta una franja hacia la parte himenial que culmina en un borde con tono anaranjado, amarillo o marrón claro. Contexto dúplex, de 10 mm de grosor, con bandas oscuras de depósitos de resina, rojizo (N80 Y99 M70) hacia los tubos y marrón claro (N60 Y70 M50) hacia la parte superior. Tubos de 9 mm de longitud, marrón claro (N20 Y50 M30). Poros circulares de 4-5 por mm, lisos, color marrón claro (N40 Y30 M40). Disepimentos de 46-66 μm de grosor. Sistema hifal dimítico. Elementos cuticulares subcilíndricos a clavados, hialinos, 21-25 × 7-9 μm (fig. 6A). Hifas esqueléticas del contexto, lisas, subhialinas a marrón claro, ramificadas, 3-5 μm diam. Hifas conectivas ramificadas, hialinas, 2-3 μm diam. Basidiosporas elipsoidales, truncadas en el ápice, color marrón claro, 8-10 × 6-7 μm, Q = 1.38.
Resumen taxonómico
Material examinado. México, Tabasco, Cárdenas, cabecera municipal, acahual. 09/III/2009. Areanna (UJAT 4671). Centla, El Faisán, mangle rojo. 16/XI/1998. López-Bonilla (UJAT 1181). Comalcalco, cabecera municipal, acahual. 21/XI/1998. López-Bonilla (UJAT 1397).
Distribución. África: Marruecos. Asia: India, Irán, Japón, Nepal, Sri Lanka, Turquía. América: Bolivia, Canadá, Colombia, Estados Unidos. Europa: Alemania, Austria, España, Estonia, Eslovenia, Francia, Grecia, Reino Unido, República Checa, Rusia, Polonia, Suiza (Cilerdzic et al., 2011; Karadelev y Rusevska, 2004; Moncalvo, 2000; Papp y Szabó, 2013; Patouillard, 1889). Registro nuevo para México.
Comentarios taxonómicos
Puede confundirse con Ganoderma oregonense Murrill, pero ésta difiere en lo rojizo del píleo sin llegar a negro, en el contexto homogéneo y en lo subestipitado del basidioma. Las características coinciden con la descripción de Patouillard (1889), pero difiere ligeramente con la longitud de las basidiosporas de 12-13 × 6-8 μm, mientras que en el material estudiado son de 8-10 × 6-7 μm, similar a lo descrito por Cilerdzic et al. (2011) (8.98-11.52 × 5.42-7.24 μm).
Ganoderma colossus (Fr.) C.F. Baker, Brotéria, sér. bot. 18(no. 2): 37 (1920) (fig. 1B)
Basidioma sésil a pileado-estipitado. Píleo de 180-290 × 100-120 ×20 mm, esponjoso, liviano, ligeramente laqueado, glabro, color amarillo claro (N10 Y60 M20) a rojo claro (N30 Y80 M40) con tonos anaranjados (N30 Y99 M60) en gran parte de la superficie. Margen grueso, obtuso, ondulado, estéril, concoloro al píleo. Contexto homogéneo de 35 mm de grosor, color amarillo (N10 Y60 M20), aspecto densamente lanoso. Tubos de 18 mm de longitud, color marrón (N30 Y60 M40). Poros circulares a angulares, irregulares, de 2-3 por mm, color marrón claro a amarillo (N00 Y60 M30). Disepimentos lisos, gruesos, 60.42-94.41 μm de grosor. Estípite corto, 5 × 3.5 cm, cilíndrico, concoloro al píleo. Sistema hifal dimítico. Elementos cuticulares claviformes, marrón claro, lisos, 24-35 μm (fig. 6B). Hifas esqueléticas raramente ramificadas, lisas, subhialinas, 4-7 μm diam. Hifas conectivas ramificadas, hialinas, 2-4 μm diam. Basidiosporas elipsoidales a ovoides, truncadas o parcialmente redondeadas en el ápice, color marrón claro, 12-18 × 8-12 μm, Q = 1.5.

Resumen taxonómico
Material examinado. México, Tabasco, Centla, El Faisán, Tintal. 30/IX/1998. Cappello GS (UJAT 1114). Centro, Parrilla II, vegetación urbana. 16/VI/2006. Rodríguez-Quero (4861 UJAT). Comalcalco, Ejido Río playa, cacaotal. 26/IX/1998. Marín-Romero (UJAT 1230). Cunduacán, cabecera municipal. 03/XI/2009. Olmedo-López (UJAT 4860).
Distribución. África: Camerún, Nigeria, Senegal, Surinam, Zaire. América: Brasil, Costa Rica, Estados Unidos. Asia: China, India, Pakistán, Taiwán, Vietnam. (Gomes-Silva et al., 2011; Hapuarachchi et al., 2019; Hong y Jung, 2004; Loyd et al., 2018; Murrill, 1905; Parihar et al., 2013; Ryvarden, 2000; Steyaert, 1972; Welti y Courtecuisse, 2010; Wu y Zhang, 2003; Zhou et al., 2015). Para México, en los estados de Chiapas, Oaxaca, Quintana Roo, Tabasco, Veracruz y Yucatán (Ramos y Cappello, 2009; Raymundo y Valenzuela, 2003; Torres-Torres et al., 2015; Welden y
Guzmán, 1978).
Comentarios taxonómicos
Ganoderma colossus puede considerarse como la única especie con basidioma grande, esponjoso, ligero, de color amarillo claro a anaranjado, y basidiosporas grandes, lo que permite diferenciarla del resto de las especies del género. Ryvarden (2000) refiere que esta especie presenta basidioma sésil, mientras que Parihar et al. (2013) y Torres-Torres et al. (2015) lo describieron de sésil a estipitado, lo cual coincide con los ejemplares estudiados. Por estas características únicas es imposible confundirla con alguna otra especie del género Ganoderma.
Ganoderma curtisii (Berk.) Murrill, N. Amer. Fl. (New York) 9(2): 120 (1908) (fig. 1C)
Basidioma pileado-estipitado. Píleo de 140 ×100 × 10 mm, laqueado, dimidiado, semicircular, leñoso, rugoso, color marrón (N90 Y90 M99) con tonos amarillos (N60 Y90 M50). Margen delgado, estéril, lobulado, flexionado ligeramente hacia la superficie del píleo. Contexto dúplex de 10 mm de grosor, de marrón a rojizo (N90 Y90 M70) con tonos amarillos (N70 Y90 M50), con bandas continuas de depósitos de resina. Tubos de 8 mm de longitud, color marrón (N70 Y80 M50). Poros angulares de 4-5 por mm, dentados. Disepimentos delgados, escabrosos, 35-40 μm de grosor, marrón claro (Y30 M10 C10). Estípite semicilíndrico, excéntrico, rojizo a negro (N80 Y99 M70), laqueado, 12 × 3 cm. Sistema hifal trimítico. Elementos cuticulares claviformes, subhialinos, lisos, 24-30 × 7-8 μm (fig. 6C). Hifas generativas no observadas. Hifas esqueléticas escasamente ramificadas, color marrón claro, 7.11-8 μm diam. Hifas conectivas hialinas, lisas, 3.8-5.9 μm diam. Basidiosporas elipsoidales a oblongas, marrón claro, truncadas en el ápice, 8.34-11.27 × 5.73-7.33 μm, Q = 1.50.
Resumen taxonómico
Material examinado. México, Tabasco, Centro, Parque Museo La Venta, selva mediana. 28/IX/2002. Cappello GS (UJAT 4883); 11/XI/2019. García-Manuel (UJAT 5108); ranchería Curahueso, huerto familiar. 25/IX/2018. Cappello GS (UJAT 4908); Av. Universidad s/n UJAT, vegetación urbana. 14/VI/1999. Morales (UJAT 1400); DACBIOL km 0.5 carretera Villahermosa-Cárdenas, Tintal. 04/X/2019. García-Manuel (UJAT 5104). Jalapa, Víctor Fernández Manero 1ra secc., acahual. 07/VIII/2004. Camacho (UJAT 4911). Macuspana, Bitzales 7ma secc. vegetación urbana. 08/X/2016. Castro-Torres (UJAT 5106); Parque Estatal Agua Blanca, selva mediana. 18/X/2019. García-Manuel (UJAT 5149).
Distribución. África: Ghana, sierra Leona, Zambia, Zimbabue. América: Brasil, Canadá, República Domi-
nicana, Ecuador, Estados Unidos, Guatemala, Perú, Asia: Filipinas, India, Irak, Irán, Japón. Europa: Francia (Bhosle et al., 2010; Kaur et al., 2017; Loyd et al., 2018; Murrill, 1908; Zhou et al., 2015). Para México, en Baja California, Estado de México, Guanajuato, Hidalgo, Jalisco, Morelos, Nuevo León, Sonora, Oaxaca, Tabasco y Yucatán (Ayala y Guzmán, 1984; Castillo y Guzmán, 1970; Guzmán, 1983; López-Peña et al., 2016; Nava-Mora y Valenzuela-Garza, 1997; Ojeda-López et al., 1986; Ramos y Cappello, 2009; Raymundo y Valenzuela, 2003; Torres-Torres et al., 2015; Torres-Torres y Guzmán-Dávalos, 2005).
Comentarios taxonómicos
Ganoderma curtisii puede confundirse morfológicamente debido a las variaciones estructurales microscópicas, lo cual se ve reflejado en el color y brillo del píleo según la madurez del ejemplar (Torres-Torres y Guzmán-Dávalos, 2005). Los autores mencionan que el contexto dúplex con bandas de resina es distintivo de la especie ya que permanece sin importar la edad y el estado del basidioma. G. ravenelii (Steyaert 1980) tiene similitud a G. curtisii, pero difiere al carecer de bandas o depósitos de resina en el contexto (Loyd et al., 2017).
Ganoderma dunense Tchotet, Rajchenb. et Jol. Roux, in Tchotet, Coetzee, Rajchenberg, Wingfield et Roux, Australas. Pl. Path. 47(4): 440 (2018) (fig. 1D)
Basidioma sésil. Píleo de 70-110 × 50-70 × 8 mm, semicircular, flabeliforme a reniforme, dimidiado, consistencia esponjosa, leñoso, sulcado al centro, superficie superior laqueada brillante, marrón (N90 Y80 M60), con tonos o manchas azules (N20 M30 C50), usualmente presenta una fina capa de basidiosporas marrón claro. Margen estéril, liso, ligeramente lobulado, obtuso, redondo, concoloro al píleo. Contexto dúplex, esponjoso, de 5 mm de grosor, con depósitos de resina, rojizo (N90 Y80 M70) hacia los tubos y marrón claro (N60 Y80 M50) hacia la superficie superior. Tubos de hasta 14 mm de longitud, marrones (N10 Y70 M40). Poros circulares de 3-4 por mm, de color marrón a amarillo claro (N10 Y50 M10), se manchan de marrón al maltratarse. Disepimentos, 46-120 μm de grosor. Sistema hifal trimítico. Elementos cuticulares claviformes, marrón claro, 20-33 × 5-9 μm (fig. 6D). Hifas generativas no observadas. Hifas esqueléticas raramente ramificadas, marrón claro, 5-7 μm diam. Hifas conectivas ramificadas, 2.5-3.6 μm diam. Basidiosporas elipsoidales a ovoides, truncadas en el ápice, marrón claro, 10.5-12.2 × 7-8.3 μm, Q = 1.48.
Resumen taxonómico
Material examinado. México, Tabasco, Huimanguillo, Malpasito, selva mediana. 01/II/2011. Valera (UJAT 3045). Jalapa, Víctor Fernández Manero 1ra secc., selva mediana. 21/II/2005. Carreño-Ruiz (UJAT 3226). Macuspana, Parque Estatal Agua Blanca, selva mediana. 25/VI/2011. Lázaro (UJAT 3258); 28/I/2014. Sánchez-Litzajaya (UJAT 3965). Tenosique, ranchería Santo Tomás, selva alta. 28/VII/1988. Cappello GS (UJAT 604).
Distribución. África: Cabo Occidental (George) (Tchotet Tchoumi et al., 2018). Registro nuevo para el continente americano.
Comentarios taxonómicos
La descripción de la especie coincide con el registro de Tchotet Tchoumi et al. (2018), quienes refieren que G. dunense es similar a las especies del complejo G. lucidum P. Karst. Puede confundirse con G. aridicola J.H. Xing y B.K. Cui, pero se diferencia por presentar mayor tamaño de los poros (3-4/ mm vs. 6-8/ mm).
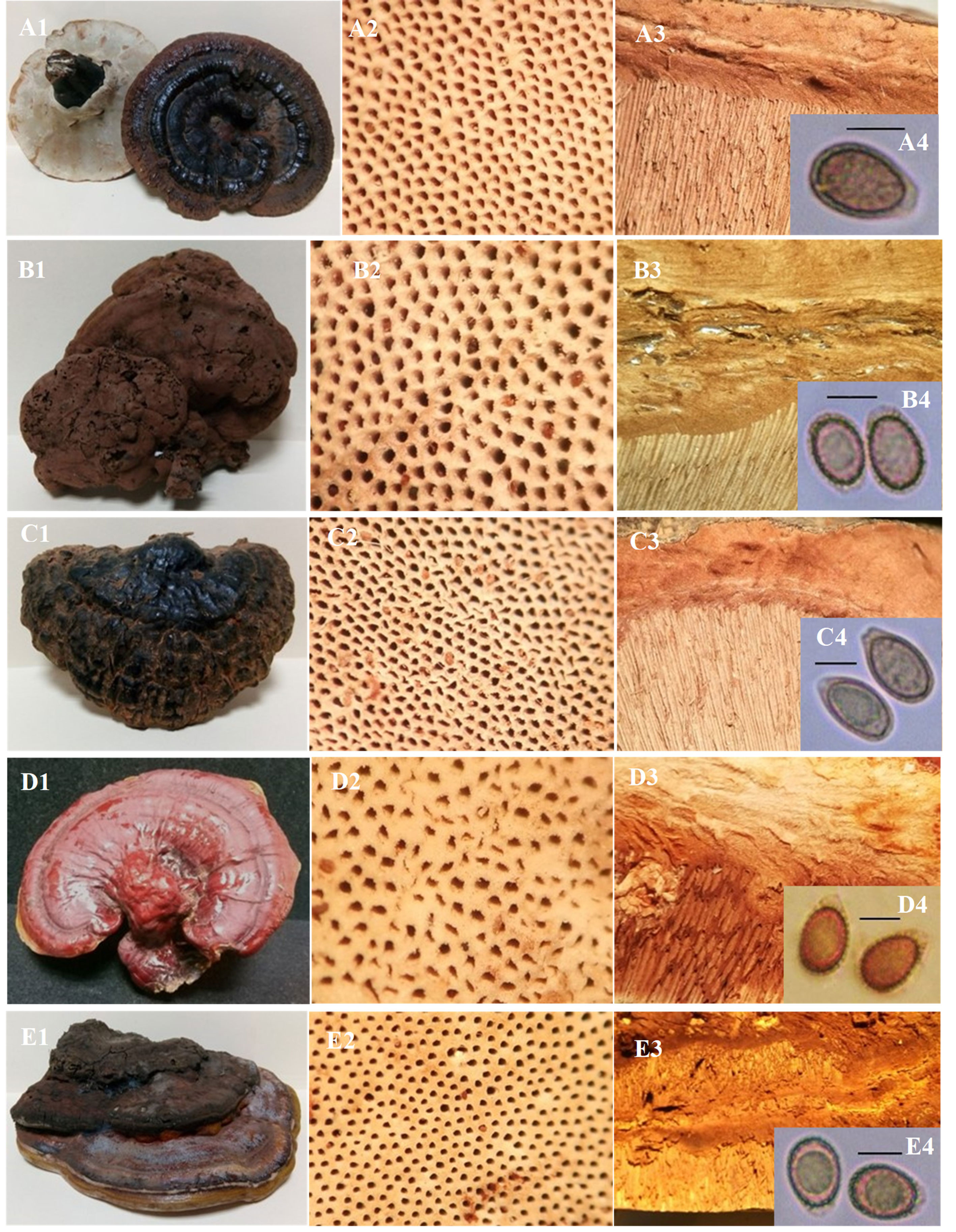
Ganoderma multipileum Ding Hou [as ‘multipilea’], Quarterly Journal of the Taiwan Museum 3: 101 (1950) (fig. 2A)
Basidioma sésil. Píleo de 90-120 × 40-60 × 10 mm, reniforme a flabelado, dimidiado, sulcado, corrugado, laqueado de ligero a fuerte, presenta tonos rojizos, anaranjados (N90 Y90 M80; N60 Y90 M80) y amarillentos. Margen obtuso, estéril, con tonos de blanco amarillento y anaranjado (N60 Y90 M80). Contexto no completamente homogéneo de 15 mm de grosor, color marrón con tonos amarillos (N10 Y80 M40). Tubos de 10 mm de longitud, marrón claro (N10 Y80 M20). Poros circulares de 5-6 por mm, de marrón claro a amarillo claro (N10 Y80 M10). Disepimentos lisos, amarillos, 38.13-88.75 μm de grosor. Sistema hifal trimítico. Elementos cuticulares ausentes. Hifas esqueléticas rara vez ramificadas, de marrón a amarillo al reaccionar con Melzer, dextrinoides, 4.21-7.62 μm diam. Hifas conectivas ramificadas, hialinas, 2.16-3.18 μm diam. Basidiosporas elipsoidales, truncadas en el ápice, marrón claro, finamente equinuladas, 8-9 × 5-6 μm, Q = 1.54.
Resumen taxonómico
Material examinado. México, Tabasco, Balancán, ranchería la Discordia, vegetación urbana. 04/VIII/2008. Hernández-Gabriela (UJAT 4879). Cárdenas, cabecera municipal, vegetación urbana. 04/IV/2019. Cappello GS (UJAT 4844). Centro, cabecera municipal, vegetación urbana. 20/II/2019. García-Manuel (UJAT 5155); 05/II/2019 García-Manuel (UJAT 5110); 21/II/2018. Cappello GS (UJAT 4852); 17/IX/2007. Torres-Pérez (UJAT 4880); 08/VI2006. Cappello GS (UJAT 3206); 08/08/2006. Hernández Piedra (UJAT 4922); Yumka’ selva mediana. 08/VII/2010. Magaña GE (UJAT 253); 12/X/1998. Ceferino-Petrona (UJAT 1479); 21/X/1998. Ceferino-Contreras (UJAT 1037). Comalcalco, cabecera municipal, acahual. 02/X/000. López-Bonilla (UJAT 1392); cacaotal. 15/II/1997. Marín-Romero (UJAT 969); 14/IX/1997. Marín-Romero (UJAT 1226); 24/I/1998. Marín-Romero (UJAT 1205); huerto familiar. 20/IX/1998. Marín-Romero (UJAT 1195); ranchería Nicolás Bravo 3ra sec., acahual. 11/III/2008. Martínez-Rodríguez (UJAT 4917); ranchería Centro Tular 2ª sec., acahual. 07/III/1999. Izquierdo-Pérez (UJAT 1398). Cunduacán, poblado Huimango 2da sec., vegetación urbana. 30/IX/2019. García-Manuel (UJAT 5005). Emiliano Zapata, cabecera municipal, acahual. 06/XI/2005. Camacho (UJAT 3192). Jalpa de Méndez, ranchería Chacalapa 1ra. secc. cacaotal. 24/IX/2008. Hernández Vargas (UJAT 4232). Macuspana, Parque Estatal Agua Blanca, selva mediana. 05/VII/2011. Fajardo (UJAT 3243); 27/III/2007. García (UJAT 3155); 21/II/2005. Gonzalo-Macías (UJAT 3290); 08/VIII/2006. Goque-Díaz (UJAT 4873). Nacajuca, cabecera municipal. 13/I/1998. Cappello GS (UJAT 1012); 11/I/1987. Sánchez-Ruiz (UJAT 25); ranchería La Cruz 1ra secc., acahual. 17/IX/2008. Cappello GS (UJAT 4998); 25/IX/2009. Hernández (UJAT 281). Tacotalpa, sierra Tapijulapa, selva mediana. 24/II/2012. Trinidad López (UJAT 4921).
Distribución. África: Costa de Marfil, Uganda. Asia: China, India, Singapur, Taiwán (Cao et al., 2012; Hapuarachchi et al., 2019; Wang et al., 2009; Zhou et al., 2015; Zohmangaiha et al., 2019). Registro nuevo para el continente americano.
Comentarios taxonómicos
Ganoderma lingzhi (Sheng H. Wu, Y. Cao et Y.C. Dai 2012) es similar a G. multipileum, pero difiere al presentar esporas no tan equinuladas y poros con tonos amarillo azufre (Cao et al., 2012). G. multipileum es el nombre válido para G. lucidum en Asia tropical según Wang et al. (2009) basado en secuencias nucleotídicas de ITS+IGS+RPB2. Se caracteriza por la presencia de múltiples píleos en el basidioma. La especie presenta diferencias mínimas en el tamaño de las basidiosporas, Wang et al. (2009) las reportan de 8-13.5 × 5.5-7.5 μm y Zhou et al. (2015) de 8.8-10.5 × 5.5-7 μm mientras que en el material estudiado son de 8-9 × 5-6 μm.
Ganoderma multiplicatum (Mont.) Pat., Bull. Soc. mycol. Fr. 5(23): 74 (1889) (fig. 2B)
Basidioma sésil. Píleo de 180 × 100 × 10 mm, laqueado, semicircular, de plano a convexo, sulcado concéntricamente, dimidiado, leñoso, rojizo hacia la base (N90 Y80 M90) con tonos anaranjados hacia el margen (N40 Y80 M50), presenta una zona diferenciada e hinchada en la base. Margen grueso, estéril, lobulado, color blanco en fresco a amarillo claro en seco. Contexto dúplex de 20 mm de grosor, fibroso, con depósitos de resina, rojizo hacia los tubos (N90 Y80 M80) y marrón claro (N70 Y80 M50) hacia la parte superior. Tubos de 8 mm de longitud marrón claro (N10 Y30 M10). Poros circulares, 5-6 por mm, marrón claro (N10 Y50 M30), se manchan de marrón rojizo al maltratarse. Sobre la superficie himenial presentan color blanco a gris en fresco y marrón claro (N70 Y80 M50) cuando seco. Disepimentos, 47-84 µm de grosor. Sistema hifal dimítico. Elementos cuticulares no observados. Hifas esqueléticas de color marrón, 5-9 µm diam. Hifas conectivas, 3-4 µm diam. Basidiosporas elipsoidales, truncadas, equinuladas, 9-11 × 6-7.5 µm, Q = 1.48.

Resumen taxonómico
Material examinado. México, Tabasco, Centla, Simón Sarlat, selva mediana. 18/IX/1999. López Bonilla (UJAT 1090); manglar. 11/IX/1998. López Bonilla (UJAT 1078). Centro, cabecera municipal, vegetación urbana. 03/X/2011. Cappello GS (UJAT 4901); 29/V/2001 Aguirre Velazco (UJAT 4885); 16/XI/2009. Mendoza-Hernández (UJAT 4887). Comalcalco, ranchería Zapotal 2ª sec., acahual. 24/I/1998. Marín-Romero (UJAT 2931). Jalapa, acahual. 04/VIII/2004. Camacho (UJAT 4877). Macuspana, Parque Estatal Agua Blanca, selva mediana. 25/II/2012. Carreño-Ruíz (UJAT 4878). Paraíso, cacaotal-potreritos. 13/X/2001. López-Bonilla (UJAT 4881); Ejido las Flores, acahual. 26/IX/2006. Hernández Sánchez (UJAT 3254). Tacotalpa, sierra Tapijulapa, acahual. 08/VI/2003. Díaz-Contreras (UJAT 4884).
Distribución. América: Brasil, Guyana. Asia: Birmania, China, India, Tahití (Bhosle et al., 2010; Gomes-Silva et al., 2011; Gottlieb y Wright, 1999a; Hapuarachchi et al., 2019; Patouillard, 1889; Ryvarden, 2000; Steyaert, 1980; Torres-Torres et al., 2012). Registro nuevo para México.
Comentarios taxonómicos
Ganoderma multiplicatum presenta un píleo hinchado hacia la base de color rojizo a negro con tonos naranjas hacia el margen, y poros de color blanco a gris en fresco tornándose marrón claro al secarse. La descripción coincide con Hapuarachchi et al. (2019) y Torres-Torres et al. (2012). G. multiplicatum fue considerada similar a G. chalceum Steyaert y G. subamboinense Bazzalo y J.E. Wright, pero Correia de Lima et al. (2014) determinaron que se trata de una especie separada con base en secuencias nucleotídicas de las regiones ITS y LSU (rDNA).
Ganoderma neojaponicum Imaseki Bull. Tokyo Sci. Mus. 1: 37 (1939) (fig. 2C)
Basidioma pileado-estipitado. Píleo de 10-70 ×15-55 × 1-5 mm, suborbicular, reniforme a dimidiado, superficie lisa, laqueada y brillante, rojiza con tonos amarillo claro cuando joven (N60 Y90 M90), cuando maduro rojizo oscuro (N90 Y90 M90), con margen delgado. Contexto homogéneo, 4 mm de grosor, con bandas continuas de laca, de marrón a amarillo (N10 Y80 M50). Tubos 4 mm de longitud, concoloros al contexto. Poros circulares de 4 por mm. Disepimentos lisos, 35-40 μm de grosor, marrones (N10 Y40 M10), se manchan de marrón al maltratarse. Estípite negruzco, excéntrico, laqueado, semicilíndrico, tortuoso, 13-21 × 1-2 cm. Sistema hifal trimítico. Elementos cuticulares claviformes, hialinos, de pared gruesa con protuberancias laterales eventualmente, 35 × 10 μm (fig. 6E). Sistema hifal trimítico. Hifas generativas hialinas, 5 μm diam. Hifas esqueléticas del contexto, marrón claro, 5-7 μm diam. Hifas conectivas hialinas, lisas, 3.5-6.2 μm diam. Basidiosporas elipsoidales, marrón claro, truncadas en el ápice, 9-11 × 7-8 μm, Q = 1.33.
Resumen taxonómico
Material examinado. México, Tabasco, Macuspana, Parque Estatal Agua Blanca, selva mediana. 12/VIII/2011. Ruiz-Villareal (UJAT 3083); 12/VIII/2011. Fajardo-Alvarado (UJAT 3082). Tacotalpa, sierra Poaná, acahual. 20/VII/2007. Díaz-Contreras (UJAT 1810).
Distribución. Asia: China, Corea, Japón, Laos, Malasia, Myanmar, Taiwán (Hapuarachchi et al., 2019; Hattori y Ryvarden, 1994; Tan et al., 2015). Registro nuevo para el continente americano.
Comentarios taxonómicos
La especie pertenece al complejo G. lucidum, y se caracteriza por el basidioma rojizo-oscuro brillante en estado maduro tal como lo resaltan Hattori y Ryvarden (1994), Hsieh y Yeh (2004) e Imazeki et al. (1988). Los autores refieren el tamaño de las basidiosporas de 9.5-13 × 6-8 µm y un estípite delgado que puede llegar hasta 27 cm (21 cm en el ejemplar examinado). Puede confundirse con G. calidophilum J.D. Zhao, L.W. Hsu y X.Q. Zhang, pero difiere en el menor tamaño del estípite (13 cm) y mayor densidad de los poros (5-6/ mm).
Ganoderma oerstedii (Fr.) Torrend, Bull. Torrey bot. Club 29: 606 (1902) (fig. 2D)
Basidioma sésil. Píleo de 60-160 × 50-100 × 3 mm, leñoso, flabeliforme a semicircular, liso, laqueado, con una capa brillante que con el tiempo se vuelve quebradiza desprendiéndose de la cutícula, sulcado concéntricamente, marrón a negro hacia la base (N99 M80 C99) y rojizo hacia el margen (N90 Y80 M90), una vez perdida la capa brillante mantiene un tono marrón claro (N70 Y90 M60). Margen estéril, obtuso, glabro, ondulado, de blanco a amarillo claro. Contexto dúplex de 10 mm de grosor, con bandas continuas de depósitos de resina en color marrón; contexto de color rojizo hacia los tubos (N80 Y90 M60) y de marrón a amarillo claro hacia la parte superior (N60 Y80 M50). Tubos de 9 mm de longitud, marrón claro (Y80 M30 C10). Poros circulares a angulares, irregulares, 4-5 por mm, con tonos de marrón a rojizos al maltratarse (Y80 M40 C20). Disepimentos lisos, 29.42-52.89 μm de grosor. Sistema hifal trimítico. Elementos cuticulares clavados, 23-45 × 7-10 μm, escasamente con protuberancias laterales (fig. 6F). Sistema hifal trimítico. Hifas generativas de color rojizas al reaccionar con Melzer, dextrinoides, 3.53-5.75 μm diam. Hifas esqueléticas lisas, 8-11 μm diam. Hifas conectivas hialinas, 4-8 μm diam. Basidiosporas elipsoidales a oblongas, hialinas a subhialinas, truncadas en el ápice, 8.52-10.29 × 5.92-7.82 μm, Q = 1.36.
Resumen taxonómico
Material examinado. México, Tabasco, Centro, DACBIOL km 0.5 carretera Villahermosa-Cárdenas, acahual. 11/VIII/2004. Camacho (UJAT 4864); Parque Museo La Venta, selva mediana. 11/XI/1991. Rodríguez-Bahena (UJAT 599). Jalapa, Víctor Fernández Manero 1ra secc., acahual. 07/XI/2005. Camacho (UJAT 4850). Nacajuca, Bosques de Saloya, acahual. 19/X/2010. Damaris Cerón (UJAT 4991).
Distribución. América: Argentina, Brasil, Colombia, Costa Rica, Ecuador, Trinidad Tobago (Baltazar y Gibertoni, 2009; Gottlieb y Wright, 1999a; Ryvarden, 2000). Para México en los estados de Guerrero, Chiapas, Jalisco, Morelos, Oaxaca, Sinaloa, Sonora y Veracruz (López-Peña et al., 2016; Torres-Torres et al., 2015). Registro nuevo para Tabasco.
Comentarios taxonómicos
Se caracteriza por poseer una cutícula brillante en el píleo, misma que se vuelve quebradiza cuando viejo y tiende a perderla paulatinamente. La especie puede presentar ligeras variaciones en el tamaño de las basidiosporas, como en los registros de Gottlieb y Wright (1999a) (10-12 × 6-9 μm), Mendoza et al. (2011) (11-13 × 8-10 μm), Ryvarden (2000) (12-15 × 8-10 μm) y Torres-Torres et al. (2015) (10.4-13.6 × 7.2-8.8 µm), pero en el material estudiado (8.52-10.29 × 5.92-7.82 μm) son similares a lo descrito por López-Peña et al. (2016) (8.3-10 × 5.1-6.3 μm). Mendoza et al. (2011) consideraron que G. oerstedii es similar a G. nitidum Murrill pero se diferencian por presentar un menor tamaño de los poros (4-5/ mm).
Ganoderma orbiforme (Fr.) Ryvarden [as ‘orbiformum’], Mycologia 92(1): 187 (2000) (fig. 3A)
Basidioma pileado-estipitado. Píleo de 60 × 35 × 2 mm, flabeliforme a semicircular, laqueado brillante, color marrón oscuro (N99 M70 C99) a negro hacia la base y parte central, hacia el margen marrón claro (N80 Y70 M70) con tonos anaranjados, amarillos y blanco; al centro de la superficie presenta unas bandas concéntricas en color rojo claro. Margen estéril, liso, de blanco a amarillo (N30 Y70 M40). Contexto no completamente homogéneo de 10 mm de grosor, de marrón a amarillo claro (N30 Y70 M40) pero en su mayoría marrón claro (N60 Y70 M50), con escasos depósitos de resina. Tubos de 7 mm longitud, concoloros al contexto. Poros circulares a angulares de 4-6 por mm, lisos, amarillos cuando joven (Y80 M10 C10) tornándose marrón claro (Y70 M40 C10) al madurar. Disepimentos, 59-114 μm de grosor. Estípite, 5 × 1.5 cm., subcilíndrico, concoloro al píleo. Sistema hifal dimítico. Elementos cuticulares subcilíndricos, marrón claro, 20-25 × 7-13 μm (fig. 6G). Hifas esqueléticas lisas, 4-7 μm diam. Hifas conectivas muy ramificadas, 3-5 μm diam. Basidiosporas elipsoidales, truncadas en el ápice, marrón claro en KOH, 7-9 × 4-6 μm, Q = 1.6.
Resumen taxonómico
Material examinado. México, Tabasco, Cárdenas, Poblado C-29, Canacoite. 18/II/2009. Gómez-García (UJAT 4903). Centro, Parque Museo La Venta, selva mediana. 10/I/1988. Fucugauchi (UJAT 475); DACBIOL km 0.5 carretera Villahermosa-Cárdenas, acahual. 30/IX/2005. Martínez- Zarrazaga (UJAT 2272). Jalapa, Víctor Fernández Manero 2da secc., acahual. 05/I/2006. Camacho (UJAT 3281); 21/II/2005. Canal-Jiménez (UJAT 3187); pastizal. 08/VI/2006. Camacho (UJAT 3368). Macuspana, Parque Estatal Agua Blanca, selva mediana. 17/VI2008. Cappello GS (UJAT 384); 09/III/2011. Fajardo-Alvarado (UJAT 3110); Apasco, desviación a ranchería Chivalito, acahual. 09/III/2011. Fajardo-Alvarado (UJAT 3259). Nacajuca, cabecera municipal, vegetación urbana. 11/VII/2006. Tun (UJAT 3188). Tacotalpa, Estación sierra Poaná, selva mediana. 22/II/2003. Díaz-Contreras (UJAT 3086).
Distribución. América: Brasil, Asia: China, Filipinas, Guinea, India, Indonesia, Laos, Singapur y Tailandia (Bhosle et al., 2010; Gomes-Silva et al., 2011; Hapuarachchi et al., 2019; Ryvarden, 2000; Torres-Torres et al., 2012; Wang et al., 2014). Registro nuevo para México.
Comentarios taxonómicos
Ganoderma orbiforme se distingue por el basidioma rígido, píleo de débil a fuertemente laqueado, contexto de color marrón variable, basidiosporas elipsoidales a ovoides, y superficie de los poros amarillos cuando joven. La descripción es similar al registro de Ryvarden (2000), Torres-Torres et al. (2012) y Wang et al. (2014). Pudiera confundirse con G. tsugae, pero difiere en el tamaño de las basidiosporas (9-12 × 5.9-7.5 μm).
Ganoderma oregonense Murrill, N. Amer. Fl. (New York) 9(2): 119 (1908) (fig. 3B)
Basidioma pileado-subestipitado. Píleo de 130 × 70 × 5 mm, reniforme a flabeliforme, leñoso, contraído en la base, sulcado concéntricamente, superficie superior lisa, laqueado, color rojizo (N99 Y70 M90), presenta un surco definido hacia el margen. Margen estéril, obtuso, liso, marrón claro. Contexto no completamente homogéneo, de 7 mm de grosor, con depósitos de resina, tonos de color rojizo (N90 Y70 M80) y marrón claro (N70 Y80 M50). Tubos de 10 mm de longitud, marrón claro (Y60 M40 C20). Poros circulares a angulares, 3-4 por mm, marrones (Y60 M20 C10) sobre la superficie himenial. Disepimentos, 50-79 μm de grosor. Estípite, 5 × 4 cm subcilíndrico, concoloro al píleo, grueso. Sistema hifal trimítico. Elementos cuticulares, 31-54 × 7-10 μm (fig. 6H). Hifas generativas, 2.9-4.7 μm diam. Hifas esqueléticas marrón claro, 3.9-9.8 μm diam. Hifas conectivas ramificadas, 3.5-5.8 μm diam. Basidiosporas elipsoidales, subagudas en el ápice, subhialinas a marrón claro, 7.18-10.26 × 6-8 μm, Q = 1.24.
Resumen taxonómico
Material examinado. México, Tabasco, Cunduacán, R/a Pichucalco 1ra. secc., vegetación urbana. 18/IX/2008. Eduardo-Magaña (UJAT 5156). Macuspana, Parque Estatal Agua Blanca, selva mediana. 25/VIII/2011. Gallardo (UJAT 776); 11/XI/2019. García-Manuel (UJAT 5150). Nacajuca, cabecera municipal, acahual. 25/IX/2000. Cappello GS (UJAT 4973).
Distribución. América: Canadá, Estados Unidos. Asia: China (Hapuarachchi et al., 2019; Hong y Jung, 2004; Loyd et al., 2018; Murrill, 1908; Zhou et al., 2015). Para México en Hidalgo, Estado de México y Veracruz (Torres-Torres et al., 2015). Registro nuevo para Tabasco.
Comentarios taxonómicos
Las características del material estudiado coinciden con la descripción de Murrill (1908) y Torres-Torres et al. (2015). Puede confundirse con Ganoderma tsugae, pero ésta difiere por el tono anaranjado, amarillo y blanco en el margen, tubos de menor tamaño (5 mm de longitud) y disepimentos de menor grosor 24-39 μm.
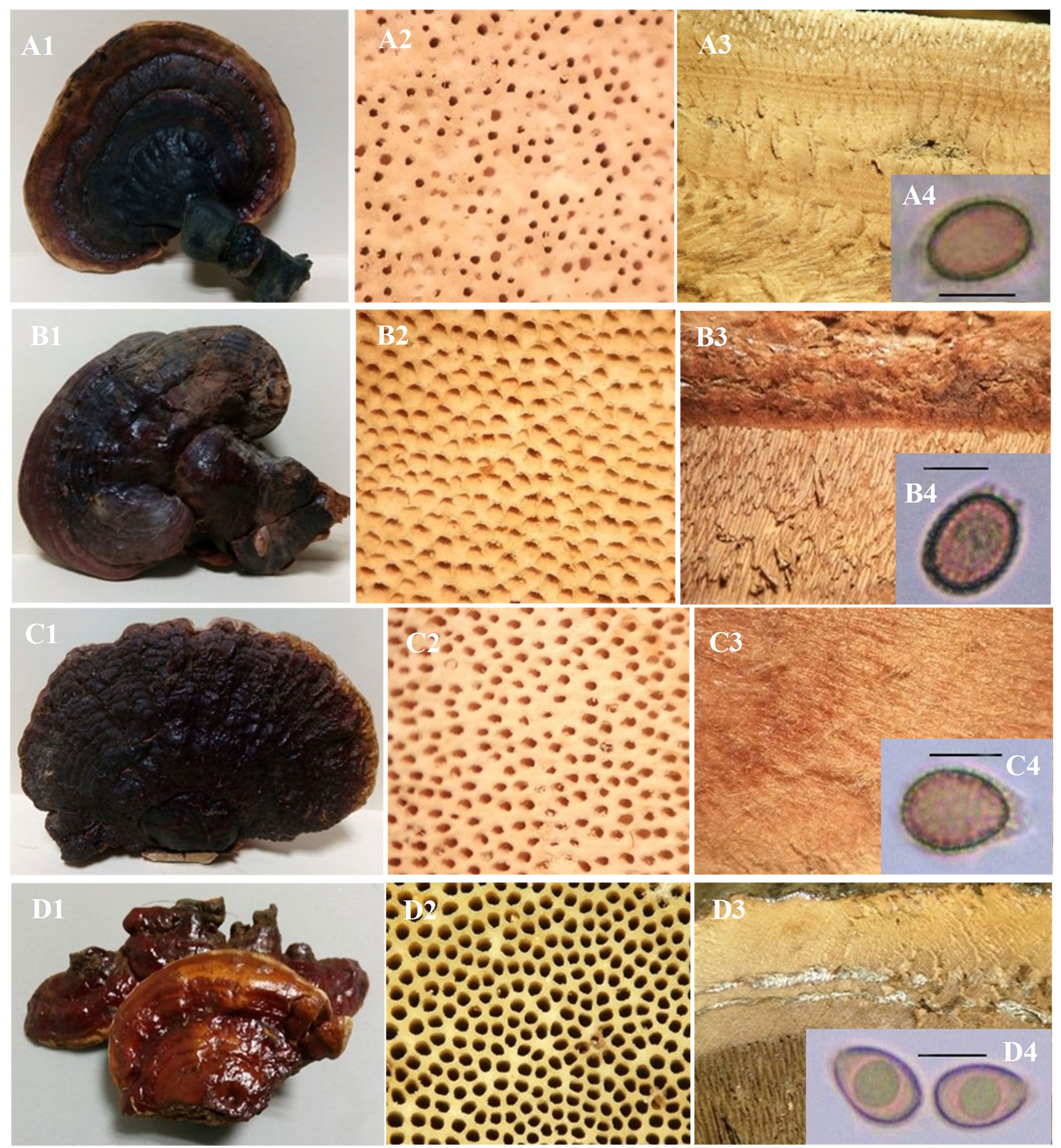
Ganoderma resinaceum Boud, in Patouillard, Bull. Soc. mycol. Fr. 5(2, 3): 72 (1889) (fig. 3C)
Basidioma pileado-estipitado. Píleo de 80-90 ×70-80 × 10 mm, semicircular a flabeliforme, esponjoso, superficie laqueada, sulcada, rojiza (N80 Y90 M90), con capas de resina en color anaranjado a marrón (N60 Y80 M60). Margen estéril medianamente grueso, obtuso, liso, concoloro al píleo. Contexto dúplex de 7 mm de grosor, marrón claro hacia la superficie cuticular (N30 Y80 M40) y marrón oscuro hacia los tubos (N80 Y80 M50). Tubos de 2-3 mm de longitud, de marrón a amarillo claro (N30 Y90 M40). Poros angulares a circulares, 4-5 por mm, lisos, marrones (N30 Y80 M40). Disepimentos lisos, 30.42-43.86 μm de grosor. Estípite, 5 × 3 cm, rojizo a negro (N80 Y90 M90), brillante, laqueado, claviforme, estrecho en la base y amplio hacia el ápice. Sistema hifal trimítico. Elementos cuticulares subcilíndricos a subclavados, de marrón claro a amarillos con tonos oscuros hacia el ápice, lisos, 27.15-28.8 × 7.11-8.38 μm (fig. 6I). Hifas generativas de hasta 3 μm diam. Hifas esqueléticas rara vez ramificadas, marrón claro, 6.07-8.43 μm diam. Hifas conectivas hialinas, ramificadas, 3.01-4.24 μm diam. Basidiosporas elipsoidales a ovoides, subhialinas, truncadas en el ápice, 8-10.5 × 6-7.5 μm, Q = 1.37.
Resumen taxonómico
Material examinado. México, Tabasco, Cárdenas, Hacienda Cacao km 10, selva alta. 22/VI/1987. Sol (UJAT 12). Centro, Villa Ocuiltzapotlan, vegetación urbana. 13/XII/1999. Marín-Romero (UJAT 1231); Recinto Memorial, Periférico s/n., vegetación urbana. 08/IX/2006. Tun (UJAT 404); Parque Museo La Venta, selva mediana. 18/I/2002. Isidro-Gómez (UJAT 51); Yumka’, selva mediana. 23/XI/2008. Osorio-Hernández (UJAT 5011); 22/IX/1988. Hernández-Trejo (UJAT 643); 25/IX/2008. Bautista (UJAT 4910); DACBiol km 0.5 carretera Villahermosa-Cárdenas, Tintal. 23/VIII/2019. García-Manuel (UJAT 4923). Comalcalco, Ejido Río Playa, cacaotal. 26/IX/1998. Ceferino-Contreras (UJAT 1196); vegetación urbana. 01/XI/2002. Díaz-Contreras (UJAT 1724). Huimanguillo, Agua Selva, selva alta. 26/V/2003. Becerra-Hernández (UJAT 1762); cabecera municipal, vegetación urbana. 23/IV/2019. García-Manuel (UJAT 4846); ranchería Paredón 1ra secc., vegetación urbana. 17/I/1988. Jiménez (UJAT 282). Jalapa, cabecera municipal, vegetación urbana. 21/II/2009. Luciano-De La Cruz (UJAT 3336); Víctor Fernández Manero 1ra secc., acahual. 21/II/2005. Verástegui-Hernández (UJAT 3225). Jalpa de Méndez, ranchería Chacalapa 1ra secc., cacaotal. 10/VIII/2014. López-Vázquez (UJAT 4902). Macuspana, Parque Estatal Agua Blanca, selva mediana. 04/XI/2009. Cappello GS (UJAT 4858); 04/VII/2019. Narvaez-Ilagor (UJAT 4886); 01/III/2012. Lara Palacio (UJAT 3175); Apasco, camino a ranchería Chivalito, acahual. 25/II/2011. Fajado-Alvarado (UJAT 3250); 17/X/2019. Carreño-Ruiz (UJAT 5105); ranchería San Antonio. 10/V/2007. García-Chable (UJAT 4854). Nacajuca, Bosques de Saloya, pastizal. 02/I/1997. Cappello GS (UJAT 1013); 27/VII/1999. Ceferino-Contreras (UJAT 1022); cabecera municipal, acahual. 15/XI/2002. Cappello GS (UJAT 2300); ranchería La Cruz 1ra secc., vegetación urbana. 17/IX/2008. Ovando-Hernández (UJAT 5152). Tacotalpa, Ejido Agua Blanca, sierra Tapijulapa, selva alta. 29/III/2003. Díaz-Contreras (UJAT 2017); 29/VI/2003. Díaz-Contreras (UJAT 2161); Oxolotán. 25/X/2008. González (UJAT 4909). Teapa, Monumento Natural Grutas de Cocona, selva alta. 20/VIII/2019. Domínguez González (UJAT 4920); ranchería San Antonio, cacaotal. 22/V/1999. Pérez de la cruz (UJAT 1152); Puyacatengo, acahual. 04/II/2005. Alejandra-Córdova (UJAT 4889).
Distribución. América: Brasil, Cuba, Estados Unidos, Venezuela África: Camerún, Congo, Togo, Uganda. Asia: China, India. Europa: Bélgica, Francia, Galia Central, Países Bajos (Ayissi y Mossebo, 2014; Bhosle et al., 2010; Cabarroi-Hernández et al., 2019; Gomes-Silva et al., 2011; Hapuarachchi et al., 2018; Kaur et al., 2018; Patouillard, 1889; Ryvarden, 2000; Steyaert, 1972; Torres-Torres et al., 2012; Zhou et al., 2015). Para México en los estados de Colima, Oaxaca y Tabasco (Ramos y Cappello, 2009; Raymundo y Valenzuela 2003; Torres-Torres et al., 2015).
Comentarios taxonómicos
Ganoderma resinaceum se distingue por el basidioma circular a flabeliforme, y por la presencia de laca brillante en la superficie pilear. Puede presentar ligeras variaciones en el tamaño de las basidiosporas, Kaur et al. (2018) las registraron de 8.5-11.5 × 5.5-6.8 μm muy similar a los ejemplares estudiados (8-10.5 × 5-7.5 μm), pero difiere de las descritas por Torres-Torres et al. (2012) de 11.2-12.5 × 6.5-7.4 μm. Se diferencia de G. curtisii, por la profundidad de los tubos de hasta 8 mm, mientras que G. resinaceum los presenta de 2-3 mm de profundidad.
Ganoderma sessiliforme Murrill, Bull. New York Bot. Gard. 8: 149 (1912) (fig. 3D)
Basidioma sésil. Píleo de 50-90 × 30-55 × 5 mm, de reniforme a semicircular, conchado, dimidiado, sulcado, laqueado brillante, corrugado, rojizo (N80 Y90 M90). Margen obtuso, estéril, liso, de blanco, marrón claro o amarillo (N80 Y80 M50). Contexto dúplex de 8 mm de grosor, con depósitos de resina marrón oscuro; color del contexto marrón con tonos amarillos (N80 Y80 M50) hacia la parte superior del píleo y marrón claro hacia la parte himenial. Tubos de 5 mm de longitud, marrones (N70 Y80 M40). Poros circulares a angulares, de 4-5 por mm, amarillos con tonos verdosos (Y50 M10 C10), se manchan de marrón al maltratarlos. Disepimentos lisos, 29.5-49.6 μm de grosor. Sistema hifal dimítico. Elementos cuticulares de subcilíndricos a clavados, marrón claro, lisos, 29.26-34.71 × 6.04-10.89 μm (fig. 6J). Hifas esqueléticas ramificadas, lisas, marrón claro, 4.16-7.12 μm diam. Hifas conectivas muy ramificadas, hialinas, 1.8-3.3 μm diam. Basidiosporas de elipsoidales a oblongas con el ápice truncado, marrón claro, 7.71-8.66 × 4.71-5.74 μm, Q = 1.56.
Resumen taxonómico
Material examinado. México, Tabasco, Centro, Yumka’, selva mediana. 22/X/1988. Hernández-Trejo (UJAT 588).
Distribución. América: Argentina, Brasil. Asia: India (Bhosle et al., 2010; Gottlieb y Wright, 1999a; Torres-Torres et al., 2012). Para México en los estados de Morelos y Sonora (López-Peña et al., 2016; Murrill, 1912; Torres-Torres et al., 2012, 2015). Registro nuevo para Tabasco.
Comentarios taxonómicos
Se distingue por un basidioma en forma de concha, laqueado brillante, por lo general en ejemplares jóvenes. Contexto con depósitos de resina marrón oscuro y basidiosporas pequeñas (Torres-Torres et al., 2012). Existen pocos registros de la especie tal como lo muestra la distribución, por lo que se considera relevante su hallazgo. Puede confundirse con G. oerstedii, pero difiere en el tamaño menor de las basidiosporas (7.71-8.66 × 4.71-5.74 μm).
Ganoderma sichuanense J.D. Zhao & X.Q. Zhang, in Zhao, Xu & Zhang, Acta Mycol. Sin. 2(3): 159 (1983) (fig. 4A)
Basidioma pileado-estipitado. Píleo de 215 × 120 × 2 mm, circular, dimidiado, cóncavo a convexo, consistencia leñosa, sulcado, rugoso, contraído hacia el estípite, laqueado brillante, rojizo (N90 Y80 M80) a marrón oscuro (N99 M80 C80), presenta capa fina de basidiosporas de color marrón. Margen estéril, liso, obtuso, delgado, concoloro al píleo. Contexto dúplex, esponjoso-fibroso, de 5 mm de grosor, con pequeños depósitos de resina, rojizo (N80 Y90 M60) hacia los tubos y marrón claro (N70 Y70 M50) hacia la superficie superior. Tubos de 10 mm de longitud, marrón claro (Y50 M50 C20). Poros suborbiculares de 4-5 por mm, concoloro a los tubos, se manchan de marrón oscuro al maltratarse. Disepimentos, 35-69 μm de grosor. Estípite, 5-7 × 2-3.5 cm, negruzco, central, laqueado, brillante, semicilíndrico. Sistema hifal trimítico. Elementos cuticulares subclavados a cilíndricos, marrón claro, 23-38 × 6-10 μm (fig. 6K). Hifas generativas no observadas. Hifas esqueléticas rara vez ramificadas, 4-7 μm diam. Hifas conectivas ramificadas, 2-5 μm diam. Basidiosporas elipsoidales a ovoides, truncadas en el ápice, marrones, 8.8-11.4 × 5.6-7.1 μm, Q = 1.59.
Resumen taxonómico
Material examinado. México, Tabasco, Nacajuca, ranchería Arroyo, selva mediana. 15/VIII/1989. González (UJAT 5072).
Distribución. Asia: China, Corea y Tailandia (Cao et al., 2012; Jargalmaa et al., 2017; Thawthong et al., 2017; Wang et al., 2012; Yao et al., 2013; Zhou et al., 2015. Registro nuevo para el continente americano.
Comentarios taxonómicos
Se caracteriza por presentar la superficie del píleo rugosa y la parte himenial de color amarillo claro a marrón claro. La descripción concuerda con Thawthong et al. (2017), Wang et al. (2012) y Yao et al. (2013). Wang et al. (2012) consideraron a G. sichuanense como G. lingzhi basado en secuencias nucleotídicas de ITS+IGS+RPB2, sin embargo, difieren en el color de la superficie de los poros, mientras que G. sichuanense los presenta amarillo pálido G. lingzhi los presenta amarillo brillante (azufre), así como la forma de las basidiosporas, en G. lingzhi son elipsoides toscamente equinuladas y con anillos cortos mientras que G. sichuanense de ovoides a elipsoides, finamente equinuladas.
Ganoderma stipitatum (Murrill) Murrill, N. Amer. Fl. (New York) 9(2): 122 (1908) (fig. 4B)
Basidioma pileado-estipitado. Píleo de 140 × 90 × 20 mm, dimidiado, aplanado, margen delgado, superficie lisa, laqueada, corrugada, marrón con tonos amarillos hacia el margen (N60 Y90 M50). Contexto dúplex, 12 mm de grosor, con bandas continuas de depósitos de resina, de marrón a rojizo (N20Y80 M30) hacia los tubos y marrón claro a amarillo claro (N20 Y70 M20) hacia el píleo. Tubos de 6 mm de longitud, marrones (N20 Y40 M20). Poros circulares de 4 por mm, marrón claro (N20 Y40 M10). Disepimentos, 34-41 μm de grosor. Estípite lateral, 4 × 2.5 cm, marrón, cilíndrico, laqueado, liso. Sistema hifal dimítico. Elementos cuticulares claviformes, lisos, de pared gruesa, 28-35 × 4-7 μm (fig. 6.L). Hifas generativas, 3-4 μm diam, septadas, subhialinas, con escasas fíbulas y ramificaciones. Hifas esqueléticas de hasta 7 μm diam, marrón claro. Basidiosporas elipsoidales, truncadas en el ápice, 8-10 × 5-7 μm, Q = 1.5.
Resumen taxonómico
Material examinado. México, Tabasco, Balancán, Ejido la Cuchilla, acahual. 10/X/2009. García-Hernández (UJAT 4912). Centro, DACBiol, km 0.5 carretera Villahermosa-Cárdenas, acahual. 28/II/2011. María de los A. Sánchez (UJAT 4993); Yumka’, selva mediana. 08/IX/2011. Hernández-García (UJAT 4847). Cunduacán, cabecera municipal, vegetación urbana. 08/IV/2019. García-Manuel (UJAT 4853).
Distribución. América: Bolivia, Brasil, Colombia, Costa Rica, Honduras, Nicaragua, Perú, Surinam, Venezuela. Asia: India (Bhosle et al., 2010; Cabarroi-Hernández et al., 2019; Gomes-Silva et al., 2011; Kaur et al., 2018; Murrill, 1908; Ryvarden, 2000; Steyaert, 1972; Welti y Courtecuisse, 2010). Registro nuevo para México.
Comentarios taxonómicos
Ganoderma resinaceum es similar a G. stipitatum, pero se diferencia por el menor tamaño del contexto (7 mm) y longitud en los tubos (2-3 mm). G. stipitatum se distingue por el basidioma laqueado, estípite lateral y bandas resinosas en el contexto. Se ha registrado con diferencias mínimas en el tamaño de las basidiosporas, Bhosle et al. (2010) de 7-10 × 6-8 µm, Ryvarden (2000) de 7-9.5 × 5-6.5 µm y Kaur et al. (2018) de 8.5-10 × 5.5-6.2 µm, esta última es similar a la del estudio (8-10 × 5-7 μm).
Ganoderma subresinosum (Murrill) C.J. Humphrey, Mycologia 30(3): 332 (1938) (fig. 4C)
Basidioma sésil. Píleo de 95-225 × 72-127 × 3 mm, semicircular a flabeliforme, laqueado, sulcado, corrugado radialmente sobre la superficie, color marrón oscuro a negro (N99 M80 C99). Margen de 3 mm, obtuso, estéril, ondulado, de marrón claro a anaranjado claro (Y50 M40 C20). Contexto dúplex, fibroso de 10 mm de grosor, rojizo (N70 Y80 M70) hacia los tubos y marrón claro (N60 Y80 M50) hacia la parte superior. Tubos de 15 mm de longitud, marrón claro (Y50 M40 C20). Poros de 4-5 por mm, circulares a irregulares, concoloro a los tubos sobre la superficie himenial. Disepimentos, 44-68 μm de grosor. Sistema hifal dimítico. Elementos cuticulares claviformes, marrón claro, 22-38 × 7-10 μm (fig. 6.M). Hifas esqueléticas rara vez ramificadas, marrón claro, 2.8-6 μm diam. Hifas conectivas ramificadas, 2-4 μm diam. Basidiosporas hialinas, elipsoidales a elongadas, 9-12 × 5.9-7 μm, Q = 1.6.
Resumen taxonómico
Material examinado. México, Tabasco, Cárdenas, Poblado C-29, acahual. 14/X/2008. Gómez-García (UJAT 4992). Centro, cabecera municipal, vegetación urbana. 27/X/2017. Sarai-Torres (UJAT 4980); Yumka’, selva mediana. 15/I/2005. Díaz-Contreras (UJAT 381). Comalcalco, ranchería Zapotal 2da secc., acahual. 06/X/2001. López-Bonilla (UJAT 3207). Cunduacán, cabecera municipal, acahual. 11/IV/1987. Sol-Sánchez (UJAT 4976). Jalapa, cabecera municipal, acahual. 22/I/2005. Díaz-Contreras (UJAT 4981); Víctor Fernández Manero 1ra secc., acahual. 22/I/2005. Díaz-Contreras (UJAT 3255); 21/II/2005. Bautista-M (UJAT 5111). Paraíso, cacaotal-potreritos. 05/II/1999. Álvarez-Gerónimo (UJAT 1396). Teapa, cabecera municipal. 24/IV/2007. Álvarez-Gerónimo (UJAT 4979).
Distribución. Asia: Birmania, China, Filipinas, India, Indonesia, Kenia, Laos, Malasia y Zaire (Hapuarachchi et al., 2019; Steyaert, 1972; Zohmangaiha et al., 2019). Registro nuevo para México y América.
Comentarios taxonómicos
Ganoderma subresinosum presenta basidiosporas hialinas, lisas y no truncadas en el ápice. Dicha especie fue registrada como Fomes subresinosus (Murrill, 1908), Trachyderma tsunodae (Imazeki, 1952), Magoderna subresinosus (Steyaert, 1972), y Amauroderma subresinosum (Corner, 1983), actualmente es reconocida como G. subresinosum en el Index Fungorum (www.indexfungorum/org/names/Names.asp) y descrita por Hapuarachchi et al. (2019).
Ganoderma tsugae Murrill, Bull. Torrey bot. Club 29: 601 (1902) (fig. 4D)
Basidioma pileado-estipitado. Píleo de 110 × 60 × 1 mm, flabeliforme a reniforme, sulcado concéntricamente, ligeramente corrugado, laqueado, superficie cubierta por una capa brillante y quebradiza, de color rojizo (N90 Y70 M99) hasta el estípite. Margen delgado ondulado, estéril, liso, con tonos de color anaranjado, amarillo y blanco (N50 Y70 M40). Contexto parcialmente homogéneo de 7 mm de grosor, con depósitos de resina en color marrón oscuro a marrón ámbar; color del contexto marrón claro a amarillo claro (N50 Y70 M40), se mancha de negro al reaccionar con KOH. Tubos de 5 mm de longitud, marrón (N50 Y90 M60). Poros angulares, irregulares, escabrosos, de 4-5 por mm, marrones (N50 Y80 M40). Disepimentos, 24- 39 μm de grosor. Estípite, 2.5 × 2 cm, subcilíndrico, corto, excéntrico, concoloro al píleo. Sistema hifal trimítico. Elementos cuticulares subcilíndricos a subclavados, subhialinos, 24-40 × 9-12 μm (fig. 6N). Hifas generativas hialinas, 5 μm diam. Hifas esqueléticas lisas, subhialinas, ramificadas, 6-7 μm diam. Hifas conectivas hialinas, ramificadas, 3 μm diam. Basidiosporas elipsoidales, hialinas, truncadas en el ápice, 9-12 × 5.9-7.5 μm, Q = 1.56.
Resumen taxonómico
Material examinado. México, Tabasco, Cárdenas, Poblado C-29, cacaotal. 09/III/2009. Olmedo-López (UJAT 4868). Centro, Parrilla II, vegetación urbana. 19/IX/2010. González-González (UJAT 4994). Jalapa, Víctor Fernández Manero 2da secc., selva mediana. 14/VIII/2004. Camacho (UJAT 278). Macuspana, Parque Estatal Agua Blanca, selva mediana. 29/III/2003. Díaz-Contreras (UJAT 2019); Poblado Límbano Blandín, acahual. 08/VIII/2006. García-Manuel (UJAT 4905); 04/VII/2019. Cappello GS (UJAT 4988).
Distribución. América: Canadá, Estados Unidos. Asia: China, Corea, Japón y Taiwán (Cao et al., 2012, Hapuarachchi et al., 2019; Hong y Jung, 2004; Loyd et al., 2017; Moncalvo, 2000; Moncalvo et al., 1995; Murrill, 1902; Zhou et al., 2015). Para México en Durango, Estado de México, Hidalgo (Bandala et al., 1993). Registro nuevo para Tabasco.
Comentarios taxonómicos
Se distingue por el píleo flabeliforme en color rojizo brillante a ligeramente oscuro. G. tsugae se ha considerado sinónimo de G. lucidum (Haddow, 1931; Steyaert, 1977) pero Zhou et al. (2015) concluyeron que es independiente de G. lucidum basado en secuencias nucleotídicas de ITS+TEF1+RPB, morfológicamente en lo que se diferencian es en el tipo de esporas. G. lucidum presenta esporas elipsoides a piriformes, equinuladas con crestas sinuosas mientras que G. tsugae son elipsoides y moderadamente equinuladas y sin crestas.
Ganoderma zonatum Murrill, Bull. Torrey bot. Club 29: 606 (1902) (fig. 4E)
Basidioma sésil. Píleo de 240 × 160 × 6 mm, semicircular, leñoso, dimidiado, zonado concéntricamente, superficie superior lisa, laqueado brillante perdiéndose al envejecer; color rojizo a oscuro (N99 Y80 M70) hacia la base donde se ubica la zona vieja y color anaranjado intenso (N60 Y80 M70) hacia el margen donde se ubica la zona joven. Margen estéril, obtuso, liso, blanco cuando fresco tornándose amarillo al secarse. Contexto homogéneo, hasta 10 mm de grosor, con pequeños depósitos de resina, zonado, marrón (N90 Y80 M70). Tubos hasta 10 mm de longitud, estratificados, concoloro al contexto. Poros circulares, 4 por mm, de blancos a amarillos sobre la superficie himenial (Y99 M30 C10), se manchan de marrón al maltratarse. Disepimentos, 99-142 μm de grosor. Sistema hifal dimítico. Elementos cuticulares clavados a cilíndricos, 41-54 × 10-16 μm con protuberancias o ramificaciones (fig. 6P). Hifas generativas, 2-4.9 μm diam, pared delgada. Hifas esqueléticas ocasionalmente ramificadas, marrón claro, 3.3-6.8 μm diam. Basidiosporas elipsoidales a oblongas, truncadas en el ápice, marrón claro, 8.6-10 × 6-7 μm, Q = 1.43.
Resumen taxonómico
Material examinado. México, Tabasco, Centro, DACBiol, km 0.5 carretera Villahermosa-Cárdenas, acahual. 25/VIII/2019. García-Manuel (UJAT 4978); Estrellas de Buenavista, vegetación urbana. 23/IX/2019. García-Manuel (UJAT 5004). Paraíso, Ejido las Flores, vegetación urbana. 05/IX/2019. García-Manuel (UJAT 5151).
Distribución. América: Brasil, Estados Unidos. Asia: China, India (Gomes-Silva et al., 2011; Gottlieb y Wright 1999a; Hapuarachchi et al., 2019; Kaur et al., 2018; Loyd et al., 2018; Murrill, 1902; Ryvarden, 2000; Zhou et al., 2015). Para México en los estados de Baja California Sur, Puebla, Jalisco (Guzmán, 1972; Torres-Torres et al., 2015). Registro nuevo para Tabasco.
Comentarios taxonómicos
Se caracteriza al presentar zonas estratificadas diferenciadas. La descripción es similar al registro de Murrill (1902), Ryvarden (2000) y Torres-Torres et al. (2015). La especie pertenece al complejo G. lucidum. Pudiera confundirse con G. resinaceum, pero este se diferencia porque presenta contexto dúplex y la longitud de los tubos es menor (2-3 mm).
Subgénero Elfvingia
Ganoderma applanatum (Pers.) Pat., Hyménomyc. Eur. (Paris): 143 (1887) (fig. 5A)
Basidioma sésil. Píleo de 50-200 × 40-100 × 5 mm, leñoso, no laqueado, semicircular, marrón (N80 Y80 M50), con una fina capa de basidiosporas en color marrón; margen estéril, marrón claro al secarse. Contexto dúplex de 20 mm de grosor sin depósitos de resina, rojizo (N80 Y80 M60) hacia los tubos y marrón claro (N60 Y90 M60) hacia el ápice. Tubos de hasta 10 mm de longitud, marrón (Y60 M30 C10). Poros circulares a angulares, 5 por mm, concoloro a los tubos. Basidiosporas elipsoidales, subhialinas a marrón claro, 6-7 × 5-6 μm, Q = 1.2.
Resumen taxonómico
Material examinado. México, Tabasco, Centro, Yumka’, selva mediana. 28/VII/1988. Hernández-Trejo (UJAT 1232); DACBiol km 0.5 carretera Villahermosa-Cárdenas, acahual. 16/I/2004. Sosa (UJAT 1567); carretera Apasco, camino a ranchería Chivalito, selva mediana subperenifolia. 25/II/2011. Fajardo-Alvarado (UJAT 3210); fraccionamiento Bonanza, vegetación urbana. 18/X/2010. Coronel-Díaz (UJAT 4914); 12/XI/2010. Hernández-Sánchez (UJAT 4913). Comalcalco, ranchería Nicolás Bravo 3ra secc., cacaotal. 11/III/2008. López-Pérez (UJAT 4859); 11/III/2008. Morales- López (UJAT 4848); Ejido Río Playa, huerto familiar. 05/IX/1999. Cappello GS (UJAT 1092); ranchería Zapotal 2da secc., cacaotal. 20/XII/1998. Marín-Romero (UJAT 1258); acahual. 01/XI/2002. Cappello GS (UJAT 2253); Poblado Chichicapa, selva mediana. 11/III/2008. Delfín-Aguilar (UJAT 5071). Cunduacán, Fraccionamiento San Antonio, acahual. 19/VII/2003. Díaz-Contreras (UJAT 2176). Huimanguillo, Puente Mosquitero km 13, selva alta. 15/VI/1991. Hernández-Trejo (UJAT 600); Malpasito, acahual. 27/IX/2008. Valera (UJAT 3041). Jalapa, Víctor Fernández Manero 1ra secc., acahual. 21/II/2005. De la Cruz (UJAT 264); 21/II/2005. Camacho (UJAT 4855); 21/II/2005. Cruz Hernández (UJAT 1665); 14/VIII/2004. Márquez (UJAT 4982). Macuspana, Parque Estatal Agua Blanca, selva mediana. 24/II/2012. Peláez Hernández (UJAT 4223); 24/II/2011. Cruz Morales (UJAT 1662); 18/IX/2019. García-Manuel (UJAT 5103); 15/I/2011. Fajardo-Alvarado (UJAT 4849); 08/IX/2011. Cristian (UJAT 4856); 20/IV/2010. López-De la Cruz (UJAT 3262); 05/IX/2011. Fajardo-Alvarado (UJAT 3173); 04/VIII/2011. Ruíz Emanuel (UJAT 3228); 14/VII/2010. Aldana (UJAT 3248); 24/II/2011. Yañes (UJAT 3172); 17/VI/2008. Cappello GS (UJAT 4900); 24/II/2012. Valencia-Sánchez (UJAT 4916); 24/V/2014. Cappello GS (UJAT 3995); 19/I/2012. Fajardo-Alvarado (UJAT 3116); 27/III/2007. De la Cruz-Rodríguez (UJAT 3134); 27/III/2007. Mora-Federico (UJAT 3146); 20/IV/2010. Arias-Cruz (UJAT 4996); 18/X/2019. García-Manuel (UJAT 5148); 08/IX/2011. Sánchez Cornelio (UJAT 4974); 23/IV/2010. Gómez-García (UJAT 4975); 06/VII/2011. Ávalos- Lázaro (UJAT 4983); 25/II/2011. Fajardo-Alvarado (UJAT 3117); carretera Apasco, selva mediana subperennifolia. 25/II/2011. Cruz-V (UJAT 3108). Nacajuca, ranchería Belén, acahual. 23/XI/1997. Concha-Frías (UJAT 1387); cabecera municipal, vegetación urbana. 04/IX/2018. Cappello GS (UJAT 4907). Tacotalpa, Ejido el Madrigal 5a secc., acahual. 06/IV/2003. Díaz-Contreras (UJAT 2042); Estación Biológica la Florida, Tapijulapa, acahual. 28/VIII/2018. Gómez-Morales (UJAT 4977). Teapa, Puyacatengo, cacaotal. 20/VI/1987. Santiago (UJAT 54). Tenosique, Rancho el Tigre, selva alta. 04/XII/1997. Córdova (UJAT 1172).
Distribución. América: Brasil, Estados Unidos. África: Kenia. Asia: Birmania, China, Corea, India, Japón. Europa: Francia, Reino Unido (Bhosle et al., 2010; Hapuarachchi et al., 2019; Hong y Jung, 2004; Jargalmaa et al., 2017; Kaur et al., 2017; Patouillard, 1889; Singh et al., 2014; Steyaert, 1972; Torres-Torres et al., 2012). Para México en los estados de Guanajuato, Nuevo León, Oaxaca, Sonora y Tabasco (Cappello, 2006; Castillo y Guzmán, 1970; López-Peña et al., 2016; Ojeda-López et al., 1986; Raymundo y Valenzuela, 2003).

Comentarios taxonómicos
Puede confundirse con G. australe, pero difiere por la presencia de bandas de resina en el contexto y el tamaño mayor de las basidiosporas (8-9.2 × 5-6 μm) (López-Peña et al., 2016; Torres-Torres et al., 2012). Debido a sus frecuentes registros se presentan las características principales, y para una descripción detallada se sugiere consultar a Bhosle et al. (2010), Hapuarachchi et al. (2019), Kaur et al. (2017), López-Peña et al. (2016), Torres-Torres et al. (2012).
Ganoderma australe (Fr.) Pat., Bull. Soc. mycol. Fr. 5(2,3): 65 (1889) (fig. 5B)
Basidioma sésil. Píleo de 200 × 160 × 7 mm, leñoso, aplanado, no laqueado, semicircular, ligeramente hinchado en el centro, sulcado concéntricamente, marrón oscuro (N99 Y80 M40), presenta una capa fina de basidiosporas sobre la superficie y una capa quebradiza en forma de costra. Margen estéril, obtuso, liso, concoloro al píleo. Contexto dúplex de 5 mm de grosor, con bandas de depósitos de resina, color rojizo a oscuro (N99 Y80 M50) hacia los tubos y marrón claro (N90 Y80 M70) hacia el ápice. Tubos estratificados de 12 mm de longitud, marrón claro (Y60 M20 C10). Poros circulares de 4-5 por mm, lisos, de blancos a amarillos sobre la superficie himenial (Y50 M30 C10). Disepimentos, 25.31-54.1 μm de grosor. Sistema hifal trimítico. Elementos cuticulares subclavados, 20-30 × 8-10 μm (fig. 6O). Hifas generativas no observadas. Hifas esqueléticas en ocasiones ramificadas, marrón claro, 4.15-6.08 μm diam. Hifas conectivas ramificadas, marrón claro, 2.05-3.35 μm diam. Basidiosporas elipsoidales, subhialinas a marrón claro, 8-9.2 × 5-6 μm, Q = 1.55.
Resumen taxonómico
Material examinado. México, Tabasco, Macuspana, Parque Estatal Agua Blanca, selva mediana. 27/III/2007. Álvaro-Torres (UJAT 3214); 18/III/2006. Tun (UJAT 5109); 11/XI/1988. González (UJAT 3269); 01/XI/2012. Carreño-Ruíz (UJAT 5107); 13/IX/1999. Cappello-GS (UJAT 5153); 01/X/2012. Carreño-Ruíz (UJAT 3115); 08/IX/2011. Martha Elena Sánchez C. (UJAT 4974). 27/III/2007. Hernández-Paola (UJAT 3150). Tenosique, ranchería Santo Tomás, selva alta. 05/XII/1987. Sánchez-Ruíz (UJAT 375).
Distribución. América: Argentina, Brasil, Canadá, Colombia, Cuba, Venezuela. África: Sudáfrica. Asia: Birmania, China, India, Niza, Nueva Caledonia, Tailandia, Taiwán. Oceanía: Australia, Nueva Zelanda. Europa: Suiza (Gomes-Silva et al., 2011; Hapuarachchi et al., 2019; Kaur et al., 2017; Moncalvo et al., 1995; Patouillard, 1889; Pinzón-Osorio y Pinzón-Osorio, 2016; Tchotet Tchoumi et al., 2019; Torres-Torres et al., 2012; Welti y Courtecuisse, 2010). Para México en el estado de Guanajuato, Nayarit y Veracruz (Guzmán, 1972; Ojeda-López et al., 1986). Registro nuevo para Tabasco.
Comentarios taxonómicos
Puede confundirse con G. applanatum, pero este difiere al poseer basidiosporas de menor tamaño (6-7 × 5-6 μm) y sin depósitos de resina en el contexto. El material estudiado concuerda con la descripción de Gottlieb y Wright (1999b), Kaur et al. (2017) y Torres-Torres et al. (2012).
Ganoderma lipsiense (Batsch) G.F. Atk., Annls mycol. 6(3): 189 (1908) (fig. 5C)
Basidioma sésil. Píleo de 89-1500 × 67-142 × 5 mm, leñoso, no laqueado, aplanado, semicircular a reniforme, sulcado hacia el centro, superficie rugosa, marrón (N80 Y80 M50), presenta una fina capa de basidiosporas marrones. Margen estéril, lobulado, obtuso, liso, blanco cuando fresco tornándose de marrón claro a amarillo al secarse. Contexto dúplex de 7 mm de grosor, con depósitos de resina, rojizo (N80 Y80 M60) hacia los tubos y marrón claro (N60 Y90 M60) hacia el ápice. Tubos de 10 mm de longitud, marrón claro (N30 Y40 M30). Poros circulares a angulares, 5 por mm, concoloros a los tubos. Disepimentos, 26.28-90 μm de grosor. Sistema hifal trimítico. Elementos cuticulares subcilíndricos a clavados, 19 × 6.7 μm (fig. 6Q). Hifas generativas no observadas. Hifas esqueléticas rara vez ramificadas, marrón claro, 6.24-9 μm diam. Hifas conectivas, 5.5-5.8 μm diam. Basidiosporas elipsoidales, de subhialinas a marrón claro, 5-10.6 × 5-6 μm, Q = 1.41.
Resumen taxonómico
Material examinado. México, Tabasco, Centro, Parque Museo La Venta, selva mediana. 17/IX/1998. Marín-Romero (UJAT 1178). Comalcalco, ranchería Nicolás Bravo 3ra secc., cacaotal. 16/I/2002. López-Bonilla (UJAT 1393); ranchería Zapotal 2ª secc., cacaotal. 24/I/1998. López-Bonilla (UJAT 4986). Cunduacán, ranchería Pichucalco 1ra secc., acahual. 18/IX/2008. Magaña (UJAT 4997). Jalapa, Víctor Fernández Manero 1ra secc., acahual. 22/I/2005. Díaz-Contreras (UJAT 4989); 14/VIII/2004. Camacho (UJAT 4995). Macuspana, Parque Estatal Agua Blanca, selva mediana. 10/VI/2010. Aldana RJ (UJAT 3019); 23/VI/2014. Gómez- García (UJAT 4016); 23/VI/2014. Heidy (UJAT 5001). Tacotalpa, sierra Tapijulapa, acahual. 01/III/2011. León Hernández (UJAT 4924); Villa Luz, selva mediana. 22/I/2006. Cappello GS (UJAT 3256); 18/VIII/2004. Camacho (UJAT 2329); Ejido el Madrigal 5a secc., selva mediana. 06/IV/2003. Díaz-Contreras (UJAT 2041); 24/VIII/2003. Díaz-Contreras (UJAT 2174); Ejido Pomoca, acahual. 31/V/2003. Díaz-Contreras (UJAT 2187); Ejido Cerro Blanco 5ta secc., acahual. 07/VI/200. Díaz-Contreras (UJAT 2119); Ejido La Pila., acahual. 15/III/2003. Díaz-Contreras (UJAT 2332); Ejido Santa Rosa 1ra secc., acahual. 18/XI/2014. De la Cruz-Lorena (UJAT 4990); Estación Biológica la Florida, Tapijulapa, acahual. 22/VIII/2012. Fajardo-Alvarado (UJAT 4987). Teapa, ranchería San Antonio, cacaotal. 22/V/1999. Pérez de la Cruz (UJAT 1145); Puyacatengo, acahual. 04/III/2005. Díaz-Contreras (UJAT 146).
Distribución. América: Estados Unidos. Asia: India (Bhosle et al., 2010; Gottlieb y Wright, 1999b; Kaur et al., 2018). Registro nuevo para México.
Comentarios taxonómicos
La descripción de los ejemplares es similar al registro de Bhosle et al. (2010), Gottlieb y Wright (1999b) y Kaur et al. (2018). Se distingue por una cutícula quebradiza en el píleo. Puede confundirse con G. applanatum, pero difiere en el contexto por la ausencia de resinas y el tamaño mayor de las basidiosporas, 7-10 × 4-6 μm.
Ganoderma philippii (Bres. & Henn. ex Sacc.) Bres., Iconogr. Mycol. 21: tab. 1014 (1932) (fig. 5D)
Basidioma sésil. Píleo de 110-140 × 80-120 × 5 mm, no laqueado, de circular a semicircular o espatulado, subaplanado, tuberculado, dimidiado, superficie quebradiza y semejante a una placodermis, ligeramente rugoso a escrobiculado, sulcado, marrón (N80 Y90 M70), hacia el margen presenta una banda con tono anaranjado y amarilla (N00 Y90 M20). Margen lobulado, estéril, color blanco a marrón. Contexto homogéneo de 15 mm de grosor, con depósitos de resina, rojizo (N80 Y90 M80). Tubos de 10 mm de longitud marrones (Y80 M50 C50). Poros circulares a angulares de 4-5 por mm, marrón claro con tonos amarillos (Y80 M40 C40), se manchan de marrón a negro al maltratarse. Disepimentos, 33-37 µm de grosor. Sistema hifal trimítico. Elementos cuticulares no observados. Hifas generativas, 5-5.5 µm diam. Hifas esqueléticas marrones, 4-6.5 µm diam. Hifas conectivas rara vez ramificadas, 1.3-3.8 µm diam. Basidiosporas ovoides, equinuladas, 6-7.7 × 4-5 µm, Q = 1.52.
Resumen taxonómico
Material examinado. México, Tabasco, Cárdenas, cabecera municipal, acahual. 14/X/2008. Abdel-Olmedo (UJAT 4875). Centla, Ejido la Sabana, selva baja caducifolia. 09/XI/2018. García-De la Cruz (UJAT 4906). Centro, Yumka’, selva mediana. 03/X/2000. Pérez-Naranjo (UJAT 4866). Comalcalco, ranchería Zapotal 2da secc., cacaotal. 19/XI/2002. López-Bonilla (UJAT 4871); ranchería Nicolás Bravo 3ra secc., cacaotal. 11/III/2008. Pérez Montejo (UJAT 4400). Huimanguillo, Ejido Villa Guadalupe Cerro de las Flores, bosque mesófilo de montaña. 23/IX/1998. López-Bonilla (UJAT 1177); Malpasito, acahual. 22/IX/2003. Pérez-Cruz (UJAT 4865); Agua Selva, acahual. 18/XI/2002. Domínguez-G (UJAT 2327). Macuspana, Parque Estatal Agua Blanca, selva mediana. 04/VII/2019. Rueda-López (UJAT 4857); 24/II/201. Hernández-Lázaro (UJAT 3111); 19/V/2005. Cappello GS (UJAT 4872); 08/III/2019. Sánchez-García (UJAT 4867); 17/VI/2008. Cappello GS (UJAT 3252); 14/IX/2008. Cappello GS (UJAT 3135); 20/IV/2010. Rodríguez-Guzmán (UJAT 3166); 25/VI/2010. Bautista (UJAT 3245); 04/VII/2019. Cappello GS (UJAT 5003); 11/XI/2002. Macías-Valadez (UJAT 3171); 12/VIII/2011. Avalos-Lázaro (UJAT 3232); 08/IX/2011. Manríquez-Muñoz (UJAT 4904); 18/III/2006. Camacho (UJAT 4869); carretera Caparroso 1ra secc., acahual. 10/IX/2010. Aldana (UJAT 3136); Apasco, carretera a Ejido Chivalito, acahual. 11/VI/2012. Carreño-Ruíz (UJAT 4876). Paraíso, cacaotal-potreritos. 03/X/2001. López Bonilla (UJAT 1394); 19/XI/2002. López Bonilla (UJAT 2267); Tacotalpa, Centro Ecoturístico Yu-Balcah, acahual. 29/IV/2005. Sánchez-Torres (UJAT 4870).
Distribución. América: Brasil. África: Congo. Asia: Birmania, China, India, Indonesia, Malasia, Myanmar, Singapur, Vietnam. Oceanía: Nueva Guinea (Bhosle et al., 2010; Gottlieb y Wright, 1999b; Hapuarachchi et al., 2019; Hood, 2006; Steyaert, 1972). Registro nuevo para México.
Comentarios taxonómicos
Ganoderma philippii se distingue al presentar una superficie del píleo quebradiza y semejante a una placodermis, tal como lo menciona Bhosle et al. (2010), Gottlieb y Wright (1999b) y Hood (2006). Además de dichas características los ejemplares estudiados poseen hacia el margen una banda anaranjada y amarillenta, que lo diferencia del resto de las especies del subgénero Elfvingia.
Discusión
El número de especies aquí descritas (21 taxones) y los nuevos registros para México (tabla 1) son un reflejo de la diversidad del género Ganoderma presente en los ecosistemas tropicales (Gilbertson y Ryvarden, 1986; Torres-Torres et al., 2012; Zhao, 1989). Moncalvo y Ryvarden (1997) destacaron que el subgénero Ganoderma está mejor representado que el subgénero Elfvingia, lo que coincide con este estudio (17 y 4 spp., respectivamente). Del total de especies, solo 4 son cosmopolitas y una neotropical, se resalta que 16 tienen una distribución pantropical (77%) compartidas con América del Sur y Asia. Los nuevos registros para el continente americano (5 especies) amplían la distribución pantropical del género Ganoderma. A nivel estatal las especies que tienen una amplia distribución son G. applanatum y G. resinaceum ya que se encontraron en 10 municipios, seguidas por G. philippii en 8 municipios; G. multiplicatum y G. subresinosum en 7. Con una distribución restringida a 1 o 2 municipios fueron: G. australe, G. multipileum, G. neojaponicum, G. sessiliforme, G. sichuanense y G. zonatum.
La plasticidad morfológica del género Ganoderma dificulta la determinación de las especies; sin embargo, las estructuras macro y micro morfológicas resultaron relevantes en la identificación de los ejemplares tal como lo propone Torres-Torres et al. (2012). Un ejemplo de ello son los basidiomas de G. colossus, G. multipileum, G. multiplicatum, G. orbiforme, G. oregonense y G. stipitatum, los cuales se habían descrito originalmente como sésiles, y en el presente estudio se registraron tanto sésiles como estipitados (Gottlieb y Wright, 1999a; Hapuarachchi et al., 2019; Murrill, 1908; Parihar et al., 2013; Ryvarden, 2000; Torres-Torres et al., 2012, 2015; Wang et al., 2009, 2014; Zohmangaiha et al., 2019).
Entre las características macromorfológicas la presencia de depósitos de resina en el contexto permitió la separación de los taxones dentro del subgénero Elfvingia, pese a que Steyaert (1967) mencionó la poca relevancia de estas estructuras. A pesar de que G. applanatum, G. australe y G. lipsiense poseen contexto dúplex se diferencian por la presencia o ausencia de depósitos de resina, y cuando están presentes por el tipo del mismo, como por ejemplo G. australe contiene bandas continuas mientras que G. lipsiense los presenta como incrustaciones. Para México el registro de Ganoderma era de 25 especies (López-Peña et al., 2016; Raymundo y Valenzuela, 2003; Torres-Torres et al., 2015), con el presente estudio se incrementa a 36, lo que representa un aporte de 44% y se amplía el rango de distribución del género.
Clave dicotómica para las especies de Ganoderma en Tabasco.
1a Basidioma laqueado (subgénero Ganoderma)…………………………………… 2
1b Basidioma no laqueado (subgénero Elfvingia)………………… 18
2a Basidioma estipitado………………… 3
2b Basidioma sésil………………… 11
3a Basidioma con estípite cilíndrico, delgado de 3-5 mm de grosor………………… G. neojaponicum
3b Basidioma con estípite con grosor mayor a 10 mm………………… 4
4a Con píleo de margen delgado menor a un mm………………… 5
4b Con píleo de margen grueso de un mm en adelante………………… 6
5a Contexto homogéneo y 7 mm de grosor………………… G. oregonense
5b Contexto dúplex de 5 mm de grosor………………… G. sichuanense
6a Consistencia corchosa, píleo totalmente amarillo………………… G. colossus
6b Consistencia leñosa………………… 7
7a Contexto dúplex y con bandas de resina………………… 8
7b Contexto parcialmente homogéneo………………… 10
8a Contexto de 12 mm de grosor………………… G. stipitatum
8b Contexto menor a 12 mm de grosor………………… 9
9a Tubos de 8 mm de longitud………………… G. curtisii
9b Tubos de 2-3 mm de longitud………………… G. resinaceum
10a Basidiosporas de 7-9 × 4-6 μm………………… G. orbiforme
10b Basidiosporas de 9-12 × 6-7.5 μm………………… G. tsugae
11a Basidioma con píleo único………………… 12
11b Basidioma con píleo múltiple………………… G. multipileum
12a Basidioma con píleo estratificado en varias capas o zonas………………… G. zonatum
12b Basidioma con píleo no estratificado………………… 13
13a Con tubos de 15 mm de longitud………………… G. subresinosum
13b Con tubos menos de 15 mm………………… 14
14a Contexto grueso de 20 mm o más………………… G. multiplicatum
14b Contexto menor a 20 mm………………… 15
15a Píleo carnoso………………… G. carnosum
15b Píleo no carnoso………………… 16
16a Disepimentos de 46-120 μm de grosor………………… G. dunense
16b Disepimentos de 29-53 μm de grosor………………… 17
17a Basidiosporas de 8-10 × 5-8 μm………………… G. oerstedii
17b Basidiosporas de 7-8 × 4-6 μm………………… G. sessiliforme
18a Píleo corrugado………………… G. philippii
18b Píleo liso………………… 19
19a Contexto sin depósitos de resina………………… G. applanatum
19b Contexto con depósitos de resina………………… 20
20a Elementos cuticulares mayor a 20 μm de longitud………………… G. australe
20b Elementos cuticulares menor a 20 μm de longitud………………… G. lipsiense
Agradecimientos
Los autores agradecen el esfuerzo en conjunto para la realización del estudio, así mismo al espacio brindado por la División Académica de Ciencias Biológicas de la UJAT, en atención especial al laboratorio de Micología y al Herbario UJAT.
Referencias
Ayala, N. y Guzmán, G. (1984). Los hongos de la península de Baja California I. Las especies conocidas. Revista Mexicana de Micología, 4, 25–42. https://doi.org/10.33885/sf.1984.2.592
Ayissi, B. M. K. y Mossebo, D. C. (2014). Some noteworthy taxonomic variations in the complex wood-decayer Ganoderma resinaceum (Basidiomycota) with reference to collections from tropical Africa. Kew Bulletin, 69, 1–14. http://dx.doi.org/10.1007/s12225-014-9542-9
Baltazar, J. M. y Gibertoni, T. B. (2009). A checklist of aphyllophoroid fungi (Basidiomycota) recorded from the Brazilian Atlantic rain forest. Mycotaxon, 109, 439–442. http://dx.doi.org/10.5248/109.439
Bandala, V. M., Guzmán, G. y Montoya, L. (1993). Los hongos del grupo de los poliporáceos conocidos en México (Eds.). Contribuciones micológicas en homenaje al biólogo José Castillo Tovar por su labor en pro de la micología mexicana (pp. 1–55). Reporte científico Núm. Especial, 13. Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León.
Bas, C. (1969). Morphology and subdivision of Amanita and a monograph on its section Lepidella. Persoonia, 5, 285–579.
Bhosle, S., Ranadive, K., Bapat, G., Garad, S., Deshpande, G. y Vaidya, J. (2010). Taxonomy and diversity of Ganoderma from the western parts of Maharashtra (India). Mycosphere, 1, 249–262.
Cabarroi-Hernández, M., Villalobos-Arámbula, A. R., Torres-Torres, M. G., Decock, C. y Guzmán-Dávalos, L. (2019). The Ganoderma weberianum-resinaceum lineage: multi-
locus phylogenetic analysis and morphology confirm G. mexicanum and G. parvulum in the Neotropics. Mycokeys, 59, 95–131. https://doi.org/10.3897/mycokeys.59.33182
Cao, Y., Wu, S. H. y Dai, Y. C. (2012). Species clarification of the prize medicinal Ganoderma mushroom ‘‘Lingzhi’’. Fungal Diversity, 56, 49–62. https://doi.org/10.1007/s13225-012-0178-5
Cappello, G. S. (2006). Hongos del Yumka’. Guía ilustrada. Villahermosa: UJAT/ SDSPA.
Castillo, J. y Guzmán, G. (1970). Estudio sobre los poliporáceos de Nuevo León II. Comentarios taxonómicos sobre las especies conocidas y discusiones acerca de su distribución en México. Boletín de la Sociedad Mexicana de Micología, 31, 1-47. https://doi.org/10.17129/botsci.1105
Cilerdzic, J., Vukojevic, J., Stajic, M. y Hadzic, I. (2011). Morpho-physiological diversity between Lingzhi or Reishi medicinal mushroom Ganoderma lucidium (W. Curt.: Fr.) P. Karst. and G. carnosum Pat. International Journal of Medicinal Mushrooms, 13, 465–72. https://doi.org/10.1615/intjmedmushr.v13.i5.60
Corner, E. J. H. (1983). Ad Ployporaceas I. Amauroderma and Ganoderma. Nova Hedwigia, 75, 1–182.
Correia-de Lima, J. N., Baptista, G. T. y Malosso, E. (2014). Delimitation of some neotropical laccate Ganoderma (Ganodermataceae): molecular phylogeny and morphology. Revista de Biología Tropical, 62, 1197–1208.
Gilbertson, R. L. y Ryvarden, L. (1986). North American Polypores. Vol. 1. Abortiporus – Lindtneria. Oslo: Fungiflora. https://doi.org/10.1002/jobm.3620270513
Gomes-Silva, A. C., Ryvarden, L. y Gibertoni, T. B. (2011). New records of Ganodermataceae (Basidiomycota) from Brazil. Nova Hedwigia, 92, 83–94. http://dx.doi.org/10.1127/0029-5035/2011/0092-0083
Gottlieb, A. M. y Wright, J. E. (1999a). Taxonomy of Ganoderma from southern South America: subgenus Ganoderma. Mycological Research, 103, 661–673. https://doi.org/10.1017/S0953756298007941
Gottlieb, A. M. y Wright, J. E. (1999b). Taxonomy of Ganoderma from southern South America: subgenus Elfvingia. Mycological Research, 103, 1289–1298. https://doi.org/10.1017/S095375629800848X
Guzmán, G. (1972). Macromicetos mexicanos en el herbario the national fungus collections de E.U.A. Boletín de la Sociedad Botánica de México, 32, 31–55.
Guzmán, G. (1983). Los hongos de la península de Yucatán II. Nuevas exploraciones y adiciones micológicas. Biótica, 8, 71–100.
Haddow, W. R. (1931). Studies in Ganoderma. Journal of the Arnold Arboretum, 12, 25–46.
Hapuarachchi, K. K., Karunarathna, S. C., Raspé, O., De Silva, K. H. W. L., Thawthong, A., Wu, X. L. et al. (2018). High diversity of Ganoderma and Amauroderma (Ganodermataceae, Polyporales) in Hainan Island, China. Mycosphere, 9, 931–982. https://doi.org/10.5943/mycosphere/9/5/1
Hapuarachchi, K. K., Karunarathna, S. C., Phengsintham, P., Yang, H. D., Kakumyan, P., Hyde, K. D. et al. (2019). Ganodermataceae (Polyporales): diversity in Greater Mekong Subregion countries (China, Laos, Myanmar, Thailand and Vietnam). Mycosphere, 10, 221–309. http://dx.doi.org/10.5943/mycosphere/10/1/6
Hattori, T. y Ryvarden, L. (1994). Type studies in the Polyporaceae 25. Species described from Japan by R. Imazeki & A. Yasuda. Mycotaxon, 50, 27–46.
Hood, I. A. (2006). The mycology of the basidiomycetes. In K. Potter, A. Rimbawanto y C. Beadle (Eds.), Heart rot and root rot in tropical Acacia plantations (pp. 34–59). Canberra, ACIAR Proceedings.
Hong, S. J. y Jung, H. S. (2004). Phylogenetic analysis of Ganoderma based on nearly complete mitochondrial small-subunit ribosomal DNA sequences. Mycologia, 96, 742–755. https://doi.org/10.1080/15572536.2005.11832922
Hsieh, F. G. y Yeh, Z. Y. (2004). Cultural and physiological studies of Ganoderma neo-japonicum and G. zonatum. Bio Formosa, 39, 23–32.
Imazeki, R. (1952). A contribution to the fungus flora of Dutch New Guinea. Bulletin of the Government Forest Experiment Station Tokyo, 57, 87–128.
Imazeki, R., Otani, Y. y Hongo, T. (1988). Coloured illustration of fungi of Japan. Tokyo: Yamakei Publishers Co.
Index Fungorum. (2022). http://www.indexfungorum.org (consultada el 10 de junio de 2022).
Jargalmaa, S., Eimes, J. A., Park, M. S., Park, J. Y., Oh, S. Y. et al. (2017). Taxonomic evaluation of selected Ganoderma species and database sequence validation. PeerJ, 5, e3596. https://doi.org/10.7717/peerj.3596
Karadelev, M. y Rusevska, K. (2004). Eco-taxonomic research of fungi on Bistra Mountain. Proceedings of II Congress of Ecologists of the Republic of Macedonia with International Participation Special Issues of Macedonian Ecological Society. Skopje, 6, 393–397.
Kaur, G., Singh, A. P. y Dhingra, G. S. (2017). Diversity of genus Ganoderma in Punjab (India). Mycobiota, 7, 25–49. http://dx.doi.org/10.12664/mycobiota.2017.07.05
Kaur, J. B., Kaur, R., Kaur, G., Singh, A. P. y Singh, D. G. (2018). Taxonomic notes on the genus Ganoderma from Union Territory of Chandigarh. Kavaka, 51, 35–48.
Küppers, H. (2002). Atlas de los colores. Barcelona: Blume.
Lima-Júnior, N. C., Gibertoni, T. B. y Malosso, E. (2014). Delimitation of some neotropical laccate Ganoderma (Ganodermataceae): molecular phylogeny and morphology. Revista de Biología Tropical, 62, 1197–1208. https://doi.org/10.15517/rbt.v62i3.12380
López-Peña, D., Gutiérrez, A., Hernández-Navarro, E., Valenzuela, R. y Esqueda, M. (2016). Diversidad y distribución de Ganoderma (Polyporales: Ganodermata-
ceae) en Sonora, México. Botanical Sciences, 94, 431–439. https://doi.org/10.17129/botsci.463
López-Peña, D., Samaniego-Rubiano, C., Morales-Estrada, I., Gutiérrez, A., Gaitán-Hernández, R., & Esqueda, M. (2019). Morphological characteristics of wild and cultivated Ganoderma subincrustatum from Sonora, Mexico. Scientia Fungorum, 49, e1213. https://doi.org/10.33885/SF.2019.49.1213
Loyd, A. L., Smith, J. A., Richter, B. S., Blanchette, R. A. y Smith, M. E. (2017). The laccate Ganoderma of the Southeastern United States: a cosmopolitan and important genus of wood decay fungi. Plant Pathology Department. UF/IFAS Extension, 333. https://doi.org/10.32473/edis-pp333-2017
Loyd, A. L., Barnes, C. W., Held, B. W., Schink, M. J., Smith, M. E., Smith, J. A. et al. (2018). Elucidating “lucidum”: distinguishing the diverse laccate Ganoderma species of the United States. Plos One, 13, e0199738. https://doi.org/10.1371/journal.pone.0199738
Mendoza, G., Guzmán, G., Ramírez-Guillén, F., Luna, M. y Trigos, A. (2011). Ganoderma oerstedii (Fr.) Murrill (higher Basidiomycetes), a tree parasite species in Mexico: taxonomic description, rDNA study, and review of its medical applications. International Journal of Medicinal Mushrooms, 13, 545–552. https://doi.org/10.1615/INTJMEDMUSHR.V13.I6.60
Moncalvo, J. M. (2000). Systematics of Ganoderma. In J. Flood, P. D. Bridge, & M. Holderness (Eds.), Ganoderma diseases of perennial crops (pp. 23–45). Wallingford, UK: CABI Publishing.
Moncalvo, J. M. y Ryvarden, L. (1997). A nomenclatural study of Ganodermataceae Donk. Synopsis Fungorum 11. Oslo: Fungiflora.
Moncalvo, J. M., Wang, H. F. y Hseu, R. S. (1995). Gene phylogeny of the Ganoderma lucidum complex based on ribosomal DNA sequences. Comparison with traditional taxonomic characters. Mycological Research, 99, 1489–1499. https://doi.org/10.1016/S0953-7562(09)80798-3
Murrill, W. A. (1902). The Polyporaceae of North America, part I. The genus Ganoderma. Bulletin Torrey Botanical Club, 29, 599–608. https://doi.org/10.2307/2478682
Murrill, W. A. (1905). Polyporaceae of North America XII. Bulletin Torrey Botanical Club, 32, 469–493. https://doi.org/10.2307/2478463
Murrill, W. A. (1908). Polyporaceae, Part 2. North American Flora, 9, 73–131.
Murrill, W. A. (1912). Polyporaceae of Mexico. Bulletin of New York Botanical Garden, 8, 137–153.
Nava-Mora, R. y Valenzuela-Garza, R. (1997). Los macromicetos de la sierra de Nanchititla I. Polibotánica, 5, 21–36.
Ojeda-López, S., Sandoval, M. L. y Valenzuela, R. (1986). Los poliporáceos de México I. Descripción de algunas especies del Noreste de Guanajuato. Revista Mexicana de Micología, 2, 367–436.
Papp, V. y Szabó, I. (2013). Distribution and host preference of poroid Basidiomycetes in Hungary I. Ganoderma. Acta Silvatica et Lignaria Hungarica, 9, 71–83. https://doi.org/10.2478/aslh-2013-0006
Parihar, A., Hembrom, M. E. y Das, K. (2013). New distributional records of Ganoderma colossus (Ganodermataceae) from Jharkhand and Rajasthan. Indian Journal of Plant Sciences, 2, 49–53.
Patouillard, N. T. (1889). Le genere Ganoderma. Bulletin de la Société Mycologique de France, 5, 64–80.
Pinzón-Osorio, C. A y Pinzón-Osorio, J. (2016). Ampliación de la distribución geográfica de Ganoderma australe (Patouillard, 1889) (Aphyllophorales-Basidiomycota), para el departamento de Cundinamarca, Colombia. Revista Biodiversidad Neotropical, 6, 7–11. https://doi.org/10.18636/BIONEOTROPICAL.V6I1.180
Ramos, S. D. O. y Cappello, G. S. (2009). Revisión del género Ganoderma (Basidiomycota) para el estado de Tabasco. Kuxulkab’, 16, 69–75. https://doi.org/10.19136/kuxulkab.a16n29.431
Raymundo, T. y Valenzuela, R. (2003). Los Poliporáceos de México VI. Los hongos poliporoides del estado de Oaxaca. Polibotánica, 16, 79–112.
Ryvarden, L. (2000). Studies in Neotropical polypores 2: a preliminary key to Neotropical species of Ganoderma with a laccate pileus. Mycologia, 92, 180–191. https://doi.org/10.2307/3761462
Singh, R., Singh, A. P., Dhingra, G. S. y Shri, R. (2014). Taxonomy, physicochemical evaluation and chemical investigation of Ganoderma applanatum and G. brownie. International Journal of Advance Research, 2, 702–711.
Steyaert, R. L. (1972). Species of Ganoderma and related genera mainly of the Bogor and Leiden Herbaria. Persoonia, 7, 55–118.
Steyaert, R. L. (1977). Ganoderma resinaceum Boud: should it be considered as a parasite? [On Quercus rubra, in Belgium]. Parasitica (Belgium), 33, 85–90.
Steyaert, R. L. (1980). Study of some Ganoderma species. Bulletin du Jardin Botanique National de Belgique, 50, 135–186. https://doi.org/10.2307/3667780
Tan, W. C., Kuppusamy, U. R., Phan, C. W., Tan, Y. S., Raman, J., Anuar, A. M. et al. (2015). Ganoderma neo-japonicum Imazeki revisited: Domestication study and antioxidant properties of its basidiocarps and mycelia. Scientific Reports, 5, 12515. https://doi.org/10.1038/srep12515
Tchotet-Tchoumi, J. M., Coetzee, M. P. A., Rajchenberg, M., Wingfield, M. J. y Roux, J. (2018). Three Ganoderma species, including Ganoderma dunense sp. nov., associated with dying Acacia cyclops trees in South Africa. Australasian Plant Pathology, 47, 431–447. https://doi.org/10.1007/s13313-018-0575-7
Tchotet-Tchoumi, J. M., Coetzee, M. P., Rajchenberg, M. y Roux, J. (2019). Taxonomy and species diversity of Ganoderma species in the Garden Route National Park of South Africa inferred from morphology and multilocus phylogenies. Mycologia, 111, 730–747. https://doi.org/10.1080/00275514.2019.1635387
Thawthong, A., Hapuarachchi, K. K., Wen, T. C., Raspé, O., Thongklang, N., Kang, J. C. et al. (2017). Ganoderma sichuanense (Ganodermataceae, Polyporales) new to Thailand. Mycokeys, 22, 27–43. https://doi.org/10.3897/mycokeys.22.13083
Torres-Torres, M. G. y Guzmán-Dávalos, L. (2005). Notas sobre la variación morfológica de Ganoderma curtisii en México. Revista Mexicana de Micología, 21, 39–47.
Torres-Torres, M. G. y Guzmán-Dávalos, L. (2012). The morphology of Ganoderma species with a laccate surface. Mycotaxon, 119, 201–216. https://doi.org/10.5248/119.201
Torres-Torres, M. G, Guzmán-Dávalos, L. y Guggliota, A. M. (2012). Ganoderma in Brazil: known species and new records. Mycotaxon, 121, 93–132. http://dx.doi.org/10.5248/121.93
Torres-Torres, M. G., Ryvarden, L. y Guzmán-Dávalos, L. (2015). Ganoderma subgénero Ganoderma en México. Revista Mexicana de Micología, 41, 27–45.
Wang, D. M., Wu, S. H., Su, C. H., Peng, J. T., Shih, Y. H. y Chen, L. C. (2009). Ganoderma multipileum, the correct name for ‘G. lucidum’ in tropical Asia. Botanical Studies, 50, 451–458.
Wang, X. C., Xi, R. J., Li, Y., Wang, D. M. y Yao, Y. J. (2012). The species identity of the widely cultivated Ganoderma, ‘G. lucidum’ (Ling-zhi), in China. Plos One, 7, e40857. http://dx.doi.org/10.1371/journal.pone.0040857
Wang, D. M., Wu, S. H. y Yao, Y. J. (2014). Clarification of the concept of Ganoderma orbiforme with high morphological plasticity. Plos One, 9, e98733. https://doi.org/10.1371/journal.pone.0098733
Welden, L. A y Guzmán, G. (1978). Lista preliminar de los hongos, líquenes y mixomicetos de las regions de Uxpanapa, Coatzacoalcos, los Tuxtlas, Papaloapan, y Xalapa (parte de los estados de Veracruz y Oaxaca). Boletín de la Sociedad Mexicana de Micología, 12, 59–102.
Welti, S. y Courtecuisse, R. (2010). The Ganodermataceae in the French West Indies (Guadeloupe and Martinique). Fungal Diversity, 43, 103–126. http://dx.doi.org/10.1007/s13225-010-0036-2
Wu, S. H. y Zhang, X. Q. (2003). The finding of three Ganodermataceae species in Taiwan. Collection and Research, 16, 61–66.
Yao, Y. J., Wang, X. C. y Wang, B. (2013). Epitypification of Ganoderma sichuanense J. D. Zhao and X. Q. Zhang (Ganodermataceae). Taxon, 62, 1025–1031. https://doi.org/10.12705/625.10
Zhao, J. D. (1989). The Ganodermataceae in China. Bibliotheca Mycologica Band 132. Berlin, Stuttgart: J. Cramer.
Zheng, L., Jia, D., Fei, X., Luo, X. y Yang, Z. (2009). An assessment of the genetic diversity within Ganoderma strains with AFLP and ITSPCR-RFLP. Microbiology Research, 164, 312–321.
Zhou, L. W., Cao, Y., Wu, S. H., Vlasák, J., Li, D. W., Li, M. J. et al. (2015). Global diversity of the Ganoderma lucidum complex (Ganodermataceae, Polyporales) inferred from morphology and multilocus phylogeny. Phytochemistry, 114, 7–15. https://doi.org/10.1016/j.phytochem.2014.09.023
Zmitrovich, I. V. (2018). Conspectus systematis Polyporacearum v. 1.0. Folia Cryptogamica Petropolitana, 6, 3–145.
Zohmangaiha, J. M. C., Vabeikhokhei, J. Z. y Lalrinawmi, H. (2019). Ganoderma species of Mizoram, India. International Journal of Current Microbiology and Applied Sciences, 8, 2871–2879. https://doi.org/10.20546/ijcmas.2019.804.335


