Pastor Coayla-Peñaloza a, André Alexander Cheneaux-Díaz a, Claudia Viviana Moreno-Salazar a, Cynthia Elizabeth Cruz-Remache a, Eusebio Walter Colque-Rondón a, Cristina Damborenea b, *
a Universidad Nacional de San Agustín, Departamento Académico de Biología, Laboratorio de Hidrobiología, Av. Alcides Carrión s/n, Arequipa, Peru
b Consejo Nacional de Investigaciones Científicas y Técnicas, División de Zoología de Invertebrados-Universidad Nacional de La Plata, FCNyM, Paseo del Bosque s/n -B1900FWA, La Plata, Argentina
*Corresponding author: cdambor@fcnym.unlp.edu.ar (C. Damborenea)
Received: 10 October 2021; accepted: 7 September 2022
Abstract
High Andean wetlands are fragile systems, vulnerable to human activity and climate change. In the Arequipa region (Peru), there are high Andean lotic and lentic systems currently affected by livestock raising, fish farming, and dams. The aim of the study was to evaluate the aquatic invertebrate community in the Callalli-Oscollo wetlands and the possible impact of human activities. Samples were taken from November 2017 to October 2018 at 4 sampling stations in lotic environments and 2 in lentic environments. Macroinvertebrates were identified to the family level. The following were determined to evaluate community structure: richness, relative abundance, Shannon-Wiener diversity, Simpson dominance, Pielou evenness, and true diversity. The indices ABI, BMWP/Bol and nPeBMWP were applied to evaluate the ecological quality of the environments sampled. Thirty families were recorded in lotic environments, the most abundant being Chironomidae, Naididae, Limnesiidae, Elmidae, Baetidae and Lumbriculidae. The ecological quality was good, except at the station associated to the dam, where it was doubtful. Twenty-six families were recorded in lentic environments, the most abundant being Cyprididae, Naididae and Corixidae. The macroinvertebrate communities in high Andean environments reflect ecosystem conditions. Environments associated to human activity have lower ecological quality.
Keywords: Bioindicators; Aquatic macroinvertebrates
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Comunidades de macroinvertebrados bentónicos y evaluación de la calidad del agua en humedales altoandinos de Callali-Oscollo, Arequipa-Cusco, Perú
Resumen
Los humedales altoandinos son sistemas frágiles, vulnerables a la actividad humana y al cambio climático. La región de Arequipa (Perú) posee ecosistemas lóticos y lénticos altoandinos con impactos por la ganadería, piscicultura y represamiento. Se evaluó la comunidad de macroinvertebrados acuáticos de los humedales de Callalli-Oscollo y el posible efecto de las actividades humanas. Desde noviembre de 2017 a octubre de 2018 se realizaron muestreos en 4 estaciones en ambientes lóticos y 2 en lénticos. Los macroinvertebrados se identificaron a nivel de familia. Para evaluar la estructura comunitaria se registró la riqueza, la abundancia relativa, la diversidad de Shannon-Wiener, la dominancia de Simpson, la equidad de Pielou y la diversidad verdadera. Se aplicaron los índices ABI, BMWP/Bol y nPeBMWP para evaluar la calidad ecológica de los ambientes lóticos. En éstos se registraron 30 familias, siendo las más abundantes Chironomidae, Naididae, Limnesiidae, Elmidae, Baetidae y Lumbriculidae. La calidad ecológica fue buena, con excepción de la estación vinculada a la presa que fue dudosa. Los ambientes lénticos registraron 26 familias, siendo las más abundantes Cyprididae, Naididae y Corixidae. Las comunidades de macroinvertebrados en ambientes altoandinos reflejan el estado del ecosistema. Los ambientes vinculados con la actividad humana registran menor calidad ecológica.
Palabras clave: Bioindicadores; Macroinvertebrados acuáticos
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
Aquatic systems, as ecosystem services, are subject to strong environmental pressures. Land use, use of water for human activities and water pollution, affect aquatic environments, both lotic and lentic, altering their physicochemical properties and the structure and composition of the communities living in them. These ecosystems harbor a series of communities that interact with the physicochemical characteristics of the water, the atmosphere and the surrounding terrestrial environment, thereby determining the global behavior of the ecosystem. Any alteration affecting the ecosystems also impacts the communities inhabiting them (Oyague, 2005; Posada et al., 2000; Roldán, 2012). High-altitude wetlands, such as those in the high Andes, are exceptional ecosystems that are sensitive to the effects of global warming (Báez et al., 2016), the impact of agrochemicals, livestock raising activities, mining, extraction of water for different purposes, and dams (Jacobsen, 2008). Nonetheless, and given the operational costs, they are among the least studied ecosystems (Luque-Fernández et al., 2020; Nieto et al., 2016).
Benthic macroinvertebrates are highly sensitive to environmental changes, so they have traditionally been used to monitor water quality in rivers and streams (Buss et al., 2015; Ríos-Touma et al., 2014). They react to variations in water quality and respond to environmental stress with different degrees of tolerance, reflecting any changes in space or time, thereby providing an efficient, rapid instrument for evaluating water quality and ecosystem conditions (González-Zuarth et al., 2014).
Aquatic macroinvertebrates have been increasingly used as water quality indicators in Peru (Acosta, 2001; Paredes et al., 2004; among others). However, there are few studies in high Andean environments, particularly in the Arequipa region (Luque-Fernández et al., 2020; Tapia et al., 2018).
Considering the above, this study proposes to assess the water quality in lotic environments located in the Callalli-Oscollo districts using the study of aquatic macroinvertebrate community. The diversity of aquatic macroinvertebrate families was determined in lotic and lentic systems and the possible impact of human activities on the ecological quality of water was determined. This study advances the knowledge of aquatic macroinvertebrates in high-altitude ecosystems in general and high Andean ecosystems in particular.
Materials and methods
The study was conducted in the high Andean aquatic environments located near the Negrillo River, which is a tributary of the Colca River and flows through the districts of Callalli and Oscollo, delimiting the departments of Arequipa and Cusco, Peru (Fig. 1). The sampling stations were located between 4,524 and 4,580 m asl. The mean monthly temperature in the study zone ranges from 6 °C in the rainy months (late December to March) to 1 °C in low water level months, when cloud cover is lower. The average annual precipitation is 710 mm (Autoridad Nacional del Agua, 2014).
Different lotic environments (creeks) and lentic environments (small ponds) were identified. The lotic environments have cold, crystalline waters supplied by snowmelt, their beds are dominated by rocky substrates, mainly pebbles; however, the presence of organic matter is observed, mainly on the riverbanks. The lentic environments are characterized by substrates composed mainly of clay, silt, and organic matter, the latter mainly coming from fish farms in the area.
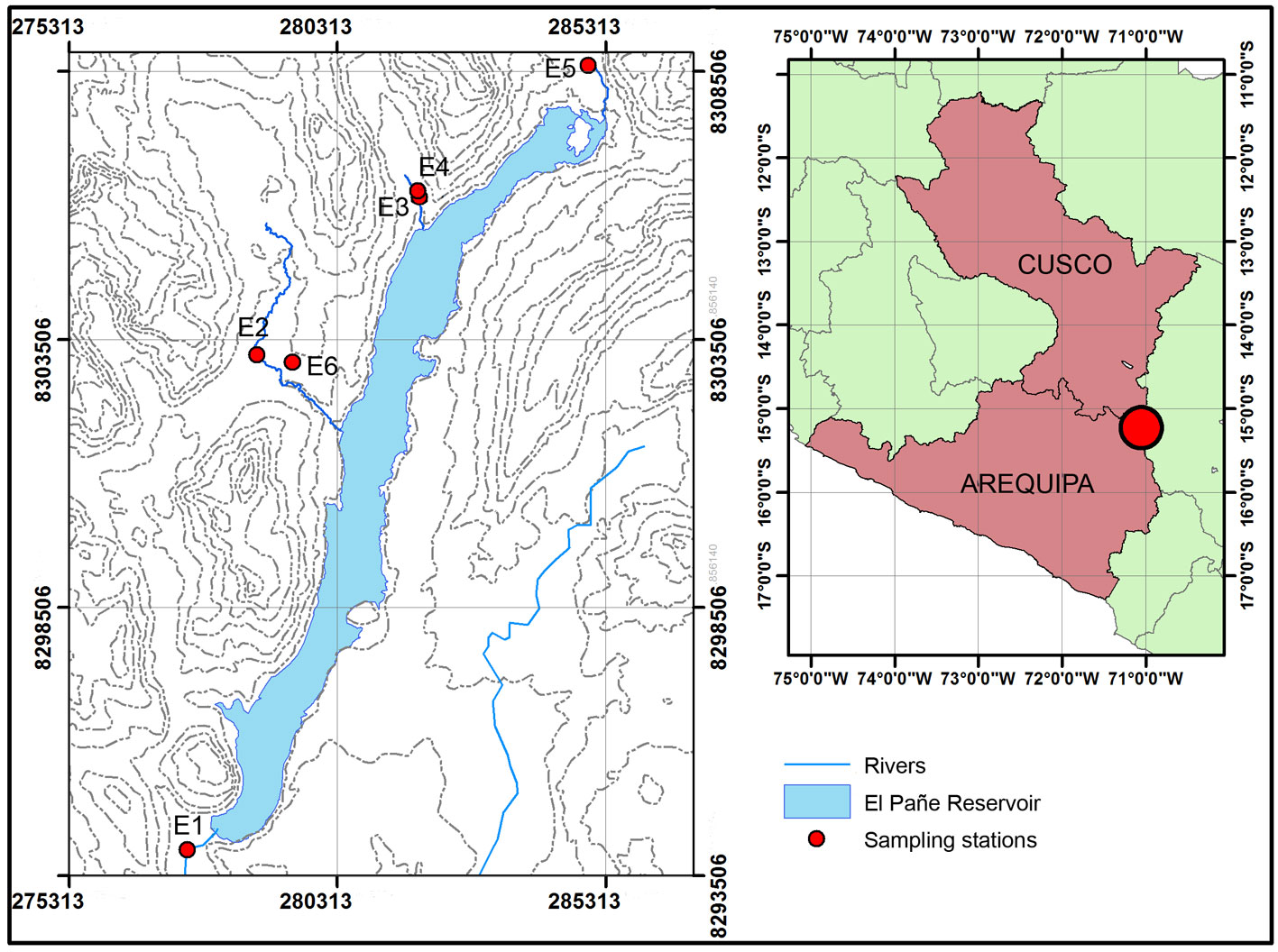
Table 1
Sampling stations in lotic and lentic environments in the districts of Callalli and Oscollo. Arequipa-Cusco, Peru.
| Stations | Characteristics | Latitude S | Longitude W |
| E1 | Creek | 15°25’17.20” | 71°04’23.40” |
| E2 | Creek | 15°20’17.60” | 71°03’37.00” |
| E3 | Creek | 15°18’46.00” | 71°01’54.70” |
| E4 | Creek | 15°18’39.13” | 71°01’55.70” |
| E5 | Pond | 15°17’23.81” | 71°00’08.53” |
| E6 | Pond | 15°20’22.10” | 71°03’15.18” |
The study is based on 6 sampling efforts conducted in November and December 2017 and April, May, July and October 2018, at 4 sampling stations in lotic environments (E1, E2, E3, and E4) and 2 in lentic environments (E5 and E6) (Fig. 1, Table 1). Station E1 is located in a stream that receives the flow from El Pañe reservoir and is affected by its dynamics; it has a rocky substrate and little vegetation; the width of the stream is not more than 7 m and the length covers 1.4 km. Station E2 has a rocky substrate composed mainly of pebbles; the width of the stream is not more than 6 m while the length is close to 0.9 km. Station E3 has a rocky substrate composed mainly of gravel and pebbles, and it also has vegetation on the banks; the width of the stream is not greater than 3 m and its length is close to 0.5 km. Station E3 has a higher development of anthropic activities, such as grazing, use of water resources by inhabitants and it is circulating near a road. Finally, in station E4, the substrate is rocky and the banks have vegetation. The width of the stream in station 4 does not exceed 3 m, while its length covers approximately 0.4 km. This station is not impacted by human activity and is the most distant point from Station 3 with access. Upstream from station 4, the flow is reduced and sampling is not possible. Station E5 corresponds to a lagoon located in the highest part of the sampling area, it is affected by the artisan farming of the introduced fish Onchorhynchus mykiss (Walbaum, 1792); the substrate is composed mainly of silt and organic matter mainly from fish farming; the depth is less than 0.8 m, while the area corresponds to approximately 1.23 km2. Station E6 corresponds to a lagoon whose substrate is mainly composed of silt and clay, the depth is not greater than 0.9 m and its area is close to 0.7 km2. As a result of the environmental characteristics and seasonal variations in the flow from the reservoir, it was not possible to sample station E1 in May and July, and station E6 in July and October.
Dissolved oxygen (DO), pH, electrical conductivity (EC) and temperature (T) were measured in the field using a Hanna HI9829 multiparameter device. Each parameter was measured 3 times per sample station on each sampling date. ANOVA was used to compare physicochemical variables according to sampling station and date.
Specimens were collected with a Surber net (500 µm mesh opening and 30 cm side frame) in lotic environments and a D-frame net (500 µm mesh, ratio 30 cm with semicircular opening) in lentic zones. For the quantitative measurement of macroinvertebrates at each station, we took 3 samples located at a distance of 100 m (3 samples × 6 stations × 6 months, 96 samples altogether).
The specimens from each sample were stored separately in labelled jars and fixed in 5% formalin. In the laboratory, the samples were washed and preserved in 70% ethanol for subsequent identification under stereomicroscope and optical microscope (Domínguez & Fernández, 2009; Roldán, 2003). Specimens were identified to family level using the keys by Domínguez and Fernández (2009), Merritt et al. (2008), Prat et al. (2011), and Thorp and Lovell (2015).
The following data were determined for each sampling station in order to analyze community structure: richness (number of families), relative abundance (number of individuals of 1 family/total number of individuals of all families)* 100, Shannon-Wiener diversity (H´) in Napierian logarithm, Simpson dominance (D), Pielou evenness (J´) and true diversity (D1) (Moreno, 2001). Based on the macroinvertebrate abundance matrix, the Bray-Curtis similarity index was used with the UPGMA algorithm (unweighted pair-group method) expressed in a cluster analysis to determine the faunal associations among temporal/spatial samples. SIMPER (similarity percentages) was used to evaluate the contribution of the species to the similarity and ANOSIM similarity analysis to determine significant differences in macrobenthos between samples. Analyses were performed using the software EXCEL® and PAST version 4.03 software (Hammer et al., 2001). The ecological quality of the lotic environments was determined by means of biotic indices such as the Andean Biological Index or ABI (Acosta et al., 2009), Biological Monitoring Working Party as BMWP/Bol (Rocabado & Goitia, 2011) and nPeBMWP (Medina-Tafur et al., 2010) considering the tolerance or sensitivity values of each family to the disturbances to the environment and the Andean region. These indices have demonstrated precision to evaluate the disturbance related to pollution from urbane source, such as the one studied here (Molina et al., 2022). To establish the tolerance values of the families, we followed Acosta et al. (2009), Rocabado and Goitia (2011) and Medina-Tafur et al. (2010).
Results
The surface temperature of the water in the lotic systems ranged from 10.88 °C at station E4 to 12.13 °C at station E2; while in ponds, it ranged from 10.47 °C to 13.16°C. Values for pH ranged from 8.18 (station E1) to 8.55 (station E4) in creeks, and 7.83 to 8.48 in ponds. The mean electrical conductivity in lotic environments ranged from 35.44 μS at station E4 to 122.03 μS at station E1. The highest values were recorded in December-2017 (E1 = 371 μS; E3 = 340 μS). In lentic environments, the mean was 33.00 μS (E6) and 140.83 μS (E5), being the higher values from December, 2017. Mean dissolved oxygen concentration ranged from 3.08 mg/l-¹ at station E3 to 3.59 mg/l-¹ at station E1; while in ponds, it ranged from 3.12 mg/l-¹ to 3.60 mg/l-¹ (Table 2). Two-way ANOVA revealed that there was statistically significant interaction between the effects of sample stations and date in the physicochemical variables considered (Table 3).
Macroinvertebrates in lotic environments
A total of 19,420 individuals were counted, distributed among 30 families. At station E1, richness was highest in December (9 families) and lowest in November (5 families). The families with greatest relative abundance at E1 were Chironomidae (56.35%), Hyalellidae (25.28%), Lumbriculidae (9.44%) and Naididae (3.45%) (Table 4). The highest abundance was recorded in July, and the lowest in April.
At station E2, richness ranged from 15 (in May) to 10 (November and April). The families with highest average relative abundance were Chironomidae (50.57%), Limnesidae (15.86%) and Naididae (14.62%) (Table 4). For these taxa, abundance was highest in May and lowest in April.
At station E3, richness ranged from 18 (November) to 8 (October). The families with the highest relative abundance at E3 were: Naididae (34.61%), Chironomidae (21.15%), Elmidae (9.99%) and Baetidae (7.51%) (Table 4). Abundance was highest in November and lowest in December.
At station E4, richness was highest (17) in May and lowest in November and April (12). The families with the highest relative abundance were Naididae (30.57%), Chironomidae (23.53%) and Lumbriculidae (23.17%) (Table 4). Abundance was highest in July and lowest in April.
Shannon’s index ranged from 0.37 (November, station E1) to 1.95 (May, station E4). True diversity was lowest at stations E1 and E2, with values close to 2 and 4 effective families, respectively. Stations E3 and E4 had the highest true diversity, with values close to 6 and 7 effective species, respectively. Dominance was highest at station E1 (0.84 in November), and lower at the other 3 stations. Evenness index was lowest (0.23) at E1 in November 2017 and highest (0.77) at E3 in October 2018 (Fig. 2).
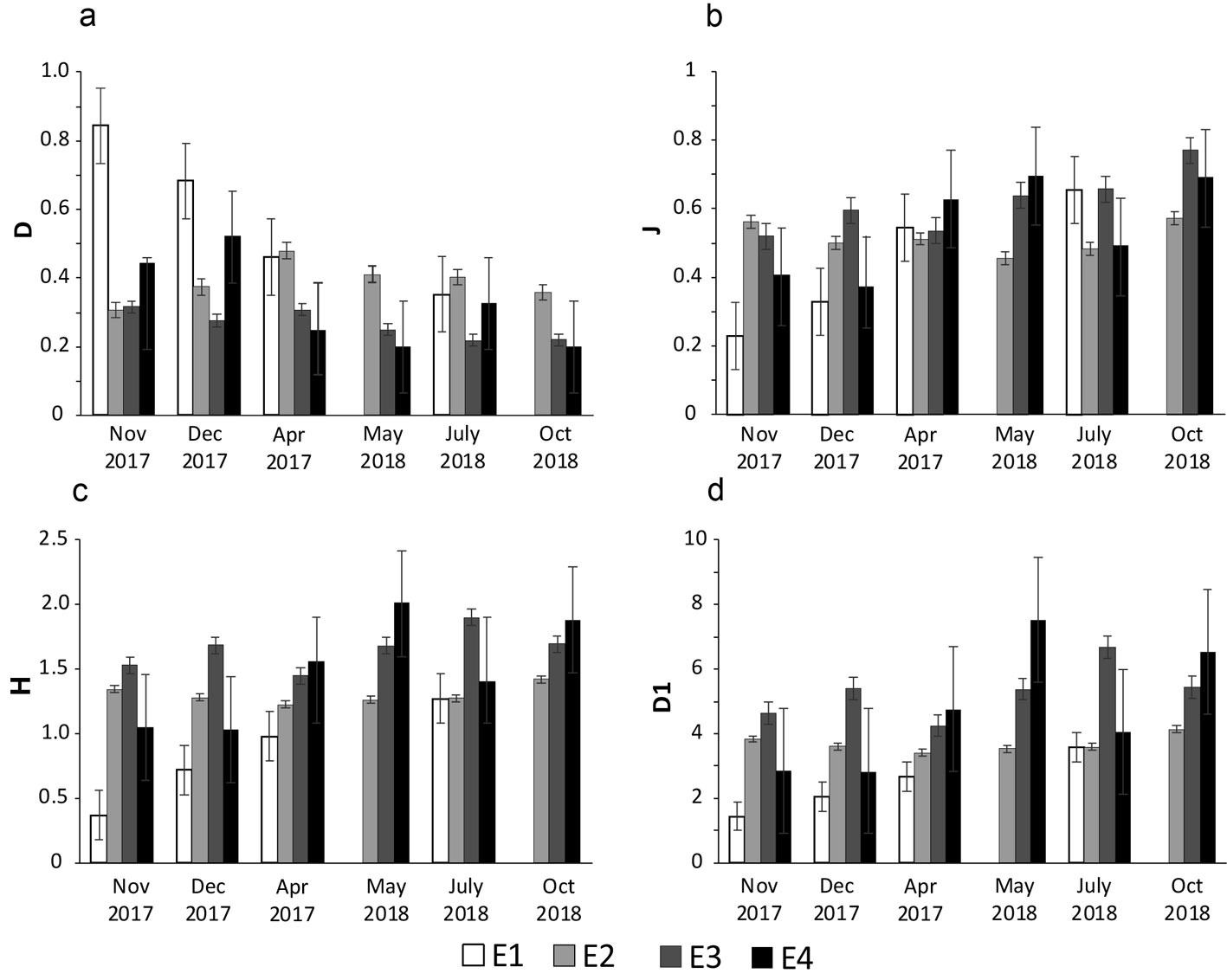
Table 2
Average values for physicochemical parameters (T, temperature; pH; EC, electrical conductivity; DO, dissolved oxygen) and standard deviation in lotic and lentic environments in Callalli-Oscollo. Arequipa-Cusco, Peru.
| Station | Type | n | T (°C) | pH | EC (µS/cm) | DO (mg/L) |
| E1 | Creek | 15 | 11.91±2.71 | 8.18±0.72 | 122.03±127.90 | 3.59±0.71 |
| E2 | Creek | 15 | 12.13±1.62 | 8.47±0.51 | 40.27±16.20 | 3.46±0.47 |
| E3 | Creek | 15 | 12.01±2.50 | 8.43±0.59 | 87.62±124.47 | 3.08±0.62 |
| E4 | Creek | 9 | 10.88±1.27 | 8.55±0.80 | 35.44±13.99 | 3.50±0.88 |
| E5 | Pond | 15 | 10.47±3.89 | 7.83±0.60 | 140.83±114.28 | 3.12±0.70 |
| E6 | Pond | 9 | 13.36±2.77 | 8.48±0.40 | 33.00±26.46 | 3.60±0.82 |
Cluster analysis (UPGMA, Bray-Curtis) values lower than 30% showed that there is heterogeneity among months and stations in the lotic systems. The November and December 2017 samples from station E1 have the highest similarity value (greater than 90%), and these 2 samples are similar to the April sample from station E1 (Fig. 3). Samples from station E2 for April, May, July and October also show high similarity (close to 80%). The rest of the samples have higher temporal similarity. Samples from stations E2, E3 and E4 in November and December are associated at a similarity of approximately 60%; samples from stations E3 and E4 in April and May, samples from stations E3 and E4 in July and samples from stations E3 and E4 in October are grouped together (Fig. 3).
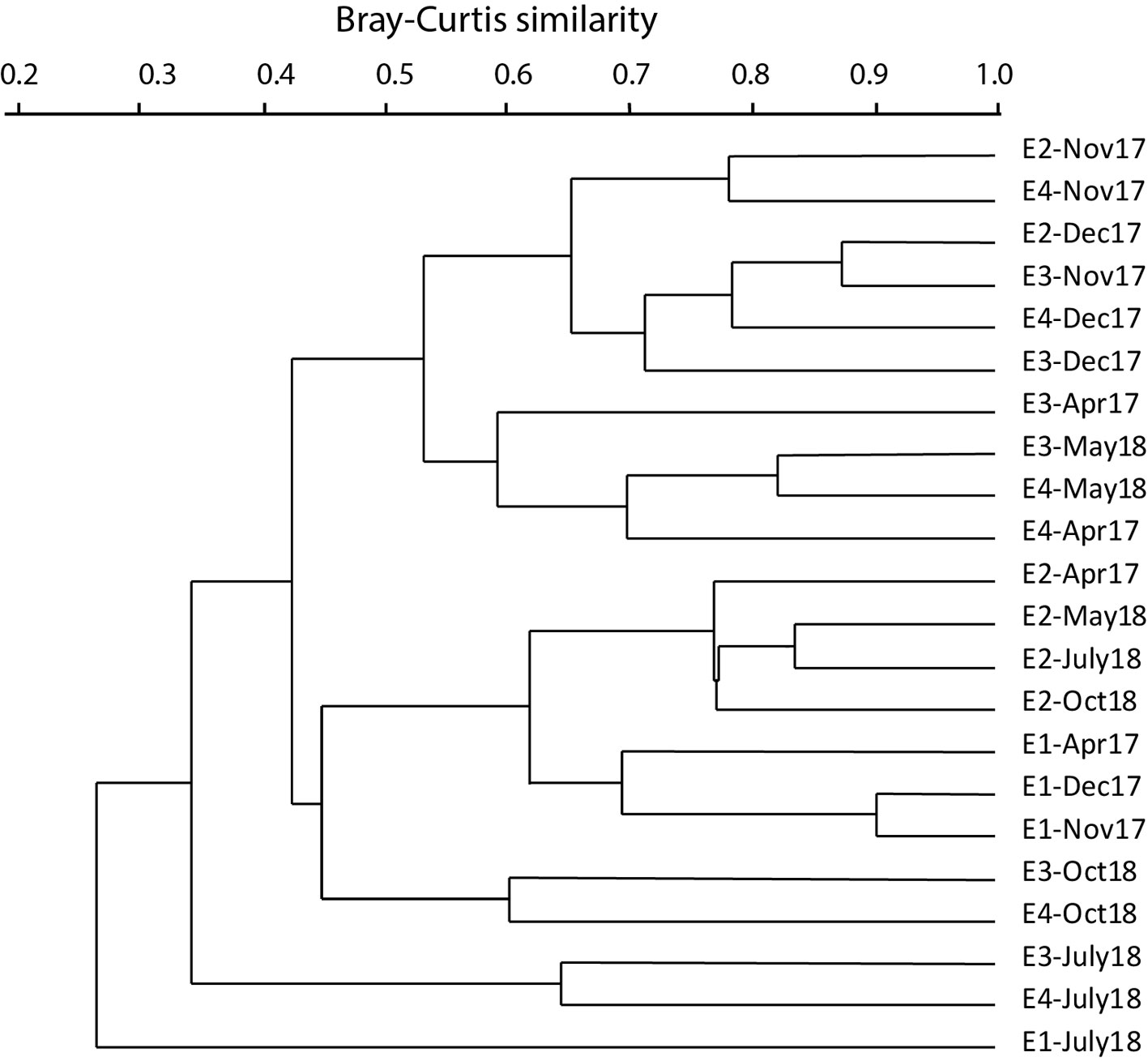
Table 3
Anova test for physicochemical variables. p < 0.0000 for all comparisons.
| DF | F | ||
| Temperature [°C] | Month | 5 | 352,260 |
| Stations | 5 | 116,611 | |
| Interaction | 16 | 127,862 | |
| pH | Month | 5 | 311,701 |
| Stations | 5 | 264,741 | |
| Interaction | 16 | 346,264 | |
| Conductivity [µS/cm] | Month | 5 | 3084,573 |
| Stations | 5 | 156,643 | |
| Interaction | 16 | 84,871 | |
| Dissolved oxygen (mg/L) | Month | 5 | 5,970,694 |
| Stations | 5 | 715,104 | |
| Interaction | 16 | 483,699 |
ANOSIM determined significant differences (R = 0.0185; p < 0.05) in macrobenthos composition between stations E1 and E3, while E3 and E4 (R = 0.946, p < 0.05) are more similar to each other, as are E1 and E2 (R = 0.248, p < 0.05). The SIMPER analysis showed that average dissimilarity of the 4 stations was 55%. The main families that contributed to that dissimilarity were Chironomidae (14.34%), Naididae (11.64%), Lumbriculidae (5.37%) and Limnesiidae (5.29%).
Table 4
Average relative abundance of aquatic macroinvertebrates in lotic environments (stations E1 to E4) and lentic environments (stations E5 to E6) in Callalli-Oscollo, Arequipa-Cusco, Peru. High values for each sampling station in bold.
| Family | E1 | E2 | E3 | E4 | E5 | E6 | |||
| Insecta | Diptera | Ceratopogonidae | 0.09 | 0.3 | |||||
| Chironomidae | 56.35 | 50.57 | 21.15 | 23.53 | 12.75 | 5.99 | |||
| Ephydridae | 1.02 | 1.67 | 0.07 | 0.08 | 0.16 | ||||
| Muscidae | 0.71 | 0.11 | 0.27 | 0.14 | 0.02 | 0.03 | |||
| Simuliidae | 0.1 | 0.33 | 0.41 | 0.29 | |||||
| Syrphidae | 1.77 | ||||||||
| Tabanidae | 0.2 | 0.16 | 0.42 | ||||||
| Tipulidae | 0.24 | 0.14 | 0.03 | 0.01 | |||||
| Coleoptera | Dytiscidae | 0.19 | |||||||
| Elmidae | 9.05 | 9.99 | 4.63 | 0.17 | 0.07 | ||||
| Ephemeroptera | Baetidae | 0.15 | 0.67 | 7.51 | 1.64 | 1.39 | 0.02 | ||
| Leptophlebiidae | 0.2 | 0.5 | 0.71 | 1.02 | |||||
| Hemiptera | Corixidae | 0.05 | 0.11 | 0.11 | 31.46 | 8.64 | |||
| Saldidae | 2.99 | 0.15 | 0.93 | 3.53 | |||||
| Odonata | Coenagrionidae | 0.06 | |||||||
| Plecoptera | Gripopterygidae | 0.02 | 0.48 | ||||||
| Trichoptera | Hydrobiosidae | 0.59 | 1.6 | 1.69 | |||||
| Hydroptilidae | 0.05 | 1.15 | 5.35 | 0.42 | |||||
| Leptoceridae | 0.09 | 0.04 | 0.03 | ||||||
| Limnephilidae | 0.04 | ||||||||
| Crustacea | Amphipoda | Hyalellidae | 25.28 | 0.07 | 0.39 | 0.34 | 3.16 | 0.13 | |
| Podocopida | Cyprididae | 0.04 | 0.04 | 12.73 | 45.56 | ||||
| Arachnida | Oribatida | Limnesiidae | 15.86 | 6.06 | 6.5 | 0.16 | 0.03 | ||
| Limnozetidae | 1.39 | 0.74 | 1.62 | 0.1 | |||||
| Oligochaeta | Lumbriculidae | 9.44 | 2.5 | 6.94 | 23.17 | 0.01 | 0.12 | ||
| Naididae | 3.45 | 14.62 | 34.61 | 30.57 | 4.62 | 25.48 | |||
| Hirudinida | Glossiphoniidae | 0.13 | 0.98 | 0.31 | 12.46 | 1.83 | |||
| Family A | 1.25 | 0.03 | |||||||
| Turbellaria | Tricladida | Dugesiidae | 0.26 | 0.36 | 0.08 | 0.04 | |||
| Typhloplanida | Typhloplanidae | 0.03 | 0.01 | ||||||
| Gastropoda | Planorbidae | 0.93 | 0.11 | 0.51 | 0.26 | 0.02 | |||
| Bivalvia | Sphaeriidae | 0.18 | 0.05 | 0.24 | |||||
| Nematoda | Longidoridae | 0.1 | 0.04 | 0.08 | 15.46 | 11.69 | |||
| Family A | 0.1 | 0.11 | 0.02 |
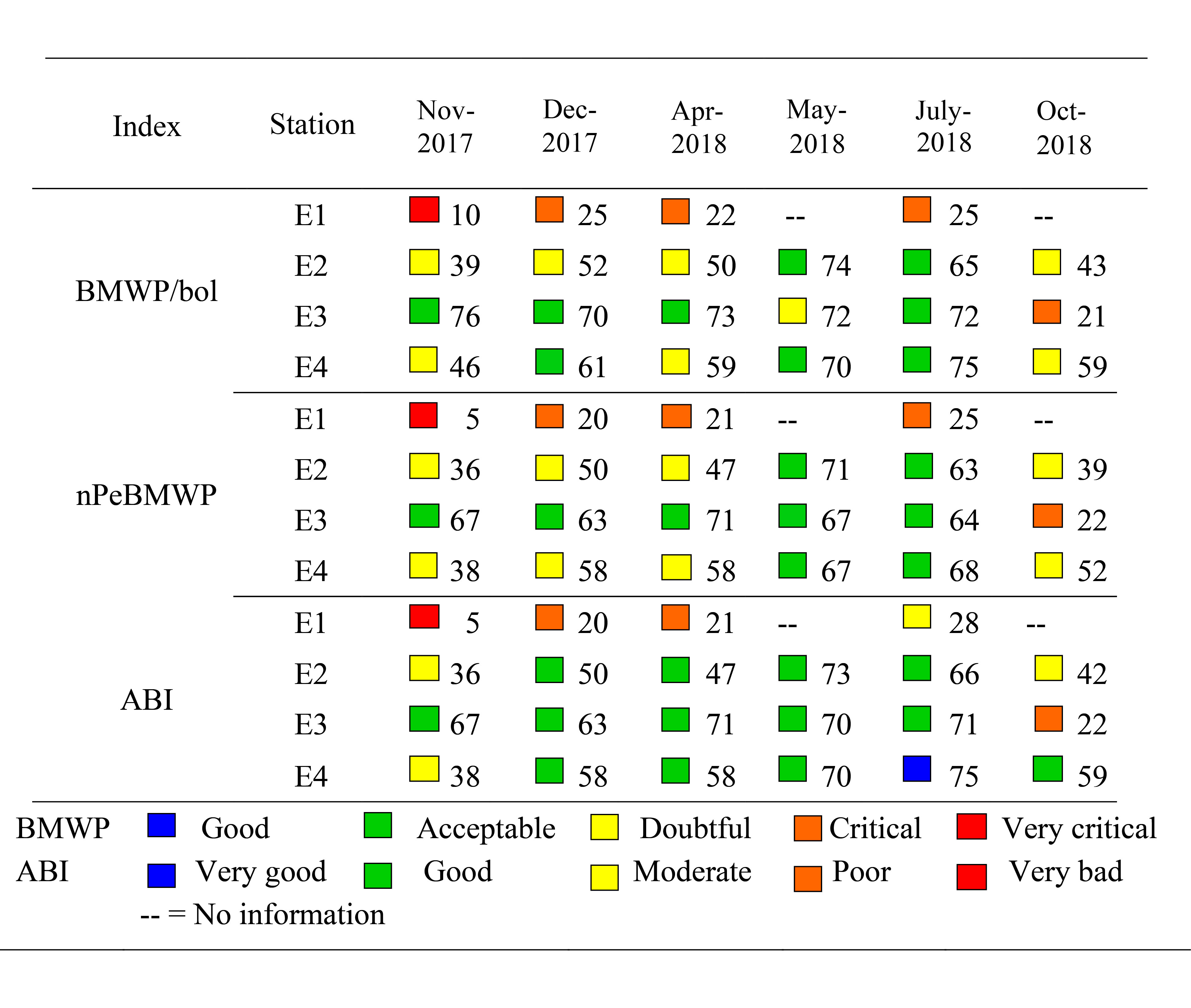
Determination of ecological quality using the biotic indices BMWP/Bol, nPeBMWP and ABI showed lowest scores for station E1, classifying it as doubtful quality according to BMWP/Bol and nPeBMWP, and very bad and poor according to ABI. For stations E2, E3 and E4, ecological quality was found to be good and acceptable according to the indices BMWP/Bol and nPeBMWP, and very good according to ABI, depending on the months evaluated (Table 5).
Macroinvertebrates in lentic environments
A total 55,553 individuals were counted, distributed among 23 families. Sampling stations in ponds E5 and E6 had richness of 21 and 18 families respectively (Table 4).
At station E5, richness ranged from 13 (July) to 4 (April). The families with high relative abundance at station E5 were Corixidae (31.46%), Longidoridae (15.46%), Chironomidae (12.75%), Cypridae (12.73%) and Glosiphoniidae (12.46%).
At station E6, richness ranged from 17 (November) to 10 (December and May). The families with highest relative abundance were Cyprididae (45.56%), Naididae (25.48%) and Longidoridae (11.69%). Abundance was highest in December and lowest in May (Table 3).
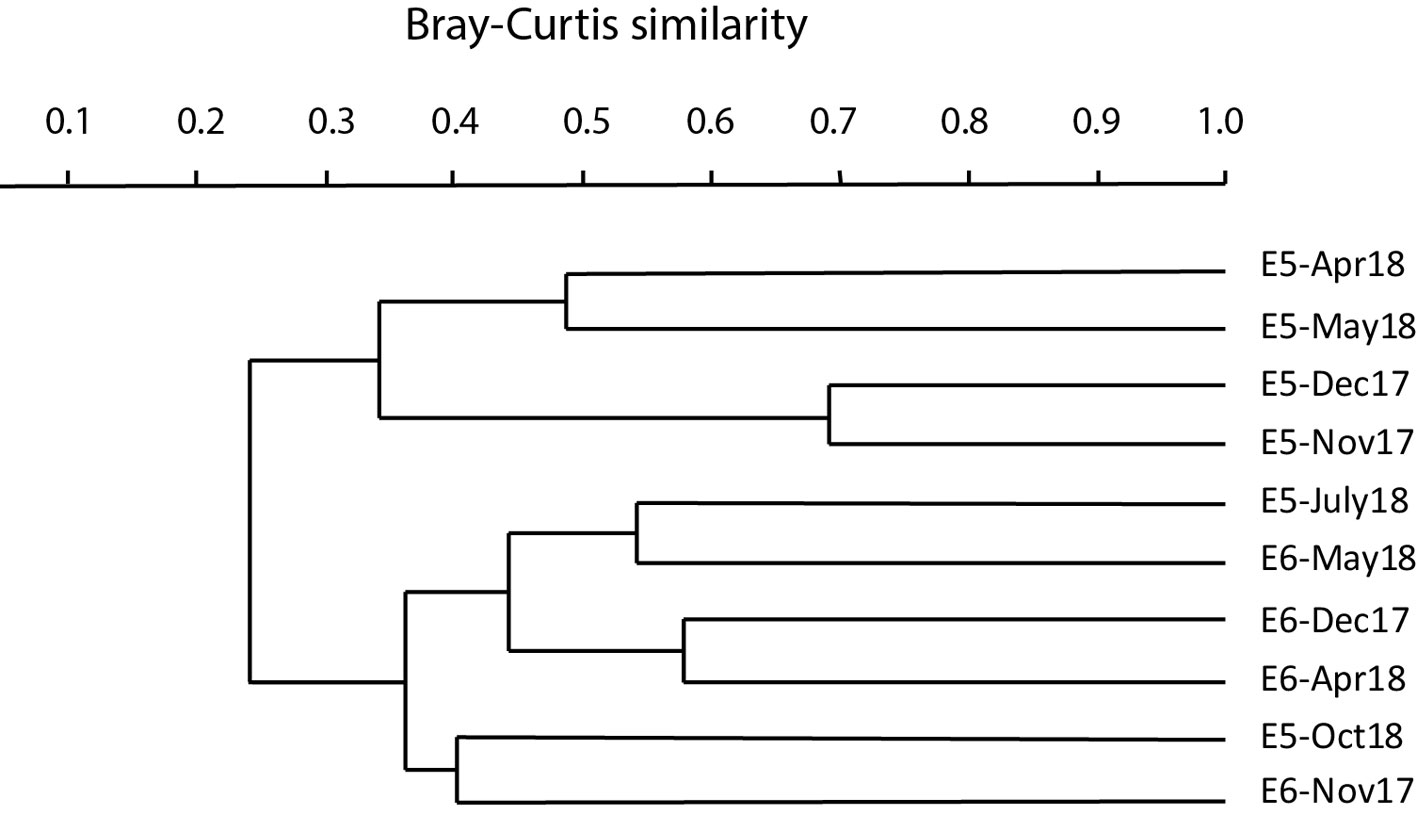
At stations E5 and E6, Shannon’s index ranged from 0.99 (E6 in December) to 1.72 (E5 in October). Both stations had 3 to 6 effective families. The dominance was low in May (0.22 at E6 and 0.27 at E5) and highest in December (0.43 at E5 and 0.51 at E6). Evenness index was lowest (0.43) at E6 in December and highest (0.79) at E5 in April (Fig. 4).
Cluster analysis (UPGMA, Bray-Curtis) shows low similarity, less than 50%, demonstrating that there is heterogeneity among months and pond stations. The highest similarity values (over 70%) were recorded for November and December 2017 at station E5. These 2 samples in turn are grouped, with less similarity, with the samples from April and May. E5 samples from July and October are grouped with E6 samples (Fig. 5).
ANOSIM determined significant differences (R = 0.3611; p < 0.05) in macrobenthos composition between stations E5 and E6. The SIMPER analysis showed that average dissimilarity of the 2 stations was 74.84%. The main families that contributed to that dissimilarity were Cyprididae (31.99%), Naididae (17.39%), Longidoridae (9.03%) and Corixidae (7.12%).
Discussion
The dissolved oxygen values recorded during this study were lower than those reported in previous studies for nearby and similar areas (Luque-Fernández et al., 2020; Molina et al., 2008). The remaining parameters were within the known values for the region. The electrical conductivity had very high values in some stations in the month of December, coinciding with the rainy season. These values are probably due to the increase in suspended sediment during those periods (Fossati et al., 2001). The taxonomic composition of the macroinvertebrate communities in the Callalli-Oscollo is similar to that reported for other similar environments (Custodio & Chanamé, 2016; Luque-Fernández et al., 2020; Molina et al., 2008; Nieto et al., 2016).

Macroinvertebrates in lotic environments
The richness recorded in lotic environments (30 families) is similar to that observed by Molina et al. (2008) for rivers in the Cordillera Real (Bolivia) and by Custodio and Chanamé (2016) for rivers in Junín (Peru). The data recorded in the current study therefore reinforce the hypotheses that claim that richness values such as those observed are representative for high Andean lotic ecosystems (Custodio & Chanamé, 2016). The current study reports 15 insect families at E2 and 14 at E3, both these stations being located in the highest zones (4,672 and 4,630 m asl, respectively). These values are lower than those reported for zones of similar altitude in Bolivia [20 families for rivers in the Cordillera Real at 4,400 m asl, (Molina et al., 2008); 17 families for River Hichu-Khota and River Ovejuyo at 4,320 m asl (Wasson & Marín, 1988); 18 families for River Kaluyo at 4,280 m asl. (Franken & Marín, 1992), among others]. Like the observations by Luque-Fernández et al. (2020), richness of families does not differ between dry and rainy seasons, although authors mention the presence of families at each site which are typical of each season.
Highest abundance values were found for the dry season, mainly in May, July and November. This agrees with Jacobsen and Encalada (1998) and González et al. (2013), who report that the number of organisms is higher in the dry season than in the rainy season. Molina et al. (2008) make a similar observation for rivers in Bolivia during the dry season when low values in hydrological parameters are associated to high densities and an increase in the number of taxa.
Chironomidae and Naididae were the most abundant and constant families in the lotic environments studied. Tapia et al. (2018) report that these are the families that contribute the most to the diversity of freshwater ecosystems, while Molina et al. (2008) mention oligochaetes as the most abundant organisms in high Andean ecosystems. Chironomidae is a resistant and resilient taxon to anthropogenic action that leads to the impoverishment of biological communities, and consequently, ecosystem alteration (Maroneze et al., 2011). Other families, such as Limnesidae, were recorded by Luque-Fernández et al. (2020) for the Chalhuanca region, which is close to the current study area and located at a similar altitude.
The high Shannon-Wiener index values at stations E1, E3 and E4 after the rainy seasons, are in agreement with Custodio and Chaname (2016), who report higher values during low water level months. The dam upstream from station E1, in contrast to the other sampling stations, causes alterations in the environment, such as temperature changes (Hendricks, 1984), changes in the downstream hydrological regime, causing changes in water flow and flow rate regulation, thereby leading to homogeneity of microenvironments and dragging benthic fauna (Fossati et al., 2001; Gutiérrez-Yurrita et al., 2002; Hurtado et al., 2005).
The ABI has proved to be efficient in evaluations of high Andean rivers above 2,000 m asl and areas with anthropic influence. The ABI has been applied in Colombia, Ecuador and Peru (Meneses-Campo et al., 2019).
Macroinvertebrates in lentic environments
Twenty-six families were recorded in lentic environments, the most abundant phyla being Arthropoda and Annelida. This coincides with studies by Huamán et al. (2002), Chocano-Arévalo (2005), Ludeña-Guamán and Rojas-López (2014) and Velásquez (2018), among others, in environments at elevations of 3,000 to 4,000 m asl. Twenty-three families were reported from ponds in the Maya region in Guatemala (Reyes-Morales, 2013); 22 in ponds in Bolivia (Lino et al., 2004), high Andean wetlands in Chile (Villagrán-Mella et al., 2006) and the wetland in Jaboque, Bogotá, located at 2,540 m asl (Rivera, 2011).
The highest abundance values in ponds were recorded in July (dry season) for station E5 and November (transition season) for station E6. This coincides with observations by other authors, who report greater abundance during the dry season (Gómez-Hernández & Salazar-Santana, 2015; Gutiérrez, 2005; Lino et al., 2004; Pineda & Quiroz, 2015; Rico-Sánchez et al., 2014; Velásquez, 2018). Ortiz (2005) says that the increase in abundance during the dry season is due to the decrease in flow and in total solids. Lino et al. (2004) suggest that after the period of disturbance by flooding, the favorable conditions trigger rapid development of macroinvertebrate fauna and consequent increase in their abundance during the transition season, since the organisms must attain their maximum development before the new period of high waters, when their numbers drop drastically. Another hypothesis suggests that benthic fauna density decreases when the water level rises, and increases when the water level drops (Fittkau et al., 1975; Irmler, 1975; Nessimian et al., 1998).
The families with the greatest abundance at stations E5 and E6 were Cyprididae, Naididae and Corixidae. In particular, the presence of Cyprididae was recorded in high Andean ponds in Argentina, finding a frequency of 100% (Locascio de Mitrovich et al., 2005), as well as in Lake Titicaca (Dejoux & Iltis, 1991), and in Chalhuanca (Luque-Fernández et al., 2020). In the current study, the greatest abundance of Cyprididae was recorded at station E6 in December (68.29%), which is the transition to the rainy season, so the presence of these ostracods (eggs, larval stages, juveniles, and adults) is limited to still water zones, which can be dragged towards floodable zones and modify populations on the lake shore (Dejoux & Iltis, 1991). Ostracods play a key role in the trophic web, being the main energy pathway in limnetic ecosystems (Fabián & Cruz-Pizarro, 1997). In addition, Cyprididae is considered an excellent aquatic bioindicator because it is sensitive to changes in salinity, temperature, conductivity, depth, and composition of the water (Echeverría, 2016; Meisch, 2000; Pérez et al., 2010). Moreover, the type of substrate (sand, clay, silt) and food availability also influence its distribution and abundance (Chacón & Rivas, 2009).
The high abundance of the family Naididae coincides with the observations by Ludeña-Guamán and Rojas-López (2014) and Huamán et al. (2002). Its presence is related to low oxygen concentrations and plentiful detritus, making it useful as a bioindicator of water pollution (Rivera, 2011).
The greatest abundance of the family Corixidae was at station E5 during November (transition period) and July (dry period). Corixidae prefers slow-flowing, small or medium size, shallow lentic waters, with free surface and little submerged vegetation (Roldán, 1988), which are features present at station E5. It is worth noting that Corixidae are primary consumers in aquatic environments, so their high abundance generates high availability for higher trophic levels, since they are preyed upon by other aquatic invertebrates and vertebrates (Barahona et al., 2005). At station E5, highest abundance and richness of macroinvertebrates was recorded in July, with high Shannon index (1.50). However, evenness was lowest also in July (0.57), explained by the high abundance of Corixidae (over 50%). At station E6, the highest abundance and lowest richness occurred in December, with the lowest value for the Shannon index (0.99).
The biotic indices found in the current study based on analysis of the macrobenthos community provide information on the ecological condition and enable temporal and geographic comparisons. The results show that the impact of El Pañe reservoir and livestock-raising activity is reflected by lower ecological quality than in more pristine waters. Likewise, E3 showed a lower water quality than the other stations. This is probably due to the fact that there are residents that use the resource, especially for cattle. The data contributed provide information that will enable monitoring and management of the affected areas.
Acknowledgments
The authors are grateful for the suggestions and the comments provided by the reviewers. We thank Universidad Nacional de San Agustín de Arequipa for financing the research through Contract IBA-0010-2017-UNSA.
References
Acosta, R. (2001). Patrones de diversidad espacio-temporal de insectos acuáticos y caracterización limnológica en la quebrada Cantón, Tributaria del río Rímac (Huarochirí, Lima). Libro de Resúmenes de la XLIII Convención Nacional de Entomología. Huancayo, Perú. Noviembre 2001, 75.
Acosta, R., Ríos-Touma, B., Rieradevall, M., & Prat, N. (2009). Propuesta de un protocolo de evaluación de la calidad ecológica de ríos Andinos (C.E.R.A) y su aplicación en dos cuencas en Ecuador y Perú. Limnetica, 28, 35–64. http://doi.org/10.23818/limn.28.04
Autoridad Nacional del Agua. (2014). Evaluación integral de la calidad del agua de los embalses y ríos que conforman el sistema hidráulico Chili-Arequipa. Informe Técnico 21. Retrieved from: http://repositorio.ana.gob.pe/handle/20.500.12543/2051 accessed on October 9th, 2021.
Báez, S., Jaramillo, L., Cuesta, F., & Donoso, D. A. (2016). Effects of climate change on Andean biodiversity: a synthesis of studies published until 2015. Neotropical Biodiversity, 2, 181–194. https://doi.org/10.1080/23766808.2016.1248710
Barahona, J., Millan, A., & Velasco, J. (2005). Population dynamics, growth and production of Sigara selecta (Fieber, 1848) (Hemiptera, Corixidae) in Mediterranean hypersaline stream. Freshwater Biology, 50, 2101–2113. https://doi.org/10.1111/j.1365-2427.2005.01463.x
Buss, D. F., Carlisle, D. M., Chon, T., Culp, J., Harding, J. S., Keizer-Vlek, H. E. et al. (2015). Stream biomonitoring using macroinvertebrates around the globe: a comparison of large-scale programs. Environmental Monitoring and Assessment, 187, 4132. https://doi.org/10.1007/s10661-014-4132-8
Chacón, L., & Rivas, P. (2009). Paleontología de invertebrados. Madrid: Sociedad Española de Paleontología.
Chocano-Arévalo, L. (2005). Las zonas altoandinas peruanas y su ictiofauna endémica. Revista Digital Universitaria, 6, 2–13.
Custodio, M., & Chanamé, F. (2016). Análisis de la biodiversidad de macroinvertebrados bentónicos del río Cunas mediante indicadores ambientales, Junín-Perú. Scientia Agropecuaria, 7, 33–44. http://doi.org/10.17268/sci.agropecu.2016.01.04
Dejoux, C., & Iltis, A. (1991). El lago Titicaca: síntesis de conocimiento limnológico actual. La Paz: ORSTOM. lnstitut Francais de Recherche Scientifique pour le Développement en Cooperation.
Domínguez, E., & Fernández, H. (2009). Macroinvertebrados bentónicos sudamericanos. Sistemática y Biología. Tucumán, Argentina: Fundación Miguel Lillo.
Echeverría, P. (2006). Ostrácodos (Crustacea: Ostracoda) en lagos kársticos (Montebello, Selva Lacandona y El Petén) en el norte de los neotrópicos en relación a un gradiente altitudinal: diversidad, distribución y ecología (Tesis). Universidad de San Carlos de Guatemala.
Fabián, D., & Cruz-Pizarro, L. (1997). Variaciones espaciales y temporales del zooplancton en un lago monomíctico eutrófico (lago Ton-Ton, Uruguay). Limnetica, 13, 55–68.
Fittkau, E. J., Irmler, U., Junk, W. J., Reiss, F., & Schmidt, G. W. (1975). Productivity, biomass, and populations dynamics in Amazonian water bodies. In F. B. Golley, & E. Medina (Eds.), Tropical ecological systems. Ecological studies (analysis and synthesis), Vol 11 (pp. 289–311). Berlín, Heidelberg: Springer. http://dori.org/10.1007/978-3-642-88533-4_20
Fossati, O., Wasson, J. G., Hery, C., Marin, R., & Salinas, G. (2001). Impact of sediment releases on water chemistry and macroinvertebrate communities in clear water Andean streams (Bolivia). Archiv für Hydrobiologie, 151, 33–50. http://doi.or./10.1127/archiv-hydrobiol/151/2001/33
Franken, M., & Marín, R. (1992). Influencia de una fábrica de estuco sobre un ecosistema acuático en la ciudad de La Paz. Ecología en Bolivia, 19, 73–96.
Gómez-Hernández, S. J., & Salazar-Santana, C. M. (2015). Abundancia y biomasa de macroinvertebrados en relación de cuatro tipos de sustratos y de variables fisicoquímicas en la laguna La Virginia, Páramo de Sumapaz (Tesis). Universidad de Bogotá Jorge Tadeo Lozano, Colombia.
González, N., Sánchez, S., & Mairena, A. (2013). Macro-
invertebrados bentónicos como bioindicadores de calidad de agua del trópico húmedo en las microcuencas de los alrededores de Bluefields, RAAS. Wani, 68, 53–63. https://doi.org/10.5377/wani.v68i0.1354
González-Zuarth, C. A., Vallerino, A., Pérez-Jiménez, J. C., & Low-Pfeng, A. M. (Eds.). (2014). Bioindicadores: guardianes de nuestro futuro ambiente. México D.F.: Editorial S y G Editores.
Gutiérrez, M. (2005). Composición de las comunidades de insectos acuáticos asociadas a la vegetación en la llanura de inundación de moxos (Beni-Bolivia) (Tesis). Universidad Mayor de San Andrés, Bolivia.
Gutiérrez-Yurrita, P., Morales-Ortiz, A., Oviedo A., & Ramírez, C. (2002). Distribution, spread, habitat characterization and conservation of the crayfish species (Cambaridae) in Querétaro (Central México). Freshwater Crayfish, 13, 288–297.
Hammer, O., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological Statistics Software for education and data analysis. Paleontologia Electronica, 4, 1–9.
Hendricks, D. (1984). Impacto de las represas en la calidad del agua. In T. S. Shorr (Ed.), Las represas y sus efectos sobre la salud (pp. 71–88). Ecoguía 1. México D.F.: OPS/OMS.
Huamán, P., Mariano, M., Chanco, M., & Montoya, H. (2002). Estructura del macrobentos de la laguna de Paca, Junín. Revista Peruana de Biología, 9, 29–38. https://doi.org/10.15381/rpb.v9i1.2520
Hurtado, S., Trejo, F., & Gutiérrez-Yurrita, P. (2005). Impor-
tancia ecológica de los macroinvertebrados bentónicos de la subcuenca del Río San Juan, Querétaro, México. Folia Entomologica Mexicana, 44, 271–286.
Irmler, U. (1975). Ecological studies of the aquatic soil invertebrates in three inundation forest of Central Amazonian. Amazoniana, 5, 337–409.
Jacobsen, D. (2008). Tropical high-altitude streams. In D.
Dudgeon (Ed.), Tropical stream ecology (pp. 219–256).
Londres: Elsevier Science. https://doi.org/10.1016/B978-01
2088449-0.50010-8
Jacobsen, D., & Encalada, A. (1998). The macroinvertebrate fauna of Ecuatorian highland streams in the wet and dry season. Archiv für Hydrobiologie, 142, 53–70. http://doi.org/10.1127/archiv-hydrobiol/142/1998/53
Lino, F., Fossati, O., Apaza, R., & Goitia, E. (2004). Diversidad biológica en la llanura de inundación del río Mamoré. Importancia ecológica de la dinámica fluvial. Santa Cruz, Bolivia: Centro de Ecología Simón I. Patifio.
Locascio de Mitrovich, C., Villagra-de Gamundi, A., Juárez, J., & Ceraolo, M. (2005). Características limnológicas y zooplancton de cinco lagunas de la Puna, Argentina. Ecología en Bolivia, 40, 10–24.
Ludeña-Guamán, I., & Rojas-López, V. (2014). Diferencias de las características físicas, químicas y biológicas de dos lagunas de diferente edad y formación, de la Sierra del Sur del Ecuador (Tesis). Universidad de Azuay, Ecuador.
Luque-Fernández, C. R., Pauca, G. A., Villegas Paredes, L. N., Quispe-Turpo, J. d. P., & Villasante-Benavides, J. F. (2020). Macroinvertebrates in a high Andean wetland (Chalhuanca) of southern Peru during the dry and wet season. Aquatic Research, 3, 155–166. https://doi.org/10.3153/AR20014
Maroneze, D. M., Tupinambás, T. H., Alves, C. B. M., Vieira, F., Pompeu, P. S., & Callisto, M. (2011). Fish as ecological tools to complement biodiversity inventories of benthic macroinvertebrates. Hydrobiologia, 673, 29–40. https://doi.org/10.1007/s10750-011-0747-8
Meisch, C. (2000). Freshwater Ostracoda of western and central Europe. In J. Schwoerbel, J., & P. Zwick (Eds.), Süßwasserfauna von Mitteleuropa 8/3. Heidelberg, Berlin: Spektrum Akademischer Verlag.
Medina-Tafur, C. M., Hora-Revilla, I., Ascencio-Gusmán, W., Pereda-Ruíz, R., & Gabriel-Aguilar, R. (2010). Índice Biological Monitoring Working Party (BMWP), modificado y adaptado a tres microcuencas del Alto Chicana. La Libertad, Perú. Uniciencia, 1, 5–20.
Meneses-Campo, Y., Castro-Rebolledo, M., & Jaramillo-Londoño, A. (2019). Comparación de la calidad del agua en dos ríos altoandinos mediante el uso de los índices BMWP/COL. y ABI. Acta Biológica Colombiana, 24, 299–310. https://doi.org/10.15446/abc.v24n2.70716
Merritt, R., Cummins, W., & Berg, M. (2008). An introduction to the aquatic insects of North America, Fourth edition. Dubuque: Kendall/Hunt Publishing Company.
Molina, C., Gibon, M., Pinto, J., & Rosales, C. (2008). Estructura de macroinvertebrados acuáticos en un río altoandino de la Cordillera Real, Bolivia: variación anual y longitudinal en relación a factores ambientales. Ecología Aplicada, 7, 105–116.
Molina, C. I., Pinto, J., & Achá, D. (2022). Selection of macroinvertebrate metrics for rapid assessment of the human impact by biotic conditions of Bolivian Altiplano streams. Journal of South American Earth Sciences, 113, 103638. https://doi.org/10.1016/j.jsames.2021.103638
Moreno C. E. (2001). Métodos para medir la biodiversidad. M&T Manuales y Tesis Sociedad Entomológica Aragonesa, 1.
Nessimian, J., Dorvillé, L., Sanseverino, A., & Baptista, D. (1998). Relation between flood pulse and functional composition of the macroinvertebrate benthic fauna in the lower Rio Negro, Amazonas, Brazil. Amazoniana, 15, 35–50.
Nieto, C., Malizia, A., Carilla, J., Izquierdo, A., Rodríguez, J., Cuello, S. et al. (2016). Patrones espaciales en comunidades de macroinvertebrados acuáticos de la Puna Argentina. Revista de Biología Tropical, 64, 747–762. https://doi.org/10.15517/rbt.v64i2.18801
Ortiz, L. L. (2005). La bioindicación de la calidad del agua: importancia de los macroinvertebrados en la cuenca alta del rio Juan Amarillo. Umbral Científico, 7, 5–11.
Oyague, E. (2005). Variación espacial del ensamblaje de macroinvertebrados bentónicos en la cuenca del río Ponasa (distritos de Shaboyacu, Tingo de Ponasa y Pucacaca, Provincia de Picota, Departamento de San Martín) y su utilidad como indicadores de la calidad del agua. Lima: Universidad Nacional Agraria la Molina. http://doi.org/10.13140/RG.2.2.19993.21606
Paredes, C., Iannacone, J., & Alvariño, L. (2004). Macro-
invertebrados bentónicos como indicadores biológicos
de la calidad de agua en dos ríos de Cajamarca y Amazonas, Perú. Revista Peruana de Entomología, 44, 107–118.
Pérez, L., Lorenschat, J., Bugja, R., Brenner, M., Scharf, B., & Schwalb, A. (2010). Distribution, diversity and ecology of modern freshwater ostracodes (Crustacea), and hydrochemical characteristics of Lago Petén Itzá, Guatemala. Alemania: Journal of Limnology, 69, 146–159. https://doi.org/10.4081/jlimnol.2010.146
Pineda, J., & Quiroz, G. (2015). Caracterización de la comunidad de macroinvertebrados y estimación de la calidad del agua de las lagunas de Chingaza, del medio y El Arnical en el Parque Nacional Natural Chingaza (Tesis). Universidad Santo Tomás, Bogotá.
Posada, A., Roldán, G., & Ramírez, J. (2000). Caracterización fisicoquímica y biológica de la calidad de aguas de la cuenca de la quebrada Piedras Blancas, Antioquia, Colombia. Revista de Biología Tropical, 48, 59–70.
Prat, N., Rieradevall, M., Acosta, R., & Villamarín, C. (2011). Guía para el reconocimiento de las larvas de Chironomidae (Diptera) de los ríos Altoandinos de Ecuador y Perú: clave para la determinación de los géneros. Barcelona: Departamento de Ecología, Universidad de Barcelona.
Reyes-Morales, F. (2013). Macroinvertebrados acuáticos de los cuerpos lénticos de la Región Maya, Guatemala. Revista Científica, 23, 7–16.
Rico-Sánchez, A., Rodríguez-Romero, A., López-López, E., & Sedeño-Díaz, J. (2014). Patrones de variación espacial y temporal de los macroinvertebrados acuáticos en la Laguna de Tecocomulco, Hidalgo (México). Revista de Biología Tropical, 62 S2, 81–96. https://doi.org/10.15517/rbt.v62i0.
15780
Ríos-Touma, B., Acosta, R., & Prat, N. (2014). The Andean Biotic Index (ABI): revised tolerance to pollution values for macroinvertebrate families and index performance. Revista de Biología Tropical, 62 S2, 249–273. https://doi.org/10.15517/rbt.v62i0.15791
Rivera, J. (2011). Relación entre la composición y biomasa de la comunidad de macroinvertebrados acuáticos y las variables físicas y químicas en el humedal Jaboque (Tesis). Facultad de Ciencias: Universidad Nacional de Colombia. Bogotá.
Rocabado, G., & Goitia, E. (2011). Guía para la evaluación de la calidad acuática mediante el índice BMWP/Bol. La Paz: Ministerio de Medio Ambiente y Agua de Bolivia.
Roldán, G. (1988). Guía para el estudio de los macroinverte-
brados acuáticos del departamento de Antioquia. Antioquia: Universidad de Antioquia.
Roldán, G. (2003). Bioindicación de la calidad de agua en Colombia. Medellín: Universidad de Antioquia. Colección de Ciencia y Tecnología.
Roldán, G. (2012). Macroinvertebrados como indicadores de calidad de agua. Bogotá: Imprenta Nacional de Colombia.
Tapia, L., Sánchez, T., Baylón, M., Jara, E., Arteaga, C., Maceda, D. et al. (2018). Invertebrados bentónicos como bioindicadores de calidad de agua en Lagunas Altoandinas del Perú. Ecología Aplicada, 17, 149–163. http://doi.org/
10.21704/rea.v17i2.1235
Thorp, J., & Lovell, L. (2015). Phylum Annnelida. In J. Thorp, & C. Rogers. Covich’s freshwater invertebrates, Vol. II. Key to Neartic Fauna (pp. 223–263). Amsterdam. Academic Press. https://doi.org/10.1016/C2010-0-65589-1
Velásquez, M. (2018). Macroinvertebrados bentónicos como bioindicadores de calidad de agua en lagunas de la cabecera de cuenca del río Rímac y cuenca del Mantaro de la región central del Perú, 2015. (Tesis). Universidad Nacional del Santa. Chimbote.
Villagrán-Mella, R., Aguayo, M., Parra, L. E., & González, A. (2006). Relación entre características del hábitat y estructura del ensamble de insectos en humedales palustres urbanos del centro-sur de Chile. Revista Chilena de Histo-
ria Natural, 79, 195–211. http://doi.org/10.4067/S0716-
078X2006000200006
Wasson, J. G., & Marín, R. (1988). Tipología y potencialidades biológicas de los ríos de altura en la región de La Paz (Bolivia): metodologías y primeros resultados. Memorias de la Sociedad de Ciencias Naturales de La Salle, 48, 97–122.