Aves del Alto Balsas de Guerrero: diversidad e identidad ecológica de una región prioritaria para la conservación
Leopoldo D. Vázquez-Reyes a, b, *, Víctor H. Jiménez-Arcos c, d, Samuel A. SantaCruz-Padilla d, Roberto García-Aguilera d, Abraham Aguirre-Romero d, María del Coro Arizmendi c y Adolfo G. Navarro-Sigüenza a
a Museo de Zoología, Departamento de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional Autónoma de México, Apartado postal 70-399, 04510 Ciudad de México, México
b Laboratorio de Ecología Funcional y Restauración Ecológica, Instituto de Investigaciones en Ecosistemas y Sustentabilidad, Universidad Nacional Autónoma de México. Antigua Carretera a Pátzcuaro Núm. 8701, Ex Hacienda de San José de la Huerta, 58190 Morelia, Michoacán, México
c Laboratorio de Ecología, Unidad de Biotecnología y Prototipos, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autónoma de México, Avenida de los Barrios 1, Los Reyes Ixtacala, 05490 Tlalnepantla, Estado de México, México
d Naturam Sequi A.C., Prol. 16 de Septiembre # 43, Col. Ampliación Ciudad de los Niños, 53450 Naucalpan de Juárez, Estado de México, México
*Autor para correspondencia: leopoldo.vazquez@unam.mx (L.D. Vázquez-Reyes)
Resumen
El Alto Balsas es una región importante para la biodiversidad mexicana por sus altos niveles de recambio taxonómico y riqueza de endemismos. Desafortunadamente la pérdida de hábitat por actividades humanas amenaza su conservación. El objetivo de este trabajo es describir la biodiversidad de aves del Alto Balsas de Guerrero, a partir de datos de incidencia de especies en 7 tipos de hábitat entre 2006 y 2017. Se registraron 209 especies; 53 de ellas representan recambio neto entre tipos de hábitat (βsim promedio 0.41 ± 0.28). Cincuenta y ocho especies (27.7%) tienen alguna categoría de endemismo, lo que constituye una de las avifaunas evolutivamente más mexicanas que se han registrado. Veinticinco especies (12%) están amenazadas, destacando la guacamaya verde (Ara militaris), vulnerable a nivel global. El gradiente ecológico y la historia evolutiva de la región definieron altas tasas de recambio taxonómico, una compleja identidad funcional y conectividad biogeográfica. Considerando estos aspectos, junto con la extensión territorial involucrada, el Alto Balsas de Guerrero cumple con criterios para su inclusión en el Sistema Nacional de Áreas Naturales Protegidas.
Palabras clave:
Biodiversidad; Bosque tropical seco; Diversidad beta; Funcionalidad ecológica; Endemismo; IBA; Recambio de especies
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Birds in the Alto Balsas of Guerrero: diversity and ecological identity of a priority conservation area
Abstract
The Alto Balsas region is important for Mexican biodiversity due to the high levels of taxonomic turnover and richness of endemism. Unfortunately, habitat loss owing to human activities threats its conservation. The goal of this contribution is to describe the birds biodiversity in the Alto Balsas region in Guerrero, using species incidence data in seven habitat-types between 2006 and 2017. A total of 209 bird species was recorded, 53 of which represent effective taxonomic turnover (mean βsim 0.41 ± 0.28) between habitats. Fifty-eight species (27.7%) have some level of endemism, representing one of the most evolutionarily Mexican bird assemblages recorded. Twenty-five species (12%) are threatened, highlighting the military macaw (Ara militaris), a globally threatened bird. Both evolutionary history and the ecological gradient defined high taxonomic turnover rates, a complex functional identity, as well as biogeographic connectivity. Considering this aspects, and involving territorial surface, the Alto Balsas in Guerrero meets the criteria for its inclusion in the National System of Natural Protected Areas.
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Keywords:
Biodiversity; Tropical dry forest; Beta diversity; Ecological function; Endemics; IBA; Species turnover
Introducción
La cuenca del río Balsas es una provincia biogeográfica del centro de México limitada por las elevaciones de la Faja Volcánica Transmexicana y la sierra Madre del Sur. Incluye las zonas con altitud menor a 2,000 m snm de los estados de Guerrero, Jalisco, México, Michoacán, Morelos, Oaxaca y Puebla (Morrone, 2014). Su orografía presenta drásticos cambios altitudinales que van desde tierras bajas en la desembocadura del río en el océano Pacífico, hasta 2,000 m snm en las serranías.
La avifauna de esta provincia biogeográfica posee una considerable riqueza específica, aproximadamente el 33% (379 especies) del total para México (Castro-Torreblanca et al., 2014), debido a su complejidad ecológica: aunque las tierras bajas de la cuenca están dominadas por bosques tropicales caducifolios, en las tierras altas alberga bosques de encino, con afinidad Neártica (Almazán-Núñez y Navarro, 2006; Vázquez-Reyes et al., 2017). Además, las variaciones microclimáticas locales promueven composiciones bióticas particulares y altas tasas de recambio entre áreas específicas del bosque tropical seco (Trejo y Dirzo, 2002). Esto define un elevado nivel de recambio taxonómico a través del gradiente altitudinal (Espinosa-Organista et al., 2008; Koleff et al., 2008). Los bosques tropicales subcaducifolios, palmares, vegetación riparia y ambientes acuáticos complementan la compleja matriz ecológica regional (Fernández-Nava et al., 1998; Ramírez-Albores, 2007; Ramírez-Albores y Ramírez-Cedillo, 2002).
Por otro lado, la riqueza de especies endémicas para México representa aproximadamente el 26% de su riqueza específica total (Navarro-Sigüenza et al., 2014). Por esta razón, se le considera un área de endemismo de aves de importancia global (EBA-008; Stattersfield et al., 1999). Este patrón se origina por el aislamiento orográfico de la cuenca (Morrone, 2014), los cambios paleoecológicos que moldearon la cobertura de bosques tropicales y templados, provocando efectos de aislamiento y diversificación de especies restringuidas entre final del Mioceno (Mason et al., 2018; Navarro-Sigüenza et al., 2017) y el Pleistoceno (Linares-Palomino et al., 2011; Lovette, 2005; Pennington et al., 2000). Por esta conjugación de factores ecológicos e históricos, la avifauna de la cuenca del Balsas posee un gran valor para la biodiversidad mexicana (Espinosa-Organista et al., 2008; Morrone, 2014; Ríos-Muñoz, 2013).
Actualmente, tenemos una valiosa visión general de la biodiversidad de aves en la cuenca del Balsas gracias a diversos inventarios (Almazán-Núñez y Navarro, 2006; Nova-Muñoz et al., 2011; Ramírez-Albores, 2007; Ramírez-Albores y Ramírez-Cedillo, 2002; Rojas-Soto et al., 2009) y estudios biogeográficos a partir de bases de datos (Castro-Torreblanca et al., 2014). Sin embargo, existen importantes vacíos de información; particularmente en el Alto Balsas de Guerrero, subprovincia definida por las cuencas de los ríos Amacuzac, Nexapa, Atoyac, Mixteco y Tlapaneco, al oriente de la sierra de Taxco (Meza y López-García, 1997). Hasta el año 2012, se ignoraba la presencia de la guacamaya verde, Ara militaris, en el Alto Balsas (Jiménez-Arcos et al., 2012), aún cuando se trata de una especie prioritaria en México y vulnerable a nivel global (Conabio, 2017). Además, hasta ahora no se han publicado estudios de campo enfocados en la riqueza, composición y recambio taxonómico de las aves del Alto Balsas de Guerrero.
Desafortunadamente, la pérdida de hábitat pone en grave riesgo la conservación de las aves del trópico seco mexicano, incluyendo la cuenca del Balsas (NABCI, 2016). La agricultura, la ganadería y el desarrollo de asentamientos humanos impulsan tasas de deforestación cercanas al 4% anual (Maass et al., 2010; Sánchez-Azofeifa y Portillo-Quintero, 2011). Como consecuencia, el bosque tropical caducifolio conservado representa sólo el 12.8% de su extensión original, mientras los encinares conservados cubren el 35% (Conafor, 2014). Aunado a ésto, no existen áreas naturales protegidas relevantes en la región (existen 2 reservas estatales, ninguna mayor a 450 ha (Conanp, 2018; Semaren, 2018). Esta situación limita las posibilidades para frenar la pérdida de hábitat, principal amenaza para las aves de México (Peterson y Navarro-Sigüenza, 2016).
Como respuesta a dicha problemática, este trabajo tiene como objetivo describir la biodiversidad de aves del Alto Balsas de Guerrero para aportar información útil en la toma de decisiones encaminadas a su conservación. Para ello, se presenta: el inventario taxonómico regional, la riqueza específica en los tipos de hábitat dominantes, el análisis de recambio taxonómico entre tipos de hábitat; así como el análisis de estacionalidad, endemismo, estatus de conservación, vulnerabilidad y atributos ecológicos generales de la avifauna.
Materiales y métodos
El área de estudio se localiza al noreste del estado de Guerrero (municipios de Atenango del Río, Copalillo y Olinalá), incluyendo el extremo suroeste del estado de Puebla (municipio de Santa María Cohetzala), debido a que representan un continuo de hábitat (fig. 1). Las coordenadas extremas son: 18º16’ N, 99º14’ O (noroeste); 18º16’ N, 98º47’ O (noreste); 17º53’ N, 99º14’ O (suroeste) y 17º53’ N, 98º47’ O (sureste), con una superficie de aproximadamente 2,000 km2 dentro del Alto Balsas. El clima es cálido semiseco con lluvias en verano (Aw), con precipitación aproximada de 780 mm anuales (Martínez-Gordillo et al., 1997; Meza y López-García, 1997). La zona incluye un gradiente altitudinal que va de 650 m snm, en las riberas de los ríos Balsas y Amacuzac, hasta 1,950 m snm en las zonas más elevadas (fig. 1).
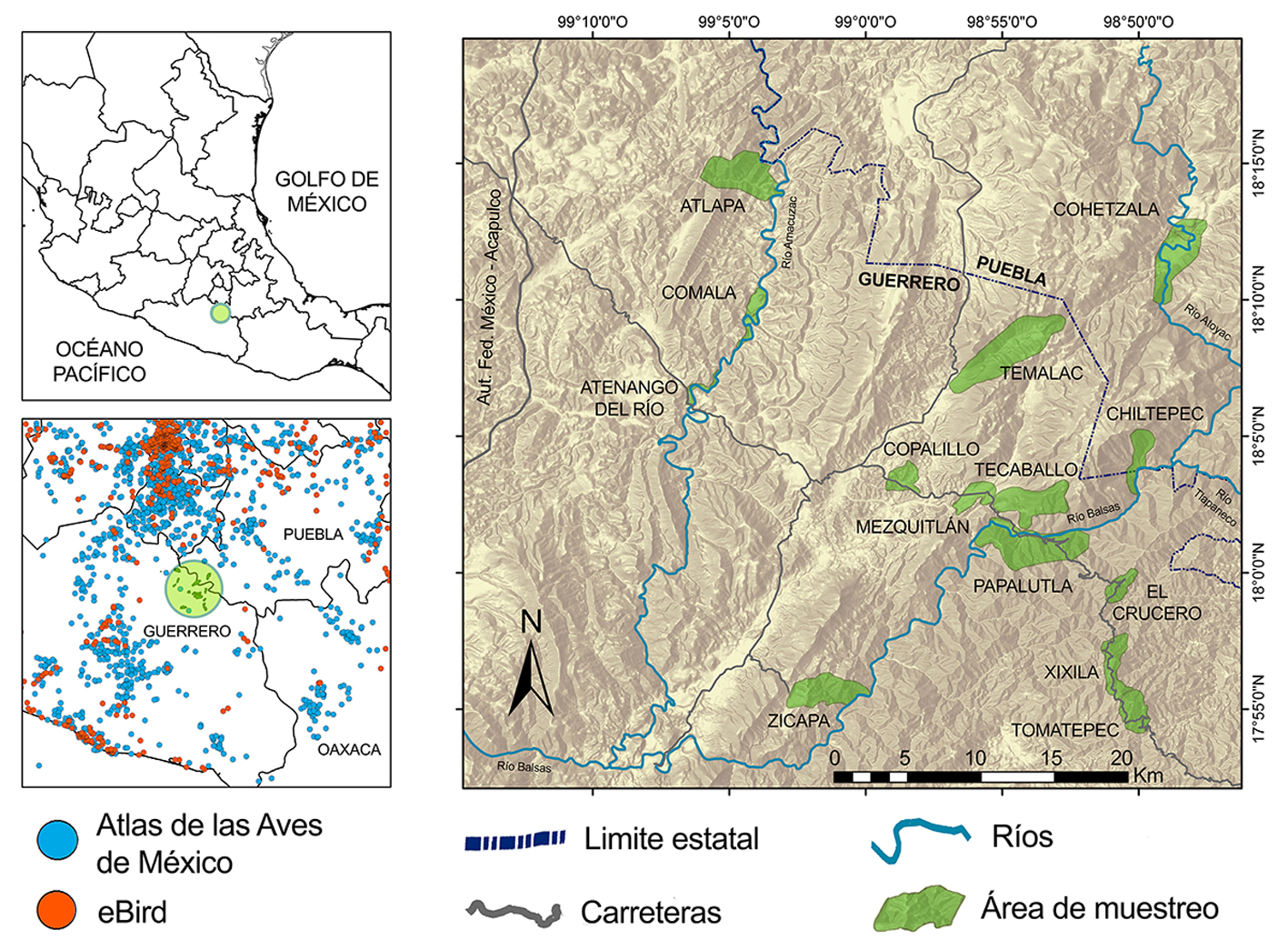
Por su latitud, altitud, temperatura y precipitación, la región corresponde a la zona de vida de bosque tropical seco (Holdridge, 1967), e incluye diferentes tipos de vegetación. Entre 650 y 1,200 m snm, la vegetación es bosque tropical caducifolio. Distribuido en laderas y valles, su estrato arbóreo se caracteriza por árboles de Bursera spp., Lysiloma spp., Cyrtocarpa procera y Ceiba aesculifolia. En barrancas y cañadas la vegetación es bosque tropical subcaducifolio, cuyo estrato arbóreo está representado por Enterolobium cyclocarpum, Ficus spp. y Lonchocarpus spp. En la ribera de los ríos, entre 650 y 750 m snm, existen elementos de bosque de galería representados por Taxodium mucronatum, Caesalpinia coriaria y E. cyclocarpum. Entre 1,500 y 1,950 m snm, el bosque de encino (Quercus spp.) es la vegetación dominante. Entre 1,200 y 1,500 m snm existen ecotonos de bosque de encino con bosque tropical caducifolio que incorporan árboles como Celtis caudata, Heliocarpus spp. y Lysiloma acapulcense. En las laderas norte existen palmares de Brahea dulcis con elementos arbóreos de bosques tropicales y de encino (Fernández-Nava et al., 1998; Martínez-Gordillo et al., 1997; Rzedowski, 2006). El hábitat acuático disponible incluye los cauces de los ríos, corrientes temporales en las barrancas, pozas naturales y artificiales. La disponibilidad de agua es variable en función de la temporada de lluvias, aunque el río y los jagüeyes mantienen agua todo el año. En la región existen asentamientos humanos que han provocado una pérdida aproximada del 18% de la cobertura forestal debido al desarrollo de agricultura de subsistencia, ganadería extensiva, extracción de madera para leña y construcción (Naturam Sequi, 2016). En el hábitat conservado, las actividades humanas se limitan al tránsito eventual de personas y ganado a través de veredas.
Este trabajo recopila los datos de incidencia de aves obtenidos entre junio de 2006 y noviembre de 2017, a través de diferentes proyectos enfocados al estudio de la biodiversidad del Alto Balsas (Jiménez-Arcos et al., 2012; Naturam Sequi, 2015, 2016; Vázquez-Reyes et al., 2017). Se consideraron los siguientes tipos de hábitat: bosque tropical caducifolio, bosque tropical subcaducifolio, bosque de encino, ecotonos forestales entre el bosque tropical caducifolio y de encino, palmares, vegetación riparia y hábitat acuático. El esfuerzo de muestreo fue de 293 días, distribuidos en 44 salidas de trabajo de campo. El muestreo de aves se realizó en diversas horas del día, dando prioridad al registro durante las primeras 4 horas después del amanecer, para aprovechar el pico de actividad de las aves diurnas. También se realizaron recorridos vespertinos y nocturnos para obtener datos de las especies crepusculares y nocturnas (Bibby et al., 2000). Siempre que fue posible, se obtuvo registro fotográfico digital. Para ello, se usaron cámaras SLR Canon EOS 30D y 7D, con objetivos EF 100-400mm f/4.5-5.6L IS USM y EF 100MM F/2.8 MACRO USM. Se usaron guías de campo de aves de México (Howell y Webb, 1995; Peterson y Chalif, 1989) y Norteamérica (Dunn y Alderfer, 2006; Robbins et al., 1966) para la identificación de las especies. También se consultaron ejemplares de referencia en la colección del Museo de Zoología de la Facultad de Ciencias de la Universidad Nacional Autónoma de México (MZFC).
En total, se consideraron datos provenientes de 5 métodos de muestreo: 1) recorridos para observación libre, entre junio de 2006 y noviembre de 2017 (190 días), incluyendo los 7 tipos de hábitat; 2) puntos de conteo, entre febrero de 2014 y mayo de 2016 (65 días). En total se realizaron 412 puntos, 204 en bosque tropical caducifolio y 208 en bosque de encino. Se tomaron en cuenta ambientes conservados, vegetación secundaria y poblados en ambos tipos de vegetación. Durante un periodo de 10 minutos se registraron las aves observadas y escuchadas dentro de un radio de 30 m. Los puntos fueron separados al menos 200 metros entre sí para mantener independencia entre los datos obtenidos (Ralph et al., 1996); 3) colecta científica, con redes de niebla (aproximadamente 3,500 horas/metro/red) y rifles de diábolos en el bosque tropical caducifolio, subcaducifolio y de encino; durante 49 días entre febrero de 2014 y marzo de 2016. Los ejemplares se taxidermizaron, colectando los tejidos de músculo, corazón e hígado para estudios moleculares; 4) grabación de cantos y vocalizaciones en todos los tipos de hábitat durante 26 días, entre febrero de 2014 y abril de 2016. Se obtuvieron archivos *.wav, con tasa de grabación de 44.1 KHz e intervalo dinámico de 16 bits con una grabadora Tascam DR-100 y micrófonos Sennheiser ME67 y Telinga TwinScience 5 con parábola, y 5) monitoreo fotográfico comunitario. Se capacitó a pobladores de las localidades de Papalutla, Xixila y Zicapa para obtener registro fotográfico digital georreferenciado de las aves observadas en recorridos libres. Este método sumó 100 días de muestreo entre septiembre de 2016 y febrero de 2017. Los ejemplares colectados (pieles y tejidos), así como grabaciones de cantos y fotografías (obtenidas por LDV-R), fueron depositados en la Colección de Aves del MZFC.
El inventario de las aves registradas (apéndice) se ordenó a nivel supraespecífico con los criterios de la AOS (2017). A nivel específico, se usaron los criterios de la propuesta taxonómica alternativa para las aves de México (Navarro-Sigüenza y Peterson, 2004). Al considerar como especies los linajes monofiléticos evolutivamente independientes, permite una mejor identificación de las áreas prioritarias para conservación por su papel en la diversificación de taxones únicos para México (Bertelli et al., 2017; Navarro-Sigüenza y Peterson, 2004; Navarro-Sigüenza et al., 2014).
Para evaluar la biodiversidad regional, consideramos la riqueza específica registrada por tipo de hábitat como diversidad α; mientras que la riqueza específica regional fue considerada como diversidad γ (Whittaker, 1972). Para calcular la riqueza esperada y modelar la eficiencia del muestreo por tipo de hábitat, se usó el estimador no paramétrico ICE (Incidence Coverage Estimator), basado en datos de incidencia de las aves. Este estimador considera a las especies raras y especies únicas para calcular la probabilidad de encontrar nuevas especies y generar una estimación de la riqueza total a partir de datos de incidencia (Moreno, 2001). El indicador fue elegido dado que tiene un desempeño estable aún con datos provenientes de diferentes tamaños muestrales (Chazdon et al., 1998). El análisis fue ejecutado en el programa EstimateS v. 9.1 (Colwell, 2013). Dado que los modelos de predicción de riqueza específica requieren unidades muestrales comparables, usamos como unidad de muestreo los días de registro de datos en campo, en lugar del número de individuos o eventos de muestro. De esta forma buscamos aprovechar las diferentes fuentes de información en el estudio, intentando reducir posibles sesgos derivados de las diferentes probabilidades de detección para las aves (Hortal y Lobo, 2002; Jiménez-Valverde y Hortal, 2003).
Se consideró al recambio taxonómico entre tipos de hábitat como diversidad β (Whittaker, 1972). Para evaluarlo, se usó el índice βsim, que mide las diferencias asociadas con las especies no compartidas, independientemente de la riqueza específica (Koleff et al., 2003). Esto permite reducir los sesgos que pueden generar tamaños de biotas diferentes en el análisis (Baselga, 2010). Para analizar las relaciones taxonómicas entre tipos de hábitat, se hizo un análisis de clasificación a la matriz de disimilitud con el método de ligamiento completo. Estos análisis se ejecutaron con la paquetería “betapart” (Baselga y Orme, 2012) del software R v. 3.2 (R Development Core Team, 2008).
Se consideraron 6 categorías de residencia a partir de la propuesta de NatureServe (Ridgely et al., 2005), disponible en la Red de Conocimientos sobre las Aves de México (Berlanga et al., 2008): residentes permanentes, migratorias locales, migratorias invernales, migratorias de verano, transitorias, y accidentales.
Se usaron 3 categorías de endemismo: endémicas, con distribución solo en México, cuasiendémicas, cuya distribución fuera de México es < 35,000 km2 y semiendémicas: migratorias cuya distribución invernal se encuentra solo en México (González-García y Gómez-de Silva, 2003).
Se usaron las categorías de riesgo de extinción de acuerdo con la NOM-059-SEMARNAT-2010 (Semarnat, 2010) y la lista roja de la UICN (Birdlife International, 2018). Para evaluar la vulnerabilidad ecológica, se usaron los resultados de la evaluación del estado de conservación de las aves de Norteamérica (NABCI, 2016). Éste se basa en datos de tendencias poblacionales y amenazas de cada especie para generar un índice con valores de 4 a 20. Generamos 3 categorías de vulnerabilidad para comparar entre tipos de hábitat: baja, valores de 4 a 8; media, valores de 9 a 13; y alta, valores de 14 a 18.
Se consideró la dieta general como un atributo relacionado con los requerimientos ecológicos de las aves. Se usaron 9 categorías: nectarívora, frugívora, granívora, carnívora, insectívora, malacófaga, piscívora, carroñera y omnívora (del Hoyo et al., 2017). Por otro lado, la vinculación con servicios ecosistémicos se consideró como atributo del efecto ecológico de las aves sobre el ambiente (Salgado-Negret y Paz, 2015). Se consideraron 9 categorías: polinización, dispersión de semillas, control poblacional de semillas, de vertebrados, de invertebrados, ingeniería ecosistémica (generación de recursos para múltiples taxa), reciclaje de cadáveres, transporte de nutrientes entre hábitat acuático y terrestre, e interacciones ecológicas múltiples (Sodhi et al., 2011). La última categoría fue asignada para aves omnívoras que pueden vincularse con varios de los servicios ecosistémicos que referimos arriba (del Hoyo et al., 2017).
Resultados
En total se registraron 209 especies de aves en el Alto Balsas de Guerrero, distribuidas en 21 órdenes, 50 familias y 135 géneros. El estimador ICE, indicó que la eficiencia de muestreo de la avifauna regional difiere entre tipos de hábitat. Los tipos de hábitat mejor muestreados fueron el bosque de encino (90.7%) y el bosque tropical caducifolio (83.8%). La eficiencia de muestreo en los tipos de hábitat restantes varía entre 70.7% y 34.5% (tabla 1). El bosque tropical caducifolio (150 especies) y el bosque de encino (133 especies) fueron los tipos de hábitat con mayor representación de la biodiversidad regional (71.7% y 63.6% respectivamente), mientras el resto tuvieron una riqueza específica ≤ 50% del total registrado (tabla 1). El inventario taxonómico obtenido se muestra en el apéndice.
|
Tabla 1 Resumen taxonómico de la avifauna por tipo de hábitat en el Alto Balsas de Guerrero. Se indica la riqueza de los taxonómicos de orden, familia, género y especie por tipo de hábitat, el valor de riqueza específica estimada (ICE) y la eficiencia del muestreo para detectar la avifauna. |
||||||||
|
Taxón |
Alto Balsas |
Hábitat acuático |
Vegetación riparia |
Tropical caducifolio |
Tropical subcaducifolio |
Ecotono forestal |
Palmar |
Bosque de encino |
|
Orden |
21 |
9 |
12 |
14 |
15 |
13 |
9 |
14 |
|
Familia |
50 |
13 |
26 |
34 |
30 |
30 |
18 |
35 |
|
Género |
135 |
20 |
40 |
96 |
47 |
69 |
23 |
82 |
|
Especie |
209 |
23 |
60 |
150 |
76 |
103 |
28 |
133 |
|
ICE |
44.57 |
95.36 |
178.86 |
112.06 |
145.69 |
81.17 |
146.64 |
|
|
ICE (%) |
51.60 |
62.92 |
83.86 |
67.82 |
70.70 |
34.50 |
90.70 |
|
Tabla 2 Valores de recambio taxonómico (βsim) de las avifaunas presentes en los tipos de hábitat en el Alto Balsas de Guerrero. |
|||||||
|
βsim (sp. únicas) |
Hábitat acuático |
Vegetación riparia |
Tropical caducifolio |
Tropical subcaducifolio |
Ecotono forestal |
Palmar |
Bosque de encino |
|
acuático (6) |
0.43 |
0.65 |
0.87 |
0.74 |
1.00 |
0.74 |
|
|
riparia (3) |
0.25 |
0.53 |
0.50 |
0.71 |
0.43 |
||
|
caducifolio (21) |
0.04 |
0.18 |
0.07 |
0.26 |
|||
|
subcaducifolio (1) |
0.29 |
0.32 |
0.26 |
||||
|
ecotono (4) |
0.18 |
0.17 |
|||||
|
palmar (0) |
0.07 |
||||||
|
encino (17) |
|||||||
|
βsim promedio |
0.74 (± 0.19) |
0.48 (± 0.15) |
0.24 (± 0.22) |
0.39 (± 0.28) |
0.34 (± 0.23) |
0.39 (± 0.38) |
0.32 (± 0.24) |
|
βsim Alto Balsas |
0.41 (± 0.28) |
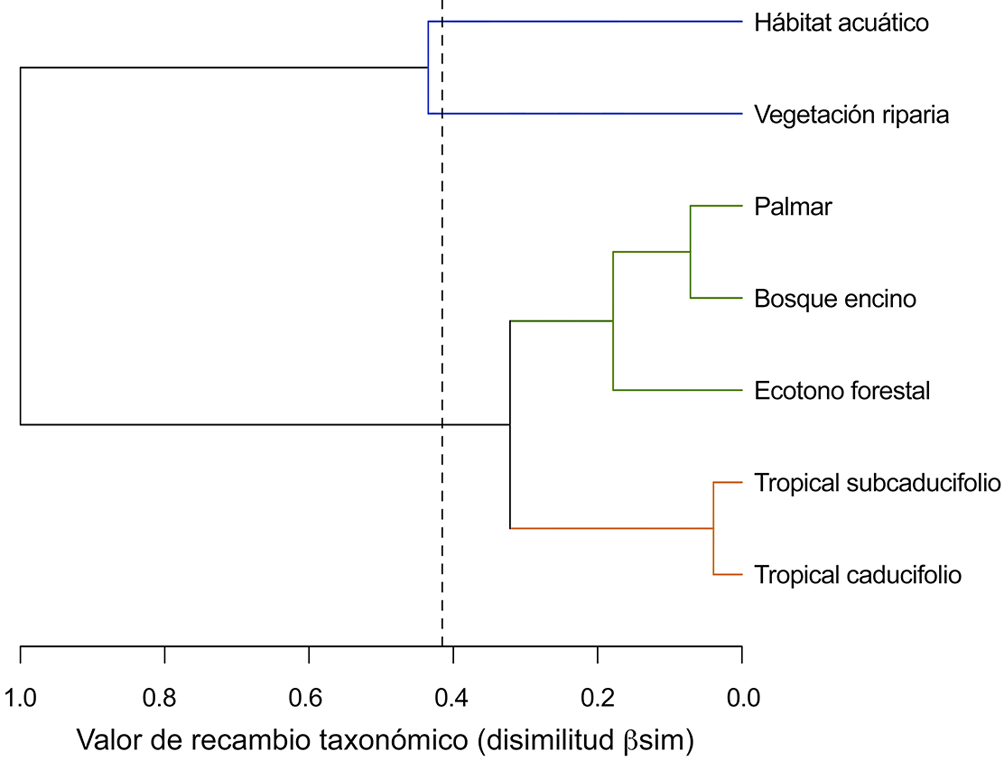
El valor promedio del recambio taxonómico regional (bsim) fue de 0.41 (± 0.28). La tabla 2 presenta la matriz de disimilitud (βsim) a nivel regional y la riqueza de especies únicas por tipo de hábitat. Los bosques tropical caducifolio (22 especies) y de encino (18 especies) aportaron más especies únicas que ningún otro tipo de hábitat (tabla 2). En total, aproximadamente el 25% de la biodiversidad regional (53 especies) representan recambio neto. El análisis de clasificación generó 2 agrupaciones principales (fig. 2). La primera incluyó al hábitat acuático y la vegetación riparia, sumando 70 especies totales y 9 únicas. La segunda agrupación se formó por los diferentes tipos de hábitat forestal (192 especies) y tuvo 2 subgrupos. El primero se formó por el palmar, el bosque de encino y los ecotonos forestales e incluyó 152 especies, 21 de ellas únicas. El segundo subgrupo se formó por los bosques tropicales caducifolio y subcaducifolio, e incluyó 153 especies totales y 22 únicas.
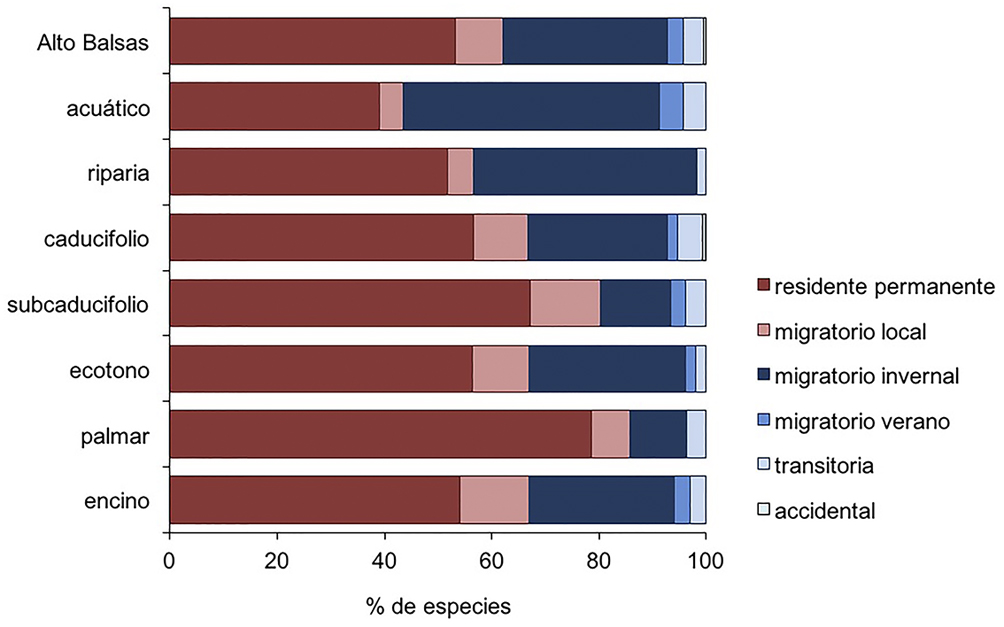
Del total regional, 112 especies fueron residentes permanentes (53.6%), 18 migratorias locales (8.6%), 64 migratorias invernales (30.6%), 6 migratorias de verano (2.8%) y 8 transitorias (3.8%). El bosque tropical caducifolio fue el hábitat con mayor riqueza de especies migratorias de larga distancia, con 49, seguido por el bosque de encino con 44, y los ecotonos entre bosques tropicales y de encino, con 34. La vegetación riparia tuvo 26 migratorias de larga distancia, el hábitat acuático 13, el bosque subcaducifolio 15, y el palmar 4 (fig. 3).
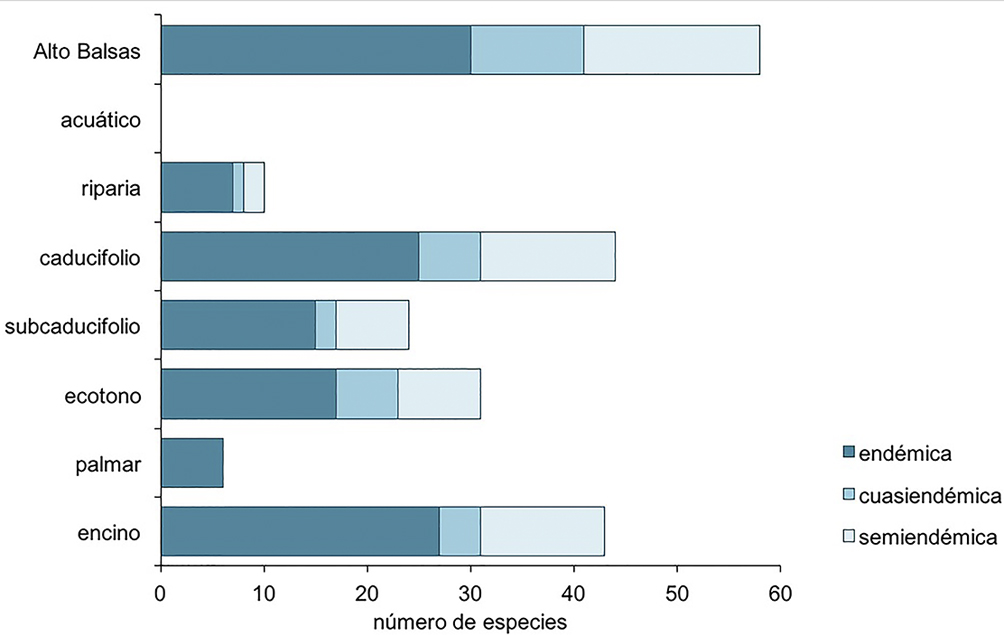
La riqueza específica del componente endémico fue de 30 especies (14.3%), 11 cuasiendémicas (5.2%), y 17 semiendémicas (8.1%), sumando 58 especies (27.7%) a nivel regional. Los tipos de hábitat con mayor riqueza de endemismo fueron el bosque caducifolio, con 44 especies, el bosque de encino, con 43 especies, seguido de los ecotonos con 31, el bosque tropical subcaducifolio con 24, la vegetación riparia con 12 y el palmar con 6. No se registró ninguna especie endémica en el hábitat acuático (fig. 4).
El 12% (25 especies) de la biodiversidad registrada se considera en riesgo de acuerdo con la legislación mexicana y la lista roja de la UICN. Las especies en mayor riesgo son: Ara militaris (guacamaya verde), en peligro de extinción en México y vulnerable a nivel global; Megascops seductus (tecolote del Balsas), amenazado en México y cerca de amenaza a nivel global; Xenotriccus mexicanus (mosquero del Balsas) y Passerina ciris (colorín sietecolores), ambas especies sujetas a protección especial en México y cerca de amenaza global. El estatus de conservación de todas las especies se presenta en el apéndice.
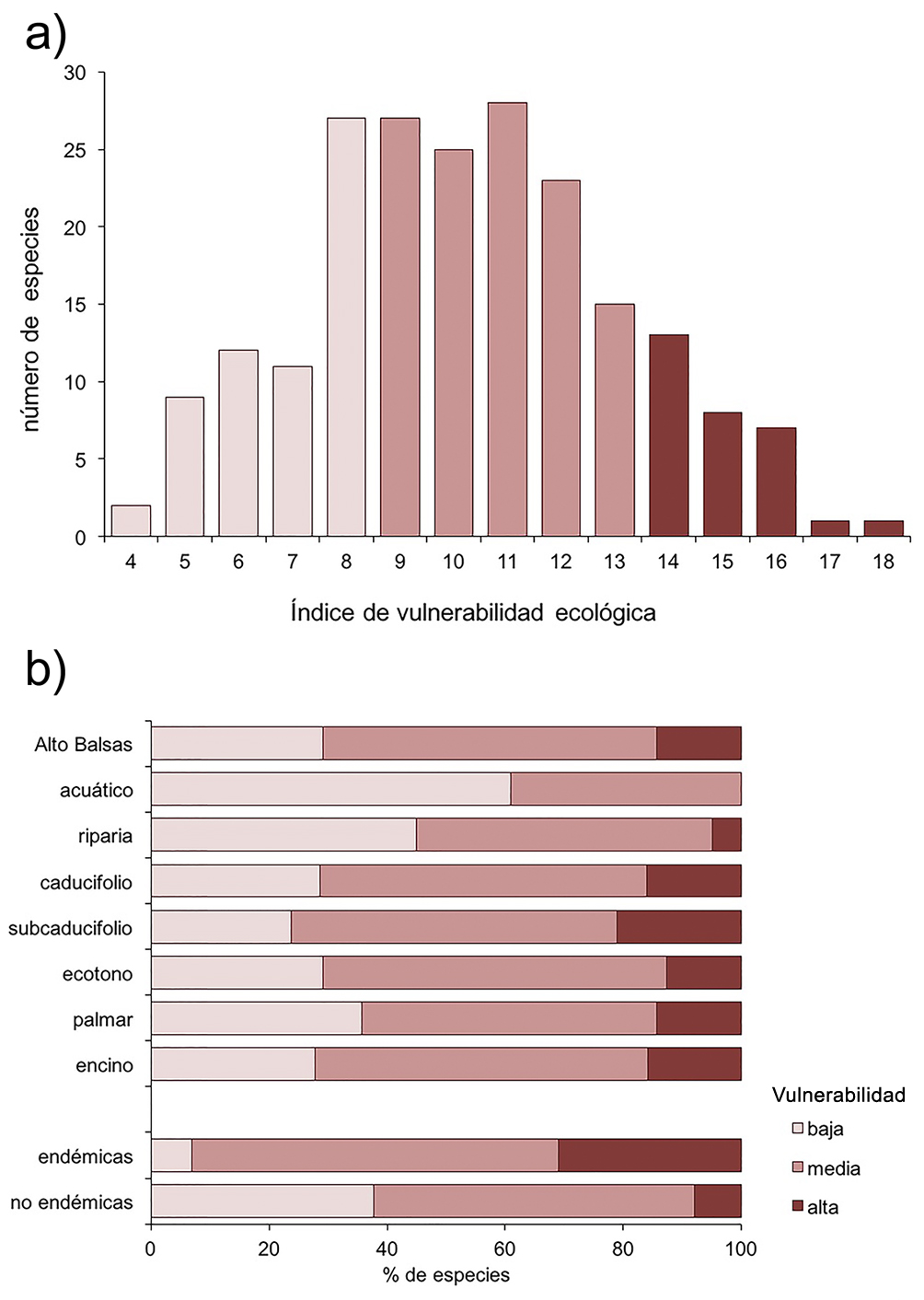
El 29.2% (61) de las especies tuvieron valores de vulnerabilidad baja, 56.5% (118) media y 14.3% (30) alta. (fig. 5a). La vegetación riparia y los ambientes acuáticos tuvieron una menor representación de especies con vulnerabilidad alta. Por otro lado, las aves endémicas tuvieron una mayor proporción de especies con alta vulnerabilidad respecto con las especies de amplia distribución (31% y 7.9% respectivamente; fig. 5b).
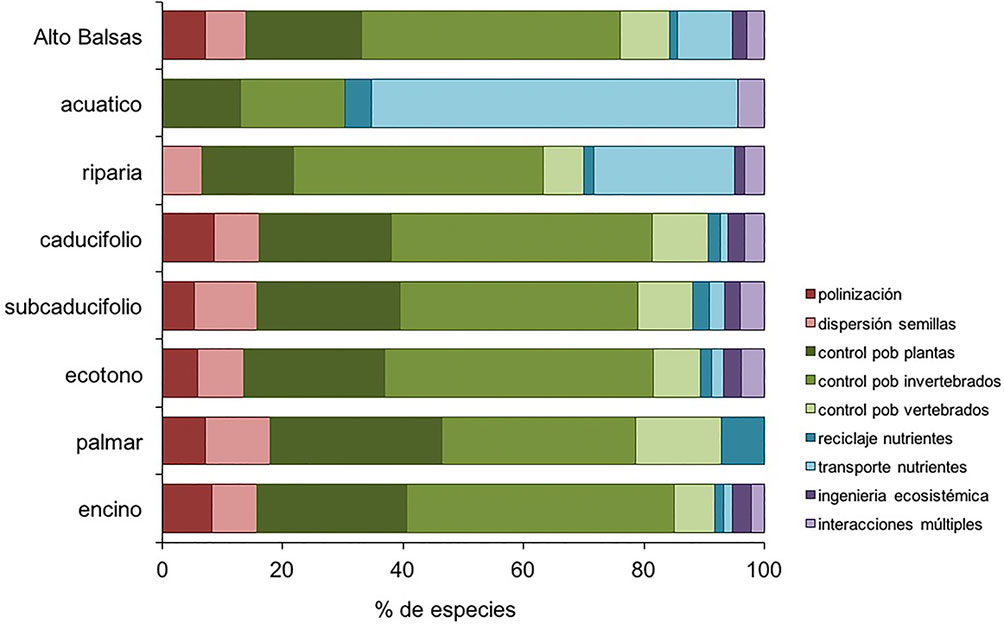
A nivel regional (fig. 6), el atributo de requerimientos ecológicos mejor representado fue la dieta insectívora con 51.2% (105 especies) del total de especies. Le siguen la dieta granívora con 16.1% (33 especies), la carnívora con 10.2% (21 especies), la nectarívora con 6.8% (14 especies), la frugívora y la piscívora, cada una con 5.4% (11 especies), la omnívora con 2.9% (6 especies), carroñera, con 1.5% (3 especies) y la malacófaga con 0.5% (1 especie). Respecto al efecto ecológico, el atributo mejor representado fue el control poblacional de invertebrados, con 43.9% (90 especies), seguido del control poblacional de semillas con 19.0% (39 especies), transporte de nutrientes del ambiente acuático al terrestre con 8.8% (18 especies), control de poblaciones de vertebrados con 8.3% (17 especies), polinización de plantas con 6.8% (14 especies), dispersión de semillas con 6.3% (13 especies), interacciones múltiples con 2.9% (6 especies), ingeniería ecosistémica con 2.4% (5 especies) y reciclaje de cadáveres con 1.5% (3 especies) (fig. 7). En el apéndice 1 se indican los atributos de dieta y vinculación con servicios ecosistémicos considerados para cada especie registrada.
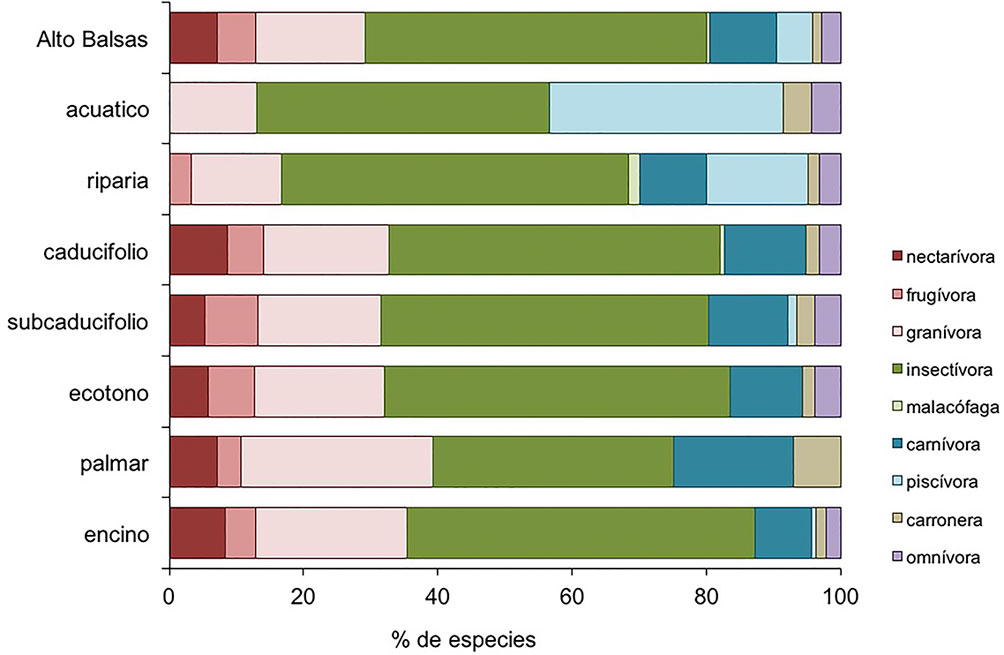
Los ambientes forestales (caducifolio, subcaducifolio, encinar y ecotonos) tuvieron patrones similares en la representación de atributos de requerimientos y efectos ecológicos (fig. 7). En contraste, en el palmar hubo una mayor representación de aves carnívoras y carroñeras y menor representación de frugívoras e insectívoras respecto a los ambientes forestales. Además, no se registraron aves omnívoras ni piscívoras (fig. 7). La dieta malacófaga sólo se registró en el bosque caducifolio y la vegetación riparia.
Respecto a la vinculación con servicios ecosistémicos, el palmar tuvo una mayor representación de los atributos de control poblacional de vertebrados y reciclaje de cadáveres; mientras que los atributos de ingeniería ecosistémica, transporte de nutrientes e interacciones múltiples no fueron registrados (fig. 7). La vegetación riparia y el hábitat acuático fueron diferentes a los demás tipos de hábitat por la baja representación de las dietas frugívora y nectarívora. Los atributos de polinización, dispersión de semillas e ingeniería ecosistémica no estuvieron representados en el hábitat acuático; mientras que la dieta piscívora y del transporte de nutrientes tuvieron una mayor representación respecto a los ambientes forestales (fig. 7).
Discusión
La biodiversidad de aves del Alto Balsas de Guerrero representa aproximadamente el 18% del total nacional, distribuida en el 0.1% del territorio mexicano. Con esto, supera a otras zonas del Alto Balsas (Puebla, 128 especies, Ramírez-Albores, 2007; Morelos, 177 especies, Ramírez-Albores y Ramírez-Cedillo, 2002) y del Balsas Medio (Guerrero, 164 especies, Almazán-Núñez y Navarro, 2006; 124 especies, Rojas-Soto et al., 2009, y 105 especies, Nova-Muñoz et al., 2011). De hecho, la riqueza específica de las aves terrestres del Alto Balsas de Guerrero es similar a la cuenca baja del Balsas (Villaseñor-Gómez et al., 2000) y la región de Chamela (Arizmendi et al., 1990), en la costa occidente de México; región donde los bosques tropicales secos alcanzan su mayor diversidad de aves (Feria-Arroyo y Peterson, 2002; Vega-Rivera et al., 2010).
Esta alta diversidad avifaunística regional se explica, desde el punto de vista ecológico, tanto por la elevada diversidad beta entre el hábitat con influencia Neotropical y Neártica (Espinosa-Organista et al., 2008; Koleff et al., 2008), como por variaciones microclimáticas locales que elevan el recambio entre áreas específicas del bosque tropical seco (Trejo y Dirzo, 2002). La avifauna de los bosques caducifolio y subcaducifolio fue definida por especies tropicales como Megascops seductus, Glaucidium griscomi y Cassiculus melanicterus (Vega-Rivera et al., 2010). En contraste, los bosques de encino, ecotonos y el palmar incorporaron especies típicas de bosques templados con influencia biogeográfica Neártica, como Cyrtonyx montezumae, Glaucidium gnoma, Trogon mexicanus, Baeolophus wollweberi, Melanerpes formicivorus (Ramírez-Bastida et al., 2015). Se trata de un patrón tanto en vertebrados terrestres (Ochoa-Ochoa et al., 2014) como con la flora de los bosques montanos del centro del país (Becerra, 2005; Espinosa-Organista et al., 2008; Rzedowski et al., 2005; Valencia, 2004). Por otro lado, las aves dependientes de los ambientes acuáticos (17 especies) representaron el 32% del recambio taxonómico regional. Esto contrasta con el bajo porcentaje de la riqueza específica regional que representan (7.3%). Sin embargo, considerando la baja eficiencia del muestreo en el hábitat acuático, es esperable que su diverisidad alfa aumente a casi 40 especies (Ramírez-Bastida et al., 2008), aumentando también la diversidad beta y gamma regionales.
En contraste, el bosque subcaducifolio, los ecotonos forestales y el palmar tuvieron pocas especies únicas, dado al uso del hábitat forestal como un continuo (Ramírez-Bastida et al., 2015; Vega-Rivera et al., 2010). Ortalis poliocephala, Ciccaba virgata, Xiphorhynchus flavigaster y Calocitta formosa son ejemplos de aves tropicales (Stotz et al., 1996) que usaron los ecotonos entre bosques tropical y de encino en tierras altas (tabla 2). Esto influyó en que el bosque subcaducifolio, los ecotonos forestales y el palmar tuvieran pocas especies únicas.
De las 78 migratorias de larga distancia registradas, 65 son aves terrestres. Esta riqueza ha sido explicada por similitudes ecológicas entre el gradiente altitudinal del centro occidente de México y el hábitat reproductivo de las aves. Con ello, el Alto Balsas de Guerrero es una de las zonas con mayor riqueza de migratorias en el Neotrópico (Hutto, 1992, 2010).
El componente migratorio influyó en el recambio taxonómico debido al uso diferencial del gradiente de altitud de algunas especies (Hutto, 1986). Setophaga petechia se registró en bosques tropicales; mientras que Setophaga townsendi se registró en bosques de encino y ecotonos. Las aves Charadiiformes (chorlos y playeros) y Pelecaniformes (garzas) migratorias fueron muy importantes para el hábitat acuático y la vegetación riparia, un patrón común en los ambientes ribereños de México (Navarro-Sigüenza et al., 2014).
Los movimientos locales asociados con la disponibilidad de recursos influyen en la composición y recambio de la avifauna (Vega-Rivera et al., 2010). Ara militaris se desplaza de los bosques tropicales a los ecotonos y bosques de encino para alimentarse entre noviembre y febrero (Naturam Sequi, 2016). Chloroceryle americana, Sayornis nigricans y Parkesia motacilla se internan en las barrancas del bosque subcaducifolio para alimentarse de peces e invertebrados acuáticos durante la temporada de lluvias, cuando la escorrentía forma cauces temporales. Finalmente, Amazilia viridifrons e Hylocharis leucotis, características de bosques tropicales y encinares respectivamente (Ornelas y Arizmendi, 1995; Vega-Rivera et al., 2010; Ramírez-Bastida et al., 2015) fueron registrados en extremos opuestos de su distribución altitudinal (ver apéndice), posiblemente buscando recursos alimenticios (Arizmendi y Ornelas, 1990; Lara, 2006; Vega-Rivera et al., 2010). La información acerca de movimientos locales es clave para entender la ecología de los bosques que frecuentemente es omitido en estudios descriptivos (Boyle, 2017).
En cuanto a endemismos e identidad biológica mexicana, la riqueza de especies endémicas registrada hace del Alto Balsas un área prioritaria para la conservación a escala global (Arizmendi, 2002; Bertelli et al., 2017; Navarro-Sigüenza y Peterson, 2004; Peterson y Navarro-Sigüenza, 2000). En términos históricos, la evolución de endemismos en los bosques tropicales secos del occidente de México se explica por varios factores: el efecto de aislamiento que la Faja Volcánica Transmexicana y la sierra Madre del Sur producen sobre la cuenca del Balsas (Becerra, 2005; Morrone, 2014); el levantamiento del istmo de Tehuantepec durante el Mioceno (Mason et al., 2018; Navarro-Sigüenza et al., 2017) y los cambios paleoclimáticos derivados del avance y retroceso de glaciares del Pleistoceno (Arbeláez-Cortés, Milá et al., 2014; Arbeláez-Cortés, Roldán-Piña et al., 2014; Montaño-Rendón et al., 2015; Navarro-Sigüenza et al., 2017). En conjunto, estos procesos moldearon la distribución de los bosques templados y tropicales, generando efectos de aislamiento y contacto periódicos que elevaron las tasas de diversificación de las aves (Lovette, 2005). Como consecuencia, los actuales bosques tropicales caducifolios y de encino del centro-occidente de México están entre los ambientes con mayor riqueza de endemismos. De esta forma, procesos históricos y ecológicos definieron la identidad de una de las avifaunas evolutivamente más mexicanas que existen (Navarro-Sigüenza et al., 2014).
La incidencia simpátrica de especies diagnósticas para diferentes provincias biogeográficas y áreas de endemismo señala al Alto Balsas de Guerrero como un área de conectividad en el centro de México, aumentando su relevancia en la planeación de estrategias de conservación (Peterson y Navarro-Sigüenza, 2000). A escala del Neotrópico, Megascops seductus, Campylorhynchus jocosus, Cynanthus sordidus y Peucaea humeralis identifican a la provincia cuenca del Balsas (Morrone, 2014). Por otro lado, endémicas y cuasiendémicas que habitan tierras altas, como Cyrtonyx sallaei, Glaucidium gnoma, Lepidocolaptes leucogaster, Ptiliogonys cinereus y Piranga hepatica son evidencia de contacto con los bosques templados de la Zona de Transición Mexicana (Morrone, 2014; Ramírez-Bastida et al., 2015). Amazilia viridifrons (Vega-Rivera et al., 2010) y Cardinalis cardinalis (Ramírez-Albores, 2010) indican conectividad con la Sierra Madre del Sur y la costa del Pacífico. Además, aves tropicales (Nyctibius jamaicensis, Chondrohierax uncinatus, Buteogallus urubitinga y Ara militaris) y costeras (Dendrocygna autumnalis, Mycteria americana y Megaceryle torquata) indican la conectividad entre las Tierras Bajas del Pacífico y el centro del país (Jiménez-Arcos et al., 2012; Ochoa-Arteaga et al., 2016; Vega-Rivera et al., 2010; Urbina-Torres, 2016).
En lo referente a la identidad funcional, aunque el empleo de indicadores simples tiene limitaciones en la interpretación de la funcionalidad ecológica asociada a las aves (Luck et al., 2012), los resultados obtenidos muestran que las aves hacen un importante aporte a los procesos de regulación, soporte ecológico y resiliencia de los sistemas naturales en el Alto Balsas (Fischer et al., 2007; Sodhi et al., 2011; Wenny et al., 2011).
En primer lugar, participan en la dinámica forestal, pues funcionan como conectores genéticos que influyen en el éxito reproductivo de las especies vegetales al influir en la polinización, mediada principalmente por la familia Trochilidae (Arizmendi y Rodríguez-Flores, 2012). También se vinculan con el reclutamiento poblacional de especies vegetales al dispersar semillas. Los mosqueros Myiarchus son los principales dispersores para Bursera, un género clave del hábitat forestal regional (Almazán-Núñez et al., 2016; Ramos-Ordoñez y Arizmendi, 2011). Numerosas especies funcionan como conectores en procesos de regulación ecosistémica, a través del atributo de control poblacional (Sodhi et al., 2011). La depredación de semillas puede moldear el reclutamiento de las plantas que las aves consumen, por ejemplo, Ara militaris sobre árboles del dosel y cactáceas columnares (Contreras-González y Arizmendi, 2014); además, las aves depredadoras de animales son un componente importante de las cascadas tróficas que regulan el flujo de materia y energía (Şekercioğlu et al., 2004; Sodhi et al., 2011). Particularmente, la mayor representación del atributo de control de poblaciones de vertebrados registrada en el palmar podría implicar una presión de depredación diferencial mediada por aves, respecto a los bosques tropicales o de encino. Esto podría explicarse con la menor cobertura vegetal del palmar, que aumenta la visibilidad de las presas y el éxito de captura (Dickman et al., 1991; Pietrek et al., 2009). Por otro lado, los pájaros carpinteros (familia Picidae) son importantes como ingenieros ecositémicos, al generar cavidades en el hábitat forestal (Cockle et al., 2011). Cathartes aura, Coragyps atratus y Caracara cheriway aportan el servicio de regulación ecológica de reciclaje de cadáveres, reduciendo la posibilidad de brotes patógenos y reintegrando materia orgánica al flujo de energía del ecosistema (Şekercioğlu et al., 2004). Finalmente, las aves acuáticas representan un vínculo importante entre el hábitat acuático y el terrestre, transportando nutrientes hacia el hábitat forestal (Şekercioğlu, 2004). Al involucrarse estrechamente en la provisión de servicios ecosistémicos, la biodiversidad de aves es importante para el bienestar de las poblaciones humanas (Cardinale et al., 2012; Díaz et al., 2004).
La principal amenaza para las aves en el Alto Balsas es la pérdida de hábitat (Vega-Rivera et al., 2010; Maass et al., 2010). Aunque la región mantiene extensiones forestales conservadas, al menos el 18% de la cobertura vegetal ha cambiado por suelos agropecuarios y asentamientos humanos (Naturam Sequi, 2016). Esto amenaza particularmente a las especies de distribución retringuida (McKinney y Lockwood, 1999; NABCI, 2016).
Megascops seductus y Glaucidium griscomi son endémicas que dependen de cavidades distribuidas en áreas específicas del bosque subcaducifolio (Monterrubio-Rico y Escalante-Pliego, 2006; Vázquez-Reyes y Renton, 2015). El Mosquero del Balsas, Xenotriccus mexicanus es un ave muy poco conocida y amenazada por la deforestación (Vega-Rivera et al., 2010). Migratorias como Contopus cooperi, Vireo bellii y Passerina ciris también enfrentan esta amenaza (Berlanga et al., 2010). Accipitriformes, Falconiformes y Strigiformes suman el 48% de las especies protegidas por la legislación nacional debido a la pérdida de hábitat y contaminación con residuos químicos (Enríquez y Vázquez-Pérez, 2015; Thiollay, 2017). Además, la presión humana directa también afecta a las poblaciones de aves (Naranjo y Cuarón, 2010). La captura ilegal y la pérdida de hábitat (Rivera-Ortíz et al., 2013) han colocado a Ara militaris en peligro de extinción, y amenazan a otras especies.
Las especies invasoras como Columba livia, Streptopelia decaocto (una invasión reciente en la región, Blancas-Calva et al., 2014) y Passer domesticus deben su presencia a las actividades humanas que aumentan artificialmente la conectividad biogeográfica (Hulme, 2009). Estas aves son un factor de riesgo al provocar exclusión competitiva y funcionar como vectores de enfermedades (Simberloff et al., 2013). Thraupis episcopus, se consideró un visitante accidental que podría haber sido traslocado por huracanes desde su área de distribución en el golfo de México (Faaborg y Gauthreax, 2014). Sin embargo, hay registros de T. episcopus usando ambientes perturbados en la costa de Guerrero (Blancas-Calva et al., 2017), por lo que podría tratarse de un proceso incipiente de dispersión facilitado por la transformación del hábitat (Rodríguez-Ruíz et al. 2011).
Estudios previos han mostrado que la perturbación antropogénica en el Alto Balsas ha impulsado tanto la pérdida local de especies nativas, como la incidencia de invasiones biológicas, erosionando las diferencias taxonómicas entre las avifaunas de los bosques tropicales caducifolios y de encino (Vázquez-Reyes et al., 2017). Este fenómeno, conocido como homogeneización biótica, es un componente clave de la crisis global de biodiversidad que afecta a la región (McGill et al., 2015).
Históricamente, la cuenca del Balsas es una de las áreas con menor inversión para conservación en el país (Bezaury, 2010). Aunque el registro de Ara militaris (Jiménez-Arcos et al., 2012) despertó el interés por la región (Naturam Sequi, 2015, 2016) y fue designada IBA (MX264, Birdlife International, 2017), este nombramiento es insuficiente para frenar la pérdida de biodiversidad. La conservación de la biodiversidad del Alto Balsas depende de la inversión de recursos para investigación, conservación y restauración de la cobertura forestal, mantenimiento de la funcionalidad ecosistémica y desarrollo de manejo agropecuario sustentable (Chazdon et al., 2009; List et al., 2017; Peterson y Navarro-Sigüenza, 2016; Prieto-Torres et al., 2016). Será indispensable involucrar la complejidad sociocultural de la región, altamente marginada, y hasta ahora dependiente de la agricultura y la ganadería (Good-Eshelman y Barrientos-López, 2004). Una opción para reducir la presión sobre los sistemas naturales es el eventual desarrollo de ecoturismo comunitario basado en la observación de aves (Peterson y Navarro-Sigüenza, 2000). Sin embargo, antes será necesario lograr que la población conozca y apropie a la biodiversidad como un recurso que debe ser conservado, así como garantizar que las actividades económicas beneficien específicamente a las comunidades locales (Sánchez-González y Ortega-Álvarez, 2015).
Finalmente, aunque es necesario desarrollar estudios de otros grupos taxonómicos, nuestros resultados indican que el Alto Balsas de Guerrero cumple criterios para la designación de reserva de la biosfera (LGEEPA, 2017; UNESCO, 2017). Posee una complejidad funcional relevante a través de un gradiente ecológico y altitudinal; conectividad entre provincias biogeográficas, una notable representación de taxa endémicos y amenazados, así como una elevada diversidad beta, todo dentro de un área mayor a 200,000 ha. Obtener dicho nombramiento favorecería la creación de un marco normativo que conjugue a comunidades locales, academia, ONGs y al sector gubernamental en el manejo sustentable y conservación de su patrimonio biológico.
Agradecimientos
LDV-R y VHJ-A contaron con apoyo de las becas Conacyt 220265 y 377215 durante sus estudios de doctorado en el Posgrado en Ciencias Biológicas de la UNAM. El trabajo de campo fue financiado por los proyectos Conacyt 152060-B (AGN-S); Conanp PROCER/DRCEN/003/2015 y PROCER/DRCEN/06/2016 (LDV-R, Naturam Sequi); Naturalista Conabio (Naturam Sequi), así como de los apoyos PAEP 2014 y 2016 del PCBiol UNAM (LDV-R). La Secretaría de Medio Ambiente y Recursos Naturales otorgó el permiso de colecta científica FAUT-0169. Agradecemos a las autoridades locales de Papalutla, Mezquitlán, Copalillo, Zicapa, Atlapa, Xixila, Tomatepec y Cohetzala por las facilidades brindadas. Juan Esteban, Josué Esteban, Mario Flores, José Enrique Rosendo, René Rosendo, Richard Flores, Eliseo Sánchez, los miembros del Comisariado de Bienes Ejidales de Zicapa, así como estudiantes del Taller de Biodiversidad y Sistemática de Vertebrados Terrestres de la Facultad de Ciencias y del Departamento de Zoología de la FES Iztacala, quienes realizaron valioso trabajo de campo. Agradecemos a Justino Morales, Josefina Pablo de Aquino, Isabel Pablo de Aquino, Adalberto Rosendo, Celso Rosendo, José María Rosendo y Juan Cortés por el alojamiento y valiosas facilidades. Alejandro Gordillo, Fanny Rebón, César Ríos, Marco F. Ortiz, Violeta Andrade, Alfonsina Hernández y Fernando León apoyaron en la consulta de ejemplares de la Colección del Aves del MZFC. Luis Enrique Sánchez desarrolló el mapa de la zona de estudio. Raúl Cueva (FES Iztacala), Fausto Méndez (Instituto de Biología), Roberto Saldaña (Biosphera Picture A.C.), Ociel Ramos Ramírez, la (Dirección Regional Centro y Eje Neovolcánico de la Conanp, la S.C. ANP Cerro Tecaballo) y Bernardo Rosendo (ICAT Olinalá) brindaron valioso apoyo logístico. Héctor Godínez, Patricia Ramírez, Horacio Paz, Luis A. Sánchez, Ek de Val y dos revisores anónimos aportaron valiosas críticas que mejoraron el manuscrito.
|
Apéndice. Inventario de las aves del Alto Balsas de Guerrero. Arreglo taxonómico supraespecífico de acuerdo con la AOS (2017). Los nombres específicos corresponden a la taxonomía de las aves de México de Navarro-Sigüenza y Peterson (2004). Las especies exóticas se señalan delante del nombre científico. Tipos de hábitat = ACU: ambiente acuático, RIP: vegetación riparia, BTC: bosque tropical caducifolio, BTS: bosque tropical subcaducifolio, EET: ecotonos encinar-bosques tropicales, PAL: palmar, BE: bosque de encino. Estatus de residencia = Rp: residente permanente, Mloc: migratoria local, Mv: migratoria de verano, T: transitoria, Acc: accidental. Endemismo = END: endémica, CUA: cuasiendémica, SEMI: semiendémica. Dieta: NEC: nectarívora, FRU: frugívora, GRA: granívora, CARN: carnívora, INS: insectívora, MALA: malacófaga, PISC: piscívora, CARR: carroñera, OMN: omnívora. Vinculación servicios ecosistémicos = POL: polinización, DIS. SEM.: dispersión de semillas, C. POB. PLA.: control poblacional de plantas, C. POB. INV.: control poblacional de invertebrados, C. POB. VERT.: control poblacional de vertebrados, ING ECO: ingeniería ecosistémica, REC. NUTR.: reciclaje de nutrientes, TRANS NUTR.: transporte de nutrientes, INT. MULT.: interacciones ecológicas múltiples. NOM-059 = sc: sin clasificación, Pr: sujeta a protección especial, A: amenazada, P: en peligro de extinción. IUCN (Birdlife International, 2018) = LC: least concern (preocupación menor), NT: near threatened (casi amenazada), VU: vulnerable (vulnerable). NABCI (2016)= índice de vulnerabilidad ecológica, con valores de 4 a 18. |
|||||||||||
|
Taxones |
ACU |
RIP |
BTC |
BTS |
EET |
PAL |
BE |
RES. |
END. |
DIETA / SERV. ECO. |
NOM-059/IUCN/NABCI |
|
ANSERIFORMES |
|||||||||||
|
Anatidae |
|||||||||||
|
Dendrocygna autumnalis |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 8 |
|||||||
|
GALLIFORMES |
|||||||||||
|
Cracidae |
|||||||||||
|
Ortalis poliocephala |
X |
X |
X |
X |
X |
Rp |
END |
FRU / DIS. SEM. |
sc / LC / 15 |
||
|
Odontophoridae |
|||||||||||
|
Philortyx fasciatus |
X |
X |
X |
Rp |
END |
GRA / C. POB. PLA. |
sc / LC / 12 |
||||
|
Cyrtonyx sallei |
X |
Rp |
END |
GRA / C. POB. PLA. |
Pr / LC / 13 |
||||||
|
PODICIPEDIFORMES |
|||||||||||
|
Podicipedidae |
|||||||||||
|
Tachybaptus dominicus |
X |
Mloc |
INS / TRANS. NUTR. |
Pr / LC / 8 |
|||||||
|
COLUMBIFORMES |
|||||||||||
|
Columbidae |
|||||||||||
|
Columba livia exótica |
X |
X |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 6 |
|||||
|
Streptopelia decaocto exótica |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 5 |
|||||||
|
Columbina inca |
X |
X |
X |
X |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 8 |
|||
|
Columbina passerina |
X |
X |
X |
X |
X |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 9 |
||
|
Leptotila verreauxi |
X |
X |
X |
X |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 8 |
|||
|
Zenaida asiatica |
X |
X |
X |
X |
X |
Mloc |
GRA / C. POB. PLA. |
sc / LC / 8 |
|||
|
Zenaida macroura |
X |
X |
X |
X |
Mloc |
GRA / C. POB. PLA. |
sc / LC / 5 |
||||
|
CUCULIFORMES |
|||||||||||
|
Cuculidae |
|||||||||||
|
Piaya mexicana |
X |
X |
X |
X |
X |
X |
Rp |
END |
INS / C. P. INV |
sc / LC / 9 |
|
|
Morococcyx erythropygus |
X |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 14 |
||||
|
Geococcyx velox |
X |
X |
X |
X |
Rp |
CARN / C. P. INV |
sc / LC / 10 |
||||
|
Crotophaga sulcirostris |
X |
X |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 7 |
|||
|
CAPRIMULGIFORMES |
|||||||||||
|
Caprimulgidae |
|||||||||||
|
Chordeiles acutipennis |
X |
Mi |
INS / C. P. INV |
sc / LC / 8 |
|||||||
|
Chordeiles minor |
X |
Mv |
INS / C. P. INV |
sc / LC / 11 |
|||||||
|
Antrostomus ridgwayi |
X |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 12 |
||||
|
NYCTIBIIFORMES |
|||||||||||
|
Nyctiibidae |
|||||||||||
|
Nyctibius jamaicensis |
X |
Rp |
INS / C. P. INV |
sc / LC / 14 |
|||||||
|
APODIFORMES |
|||||||||||
|
Apodidae |
|||||||||||
|
Aeronautes saxatalis |
X |
Rp |
INS / C. P. INV |
sc / LC / 11 |
|||||||
|
Trochilidae |
|||||||||||
|
Heliomaster constantii |
X |
X |
X |
X |
Mloc |
NEC / POL |
sc / LC / 12 |
||||
|
Tilmatura dupontii |
X |
Rp |
NEC / POL |
A / LC / 16 |
|||||||
|
Calothorax lucifer |
X |
Mi |
CUA |
NEC / POL |
sc / LC / 14 |
||||||
|
Calothorax pulcher |
X |
X |
Rp |
END |
NEC / POL |
sc / LC / 16 |
|||||
|
Archilochus colubris |
X |
X |
X |
X |
T |
NEC / POL |
sc / LC / 8 |
||||
|
Archilochus alexandri |
X |
X |
X |
T |
CUA |
NEC / POL |
sc / LC / 11 |
||||
|
Selasphorus rufus |
X |
X |
Mi |
NEC / POL |
sc / LC / 13 |
||||||
|
Selasphorus sasin |
X |
Mi |
CUA |
NEC / POL |
sc / LC / 15 |
||||||
|
Chlorostilbon auriceps |
X |
X |
Rp |
END |
NEC / POL |
sc / LC / 14 |
|||||
|
Cynanthus sordidus |
X |
X |
X |
X |
X |
Rp |
END |
NEC / POL |
sc / LC / 12 |
||
|
Cynanthus latirostris |
X |
Rp |
CUA |
NEC / POL |
sc / LC / 10 |
||||||
|
Amazilia beryllina |
X |
X |
Mloc |
END |
NEC / POL |
sc / LC / 10 |
|||||
|
Amazilia violiceps |
X |
X |
X |
X |
Rp |
CUA |
NEC / POL |
sc / LC / 10 |
|||
|
Amazilia viridifrons |
X |
X |
Mloc |
END |
NEC / POL |
A / LC / 14 |
|||||
|
Hylocharis leucotis |
X |
X |
Mloc |
NEC / POL |
sc / LC / 11 |
||||||
|
CHARADIIFORMES |
|||||||||||
|
Recurvirostridae |
|||||||||||
|
Himantopus mexicanus |
X |
Rp |
INS / TRANS. NUTR. |
sc / LC / 11 |
|||||||
|
Charadriidae |
|||||||||||
|
Charadrius collaris |
X |
Rp |
INS / TRANS. NUTR. |
sc / LC / 12 |
|||||||
|
Charadrius vociferus |
X |
X |
Rp |
INS / TRANS. NUTR. |
sc / LC / 9 |
||||||
|
Scolopacidae |
|||||||||||
|
Actitis macularius |
X |
X |
Mi |
INS / TRANS. NUTR. |
sc / LC / 9 |
||||||
|
CICONIIFORMES |
|||||||||||
|
Ciconiidae |
|||||||||||
|
Mycteria americana |
X |
T |
PISC / TRANS. NUTR. |
Pr / LC / 13 |
|||||||
|
SULIFORMES |
|||||||||||
|
Phalacrocoracidae |
|||||||||||
|
Phalacrocorax brasilianus |
X |
Rp |
PISC / TRANS. NUTR. |
sc / LC / 8 |
|||||||
|
PELECANIFORMES |
|||||||||||
|
Ardeidae |
|||||||||||
|
Ardea herodias |
X |
X |
Mi |
PISC / TRANS. NUTR. |
sc / LC / 7 |
||||||
|
Ardea alba |
X |
X |
Mi |
PISC / TRANS. NUTR. |
sc / LC / 7 |
||||||
|
Egretta thula |
X |
X |
Mi |
PISC / TRANS. NUTR. |
sc / LC / 8 |
||||||
|
Butorides virescens |
X |
X |
Mi |
PISC / TRANS. NUTR. |
sc / LC / 11 |
||||||
|
Nycticorax nycticorax |
X |
X |
Mi |
PISC / TRANS. NUTR. |
sc / LC / 10 |
||||||
|
CATHARTIFORMES |
|||||||||||
|
Cathartidae |
|||||||||||
|
Coragyps atratus |
X |
X |
X |
X |
X |
Rp |
CARR / REC. NUTR. |
sc / LC / 5 |
|||
|
Cathartes aura |
X |
X |
X |
X |
X |
X |
Rp |
CARR / REC. NUTR. |
sc / LC / 5 |
||
|
ACCIPITRIFORMES |
|||||||||||
|
Pandionidae |
|||||||||||
|
Pandion haliaetus |
X |
X |
Mi |
PISC / TRANS. NUTR. |
sc / LC / 7 |
||||||
|
Accipitridae |
|||||||||||
|
Chondrohierax uncinatus |
X |
X |
Rp |
MALA / C. P. INV |
Pr / LC / 12 |
||||||
|
Accipiter striatus |
X |
X |
X |
Mi |
CARN / C. POB. VERT |
Pr / LC / 7 |
|||||
|
Accipiter cooperii |
X |
X |
X |
X |
Mi |
CARN / C. POB. VERT |
Pr / LC / 8 |
||||
|
Buteogallus anthracinus |
X |
X |
X |
X |
X |
Rp |
CARN / TRANS. NUTR. |
Pr / LC / 11 |
|||
|
Buteogallus urubitinga |
X |
X |
Rp |
CARN / TRANS. NUTR. |
Pr / LC / 12 |
||||||
|
Geranoaetus albicaudatus |
X |
X |
X |
X |
Rp |
CARN / C. POB. VERT |
Pr / LC / 10 |
||||
|
Buteo plagiatus |
X |
X |
X |
Rp |
CARN / C. POB. VERT |
sc / LC / 8 |
|||||
|
Buteo lineatus |
X |
Mi |
CARN / C. POB. VERT |
Pr / LC / 8 |
|||||||
|
Buteo platypterus |
X |
Mi |
CARN / C. POB. VERT |
Pr / LC / 8 |
|||||||
|
Buteo brachyurus |
X |
Rp |
CARN / C. POB. VERT |
sc / LC / 10 |
|||||||
|
Buteo albonotatus |
X |
X |
X |
Mi |
CARN / C. POB. VERT |
Pr / LC / 10 |
|||||
|
Buteo jamaicensis |
X |
X |
X |
X |
X |
Rp |
CARN / C. POB. VERT |
sc / LC / 6 |
|||
|
STRIGIFORMES |
|||||||||||
|
Tytonidae |
|||||||||||
|
Tyto alba |
X |
X |
X |
X |
Rp |
CARN / C. POB. VERT |
sc / LC / 9 |
||||
|
Strigidae |
|||||||||||
|
Megascops seductus |
X |
X |
Rp |
END |
INS / C. P. INV |
A / NT / 17 |
|||||
|
Bubo virginianus |
X |
X |
X |
X |
Rp |
CARN / C. POB. VERT |
sc / LC / 6 |
||||
|
Glaucidium gnoma |
X |
Rp |
CUA |
CARN / C. POB. VERT |
sc / LC / 11 |
||||||
|
Glaucidium griscomi |
X |
X |
X |
X |
Rp |
END |
CARN / C. POB. VERT |
A / LC / 16 |
|||
|
Glaucidium brasilianum |
X |
Rp |
CARN / C. POB. VERT |
sc / LC / 8 |
|||||||
|
Micrathene whitneyi |
X |
X |
Mi |
CUA |
INS / C. P. INV |
sc / LC / 13 |
|||||
|
Athene cunicularia |
X |
Mi |
INS / C. P. INV |
sc / LC / 12 |
|||||||
|
Ciccaba virgata |
X |
X |
X |
Rp |
CARN / C. P. INV |
sc / LC / 11 |
|||||
|
TROGONIFORMES |
|||||||||||
|
Trogonidae |
|||||||||||
|
Trogon ambiguus |
X |
X |
X |
Rp |
CUA |
FRU / DIS. SEM. |
sc / LC / 14 |
||||
|
Trogon mexicanus |
X |
Rp |
FRU / DIS. SEM. |
sc / LC / 14 |
|||||||
|
CORACIIFORMES |
|||||||||||
|
Momotidae |
|||||||||||
|
Momotus mexicanus |
X |
X |
X |
X |
X |
Rp |
CUA |
INS / C. P. INV |
sc / LC / 12 |
||
|
Alcedinidae |
|||||||||||
|
Megaceryle torquata |
X |
Rp |
PISC / TRANS. NUTR. |
sc / LC / 8 |
|||||||
|
Megaceryle alcyon |
X |
X |
Mi |
PISC / TRANS. NUTR. |
sc / LC / 11 |
||||||
|
Chloroceryle americana |
X |
X |
Rp |
PISC / TRANS. NUTR. |
sc / LC / 10 |
||||||
|
PICIFORMES |
|||||||||||
|
Picidae |
|||||||||||
|
Melanerpes formicivorus |
X |
X |
Rp |
INS / ING ECO |
sc / LC / 9 |
||||||
|
Melanerpes chrysogenys |
X |
X |
X |
X |
X |
Rp |
END |
INS / ING ECO |
sc / LC / 12 |
||
|
Melanerpes hypopolius |
X |
X |
Rp |
END |
INS / ING ECO |
sc / LC / 16 |
|||||
|
Picoides scalaris |
X |
X |
X |
Rp |
INS / ING ECO |
sc / LC / 9 |
|||||
|
Campephilus guatemalensis |
X |
X |
Rp |
INS / ING ECO |
Pr / LC / 14 |
||||||
|
FALCONIFORMES |
|||||||||||
|
Falconidae |
|||||||||||
|
Herpetotheres cachinnans |
X |
X |
Rp |
CARN / C. POB. VERT |
sc / LC / 10 |
||||||
|
Caracara cheriway |
X |
X |
Rp |
CARR / REC. NUTR. |
sc / LC / 8 |
||||||
|
Falco sparverius |
X |
X |
X |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 11 |
||
|
Falco columbarius |
X |
Mi |
CARN / C. POB. VERT |
sc / LC / 7 |
|||||||
|
Falco peregrinus |
X |
X |
X |
X |
Mi |
CARN / C. POB. VERT |
Pr / LC / 9 |
||||
|
PSITTACIFORMES |
|||||||||||
|
Psittacidae |
|||||||||||
|
Eupsittula canicularis |
X |
X |
X |
Rp |
FRU / C. POB. PLA. |
Pr / LC / 14 |
|||||
|
Ara militaris |
X |
X |
X |
X |
Rp |
FRU / C. POB. PLA. |
P / VU / 18 |
||||
|
PASSERIFORMES |
|||||||||||
|
Furnariidae |
|||||||||||
|
Xiphorhynchus flavigaster |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 13 |
|||||
|
Lepidocolaptes leucogaster |
X |
X |
Rp |
END |
INS / C. P. INV |
sc / LC / 15 |
|||||
|
Tyrannidae |
|||||||||||
|
Camptostoma imberbe |
X |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 10 |
||||
|
Myiopagis viridicata |
X |
X |
X |
Mloc |
INS / C. P. INV |
sc / LC / 11 |
|||||
|
Xenotriccus mexicanus |
X |
X |
X |
Rp |
END |
INS / C. P. INV |
Pr / NT / 16 |
||||
|
Contopus cooperi |
X |
X |
T |
INS / C. P. INV |
sc / NT / 13 |
||||||
|
Contopus pertinax |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 12 |
|||||
|
Contopus sordidulus |
X |
X |
X |
X |
T |
INS / C. P. INV |
sc / LC / 11 |
||||
|
Empidonax traillii |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 10 |
||||||
|
Empidonax minimus |
X |
Mi |
INS / C. P. INV |
sc / LC / 10 |
|||||||
|
Empidonax wrightii |
X |
X |
Mi |
CUA |
INS / C. P. INV |
sc / LC / 9 |
|||||
|
Empidonax oberholseri |
X |
X |
Mi |
CUA |
INS / C. P. INV |
sc / LC / 11 |
|||||
|
Empidonax affinis |
X |
Rp |
CUA |
INS / C. P. INV |
sc / LC / 13 |
||||||
|
Empidonax occidentalis |
X |
X |
X |
Rp |
CUA |
INS / C. P. INV |
sc / LC / 11 |
||||
|
Sayornis nigricans |
X |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 9 |
||||
|
Pyrocephalus rubinus |
X |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 5 |
||||
|
Myiarchus tuberculifer |
X |
X |
X |
X |
Mloc |
INS / DIS. SEM. |
sc / LC / 9 |
||||
|
Myiarchus cinerascens |
X |
X |
Mi |
INS / DIS. SEM. |
sc / LC / 9 |
||||||
|
Myiarchus nuttingi |
X |
X |
X |
X |
X |
Rp |
INS / DIS. SEM. |
sc / LC / 13 |
|||
|
Myiarchus tyrannulus |
X |
X |
X |
Mloc |
INS / DIS. SEM. |
sc / LC / 9 |
|||||
|
Pitangus sulphuratus |
X |
X |
X |
X |
Rp |
OMN / INT. MULT |
sc / LC / 5 |
||||
|
Myiozetetes similis |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 5 |
||||||
|
Myiodynastes luteiventris |
X |
X |
Mv |
INS / C. P. INV |
sc / LC / 11 |
||||||
|
Tyrannus melancholicus |
X |
X |
X |
X |
X |
X |
Rp |
INS / DIS. SEM. |
sc / LC / 4 |
||
|
Tyrannus vociferans |
X |
X |
Mloc |
CUA |
INS / DIS. SEM. |
sc / LC / 9 |
|||||
|
Tyrannus crassirostris |
X |
X |
X |
X |
Rp |
CUA |
INS / DIS. SEM. |
sc / LC / 12 |
|||
|
Tyrannus verticalis |
X |
X |
X |
X |
X |
Mi |
INS / DIS. SEM. |
sc / LC / 9 |
|||
|
Tityridae |
|||||||||||
|
Pachyramphus albiventris |
X |
X |
Rp |
CUA |
INS / C. P. INV |
sc / LC / 11 |
|||||
|
Laniidae |
|||||||||||
|
Lanius ludovicianus |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 11 |
||||||
|
Vireonidae |
|||||||||||
|
Vireo hypochryseus |
X |
X |
Mloc |
END |
INS / C. P. INV |
sc / LC / 13 |
|||||
|
Vireo bellii |
X |
T |
INS / C. P. INV |
sc / NT / 15 |
|||||||
|
Vireo cassini |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 14 |
||||||
|
Vireo plumbeus |
X |
Mi |
INS / C. P. INV |
sc / LC / 10 |
|||||||
|
Vireo swainsonii |
X |
Mi |
INS / C. P. INV |
sc / LC / 8 |
|||||||
|
Corvidae |
|||||||||||
|
Calocitta formosa |
X |
X |
X |
Rp |
OMN / INT. MULT |
sc / LC / 12 |
|||||
|
Aphelocoma woodhouseii |
X |
X |
Rp |
OMN / INT. MULT |
sc / LC / 9 |
||||||
|
Corvus sinuatus |
X |
X |
X |
X |
Rp |
OMN / INT. MULT |
sc / LC / 6 |
||||
|
Hirundinidae |
|||||||||||
|
Tachycineta thalassina |
X |
X |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 8 |
|||
|
Stelgidopteryx serripennis |
X |
X |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 9 |
|||
|
Petrochelidon pyrrhonota |
X |
X |
X |
X |
X |
Mv |
INS / C. P. INV |
sc / LC / 7 |
|||
|
Hirundo rustica |
X |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 8 |
||||
|
Paridae |
|||||||||||
|
Baeolophus wollweberi |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 12 |
||||||
|
Trogloditydae |
|||||||||||
|
Salpinctes obsoletus |
X |
Rp |
INS / C. P. INV |
sc / LC / 12 |
|||||||
|
Catherpes mexicanus |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 11 |
|||||
|
Troglodytes brunneicollis |
X |
Rp |
CUA |
INS / C. P. INV |
sc / LC / 5 |
||||||
|
Campylorhynchus gularis |
X |
Rp |
END |
INS / C. P. INV |
sc / LC / 14 |
||||||
|
Campylorhynchus jocosus |
X |
Rp |
END |
INS / C. P. INV |
sc / LC / 16 |
||||||
|
Pheugopedius felix |
X |
X |
X |
Rp |
END |
INS / C. P. INV |
sc / LC / 14 |
||||
|
Thryophilus pleurostictus |
X |
X |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 15 |
|||
|
Polioptilidae |
|||||||||||
|
Polioptila caerulea |
X |
X |
X |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 7 |
||
|
Polioptila albiloris |
X |
X |
X |
X |
X |
Mloc |
INS / C. P. INV |
sc / LC / 15 |
|||
|
Regulidae |
|||||||||||
|
Regulus calendula |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 6 |
||||||
|
Turdidae |
|||||||||||
|
Catharus aurantiirostris |
X |
X |
Mv |
INS / C. P. INV |
sc / LC / 12 |
||||||
|
Catharus frantzii |
X |
Rp |
INS / C. P. INV |
A / LC / 16 |
|||||||
|
Catharus ustulatus |
X |
X |
T |
INS / C. P. INV |
sc / LC / 10 |
||||||
|
Catharus guttatus |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 6 |
||||||
|
Turdus rufopalliatus |
X |
X |
X |
X |
X |
Rp |
END |
INS / C. P. INV |
sc / LC / 10 |
||
|
Mimidae |
|||||||||||
|
Melanotis caerulescens |
X |
X |
X |
Rp |
END |
INS / C. P. INV |
sc / LC / 12 |
||||
|
Toxostoma curvirostre |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 9 |
||||||
|
Mimus polyglottos |
X |
Rp |
INS / C. P. INV |
sc / LC / 8 |
|||||||
|
Bombycillidae |
|||||||||||
|
Bombycilla cedrorum |
X |
Mi |
FRU / DIS. SEM. |
sc / LC / 6 |
|||||||
|
Ptilogonatidae |
|||||||||||
|
Ptiliogonys cinereus |
X |
Mloc |
CUA |
INS / C. P. INV |
sc / LC / 12 |
||||||
|
Passeridae |
|||||||||||
|
Passer domesticus exótica |
X |
X |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 8 |
|||||
|
Fringillidae |
|||||||||||
|
Euphonia elegantissima |
X |
X |
Rp |
FRU / DIS. SEM. |
sc / LC / 13 |
||||||
|
Haemorhous mexicanus |
X |
X |
X |
X |
Rp |
END |
GRA / C. POB. PLA. |
sc / LC / 6 |
|||
|
Spinus psaltria |
X |
X |
X |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 8 |
||||
|
Passerellidae |
|||||||||||
|
Aimophila rufescens |
X |
X |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 10 |
|||||
|
Aimophila ruficeps |
X |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 11 |
||||||
|
Melozone kieneri |
X |
X |
Rp |
END |
GRA / C. POB. PLA. |
sc / LC / 13 |
|||||
|
Peucaea acuminata |
X |
X |
X |
X |
Rp |
END |
GRA / C. POB. PLA. |
sc / LC / 11 |
|||
|
Peucaea humeralis |
X |
X |
X |
X |
X |
Rp |
END |
GRA / C. POB. PLA. |
sc / LC / 15 |
||
|
Spizella passerina |
X |
Mv |
GRA / C. POB. PLA. |
sc / LC / 8 |
|||||||
|
Spizella pallida |
X |
Mi |
CUA |
GRA / C. POB. PLA. |
sc / LC / 10 |
||||||
|
Pooecetes gramineus |
X |
X |
Mi |
GRA / C. POB. PLA. |
sc / LC / 11 |
||||||
|
Chondestes grammacus |
X |
X |
X |
Mi |
GRA / C. POB. PLA. |
sc / LC / 10 |
|||||
|
Melospiza lincolnii |
X |
X |
Mi |
GRA / C. POB. PLA. |
sc / LC / 7 |
||||||
|
Icteriidae |
|||||||||||
|
Icteria virens |
X |
X |
T |
INS / C. P. INV |
sc / LC / 9 |
||||||
|
Icteridae |
|||||||||||
|
Cassiculus melanicterus |
X |
Rp |
CUA |
OMN / INT. MULT |
sc / LC / 12 |
||||||
|
Icterus wagleri |
X |
X |
X |
X |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 12 |
||
|
Icterus spurius |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 9 |
|||||
|
Icterus cucullatus |
X |
Mi |
CUA |
INS / C. P. INV |
sc / LC / 10 |
||||||
|
Icterus pustulatus |
X |
X |
X |
X |
X |
Rp |
END |
INS / C. P. INV |
sc / LC / 10 |
||
|
Icterus bullockii |
X |
X |
X |
Mi |
CUA |
INS / C. P. INV |
sc / LC / 11 |
||||
|
Molothrus aeneus |
X |
X |
X |
X |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 6 |
|||
|
Molothrus ater |
X |
X |
X |
X |
Mi |
GRA / C. POB. PLA. |
sc / LC / 7 |
||||
|
Quiscalus mexicanus |
X |
X |
X |
X |
Rp |
OMN / INT. MULT |
sc / LC / 8 |
||||
|
Parulidae |
|||||||||||
|
Parkesia motacilla |
X |
X |
X |
Mi |
INS / TRANS. NUTR. |
sc / LC / 12 |
|||||
|
Mniotilta varia |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 10 |
|||||
|
Oreothlypis celata |
X |
Mi |
INS / C. P. INV |
sc / LC / 9 |
|||||||
|
Oreothlypis luciae |
X |
Mi |
CUA |
INS / C. P. INV |
sc / LC / 13 |
||||||
|
Oreothlypis ruficapilla |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 9 |
|||||
|
Oreothlypis virginiae |
X |
X |
X |
Mi |
CUA |
INS / C. P. INV |
sc / LC / 14 |
||||
|
Geothlypis poliocephala |
X |
Rp |
INS / C. P. INV |
sc / LC / 11 |
|||||||
|
Geothlypis tolmiei |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 11 |
||||||
|
Setophaga aestiva |
X |
Mv |
INS / C. P. INV |
sc / LC / 6 |
|||||||
|
Setophaga auduboni |
X |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 6 |
||||
|
Setophaga nigrescens |
X |
X |
Mi |
CUA |
INS / C. P. INV |
sc / LC / 11 |
|||||
|
Setophaga townsendi |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 10 |
||||||
|
Setophaga virens |
X |
Mi |
INS / C. P. INV |
sc / LC / 9 |
|||||||
|
Basileuterus rufifrons |
X |
X |
X |
Mloc |
END |
INS / C. P. INV |
sc / LC / 12 |
||||
|
Cardellina pusilla |
X |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 10 |
||||
|
Myioborus pictus |
X |
X |
Rp |
INS / C. P. INV |
sc / LC / 13 |
||||||
|
Myioborus miniatus |
X |
Rp |
INS / C. P. INV |
sc / LC / 11 |
|||||||
|
Cardinalidae |
|||||||||||
|
Piranga hepatica |
X |
X |
Rp |
CUA |
INS / C. P. INV |
sc / LC / 8 |
|||||
|
Piranga rubra |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 9 |
|||||
|
Piranga ludoviciana |
X |
X |
X |
X |
X |
Mi |
INS / C. P. INV |
sc / LC / 8 |
|||
|
Cardinalis cardinalis |
X |
Rp |
FRU / C. POB. PLA. |
sc / LC / 5 |
|||||||
|
Pheucticus chrysopeplus |
X |
X |
X |
X |
Rp |
END |
FRU / C. POB. PLA. |
sc / LC / 14 |
|||
|
Pheucticus ludovicianus |
X |
Mi |
FRU / C. POB. PLA. |
sc / LC / 11 |
|||||||
|
Pheucticus melanocephalus |
X |
X |
X |
X |
Mloc |
CUA |
FRU / C. POB. PLA. |
sc / LC / 9 |
|||
|
Passerina caerulea |
X |
X |
X |
Rp |
GRA / C. POB. PLA. |
sc / LC / 8 |
|||||
|
Passerina amoena |
X |
X |
X |
X |
Mi |
CUA |
GRA / C. POB. PLA. |
sc / LC / 9 |
|||
|
Passerina cyanea |
X |
X |
X |
X |
Mi |
GRA / C. POB. PLA. |
sc / LC / 9 |
||||
|
Passerina leclancherii |
X |
X |
X |
X |
X |
X |
Rp |
END |
GRA / C. POB. PLA. |
sc / LC / 15 |
|
|
Passerina versicolor |
X |
X |
X |
X |
X |
Mloc |
GRA / C. POB. PLA. |
sc / LC / 13 |
|||
|
Passerina ciris |
X |
X |
X |
Mi |
GRA / C. POB. PLA. |
Pr / NT / 12 |
|||||
|
Spiza americana |
X |
Mi |
GRA / C. POB. PLA. |
sc / LC / 10 |
|||||||
|
Thraupidae |
|||||||||||
|
Thraupis episcopus |
X |
Acc |
FRU / DIS. SEM. |
sc / LC / 7 |
|||||||
|
Volatinia jacarina |
X |
X |
X |
Mloc |
GRA / C. POB. PLA. |
sc / LC / 4 |
|||||
|
Sporophila torqueola |
X |
X |
|
|
X |
Rp |
END |
GRA / C. POB. PLA. |
sc / LC / 6 |
Referencias
Almazán-Núñez, R. C., Eguiarte, L. E., Arizmendi, M. C. y Corcuera, P. (2016). Myiarchus flycatchers are the primary seed dispersers of Bursera longipes in a Mexican dry forest. PeerJ, 4, e2126.
Almazán-Núñez, R. C. y Navarro, A. G. (2006). Avifauna de la subcuenca del río San Juan, Guerrero, México. Revista Mexicana de Biodiversidad, 77, 103–114.
AOS (American Ornithological Society). (2017). Checklist of North and Middle America birds. Recuperado el 11 enero, 2018 de: http://checklist.aou.org/
Arbeláez-Cortés, E., Milá, B. y Navarro-Sigüenza, A. G. (2014). Multilocus analysis of intraspecific differenciation in three endemic bird species from northern Neotropical dry forest. Molecular Phylogenetics and Evolution, 70, 362–377.
Arbeláez-Cortés, E., Roldán-Piña, D. y Navarro-Sigüenza, A. G. (2014). Multilocus phylogeography and morphology give insigths in the recent evolution of a Mexican endemic songbird: Vireo hypochryseus. Journal of Avian Biology, 45, 253–263.
Arizmendi, M. C. (2002). Estableciendo prioridades para la conservación de las aves. En H. Gómez-de Silva y A. Oliveras-de Ita. (Eds.), Conservación de aves. Experiencias en México (pp. 133–149). México D.F.: CIPAMEX.
Arizmendi, M. C., Berlanga, H., Márquez-Valdelamar, L., Navarijo, L. y Ornelas, F. (1990). Avifauna de la región de Chamela, Jalisco. Cuadernos (4). México, D.F.: Instituto de Biología, UNAM.
Arizmendi, M. C. y Ornelas, J. F. (1990). Hummingbirds and their floral resources in a tropical dry forest in Mexico. Biotropica, 22, 172–180.
Arizmendi, M. C. y Rodríguez-Flores, C. I. (2012). How many plant species do hummingbirds visit? Ornitología Neotropical, 23, 49–54.
Baselga, A. (2010). Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography, 19, 134–143.
Baselga, A. y Orme, C. D. L. (2012). betapart: an R package for the study of beta diversity. Methods in Ecology and Evolution, 3, 808–812.
Becerra, J. X. (2005). Timing the origin and expansion of the Mexican tropical dry forest. Proceedings of the National Academy of Sciences, 102, 10919–10923.
Berlanga, H., Kennedy, J. A., Rich, T. D., Arizmendi, M. C., Beardmore, C. J., Blancher, P. J. et al. (2010). Conservando a nuestras aves compartidas: la vision trinacional de Compañeros en Vuelo para la conservación de las aves terrestres. Ithaca, N.Y.: Cornell Lab of Ornithology.
Berlanga, H., Rodríguez-Contreras, V., Oliveras-de Ita, A., Escobar, M., Rodríguez, L., Vieyra, J. et al. (2008). Red de conocimientos sobre las aves de México (AVESMX). Ciudad de México: Conabio.
Bertelli, S., Szumik, C., Goloboff, P. A., Giannini, N. P., Navarro-Sigüenza, A. G., Peterson, A. T. et al. (2017). Mexican land birds reveal complexity in fine-scale patterns of endemism. Journal of Biogeography, 44, 1836–1846.
Bezaury, J. B. (2010). Las selvas secas del Pacífico mexicano en el contexto mundial. En G. Ceballos, L. Martínez, A. García, E. Espinoza, J. Bezaury-Creel y R. Dirzo (Eds.), Diversidad, amenazas y áreas prioritarias para la conservación de las selvas secas del Pacífico de México. (pp. 21–40). México D.F.: Fondo de Cultura Económica, Conabio.
Bibby, C. J., Burgess, N. D., Hill, D. A. y Mustoe, S. (2000). Bird census techniques. London: Academic Press.
BirdLife International. (2017). Important bird and biodiversity area factsheet: Papalutla, Sierra de Tecaballo. Recuperado el 19 de abril, 2017 de: http://datazone.birdlife.org/site/factsheet/papalutla-sierra-de-tecaballo-iba-mexico
Birdlife International (2018). Species factsheet: Ara militaris. Recuperado el 11 de enero, 2018 de: http://datazone.birdlife.org/species/factsheet/Military-Macaw
Blancas-Calva, E., Blancas-Hernández, J. A., Castro-Torreblanca, M. y Cano-Nava, G. (2017). La tángara azul gris (Thraupis episcopus) en el estado de Guerrero, México. Huitzil, 18, 232–237.
Blancas-Calva, E., Castro-Torreblanca, M. y Blancas-Hernández, J. C. (2014). Presencia de las palomas turca (Streptopelia decaocto) y africana de collar (Streptopelia roseogrisea) en el estado de Guerrero, México. Huitzil, 15, 10–16.
Boyle, W. A. (2017). Altitudinal migration in North America. The Auk, 134, 443–465.
Cardinale, B. J., Duffy, J, E., González, A., Hooper, D. U., Perrings, C., Venail, P. et al. (2012). Biodiversity loss and its impact on humanity. Nature, 486, 59–67.
Castro-Torreblanca, M., Blancas-Calva, E., Rodríguez-Mirón, G. M. y Espinosa-Organista, D. N. (2014). Patrones espaciales de distribución y diversidad de la avifauna en la provincia del Balsas. Revista Mexicana de Biodiversidad, 85, 823–830.
Chazdon, R. L., Colwell, R. K., Denslow, J. S. y Guariguata, M. R. (1998). Statistical methods for estimating species richness of woody regeneration in primary and secondary rain forests of NE Costa Rica. En F. Dallmeier y J. A. Comiskey (Eds.), Forest biodiversity research, monitoring and modeling: conceptual background and Old World case studies (pp. 285–309) París: Parthenon Publishing.
Chazdon, R. L., Harvey, C. A., Komar, O., Griffith, D. M., Ferguson, B. G., Martínez-Ramos, M. et al. (2009). Beyond reserves: a research agenda for conserving biodiversity in human-modified tropical landscapes. Biotropica, 41, 142–153.
Cockle, K. L., Martin, K. y Wesolowski, T. (2011). Woodpeckers, decay, and the future of cavity-nesting vertebrate communities worldwide. Frontiers in Ecology and the Environment, 9, 377–382.
Colwell, R. K. (2013). EstimateS: statistical estimation of species richness and shared species from samples. Version 9.1. Disponible en: http://purl.oclc.org/estimates
Conabio (Comisión Nacional para el conocimiento y uso de la Biodiversidad). (2017). Especies prioritarias. Programa de conservación de especies en riesgo. Recuperado el 19 abril, 2017 de http://www.biodiversidad.gob.mx/especies/espPrioritaria.html
Conafor (Comisión Nacional Forestal). (2014). Inventario estatal forestal y de suelos. Guerrero 2013. Zapopan, México D.F.: Conafor.
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2018). Áreas Naturales Protegidas Federales de México. Recuperado el 10 enero, 2018 de: http://sig.conanp.gob.mx/website/siganp/
Contreras-González, A. M. y Arizmendi, M. C. (2014). Pre-dispersal seed predation of the columnar cactus (Neobuxbaumia tetetzo, Cactaceae) by birds in central Mexico. Ornitología Neotropical, 25, 373–387.
del Hoyo, J., Elliott, A., Sargatal, J., Christie, D. A. y de Juana, E. (2017). Handbook of the birds of the world alive. Barcelona: Lynx Editions. Recuperado el 1 de marzo, 2017 de: http://www.hbw.com/
Díaz, S., Fargione, J., Chapin III, F. S. y Tilman, D. (2004). Biodiversity loss threatens human well-being. Plos Biology, 4, 1300–1305.
Dickman, C. R., Prevadec, M. y Lynam, A. J. (1991). Differential predation of size and sex classes of mice by the barn owl, Tyto alba. Oikos, 62, 67–76.
Dunn, J. L. y Alderfer, J. (2006). National Geographic field guide to the birds of North America, 6a Ed. Washington, D.C.: National Geographic.
eBird (2017). Recuperado el 10 abril, 2017 de: http://ebird.org/content/averaves
Enríquez, P. L. y Vázquez-Pérez, J. R. (2015). Los búhos de México. En P. L. Enríquez (Eds.), Los búhos neotropicales. Diversidad y conservación (pp. 475–512). México D.F.: El Colegio de la Frontera Sur.
Espinosa-Organista, D. N., Ocegueda-Cruz, S., Aguilar-Zúñiga, C., Flores-Villela, O., Llorente-Bousquets, J. y Vázquez-Benitez, B. (2008). El conocimiento biogeográfico de las especies y su regionalización natural. En R. Dirzo, R. González e I. March. (Eds.). Capital natural de México, vol. I: Conocimiento actual de la biodiversidad (pp. 33–65). Mexico D.F.: Conabio.
Faaborg, J. y Gauthreax, S. A. (2014). Weather conditions promoting the exceptional mid-October 2005 avian migration in the eastern Caribbean. The Journal of Caribbean Ornithology, 27, 9–14.
Feria-Arroyo, T. P. y Peterson, A. (2002). Prediction of bird community composition based on point-occurrence data and inferential algorithms: a valuable tool in biodiversity assessments. Diversity and Distributions, 8, 49–56.
Fernández-Nava, R., Rodríguez-Jiménez, C., Arreguín-Sánchez, M. L. y Rodríguez-Jimémez, A. (1998). Listado florístico de la cuenca del río Balsas, México. Polibotánica, 9, 1–151.
Fischer, J., Lindenmayer, D. B., Blomberg, S. P., Montague-Drake, R., Felton, A. y Stein, J. A. (2007). Functional richness and relative resilience of bird communities in regions with different land use intensities. Ecosystems, 10, 964–974.
González-García, F. y Gómez-de Silva, H. (2003). Especies endémicas: riqueza, patrones de distribución y retos para su conservación. En H. Gómez de Silva y A. Oliveras-de Ita. (Eds). Conservación de aves: experiencias en México. (pp. 150–194). Mexico D.F.: CIPAMEX/ Conabio/ NFWF.
Good-Eshelman, C. y Barrientos-López, G. (2004). Nahuas del Alto Balsas. Mexico D.F.: Comisión Nacional para el Desarrollo de los Pueblos Indígenas.
Holdridge, L. R. (1967). Life zone ecology. San José: Tropical Science Center.
Hortal, J. y Lobo, J. M. (2002). Una metodología para predecir la distribución espacial de la diversidad biológica. Ecología, 16, 405–432.
Howell, S. N. G. y Webb, S. (1995). A guide to the birds of Mexico and northern Central America. Oxford, RU: Oxford University Press.
Hulme, P. E. (2009). Trade, transport and trouble: managing invasive species pathways in an era of globalization. Journal of Applied Ecology, 46, 10–18.
Hutto, R. L. (1986). Migratory landbirds in western Mexico: a vanishing habitat. Western Wildlands, 11, 12–16.
Hutto, R. L. (1992). Habitat distributions of migratory landbird species in western Mexico. En J. M. Hagan III y D. W. Johnston (Eds). Ecology and conservation of Neotropical migrant landbirds. (pp. 221–239). Washington D.C.: Smithsonian Institution Press.
Hutto, R. L. (2010). Aspectos ecológicos únicos asociados con las aves migratorias de larga distancia del occidente de México. En G. Ceballos, L. Martínez, A. García, E. Espinoza, J. Bezaury-Creel y R. Dirzo. (Eds.). Diversidad, amenazas y áreas prioritarias para la conservación de las selvas secas del Pacífico de México. (pp. 215–234). México D.F.: Fondo de Cultura Económica/ Conabio.
Jiménez-Arcos, V. H., Santa Cruz-Padilla, S. A., Escalona-López, A., Arizmendi, M. C. y Vázquez, L. (2012). Ampliación de la distribución y presencia de una colonia reproductiva de la guacamaya verde (Ara militaris) en el alto Balsas de Guerrero, México. Revista Mexicana de Biodiversidad, 83, 864–867.
Jiménez-Valverde, A. y Hortal, J. (2003). Las curvas de acumulación de especies y la necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de Aracnología, 8, 151–161.
Koleff, P., Gaston, K. J. y Lennon, J. J. (2003). Measuring beta diversity for presence-absence data. Journal of Animal Ecology, 72, 367–382.
Koleff, P., Soberón, J., Arita, H. T., Dávila, P., Flores-Villela, O., Golubov, J. et al. (2008). Patrones de diversidad espacial en grupos selectos de especies. En R. Dirzo, R. González e I. March (Eds.), Capital natural de México, Vol. I: conocimiento actual de la biodiversidad (pp. 323–364). México D.F.: Conabio.
Lara, C. (2006). Temporal dynamics of flower use by hummingbirds in a highland temperate forest in Mexico. Ecoscience, 13, 23–29.
LGEEPA (Ley General del Equilibrio Ecológico y la Protección al Ambiente). (2017). Diario Oficial de la Federación última reforma publicada el 24 de enero de 2017. México.
Linares-Palomino, R., Oliveira-Filho, A. T. y Pennington, R. T. (2011). Neotropical seasonally dry forests: diversity, endemism, and biogeography of woody plants. En R. Dirzo, H. S. Young y G. Ceballos (Eds.). Seasonally dry tropical forests. Ecology and conservation (pp. 3–22). Washington D.C.: Island Press.
List, R., Rodríguez, P., Pelz-Serrano, K., Benítez-Malvido, J. y Lobato, J. M. (2017). La conservación en México: exploración de logros, retos y perspectivas desde la ecología terrestre. Revista Mexicana de Biodiversidad, 88, 65–75.
Lovette, I. J. (2005). Glacial cycles and tempo in avian speciation. Trends in Ecology and Evolution, 20, 57–59.
Luck, G. W., Lavorel, S., Mcintyre, S. y Lumb, K. (2012). Improving the application of vertebrate trait-based frameworks to the study of ecosystem services. Journal of Animal Ecology, 81, 1065–1076.
Maass, M., Búrquez, A., Trejo, I., Valenzuela, D., González, M. A., Rodríguez, M. et al. (2010). Amenazas. En G. Ceballos, L. Martínez, A. García, E. Espinoza, J. Bezaury-Creel y R. Dirzo. (Eds.). Diversidad, amenazas y áreas prioritarias para la conservación de las selvas secas del Pacífico de México. (pp. 321–348). México D.F.: Fondo de Cultura Económica/ Conabio.
Martínez-Gordillo, M., Valencia-Avalos, S. y Calónico-Soto, J. (1997). Flora Papalutla, Guerrero y de sus alrededores. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Botánica, 68, 107–133.
Mason, N. A., Olvera-Vital, A., Lovette, I. J. y Navarro-Sigüenza, A. G. (2018). Hidden endemism, deep polyphyly, and repeated dispersal across the Itsmus of Tehuantepec: diversification of the White-collared Seedeater complex (Thraupidae: Sporophila torqueola). Ecology and Evolution, 8, 1867–1881.
McGill, B. J., Dornelas, M., Gotelli, N. J. y Magurran, A. E. (2015). Fifteen forms of biodiversity trend in the Anthropocene. Trends in Ecology and Evolution, 30, 104–113.
McKinney, M. L. y Lockwood, J. L. (1999). Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends in Ecology and Evolution, 14, 450–453.
Meza, L. y López-García, J. (1997). Vegetación y mesoclima de Guerrero. Estudios florísticos en Guerrero. México, D.F.: Universidad Nacional Autónoma de México.
Montaño-Rendón, M., Sánchez-González, L. A., Hernández-Alonso, G. y Navarro-Sigüenza, A. G. (2015). Genetic differenciation in the Mexican endemic Rufous-backed Robin, Turdus rufopalliatus (Passeriformes: Turdidade). Zootaxa, 4034, 495–514.
Monterrubio-Rico, T. C. y Escalante-Pliego, P. (2006). Richness, distribution and conservation status of cavity nesting birds in Mexico. Biological Conservation, 128, 67–78.
Moreno, C. E. (2001). Métodos para medir biodiversidad. Zaragoza, España: M&T Manuales y Tesis SEA.
Morrone, J. J. (2014). Biogeographical regionalization of the Neotropical region. Zootaxa, 3782, 1–110.
NABCI (North American Bird Conservation Initiative). (2016). El estado de las aves de Norteamérica 2016. Environment and Climate Change Canada: Ottawa, Ontario. Recuperado el 18 de abril, de www.es.stateofthebirds.org
Naranjo, E. J. y Cuarón, A. D. (201)0. Uso de la fauna silvestre. En G. Ceballos, L. Martínez, A. García, E. Espinoza, J. Bezaury-Creel y R. Dirzo. (Eds.). Diversidad, amenazas y áreas prioritarias para la conservación de las selvas secas del Pacífico de México (pp. 271–284). México D.F.: Fondo de Cultura Económica/ Conabio.
Naturam Sequi (2015). Prospección de la guacamaya verde en el Alto Balsas. Reporte final. Programa de Conservación de Especies en Riesgo. México D.F.: Conanp, Semarnat.
Naturam Sequi (2016). Conservación del hábitat de la guacamaya verde en el Alto Balsas. Reporte final, Programa de Conservación de Especies en Riesgo. México D.F.: Conanp, Semarnat.
Navarro-Sigüenza, A. G. (2002). Atlas de las Aves de México: Fase II. Base de datos de SNIB-Conabio proyectos E018 y A002. México D.F.: Facultad de Ciencias, Universidad Nacional Autónoma de México.
Navarro-Sigüenza, A. G. y Peterson, T. (2004). An alternative species taxonomy of the birds of Mexico. Biota Neotropica, 4, 1–32.
Navarro-Sigüenza, A. G., Rebón-Gallardo, M. F., Gordillo-Martínez, A., Peterson, T., Berlanga-García, H. y Sánchez-González, L. (2014). Biodiversidad de aves en México. Revista Mexicana de Biodiversidad, 85, 476–495.
Navarro-Sigüenza, A. G., Vázquez-Miranda, H., Hernández-Alonso, G., García-Trejo, E. A. y Sánchez-González, L. A. (2017). Complex biogeographic scenarios revealed in the diversification of the largest woodpecker radiation in the New World. Molecular Phylogenetics and Evolution, 112, 53–67.
Nova-Muñoz, O., Almazán-Núñez, R. C., Bahena-Toribio, R., Cruz-Palacios, M. T. y Puebla-Olivares, F. (2011). Riqueza y abundancia de aves de la subcuenca de Tuxpan, Guerrero, México. Universidad y Ciencia, 27, 299–313.
Ochoa-Arteaga, A., García-Leyva, A. y Charre, G. M. (2016). Primer registro del gavilán pico gancho (Chondrohierax uncinatus) para la localidad de El Limón, sierra de Huautla, Morelos, México. Huitzil, 17, 215–219.
Ochoa-Ochoa, L., Munguía, M., Lira-Noriega, A., Sánchez-Cordero, V., Flores-Villela, O., Navarro-Sigüenza, A. et al. (2014). Spatial scale and ß-diversity of terrestrial vertebrates in Mexico. Revista Mexicana de Biodiversidad, 85, 918–930.
Ornelas, J. F. y Arizmendi, M. C. (1995). Altitudinal migration: implications for the conservation of the Neotropical migrant avifauna of western Mexico. (pp. 98–112). En M. H. Wilson y S. A. Sader (Eds.), Conservation of Neotropical migrant avifauna of western Mexico. Orono, USA.: Maine Agricultural and Forest Experiment Station.
Ortega-Álvarez, R., Sánchez-González, L. A. y Berlanga-García, H. (2015). Plumas de multitudes. Integración comunitaria en el estudio y monitoreo de aves en México. México D.F.: Conabio.
Pennington, R. T., Prado, D. E. y Pendry, C. A. (2000). Neotropical seasonally dry forests and Quaternary vegetation changes. Journal of Biogeography, 27, 261–273.
Peterson, A. T. y Navarro-Sigüenza, A. G. (2000). Western Mexico: a significant centre of avian endemism and challenge for conservation action. Cotinga, 14, 42–46.
Peterson, A. T. y Navarro-Sigüenza, A. G. (2016). Bird conservation and biodiversity research in Mexico: status and priorities. Journal of Field Ornithology, 87, 121–132.
Peterson, T. y Chalif, R. (1989). Guía de aves de México. México D.F.: Editorial Diana.
Pietrek, A. G., Walker, R. S. y Novaro, A. J. (2009). Susceptibility of lizards to predation under two levels of vegetative cover. Journal of Arid Environments, 73, 574–577.
Prieto-Torres, D. A., Navarro-Sigüenza, A. G., Santiago-Alarcón, D. y Rojas-Soto, O. R. (2016). Response of the endangered tropical dry forests to climate change and the role of Mexican Protected Areas for their conservation. Global Change Biology, 22, 365–379.
R Development Core Team. (2008). R: a language and environment for statistical computing, R Foundation for Statistical Computing, Austria. Disponible: http://www.R-project.org
Ralph, C. R., Geupel, G. R., Pyle, P., Martin, T. E., DeSante, D. F. y Milá, B. (1996). Manual de métodos de campo para el monitoreo de aves terrestres. Albany, CA: US Forest Service.
Ramírez-Albores, J. E. (2007). Bird diversity and conservation of Alto Balsas (Southwestern Puebla), Mexico. Revista de Biología Tropical, 55, 287–300.
Ramírez-Albores, J. E. (2010). Presencia del cardenal rojo (Cardinalis cardinalis) en el suroeste de Puebla, México. Huitzil, 11, 42–45.
Ramírez-Albores, J. E. y Ramírez-Cedillo, M. G. (2002). Avifauna de la región oriente de la sierra de Huautla, Morelos. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 73, 91–111.
Ramírez-Bastida, P., Navarro-Sigüenza, A. G. y Peterson, A. T. (2008). Aquatic bird distributions in Mexico: designing conservation approaches quantitatively. Biodiversity and Conservation, 17, 2525–2558.
Ramírez-Bastida, P., Vázquez, L. y Navarro-Sigüenza, A. G. (2015). Aves de los encinares mexicanos: riqueza específica, endemismo y relaciones faunísticas. En S. Romero-Rangel, E. Rojas-Zenteno y L. E. Rubio-Licona (Eds.), Encinos de México (Quercus, Fagaceae) 100 especies. (pp. 91–128). México D.F.: FES Iztacala, UNAM.
Ramos-Ordoñez, M. F. y Arizmendi, M. C. (2011). Parthenocarpy, attractiveness and seed predation by birds in Bursera morelensis. Journal of Arid Environments, 75, 757–762.
Ridgely, R. S., Allnutt, T. F., T. Brooks, McNicol, D. K., Mehlman, D. W., Young, B. E. et al. (2005). Digital distribution maps of the birds of the western hemisphere version 2.1. Arlington, Virginia, USA: NatureServe.
Ríos-Muñoz, C. A. (2013). ¿Es posible reconocer una unidad biótica entre América del Norte y del Sur? Revista Mexicana de Biodiversidad, 84, 1022-1030.
Rivera-Ortíz, F. A., Oyama, K., Ríos-Muñoz, C. A., Solórzano, S., Navarro-Sigüenza, A. G. et al. (2013). Habitat characterization and modeling of the potential distribution of the Military Macaw (Ara militaris) in México. Revista Mexicana de Biodiversidad, 84, 1200–1215.
Robbins., S. C., Bruun, B. y Zim, H. S. (1966). A guide to field identification birds of North America. Nueva York: Golden Press.
Rodríguez-Ruíz, E. R., Garza-Torres, H. A., Ríos-Muñoz, C. A. y Navarro-Sigüenza, A. G. (2011). La distribución geográfica de la tángara azul-gris (Thraupis episcopus) en hábitats modifcados antropogénicamente en México. Revista Mexicana de Biodiversidad, 82, 989–996.
Rojas-Soto, O. R., Oliveras-de Ita, A., Almazán-Núñez, R. C., Navarro-Sigüenza, A. G. y Sánchez-González, L. A. (2009). Avifauna de Campo Morado, Guerrero. Revista Mexicana de Biodiversidad, 80, 741–749.
Rzedowski, J. (2006). Vegetación de México. Edición digital. México. Conabio. Disponible en: http://bioteca.biodiversidad.gob.mx/janium/Documentos/7369.pdf
Rzedowski, J., Medina-Lemos, R. y Calderón-de Rzedowski, G. (2005). Inventario del conocimiento taxonómico, así como de la diversidad y del endemismo regionales de las especies mexicanas de Bursera (Burseraceae). Acta Botanica Mexicana, 70, 85–111.
Salgado-Negret, B. y Paz, H. (2015). Escalando de los rasgos funcionales a procesos poblacionales, comunitarios y ecosistémicos. En B. Salgado-Negret (Ed.), La ecología funcional como aproximación al estudio, manejo y conservación de la biodiversidad: protocolos y aplicaciones (pp. 12–35). Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Sánchez-Azofeifa, G. A. y Portillo-Quintero, C. (2011). Extent and drivers of change of neotropical seasonally dry tropical forests. En R. Dirzo, H. S. Young, H. A. Mooney y G. Ceballos (Eds.), Seasonally dry tropical forests: ecology and conservation (pp. 45–58). Washington D.C.: Island Press.
Sánchez-González, L. A. y Ortega-Álvarez, R. (2015). Monitoreo comunitario de aves: un elemento fundamental en los esfuerzos de conservación de la biodiversidad. En R. Ortega-Álvarez, L. A. Sánchez-González y H. Berlanga-García. (Eds.), Plumas de multitudes. Integración comunitaria en el estudio y monitoreo de aves en México. México D.F.: Conabio.
Şekercioğlu, C. H., Daily, G. C. y Ehrlich, P. R. (2004). Ecosystem consequences of bird declines. Proceedings of the National Academy of Sciences, 101, 18042–18047.
Simberloff, D., Martin, J. L., Genovesi, P., Maris, V., Wardle, D. A., Aronson, J. et al. (2013). Impacts of biological invasions: what’s what and the way forward. Trends in Ecology and Evolution, 28, 58–66.
Semaren (Secretaría de Medio Ambiente del Estado de Guerrero). (2018). Las Áreas Naturales Protegidas en el Estado de Guerrero. Recuperado el 15 enero, 2018: http://guerrero.gob.mx/articulos/537959-2/
Semarnat (Secretaría de Medio Ambiente y Recursos Naturales). (2010). Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental. Especies nativas de México de flora y fauna silvestres. Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio. Lista de especies en riesgo. Diario Oficial de la Federación 30 de diciembre 2010. México.
Sodhi, N. S., Şekercioğlu, C. H., Barlow, J. y Robinson, S. K. (2011). Ecological functions of birds in the Tropics. En N. S. Sodhi, C. H. Şekercioğlu, J. Barlow y S. K. Robinson. Conservation of Tropical Birds. (pp. 68–108). UK: Wiley-Blackwell.
Stattersfield, A. J., Crosby, M. J., Long, A. J. y Wege, D. C. (1999). Endemic Bird Areas of the World. Priorities for biodiversity conservation. Cambridge, UK: Birdlife International.
Stotz, D. F., Fitzpatrick, J. W., Parker III, T. A. y Moskovits, D. K. (1996). Neotropical birds. Ecology and conservation. Chicago: The University of Chicago Press.
Thiollay, J. M. (2017). Hawks, eagles (Accipitridae). En J. del Hoyo, A. Elliott, J. Sargatal, D. A. Christie y E. de Juana (Eds.). Handbook of the birds of the world alive. Barcelona. Lynx Edicions. Recuperado el 18 abril, 2017 de: http://www.hbw.com/node/52213
Trejo, I. y Dirzo, R. (2002). Floristic diversity of Mexican seasonally dry tropical forests. Biodiversity and Conservation, 11, 2063–2084.
UNESCO (United Nations Educational, Scientific and Cultural Organization). (2017). Reservas de la biosfera. Recuperado el 19 abril, 2017 de: http://www.unesco.org/new/es/natural-sciences/environment/ecological-sciences/biosphere-reserves/
Urbina-Torres, F. (2016). Registros notables de aves de Morelos, México. Huitzil, 17, 163–174.
Valencia, S. (2004). Diversidad del género Quercus (Fagaceae) en México. Boletín de la Sociedad Botánica de México, 75, 33–53.
Vázquez-Reyes, L. D., Arizmendi, M. C., Godínez-Álvarez, H. O. y Navarro-Sigüenza, A. G. (2017). Directional effects of biotic homogenization of bird communities in Mexican seasonal forests. The Condor, 119, 275–288.
Vázquez-Reyes, L. y Renton, K. (2015). High density of tree-cavities and snags in tropical dry forest of western Mexico raises questions for a latitudinal gradient. Plos One, 10, e0116745.
Vega-Rivera, J. H., Arizmendi, M. C. y Morales-Pérez, L. (2010). Aves. En G. Ceballos, L. Martínez, A. García, E. Espinoza, J. Bezaury-Creel y R. Dirzo. (Eds.). Diversidad, amenazas y áreas prioritarias para la conservación de las selvas secas del Pacífico de México. (pp. 145–164). México D.F.: Fondo de Cultura Económica/ Conabio.
Villaseñor-Gómez, J. F., Villaseñor-Gómez, L. E. y Chávez, G. (2000). AICA 23 “Cuenca Baja del Balsas”. En M. C. Arizmendi y L. Márquez. (Eds.), Áreas de importancia para la conservación de las aves en México. México D.F.: CIPAMEX.
Wenny, D. G., Devault, T. L., Johnson, M. D., Kelly, D., Şekercioğlu, C. H., Tomback, D. F. et al. (2011). The need to quantify ecosystem services provided by birds. The Auk, 128, 1–14.
Whittaker, R. H. (1972). Evolution and measurement of species diversity. Taxon, 21, 213–251.


