
Caio S. Nogueira a, *, Ana Carolina F. Porto b, Samara P. Barros-Alves c,
Giuliano B. Jacobucci b, Ariádine C. Almeida b
a São Paulo State University, Faculty of Sciences, Department of Biological Sciences, Laboratory of Biology of Marine and Freshwater Shrimps, Av. Engenheiro Luiz Edmundo Carrijo Coube, 14-01, 17033-360 Bauru, São Paulo, Brazil
b Federal University of Uberlândia, Institute of Biology, Laboratory of Ecology of Aquatic Ecosystem, Rua Ceará, s/n, 38400-902 Uberlândia, Minas Gerais, Brazil
c Minas Gerais State University, Ituiutaba Unit, Study group on Crustacean Biology, Ecology, and Culture, R. Vereador Geraldo Moisés da Silva, s/n, 38302-192 Ituiutaba, Minas Gerais, Brazil
*Corresponding author: caiosnogueira@hotmail.com (C.S. Nogueira)
Received: 16 September 2020; accepted: 25 July 2021
Abstract
Growth and longevity parameters of males and females of Macrobrachium brasiliense were analyzed. Sampling was carried out monthly from July 2012 to June 2013 in Triângulo Mineiro region, Minas Gerais, Brazil. A total of 199 adult prawns (97 males and 102 females) were analyzed and measured according to the carapace length (CL, mm). Males (9.64 ± 4.17 mm, CL) were larger than females (7.7 ± 3.39 mm, CL). CL growth curves by the von Bertalanffy model are described by the equations CL = 21.0 [1 – e -0.00446 (t)] for males and CL = 20.0 [1 – e -0.00454 (t)] for females. Comparison of the growth curves by performing the F-type test, showed that there was no difference between the growth coefficients (K) of males and females (GL22; F3.16 = 2.476; p = 0.088). Longevity was estimated as 2.83 years for males and 2.78 years for females. In general, the observed longevity is similar to that of other Macrobrachium species.
Keywords: Caridea; Life strategies; Minas Gerais; von Bertalanffy
© 2022 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Crecimiento y longevidad del camarón de agua dulce Macrobrachium brasiliense (Decapoda: Palaemonidae)
Resumen
En el presente estudio se analizaron los parámetros de crecimiento y longevidad de machos y hembras de Macrobrachium brasiliense. Las muestras se obtuvieron mensualmente desde julio/2012 hasta junio/2013 en la región de Triângulo Mineiro, Minas Gerais, Brasil. Un total de 199 camarones adultos (97 machos, 102 hembras) fueron analizados y medidos según la longitud del caparazón (CL, mm). Los machos (9.64 ± 4.17 mm, CL) fueron más grandes que las hembras (7.7 ± 3.39 mm, CL). Las curvas de crecimiento para CL por el modelo de von Bertalanffy se describen mediante las ecuaciones: CL = 21.0 [1 – e -0.00446 (t)] para los machos y CL = 20.0 [1 – e -0.00454 (t)] para las hembras. Al comparar las curvas de crecimiento con la prueba tipo F, no hubo diferencias entre los coeficientes de crecimiento (K) de machos y hembras (GL22; F3.16 = 2.476; p = 0.088). La longevidad se estimó en 2.83 años para los machos y 2.78 años para las hembras. En general, la longevidad observada es similar a la de las especies de Macrobrachium.
Palabras clave: Caridea; Estrategias de vida; Minas Gerais; von Bertalanffy
Palabras clave: Caridea; Estrategias de vida; Minas Gerais; von Bertalanffy
© 2022 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
Decapod crustaceans are widely studied in many aspects, both ecologically and populationally; however, information about some species in this group beyond the aspects mentioned above is very scarce (i.e., ontogenetic development, growth, and longevity) (Pinheiro & Boss, 2016). For example, it is estimated that less than 2% of existing decapod species have known growth and longevity parameters, although it is known that the majority of the studied species have a wide variation in longevity, living from 1 to 20 years (Vogt, 2019).
The growth process of this group is complex due to the occurrence of ecdysis, in which almost the entire exoskeleton is eliminated and replaced by a new one that will have a larger size. Therefore, the growth and longevity patterns of crustaceans are estimated based mainly on the total body or carapace length and the use of the von Bertalanffy growth model (Hartnoll, 1978; Vogt, 2012). Studies suggest that the growth and longevity of decapod crustaceans can be influenced by many variables, including latitude, habitat, and sex (Vogt, 2012). In addition, they may be related to other life history variables, such as mortality and size at sexual maturity, which can vary depending on the growth pattern of species (Kim, 2005).
Macrobrachium brasiliense (Heller, 1862) has a wide distribution in South America; it is found in Colombia, Ecuador, Guianas, Peru, Venezuela, and Brazil (Amazonas, São Francisco, Paraná, and coastal basins) (Pileggi & Mantelatto, 2012). Its occurrence is related to regions of low elevation and tropical climate (García-Dávila & Magalhães, 2003). In Brazil, M. brasiliense occurs in the states of Amapá, Amazonas, Roraima, Tocantins, Pará, Bahia, Maranhão, Goiás, Mato Grosso, Mato Grosso do Sul, Paraná, São Paulo, and Minas Gerais, in lotic and lentic environments, with clear or dark waters (Coelho & Ramos-Porto, 1984; García-Dávila & Magalhães, 2003; Rossi et al., 2020; Santos et al., 2018).
The first studies on M. brasiliense had a faunistic and taxonomic focus, in addition to addressing some ecological aspects, such as the investigations by Coelho and Ramos-Porto (1984), García-Dávila et al. (2000), Vásquez et al. (2000), and García-Dávila and Magalhães (2003). Over the years, a few studies have been published on ecological aspects of this species, addressing topics such as population structure (Mantelatto & Barbosa, 2005; Oliveira et al., 2019; Pereira & Chacur, 2009), relative growth (Nogueira et al., 2019, 2020), reproductive biology (Oliveira et al., 2019), and habitat and diet preference and characterization (Melo & Nakagaki, 2013; Nogueira et al., 2018) throughout different regions of Brazil.
Growth and longevity are biological aspects still little studied within the genus Macrobrachium. There is information on these aspects only for 11 of the 246 species (Mantelatto et al., 2016): M. amazonicum (Heller, 1892) (Bentes et al., 2016; Freire et al., 2012), M. americanum Spence Bate, 1868 (Ibarra & Wehrtmann, 2020), M. digueti (Bouvier, 1895) (Ibarra & Wehrtmann, 2020), M. equidens (Dana, 1852) (Nwosu, 2008), M. hancocki Holthuis, 1950 (Ibarra & Wehrtmann, 2020), M. macrobrachion (Herklots, 1851) (Deekae & Abowei, 2010; Nwosu et al., 2007), M. occidentale Holthuis, 1950 (Ibarra & Wehrtmann, 2020), M. panamense Rathbun, 1912 (Ibarra & Wehrtmann, 2020), M. rosenbergii (de Man, 1879) (Harikrishnan & Madhusoodana, 1997), M. tenellum (Smith, 1871) (Ibarra & Wehrtmann, 2020), and M. vollenhoveni (Herklots, 1857) (Nwosu & Wolfi, 2006).
Thus, the general knowledge about aspects of population dynamics, such as growth and longevity of the genus Macrobrachium contributed with positive results for the understanding of the current situation of each population. The present study provides the first information about the growth and longevity of males and females of M. brasiliense, thus expanding the understanding of the biology of this freshwater prawn and providing data for future comparative analysis with its congeners.
Material and methods
Sampling was conducted monthly, from July 2012 to June 2013, in the Água Limpa stream (19°05’55.6” S 48°25’38.2” W; Fig. 1), approximately 30 km from Uberlândia city, Minas Gerais, Brazil. Twelve collections were carried out over the sampling period; the specific dates are provided in Table 1. The sampling site was composed of typical vegetation of the Cerrado biome, and the collections took place in a single stretch of the stream that was approximately 20 m long and 2-3 m wide. Macrobrachium brasiliense specimens were sampled by 2 collectors using a sieve of 1.0 mm (mesh). The sieves were dragged along the submerged substrate at depths from 15 to 150 cm over a period of 90 minutes in order to catch the specimens. The sampled individuals were kept in ice storage containers during transport to the laboratory, where they were transferred to labeled pots containing 70% alcohol for posterior identification, sexing, and measurement. The water temperature was measured with a thermometer. Throughout the entire sampling period, the water temperature ranged from 18 to 28 °C (22.9 ± 3.1 °C).
The specimens collected were identified according to Melo (2003) and García-Dávila and Magalhães (2003). It is important to mention that only the species M. brasiliense is recorded in the current study area. Using a stereomicroscope, sex was identified by the presence (male) or absence (female) of the appendix masculina in the second pair of pleopods (Bauer, 2004). Then, the prawns were measured with a caliper (0.1 mm precision) considering the carapace length (CL), i.e., the distance between the posterior margin of the ocular orbit and the midpoint of the posterior margin of the carapace. Specimens that did not have an appendix masculina and were smaller than the smallest male identified were considered undifferentiated juveniles and were not used in the analyses.
Growth estimates were made for males and females. Initially, frequency distributions by size classes were built using Sturges’ method (Sturges, 1926), in order to estimate the modal peaks using Peak-fit software (PeakFit v.4.06 SPSS Inc. for Windows, AISN Software Inc.), which adjusts the observed frequencies to normal distribution curves. The modal peaks were interpreted as age groups, which together over the time provide information for building the growth curves. In case of modal overlapping peaks, they were eliminated as they do not correspond to true age groups. For each adjusted normal curve, a numerical summary containing amplitude (number), mean and standard deviation was obtained. The critical F-value with a probability of 0.05 was calculated for each of the adjustments. Subsequently, the modal peak values were plotted on a scatter plot of age (days) vs. carapace length (mm), thus it was possible to determine cohorts (modal progressions).
The growth parameters of males and females were estimated according to the von Bertalanffy (1938) model, represented by the equation CLt = CL∞ [1 – e -K(t-t0)], where CLt: carapace length at time t; CL∞: asymptotic length; K: constant growth; e: base of natural logarithms; t: age of the individuals; and t0: initial age. The maximum carapace length recorded for captured individual was used as a parameter value for CL∞, and the values of constant growth (K) and theoretical age (t0) were tested in various iterations of trial and error (Fonseca & D’Incao, 2003). The growth curves were constructed through modal progresses, called cohorts in this study. The points were connected, defining different cohorts representing the population within that sampling period (Fonseca & D’Incao, 2003). For each cohort, we used the Solver routine of the Excel program, where the parameters in the von Bertalanffy model (CL∞, K, t0) were estimated based on the least-squared residuals between the lengths observed in the field and calculated by the von Bertalanffy model.
Once the growth curves were selected, age corrections in days were carried out based on time intervals between samples and the obtained t0 value. The modal peaks and the corrected ages were used to calculate an average growth curve, also through the growth model of von Bertalanffy (1938). Mean curves for males and females were compared using a hypothesis test F-type (0.05), according to Cerrato (1990). Longevity (tmax) was obtained by the reverse of the von Bertalanffy equation: tmax = t0 – (1/K) ln [1 – (CLt/CL∞)], considering the t0 parameter equal to zero and the expected length (CLt) as 99% of the asymptotic length (CLt/CL∞ = 0.99, Fonseca & D’Incao, 2003; Miazaki et al., 2019).
Table 1
Information on the dates and quantity of samples of Macrobrachium brasiliense.
| Sampling number | Months |
| 1 | 07/19/2012 |
| 2 | 08/02/2012 |
| 3 | 09/20/2012 |
| 4 | 10/18/2012 |
| 5 | 11/30/2012 |
| 6 | 12/21/2012 |
| 7 | 01/31/2013 |
| 8 | 02/28/2013 |
| 9 | 03/15/2013 |
| 10 | 04/16/2013 |
| 11 | 05/31/2013 |
| 12 | 06/28/2013 |
Results
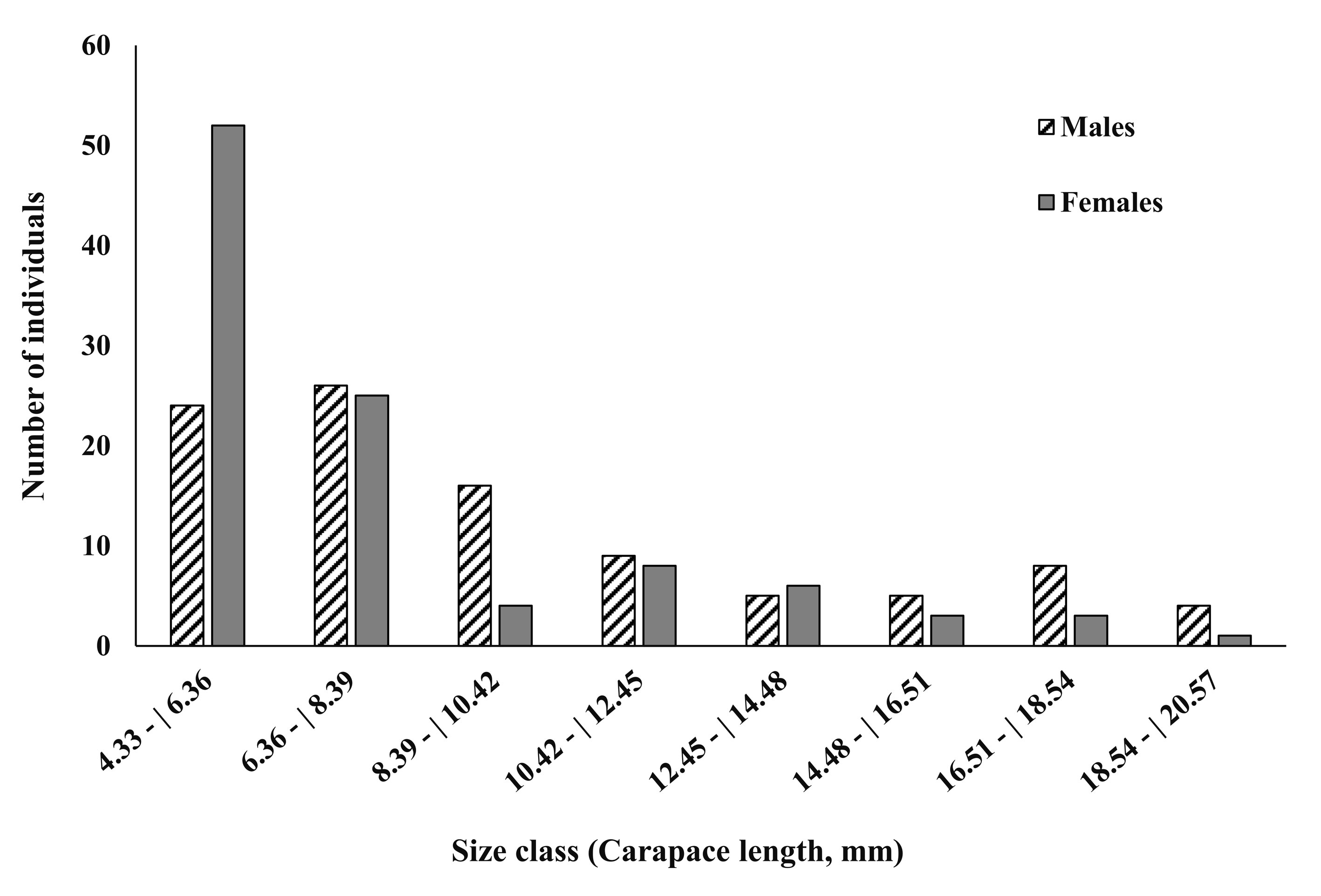
A total of 199 individuals (97 males and 102 females) were analyzed. The average CL was 9.6 ± 4.2 mm for males and 7.7 ± 3.4 mm for females. The variation of CL in males was 4.8 to 20.5 mm, while in females it was 4.8 to 20.1 mm. Eight size classes were estimated, with lower and upper limits of the first and octave size classes of 4.3 and 20.6 mm, respectively, and 2.03 mm of amplitude (Fig. 2).
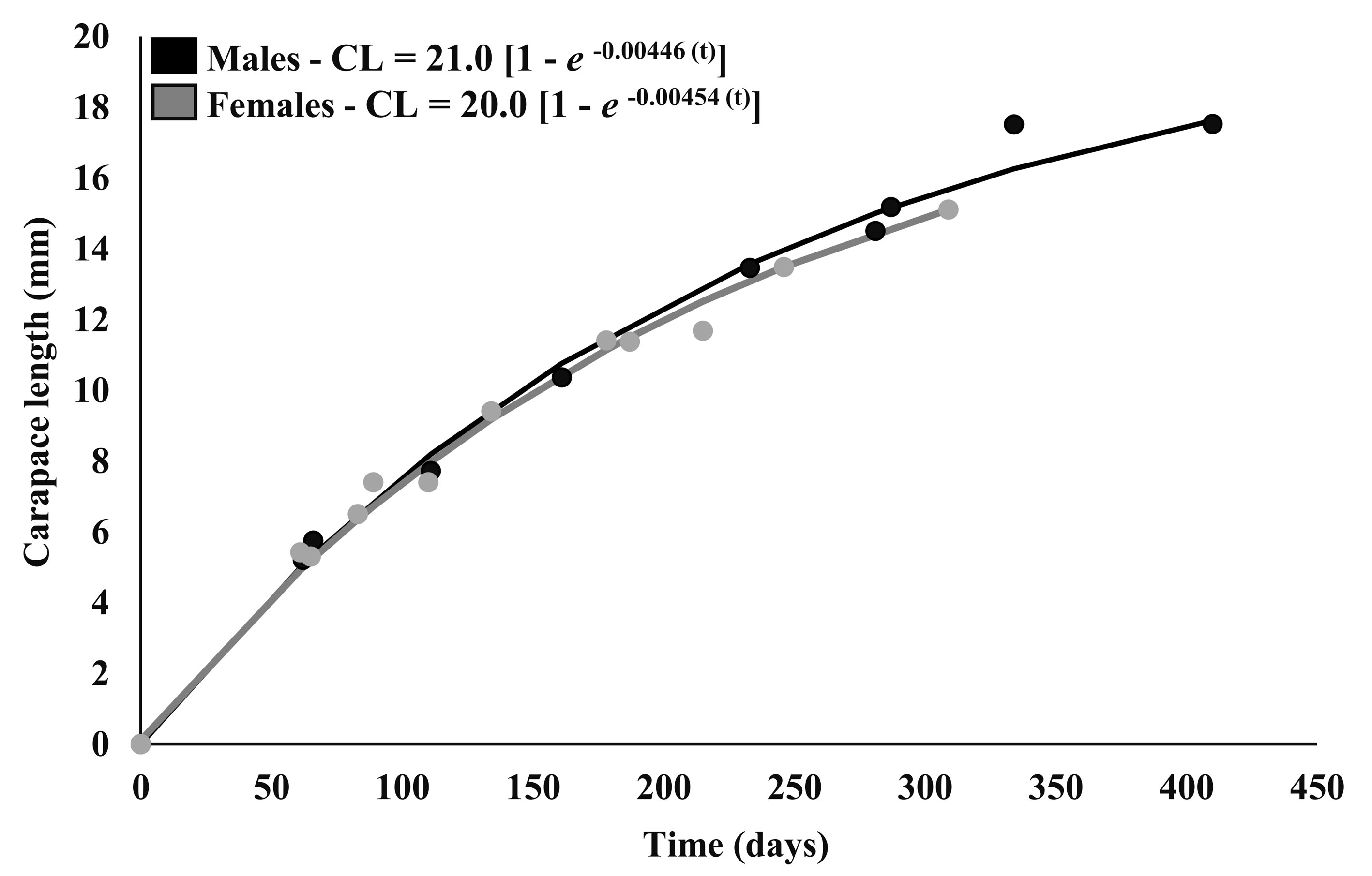
Four and 5 cohorts were analyzed for males and females, respectively. Their growth parameters are described in Table 2. The asymptotic size of males was set as 21.0 mm and females as 20.0 mm, values corresponding to the largest specimens obtained in the present study. From this result it was possible to describe the average growth curves for males CL = 21.0 [1 – e -0.00446 (t)] and females CL = 20.0 [1 – e -0.00454 (t)], calculated by the equation of the von Bertalanffy model (Fig. 3).
Comparing these growth curves using F-test there was no statistical difference between the growth coefficient (K) of males and females (GL22; F3.16 = 2.476; p = 0.088). Using the reverse von Bertalanffy equation, the longevity of M. brasiliense was estimated at 2.83 years for males and 2.78 years for females.
Table 2
Growth parameters of males and females of Macrobrachium brasiliense. Abbreviations: K, growth coefficient; t0, theoretical age at time zero.
| Groups | Cohorts | K (days-1) | t0 | Longevity (years) |
| Males | 1 | 0.0043 | -160.9909 | 2.9575 |
| 2 | 0.0044 | -61.9680 | 2.8878 | |
| 3 | 0.0049 | -65.6324 | 2.5699 | |
| 4 | 0.0054 | -80.8642 | 2.3444 | |
| 1 | 0.0043 | -109.6510 | 2.9557 | |
| 2 | 0.0048 | -82.5130 | 2.6537 | |
| Females | 3 | 0.0052 | -60.7503 | 2.4241 |
| 4 | 0.0047 | -64.9655 | 2.6623 | |
| 5 | 0.0046 | -245.6057 | 2.7653 |
Discussion
Important information on ecological aspects, growth, and longevity of Macrobrachium brasiliense were obtained. These population traits had not yet been investigated for this species. It is noticed that M. brasiliense has such aspects within the parameters already observed in other species of the same genus. In the present study, it was seen that males and females have similar growth rates and longevity values. Such information was discussed, and comparisons were made with congeners and other biological characteristics that influence these aspects (i.e., body size and habitats).
The growth coefficients (K) obtained for males and females of Macrobrachium brasiliense were very close and did not differ statistically. Individuals of both sexes have similar growth rates while they are juveniles. However, once these animals become adults, females grow more slowly than males, possibly due to the high energy investment in gonadal development compared to the somatic growth of males, as observed for Macrobrachium borellii (Nobili, 1896), M. potiuna (Müller, 1880), and M. brasiliense (Bond & Buckup, 1983; Mantelatto & Barbosa, 2005; Nogueira et al., 2019). The differences in the values of the growth coefficient (K) for crustaceans might be related to the energy relocation strategies of each sex (Freire et al., 2012; Hartnoll, 1982).
Although males and females of M. brasiliense have an average CL value with an evident difference, the maximum size that each sex reaches, observed in the present study, is not so unequal (20.5 and 20.1 mm CL for males and females, respectively). This small difference could explain the divergence in life span required to reach the maximum size of males (2.83 years) and females (2.78 years). Possibly the longevity of these individuals is similar because they inhabit small streams where all individuals are susceptible to the same environmental conditions, which can directly influence this correlated life span (Trevisan & Santos, 2011).
Age in decapods can be influenced by many factors, for example, biological traits, geographic distribution, habitat, and others. In addition, there is a negative correlation between growth rate and longevity, in which the latter can vary considerably (Vogt, 2012). In Caridea, longevity can vary from 1 to 11 years, as observed for Pandalus borealis Krøyer, 1838 (11 years; Nilssen & Aschan, 2009), Plesionika izumiae Omori, 1971 (1.5 years; Ahamed & Ohtomi, 2012), Palaemon pandaliformis (Stimpson, 1871) (1-1.5 years; Paschoal et al., 2016), and Exhippolysmata oplophoroides (Holthuis, 1948) (2 years; Pescinelli et al., 2018). Thus, this longevity amplitude is also recorded for congenerics such as M. brasiliense (2.78 and 2.83 years) in the present study, M. hainanense (Parisi, 1919) (4 years; Mantel & Dudgeon, 2005), M. macrobrachion (2.48 years; Deekae & Abowei, 2010; Enin, 1995), M. vollenhoveni (1.97 years; Alhassan & Armah, 2011; Etim & Sankare, 1998), and M. amazonicum (3.86 years; Lucena-Frédou et al., 2010).
Considering some of the freshwater prawns that occur in Brazil, Freire et al. (2012) observed a longevity of approximately 4 years for M. amazonicum, with an average length of 32.5 mm CL (populations from northwestern Brazil). Souza and Fontoura (1996) estimated longevity of approximately 1.25 years for M. potiuna with individuals that had between 8 and 10.5 mm CL (populations from southern Brazil). Therefore, there is a strong relationship between maximum size and longevity, since among these 2 species mentioned and M. brasiliense (present study), M. amazonicum has the largest size and longest longevity, while M. potiuna has a smaller size and shorter longevity. In turn, M. brasiliense has average values of size and longevity between these 2 species, below M. amazonicum and above M. potiuna. Furthermore, such variation in the longevity of M. brasiliense compared to other species of the same genus may be related to interspecific factors (i.e., life history, reproductive strategies, and environmental conditions) and to the different habitats of each species.
This study provides relevant information on the growth and longevity of M. brasiliense, which together with other data available in the literature, can help understand the vulnerability of this species. Furthermore, the results of the present work may contribute to future research of comparative studies carried out in different regions to increase the knowledge in terms of the biology of M. brasiliense.
Acknowledgements
To the FAPEMIG (Fundação de Amparo à Pesquisa de Minas Gerais) and to the Instituto de Biologia of the Universidade Federal de Uberlândia for providing financial support. We are thankful to our LEEA (Laboratório de Ecologia de Ecossistemas Aquáticos) coworkers for their support during the fieldwork. CSN would like to thank the doctoral scholarship provided by Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP – #2019/00661-3). All sampling in this study was conducted in compliance with current applicable state and federal laws.
References
Ahamed, F., & Ohtomi, J. (2012). Growth patterns and longevity of the pandalid shrimp Plesionika izumiae (Decapoda: Caridea). Journal of Crustacean Biology, 32, 733–740. https://doi.org/10.1163/193724012X645853
Alhassan, E. H., & Armah, A. K. (2011). Population dynamics of the African river prawn, Macrobrachium vollenhovenii, in Dawhenya impoundment. Turkish Journal of Fisheries and Aquatic Sciences, 11, 113–119. https://doi.org/10.4194/trjfas.2011.0115
Bauer, R. T. (2004). Remarkable shrimps: adaptations and natural history of the carideans. Norman, Oklahoma: University of Oklahoma Press.
Bentes, B., Martinelli-Lemos, J. M., Lutz, I. A., Nascimento, M. S., & Isaac, V. J. (2016). Population dynamics of Macrobrachium amazonicum (Heller, 1862) (Decapoda: Palaemonidae) in a Brazilian Amazon estuary. Pan-American Journal of Aquatic Sciences, 11, 1–17.
Bond, G., & Buckup, L. (1983). O cultivo de Macrobrachium borellii (Nobili, 1896) e Macrobrachium potiuna (Müller, 1880) em laboratório (Crustacea, Decapoda, Palaemonidae). Revista Brasileira de Biologia, 43, 177–190.
Cerrato, R. M. (1990). Interpretable statistical tests for growth comparisons using parameters in the von Bertalanffy equation. Canadian Journal of Fisheries and Aquatic Sciences, 47, 1416–1426. https://doi.org/10.1139/f90-160
Coelho, P. A., & Ramos-Porto, M. (1984). Camarões de água doce do Brasil: distribuição geográfica. Revista Brasileira de Zoologia, 2, 405–410. https://doi.org/10.1590/S0101-81751984000200014
Cruz, B. R., Cunha, M. C., Bueno, A. A. P., & Jacobucci, G. B. (2021). Natural diet of Macrobrachium brasiliense (Crustacea, Decapoda) in a Cerrado stream. Iheringia. Série Zoologia, 111, e2021003. https://doi.org/10.1590/1678-4766e2021003
Deekae, S. N., & Abowei, J. F. N. (2010). Some growth parameters of “Macrobrachium macrobrachion” (Herklots, 1851) from Luubara Creek in Ogoni Land, Niger Delta, Nigeria. Research Journal of Applied Sciences, Engineering and Technology, 2, 558–567.
Enin, U. I. (1995). First estimates of growth, mortality and recruitment parameters of Macrobrachium macrobrachion Herklots, 1851 in the Cross River Estuary, Nigeria. Dana, 2, 29–38.
Etim, L., & Sankare, Y. (1998). Growth and mortality, recruitment and yield of the fresh-water shrimp, Macrobrachium völlenhovenii, Herklots 1851 (Crustacea, Palaemonidae) in the Fahe reservoir, Côte d’Ivoire, West Africa. Fisheries Research, 38, 211–223. https://doi.org/10.1016/S0165-7836(98)00161-1
Fonseca, D. B., & D’Incao, F. (2003). Growth and reproductive parameters of Kalliapseudes schubartii in the estuarine region of the Lagoa dos Patos (southern Brazil). Journal of the Marine Biological Association of the United Kingdom, 83, 931–935. https://doi.org/10.1017/S0025315403008087h
Freire, J. L., Marques, C. B., & Silva, B. B. (2012). Estrutura populacional e biologia reprodutiva do camarão-da-amazônia Macrobrachium amazonicum (Heller, 1862) (Decapoda: Palaemonidae) em um estuário da região nordeste do Pará, Brasil. Brazilian Journal of Aquatic Sciences and Technology, 16, 65–76. https://doi.org/10.14210/bjast.v16n2.p65-76
García-Dávila, C. R., Alcantára, Β. F., Vasquez, R. E., & Chujandama, S. M. (2000). Biologia reprodutiva do camarão Macrobrachium brasiliense (Heller, 1862) (Crustacea: Decapoda: Palaemonidae) em igarapés de terra firme da Amazônia Peruana. Acta Amazonica, 30, 653–653. https://doi.org/10.1590/1809-43922000304664
García-Dávila, C. R., & Magalhães, C. (2003). Revisão taxonômica dos camarões de água doce (Crustacea: Decapoda: Palaemonidae, Sergestidae) da Amazônia Peruana. Acta Amazonica, 33, 663–686. https://doi.org/10.1590/S0044-59672003000400013
Harikrishnan, M., & Madhusoodana, K. B. (1997). Growth, mortality and exploitation of male and female populations of Macrobrachium rosenbergii (de Man) in the Vembanad lake, India. Indian Journal of Fisheries, 44, 337–344.
Hartnoll, R. G. (1978). The determination of relative growth in Crustacea. Crustaceana, 34, 281–293. https://doi.org/10.1163/156854078X00844
Hartnoll, R. G. (1982). Growth. In D.E. Bliss (Ed.), The biology of Crustacea: embryology, morphology, and genetics (pp. 11–196). New York: Academic Press.
Ibarra, M. A., & Wehrtmann, I. S. (2020). Estimates of growth and longevity of six species of freshwater shrimps (Macrobrachium spp.) (Decapoda: Caridea: Palaemonidae) from Costa Rica. Journal of Crustacean Biology, 40, 45–57.
Kim, S. (2005). Population structure, growth, mortality, and size at sexual maturity of Palaemon gravieri (Decapoda: Caridea: Palaemonidae). Journal of Crustacean Biology, 25, 226–232. https://doi.org/10.1651/C-2510
Lucena-Frédou, F., Filho, J. S. R., Silva, M. C., & Azevedo, E. F. (2010). Population dynamics of the river prawn, Macrobrachium amazonicum (Heller, 1862) (Decapoda, Palaemonidae) on Combu island (Amazon estuary). Crustaceana, 83, 277–290. https://doi.org/10.1163/001121609X12596543952298
Mantel, S. K., & Dudgeon, D. (2005). Reproduction and sexual dimorphism of the palaemonid shrimp Macrobrachium hainanense in Hong Kong streams. Journal of Crustacean Biology, 25, 450–459. https://doi.org/10.1651/C-2541
Mantelatto, F. L. M., & Barbosa, L. R. (2005). Population structure and relative growth of freshwater prawn Macrobrachium brasiliense (Decapoda, Palaemonidae) from São Paulo State, Brazil. Acta Limnologica Brasiliensia, 17, 245–255.
Mantelatto, F. L. M., Pileggi, L. G., Magalhães, C., Carvalho, F. L., Rocha, S. S., Mossolin, E. C. et al. (2016). Avaliação dos Camarões Palemonídeos (Decapoda: Palaemonidae). In M. A. A. Pinheiro, & H. Boss (Eds.), Livro vermelho dos crustáceos do Brasil: avaliação 2010–2014. Porto Alegre, RS: Sociedade Brasileira de Carcinologia.
Melo, G. A. S. (2003). Manual de identificação dos Crustacea Decapoda de água doce do Brasil. São Paulo: Edições Loyola.
Melo, M. S., & Nakagaki, J. M. (2013). Evaluation of the feeding habits of Macrobrachium brasiliense (Heller, 1862) in the Curral de Arame stream (Dourados/Mato Grosso Do Sul, Brazil). Nauplius, 21, 25–33. https://doi.org/10.1590/S0104-64972013000100004
Miazaki, L. F., Simões, S. M., Castilho, A. L., & Costa, R. C. (2019). Population dynamics of the crab Hepatus pudibundus (Herbst, 1785) (Decapoda, Aethridae) on the southern coast of São Paulo state, Brazil. Journal of the Marine Biological Association of the United Kingdom, 99, 867–878. https://doi.org/10.1017/S0025315418000620
Nilssen, E. M., & Aschan, M. M. (2009). Catch, survey and life-history data for shrimp (Pandalus borealis) off Jan Mayen. Deep Sea Research Part II: Topical Studies in Oceanography, 56, 2023–2036. https://doi.org/10.1016/j.dsr2.2008.11.013
Nogueira, C. S., Costa, T. M., & Almeida, A. C. (2018). Habitat choice behavior in Macrobrachium brasiliense (Heller, 1862) (Decapoda, Palaemonidae) under laboratory conditions. Oecologia Australis, 22, 55–62. https://doi.org/10.4257/oeco.2018.2201.05
Nogueira, C. S., Oliveira, M. S., Jacobucci, G. B., & Almeida, A. C. (2019). Relative growth of freshwater prawn Macrobrachium brasiliense (Decapoda, Palaemonidae) and its implications for reproduction. Iheringia, Série Zoologia, 109, e2019005. https://doi.org/10.1590/1678-4766e2019005
Nogueira, C. S., Pantaleão, J. A. F., Almeida, A. C., & Costa, R. C. (2020). Male morphotypes of the freshwater prawn Macrobrachium brasiliense (Decapoda: Caridea: Palaemonidae). Invertebrate Biology, 139, e12279. https://doi.org/10.1111/ivb.12279
Nwosu, F. M. (2008). Growth and mortality of the rough river prawn Macrobrachium equidens Dana, 1852 (Crustacea, Palaemonidae) in Cross River Estuary, Southeast Nigeria. Journal of Food Agriculture and Environment, 6, 186–189.
Nwosu, F. M., Holzlöhner, S., & Enin, U. (2007). The exploited population of the brackish river prawn (Macrobrachium macrobrachion Herklots 1851) in the Cross River Estuary, Nigeria. Scientia Marina, 71, 115–121. https://doi.org/10.3989/scimar.2007.71n1115
Nwosu, F. M., & Wolfi, M. (2006). Population dynamics of the giant African river prawn Macrobrachium vollenhovenii Herklots 1857 (Crustacea, Palaemonidae) in the Cross River estuary, Nigeria. West Africa Journal of Applied Ecology, 9, 1–14. https://doi.org/10.4314/wajae.v9i1.45681
Oliveira, L. J. F., Sant’Anna, B. S., & Hattori, G.Y. (2019). Population biology of the freshwater prawn Macrobrachium brasiliense (Heller, 1862) in the Middle Amazon Region, Brazil. Tropical Zoology, 32, 19–36. https://doi.org/10.1080/03946975.2018.1542195
Paschoal, L. R., Guimarães, F. J., & Couto, E. C. (2016). Growth and reproductive biology of the amphidromous shrimp Palaemon pandaliformis (Decapoda: Caridea) in a Neotropical river from northeastern Brazil. Zoologia, 33, e20160060. https://doi.org/10.1590/s1984-4689zool-20160060
Pereira, M. G. C., & Chacur, M. M. (2009). Estrutura populacional de Macrobrachium brasiliense (Crustacea, Palaemonidae) do Córrego Escondido, Batayporã, Mato Grosso do Sul, Brasil. Revista de Biologia Neotropical, 6, 75–82. https://doi.org/10.5216/rbn.v6i1.12630
Pescinelli, R. A., Davanso, T. M., Pantaleão, J. A. F., Carvalho-Batista, A., Bauer, R. T., & Costa, R. C. (2018). Population dynamics, relative growth and sex change of the protandric simultaneous hermaphrodite Exhippolysmata oplophoroides (Caridea: Lysmatidae) close to an upwelling area. Journal of the Marine Biological Association of the United Kingdom, 98, 727–734. https://doi.org/10.1017/S0025315416001880
Pileggi, L. G., & Mantelatto, F. L. (2012). Taxonomic revision of doubtful Brazilian freshwater shrimp species of genus Macrobrachium (Decapoda, Palaemonidae). Iheringia, Série Zoologia, 102, 426–437. https://doi.org/10.1590/S0073-47212012005000012
Pinheiro, M. A., & Boos, H. (2016). Livro vermelho dos crustáceos do Brasil: Avaliação 2010-2014. Porto Alegre: Sociedade Brasileira de Carcinologia.
Rossi, N., Magalhaes, C., Mesquita, E. R., & Mantelatto, F. L. (2020). Uncovering a hidden diversity: a new species of freshwater shrimp Macrobrachium (Decapoda: Caridea: Palaemonidae) from Neotropical region (Brazil) revealed by morphological review and mitochondrial genes analyses. Zootaxa, 4732, 177–195. https://doi.org/10.11646/zootaxa.4732.1.9
Santos, M., Castro, P., & Magalhães, C. (2018). Freshwater shrimps (Crustacea, Decapoda, Caridea, Dendrobranchiata) from Roraima, Brazil: species composition, distribution, and new records. Check List, 14, 21–35. https://doi.org/10.15560/14.1.21
Souza, G. D., & Fontoura, N. F. (1996). Reprodução, longevidade e razão sexual de Macrobrachium potiuna (Müller, 1880) (Crustacea, Decapoda, Palaemonidae) no arroio Sapucaia, município de Gravataí, Rio Grande do Sul. Nauplius, 4, 49–60.
Sturges, H. A. (1926). The choice of a class interval. Journal of the American Statistical Association, 21, 65–66.
Trevisan, A., & Santos, S. (2011). Crescimento de Aegla manuinflata (Decapoda, Anomura, Aeglidae) em ambiente natural. Iheringia, Série Zoologia, 101, 336–342. https://doi.org/10.1590/S0073-47212011000300008
Vásquez, E., Chujandama, M., García, C., & Alcántara, F. (2000). Caracterización del hábitat del camarón Macrobrachium brasiliense en ambientes acuáticos de la carretera Iquitos-Nauta. Folia Amazónica, 10, 57–71. https://doi.org/10.24841/fa.v10i1-2.214
Vogt, G. (2012). Ageing and longevity in the Decapoda (Crustacea): a review. Zoologischer Anzeiger, 251, 1–25. https://doi.org/10.1016/j.jcz.2011.05.003
Vogt, G. (2019). A compilation of longevity data in decapod crustaceans. Nauplius, 27, e2019011. https://doi.org/10.1590/2358-2936e2019011
von Bertalanffy, L. (1938). A quantitative theory of organic growth (inquiries on growth laws. II). Human Biology, 10, 181–213.


