Riqueza, distribución geográfica y conservación de Lycianthes serie Meizonodontae (Capsiceae, Solanaceae)
Marco Antonio Anguiano-Constante a, Guadalupe Munguía-Lino b, c, Enrique Ortiz d, José Luis Villaseñor d y Aarón Rodríguez a, c, *
a Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, camino Ramón Padilla Sánchez 2100, 45200 Las Agujas, Zapopan, Jalisco, México
b Cátedra Conacyt-Universidad de Guadalajara, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, camino Ramón Padilla Sánchez 2100, 45200 Las Agujas, Zapopan, Jalisco, México
c Laboratorio Nacional de Identificación y Caracterización Vegetal, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, camino Ramón Padilla Sánchez 2100, 45110 Las Agujas, Zapopan, Jalisco, México
d Departamento de Botánica, Instituto de Biología, Universidad Nacional Autónoma de México, Apartado postal 70-233, 04510 Ciudad de México, México
*Autor para correspondencia: aaron.rodriguez@cucba.udg.mx (A. Rodríguez)
Resumen
Lycianthes serie Meizonodontae Bitter (Solanaceae) es un grupo monofilético, integrado por 8 especies y 2 variedades. El objetivo del presente trabajo fue analizar la riqueza, distribución y estado de conservación de la serie. Mediante el uso de sistemas de información geográfica (SIG), se evaluó la riqueza por latitud, longitud, altitud, división política, provincias biogeográficas, tipos de vegetación, una cuadrícula de 83 × 83 km y modelos de distribución potencial. También se estimó la distribución geográfica, así como el estado de conservación para cada taxón, de acuerdo con el criterio B2 de la UICN. Lycianthes serie Meizonodontae se distribuye desde el noreste de México hasta el centro de Costa Rica, pero México es su centro de diversificación. La riqueza de especies se concentra en el centro de México, donde converge la Faja Volcánica Transmexicana (FVT) y la Cuenca del río Balsas (CB). Por su área de ocupación (AOO), L. hintonii y L. staburckii están en peligro crítico de extinción (CR), mientras que las demás especies se consideran en preocupación menor (LC). Los modelos de distribución potencial infieren que la riqueza de Lycianthes serie Meizonodontae se localiza a lo largo de la FVT.
Palabras clave:
Faja Volcánica Transmexicana; SIG; Categorías de riesgo; Distribución potencial
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Richness, geographic distribution and conservation of Lycianthes series Meizonodontae (Capsiceae, Solanaceae)
Abstract
Lycianthes series Meizonodontae Bitter (Solanaceae) is monophyletic and includes 8 species and 2 varieties. The aim of this paper is to analyze the richness, distribution and conservation status of the series. Using geographic information systems (GIS), we evaluated the richness by latitude, longitude, elevation, political divisions, biogeographic provinces, vegetation types, a 83 × 83 km grid cell and species distribution models. Also, we estimated geographic distribution and conservation status for each taxon, according to criterion B2 of the IUCN. Lycianthes series Meizonodontae distributes from northeast of Mexico to central Costa Rica, with Mexico as its main center of diversification. The species richness concentrates in Central Mexico, where the Transmexican Volcanic Belt and the Balsas Basin converge. Based on their areas of occupancy (AOO), L. hintonii and L. starbuckii are considered as critically endangered (CR), while the rest of the species are least concern (LC). The potential distribution models suggest that the main richness of Lycianthes series Meizonodontae occurs in the Transmexican Volcanic Belt.
Keywords:
Transmexican Volcanic Belt; GIS; Risk category; Potential distribution
Introducción
La familia Solanaceae Juss. posee una distribución cosmopolita, incluyendo unos 100 géneros y 2,500 especies (Wilf et al., 2017). La mayor diversidad se localiza en Centro y Sur América (Barboza et al., 2016) y por su número de especies, los géneros más importantes son Solanum L. (1400), Lycianthes (Dunal) Hassl. (200), Cestrum L. (175), Nicotiana L. (95), Physalis L. (80) y Lycium L. (75) (Aubriot et al., 2016; D’Arcy, 1991). México es un centro de diversificación de Solanaceae, ocupando el lugar 14 entre las familias de angiospermas más diversas del país (Villaseñor, 2016). Martínez et al. (2017) estiman que en el país crecen 33 géneros y 387 especies, en tanto que Villaseñor (2016) registra la existencia de 34 géneros y 407 especies.
El género Lycianthes (Dunal) Hassl. fue separado de Solanum por Hassler en 1917. Posteriormente, Bitter (1919) realizó la primera monografía del género, en la que incluyó aproximadamente 150 especies, agrupadas en secciones y series. Por décadas, Lycianthes no fue aceptado a nivel genérico; sin embargo, evidencia morfológica (D’Arcy, 1986) apoya su reconocimiento como género. Por otro lado, análisis moleculares (Bohs y Olmstead, 1997; Olmstead, 2013; Olmstead y Palmer, 1997; Olmstead et al., 2008; Särkinen et al., 2013) sugieren su relación filogenética con Capsicum L.
Lycianthes es de afinidad Neotropical y se distingue por su cáliz 0-10 dentado y sus anteras con dehiscencia poricida apical. Es un grupo parafilético que divergió aproximadamente hace 13 Ma (Särkinen et al., 2013). Incluye alrededor de 200 especies, distribuidas en las regiones tropicales de América y Asia-Pacífico. Es uno de los 3 géneros más diversos de Solanaceae e incluye hierbas, arbustos y lianas. Su mayor diversidad se concentra en América; en México crecen 39 especies, de las cuales 14 son endémicas.
Lycianthes serie Meizonodontae Bitter fue descrita en 1919, dentro de la sección Perennans Bitter (Bitter, 1919). Después, se describieron L. rzedowskii E. Dean, L. starbuckii E. Dean y L. hintonii E. Dean (Dean, 1994, 2004). Los estudios sobre la serie incluyen aspectos taxonómicos (Dean, 1994, 1995, 2004), filogenéticos y etnobotánicos (Dean, 1995; Williams, 1993).
Lycianthes serie Meizonodontae es un grupo monofilético (Dean, 1995). Incluye 8 especies y 2 variedades: L. acapulcensis (Baill.) D´Arcy, L. ciliolata (M. Martens & Galeotti) Bitter, L. dejecta (Fernald) Bitter, L. hintonii E. Dean, L. moziniana (Dunal) Bitter, L. moziniana var. margaretiana E. Dean, L. moziniana var. oaxacana E. Dean, L. peduncularis (Schltdl.) Bitter, L. rzedowskii E. Dean y L. starbuckii E. Dean (fig. 1). Son hierbas perennes con raíz napiforme, la ramificación del simpodio puede ser monocasial o dicasial. El primer simpodio frecuentemente erecto, con múltiples hojas en arreglo espiral, el siguiente simpodio con hojas geminadas. Flores solitarias, cáliz 10-dentado, corola rotada, fruto ovoide, de color verde o púrpura y semilla obovada o reniforme, comprimida, angular o rómbico-deprimida, de coloración parda. El número cromosómico base es x = 12 (Dean, 1995, 2004). La serie se distribuye desde el norte de México hasta el centro de Costa Rica.
Las relaciones filogenéticas en Lycianthes aún no están resueltas. El análisis filogenético más reciente de la familia incluyó 37 especies de Lycianthes (Särkinen et al., 2013). Se infiere que el género no es un grupo monofilético. Existe una relación de hermandad entre un clado de Lycianthes y otro de Capsicum. A su vez, este clado comparte un ancestro común con otro clado de Lycianthes (Särkinen et al., 2013). Lycianthes serie Meizonodontae se comporta como un grupo monofilético y muestra una posible hermandad con Lycianthes sección Asaropsis (Olmstead, 2013; Särkinen et al., 2013). Con base en evidencia morfológica, Dean (1995) reconoció 2 grupos dentro de la serie. En el grupo ciliolata están incluidos L. ciliolata, L. acapulcensis, L. rzedowskii y L. starbuckii. Por otro lado, el grupo moziniana está formado por L. dejecta, L. moziniana y L. peduncularis. El análisis filogenético de Särkinen et al. (2013) también recuperó 2 clados pero con diferente composición taxonómica. El primero formado por L. acapulcensis y L. ciliolata y su grupo hermano que incluye a L. dejecta, L. moziniana, L. peduncularis y L. rzedowskii. Lycianthes hintonii y L. starbuckii no fueron consideradas en su estudio. Lycianthes hintonii es morfológicamente similar a L. rzedowskii, mientras que L. starbuckii es cercanamente relacionada a L. moziniana y L. peduncularis (Dean, 2004).
Lycianthes serie Meizonodontae tiene importancia etnobotánica. En México, los frutos de L. ciliolata, L. moziniana y L. peduncularis son consumidos y también se utilizan en ceremonias religiosas. En Chiapas, Michoacán, Oaxaca, Puebla y Tlaxcala se ofrecen a los espíritus durante el día de muertos (Dean, 1995). En Tlaxcala, Williams (1993) documentó los usos de L. moziniana como alimento, medicamento y ofrenda. Por otro lado, los frutos de L. acapulcensis y L. rzedowskii, aunque poco conocidos por su importancia etnobotánica, son consumidos en Guerrero y Michoacán. El consumo humano de los frutos ha favorecido su cultivo y tolerancia en traspatios. Ahora, se sugiere que L. ciliolata y L. moziniana son plantas en proceso de domesticación (Dean, 1995; Williams, 1993).
Los datos resguardados en las colecciones biológicas y la aplicación de nuevas tecnologías de análisis permiten conocer mejor la distribución espacial y temporal de las especies. Lo anterior es relevante para la biogeografía, la macroecología, la conservación de la biodiversidad y los efectos del cambio climático global (D’Amen et al., 2015; Powney e Isaac, 2015). Los sistemas de información geográfica (SIG) manipulan y analizan datos geoespaciales, permitiendo generar información nueva y tomar decisiones con criterios más rigurosos (Lira, 2008). El objetivo del presente trabajo es evaluar la riqueza, distribución y el estado de conservación de Lycianthes serie Meizonodontae. En este contexto, conocer áreas de endemismo y alta riqueza de especies es fundamental para identificar regiones prioritarias para la conservación. Asimismo, analizar la distribución de los grupos naturales es importante para determinar áreas geográficas basales y sugerir hipótesis sobre su especiación. Por último, el estado de conservación es un punto de partida para proponer el manejo y aprovechamiento adecuado de especies con valor para el hombre.

Materiales y métodos
Se utilizaron 711 registros georreferenciados que incluyen 8 especies y 2 variedades de Lycianthes serie Meizonodontae. Los registros fueron obtenidos de 11 colecciones nacionales (ANSM, CICY, CIIDIR, HUAA, IBUG, IEB, INEGI, MEXU, SLPM, UNL y ZEA) (Thiers, 2016). Al mismo tiempo, se consultaron las bases de datos electrónicas del Jardín Botánico de Missouri (http://www.tropicos.org) y del Southwest Environmental Information Network (http//:swbiodiversity.org/seinet/index.php). Finalmente, las publicaciones consultadas incluyen a Dean (1994, 1995 y 2004), Nee (1986) y Sierra-Muñoz et al. (2015). Para corroborar la identidad taxonómica de los ejemplares consultados, se incorporaron aquellos registros que tenían adjunto fotografía o fueron referidos en un tratamiento taxonómico para el género.
Se estimó la distribución geográfica de cada taxón con la ayuda de ArcView GIS 3.3 (ESRI, 1992). Se evaluaron los siguientes parámetros: 1) la distancia máxima entre 2 puntos (MaxD), 2) la extensión del área de ocupación (EOO), es decir, la superficie que incluye el total de los registros y que se calcula mediante un polígono convexo mínimo y 3) el área de ocupación (AOO), es decir, la zona que ocupa un taxón y que se encuentra dentro de la extensión del área de ocupación, considerada el área ideal donde un taxón prospera (UICN, 2012b).
Se cuantificó la riqueza de Lycianthes serie Meizonodontae utilizando 6 criterios: 1) división política, 2) provincias biogeográficas (Morrone, 2001), 3) tipos de vegetación (Olson et al., 2001; Rzedowski, 1978), 4) geográficos (elevación, latitud y longitud), 5) modelos de nicho ecológico, 6) cuadrícula de 83 × 83 km.
Según las recomendaciones de la UICN (2012a), el tamaño de celda para una especie bajo análisis debe ser el 10% de la distancia total de los puntos extremos conocidos. Se diseñó una cuadrícula con 609 celdas, cada celda de 83 × 83 km (6,889 km2) en QGIS 2.2.0 (2014). Para este análisis se calculó la mediana de los valores de MaxD obtenidos para cada especie. Este ejercicio resultó en la selección del tamaño de área en 45 minutos por lado (latitud y longitud), que corresponde aproximadamente a 83 km, que es el valor de la mediana obtenida.
Se modeló la distribución potencial mediante el algoritmo MaxEnt versión 3.3.3k (Phillips et al., 2006). El mínimo de registros para ejecutar modelos de distribución potencial es 3 en especies de distribución restringida (van Proosdij et al., 2015). Por esta razón, se incluyeron solo los taxones con 4 o más registros. Para este análisis, se obtuvieron las 19 capas ambientales y el modelo digital de elevación de BIOCLIM (Hijmans et al., 2005; http://www.worldclim.org). La selección de variables se definió mediante análisis de componentes principales (ACP) en la plataforma R (R Development Core Team, 2015). Las variables fueron recortadas para México y Centroamérica. Se definió el área accesible (distribución conocida) de cada taxón (Soberón y Peterson, 2005). Para acotar estas áreas, se utilizaron las ecorregiones terrestres propuestas por Olson et al. (2001). El 75% de los registros se usó para el entrenamiento del modelo y el 25% restante para validarlo. La importancia de las variables se calculó utilizando el análisis de Jackknife (Baldwin, 2009; Phillips et al., 2006). Se utilizó un umbral de corte particular para cada modelo de distribución, con un error de comisión menor al 10%. Los modelos de presencia-ausencia obtenidos representan el área potencial de distribución de cada taxón. Finalmente, los modelos fueron empalmados en ArcMap GIS 10 (ESRI, 2011) para conocer la riqueza potencial de la serie.
Para los 8 taxones, la posible categoría de riesgo se estimó con base en el criterio de distribución geográfica, según las categorías y criterios de la Lista Roja de UICN (UICN, 2012a). El algoritmo “Conservation Assessment Tools”, incluido como una extensión de ArcView (Moat, 2007) fue empleado para este fin. Los valores de AOO sirvieron para designar la categoría de riesgo, a partir de una cuadrícula de 2 × 2 km (Munguía-Lino et al., 2015; UICN, 2012a; Vargas-Amado et al., 2013; Willis et al., 2003).
Resultados
Lycianthes serie Meizonodontae incluye 10 taxones (8 especies y 2 variedades). En México crecen todos ellos, registrados en 22 de los 32 estados. Lycianthes moziniana y L. dejecta son las especies de más amplia distribución en el país, presentes en 15 y 13 entidades federativas, respectivamente. En contraste, L. hintonii y L. starbuckii se restringen a un estado y a una localidad. Los estados con la mayor riqueza son: Estado de México (7 especies) y Puebla (5). En contraste, en 5 estados solo se registra 1 especie y en los demás estados se registran 2 a 4 taxones (tabla 1).
En Centroamérica, con excepción de Belice y Panamá, solo se registró L. acapulcensis y L. ciliolata. En Guatemala crecen ambas especies, mientras que en los otros países de Centroamérica se documentó únicamente a L. acapulcensis (tabla 1).
Lycianthes serie Meizonodonte se registró en 11 provincias biogeográficas. Las provincias con el mayor número de especies fueron la Cuenca del río Balsas (CB) con 7 y la Faja Volcánica Transmexicana (FVT) con 6. Seguido de estas, se encuentra el Altiplano Mexicano (AM), la Costa Pacífica Mexicana (CPM) y la Sierra Madre del Sur (SMS), todas ellas con 4 taxones. En el Golfo de México (GM) y la Sierra Madre Oriental (SMOr) se registraron 3 taxones. Por otro lado, en las provincias de Chiapas (Chis) y la Sierra Madre Occidental (SMOc) solo crecen 2 taxones. Finalmente, en el Oriente de Centro América (OCA) y el Occidente del Istmo de Panamá (OIP) se registró 1 especie (tabla 1). Lycianthes acapulcensis y L. moziniana prosperan en 8 de las 11 provincias, siendo las especies más frecuentes, seguidas por L. ciliolata que crece en 7 provincias. Lycianthes dejecta y L. peduncularis se registraron en 5 y las otras especies (L. hintonii, L. rzedowskii y L. starbuckii) se restringen a 1 provincia (tabla 1).
A nivel regional, Lycianthes serie Meizonodontae se distribuye en 2 regiones biogeográficas y la Zona de Transición Mexicana (ZTM), aunque su distribución no es homogénea en ellas. La ZTM alberga todas las especies, mientras que en las regiones Neártica y Neotropical se registran 4. En la región Neártica se registran a L. ciliolata, L. dejecta, L. moziniana y L. peduncularis, y en la Neotropical a L. acapulcensis, L. ciliolata, L. moziniana y L. peduncularis (tabla 1).
Lycianthes serie Meizonodontae prospera en 9 tipos de vegetación. En el bosque de pino-encino crecen todas sus especies. Mientras que, en el bosque tropical caducifolio crecen 7 (L. acapulcesis, L. ciliolata, L. dejecta, L. moziniana, L. peduncularis, L. rzedowskii y L. starbuckii). En el matorral xerófilo se encontraron Lycianthes ciliolata, L. dejecta, L. moziniana y L. peduncularis (tabla 1). En el bosque mesófilo crecen L. acapulcensis, L. ciliolata, L. moziniana y L. peduncularis. En seguida se encuentran el bosque espinoso, el pastizal y la vegetación acuática y subacuática con 3 especies. Por último, en el bosque tropical perennifolio solo crecen L. acapulcensis y L. ciliolata y en el bosque tropical subcaducifolio solo se encuentra a L. acapulcensis (tabla 1).
La mayor amplitud ecológica se observa en Lycianthes moziniana, registrada en 7 tipos de vegetación. Le siguen L. dejecta y L. peduncularis en 6 tipos, y Lycianthes acapulcensis y L. ciliolata en 5 tipos diferentes. En 3 prospera L. rzedowskii. Lycianthes hintonii y L. starbuckii se restringen a 1 y 2 tipos de vegetación respectivamente (tabla 1).
Tabla 1
Riqueza y distribución de Lycianthes serie Meizonodontae por división política, provincias biogeográficas y tipos de vegetación. AM, Altiplano Mexicano; CB, Cuenca del Balsas; FVT, Faja Volcánica Transmexicana; SMOc, Sierra Madre Occidental; SMOr, Sierra Madre Oriental; SMS, Sierra Madre del Sur; Chis, Chiapas; CPM, Costa Pacífica Mexicana; OAC, Oriente de América Central; GM, Golfo de México; OIP, Occidente del Istmo de Panamá. ZTM, Zona de Transición Mexicana. BE, bosque espinoso; BMM, bosque mesófilo de montaña; BPQ, bosque de pino-encino; BTC, bosque tropical caducifolio; BTP, bosque tropical perennifolio; BTSC, bosque tropical subcaducifolio; MX, matorral xerófilo; P, pastizal; Vas, vegetación acuática y subacuática.
|
Taxón |
División política |
Región/Provincias biogeográficas |
Tipos de vegetación |
|
L. acapulcensis |
Costa Rica: Alajuela, Guanacaste, Puntarenas y San José; El Salvador: Ahuachapán, Chalatenango, San Vicente, Santa Ana y Sonsonate; Guatemala: Chiquimula y Retalhuleu; Honduras: Francisco Morazán; México: Chiapas, Colima, Guerrero, Jalisco, México, Michoacán, Morelos, Oaxaca y Puebla; Nicaragua: León y Managua |
Neotropical: CB, CPM, GM, OAC y OIP; ZTM: Chis, FVT y SMS |
BMM, BPQ, BTC, BTP y BTSC |
|
L. ciliolata |
Guatemala: Baja Verapaz, Huehuetenango, El Progreso y Quiché; México: Chiapas, Hidalgo, México, Oaxaca, Puebla, San Luis Potosí y Tabasco |
Neártica: AM; Neotropical: CB, CPM y GM; ZTM: Chis, FVT y SMS |
BMM, BPQ, BTC, BTP y MX |
|
L. dejecta |
México: Aguascalientes, Ciudad de México, Durango, Guanajuato, Hidalgo, México, Michoacán, Nuevo León, Puebla, Querétaro, San Luis Potosí, Tlaxcala y Zacatecas |
Neártica: AM; Neotropical: CB; ZTM: FVT, SMOc y SMOr |
BE, BPQ, BTC, MX, P y Vas |
|
L. hintonii |
México: Nuevo León |
ZTM: SMOr |
BPQ |
|
L. moziniana |
México: Aguascalientes, Ciudad de México, Durango, Guanajuato, Hidalgo, Jalisco, México, Michoacán, Nayarit, Nuevo León, Oaxaca, Puebla, Querétaro, San Luis Potosí, Tlaxcala, Veracruz y Zacatecas |
Neártica: AM; Neotropical: CB, CPM y GM; ZTM: FVT, SMOc, SMOr y SMS |
BE, BMM, BPQ, BTC, MX, P y Vas |
|
L. peduncularis |
México: Guanajuato, Hidalgo, México, Oaxaca, Puebla y Querétaro |
Neártica: AM; Neotropical: CB y CPM; ZTM: FVT y SMS |
BE, BMM, BPQ, BTC, MX y P |
|
L. rzedowskii |
México: México, Michoacán y Morelos |
Neotropical: CB; ZTM: FVT |
BPQ, BTC y Vas |
|
L. starbuckii |
México: México |
Neotropical: CB |
BPQ y BTC |
El mayor número de registros analizados (361) se ubican en el bosque de pino-encino. El bosque tropical caducifolio y el matorral xerófilo siguen en importancia al contener 136 y 114 registros, respectivamente. En contraste, el bosque tropical subcaducifolio y el bosque espinoso registraron 5 y 4 registros cada uno.
En un gradiente latitudinal, el mayor número de especies (7) fue encontrado a los 18° de latitud N. Sin embargo, el número de registros fue mayor entre los 19 y 20° N, donde se localizaron 6 y 5 especies, respectivamente (fig. 2A). Longitudinalmente, la riqueza máxima (7 especies) se documentó entre los 99 y 100° de longitud O. Así mismo, el número máximo de registros se encontró en esta última longitud (fig. 2B). Por último, se observó una amplitud elevacional, que abarca de los 0 a los 3,200 m. Todos los taxones crecen en un intervalo de 1,500 a 2,000 m snm (fig. 2C), pero la mayor cantidad de registros se ubicó entre 2,000 y 2,500 m snm. Las especies con la distribución altitudinal y geográfica más restringida son L. hintonii y L. starbuckii. En contraste, L. ciliolata, L. acapulcensis y L. moziniana poseen los intervalos más amplios (tabla 2).
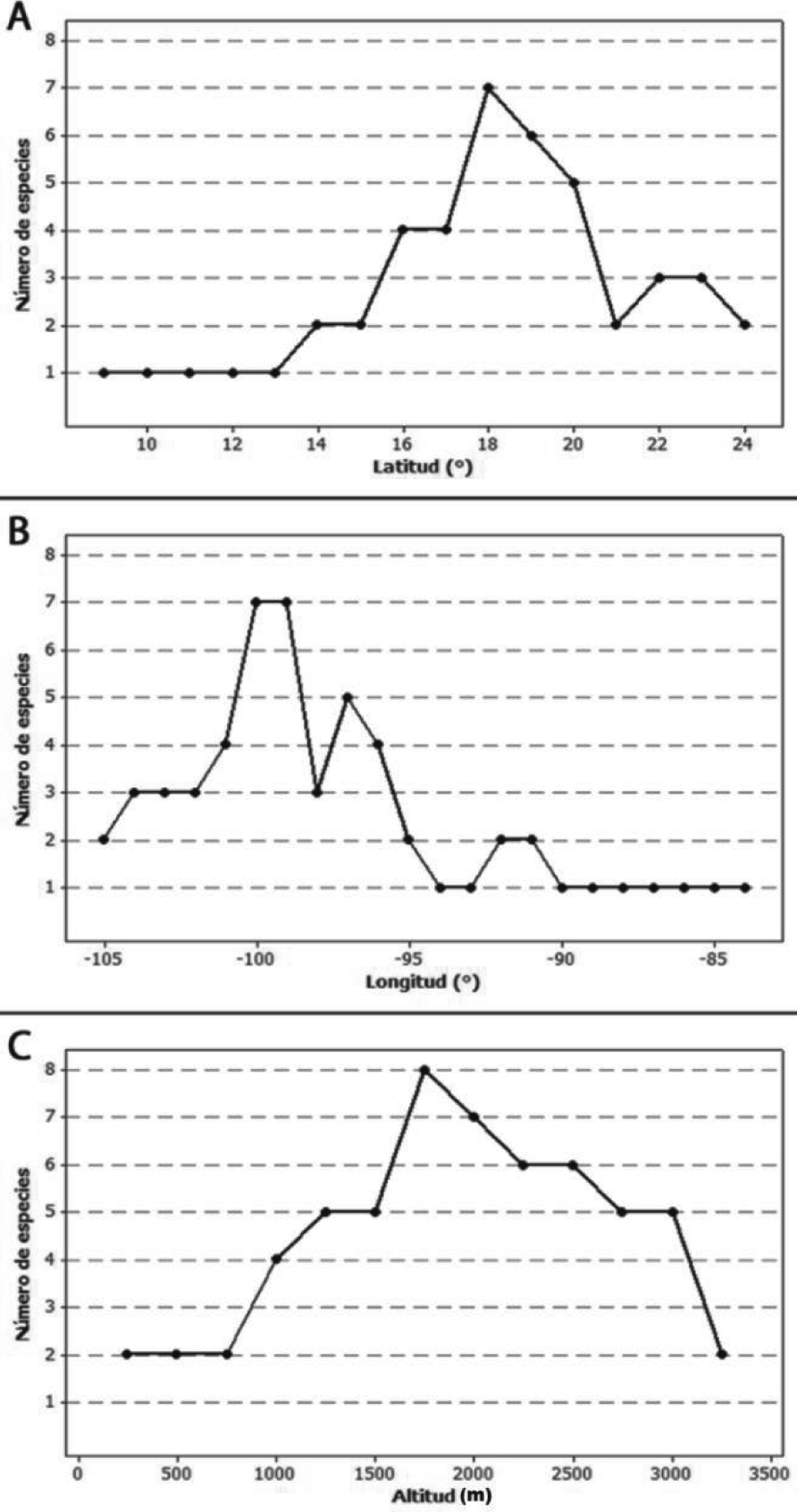
Tabla 2
Distribución de Lycianthes serie Meizonodontae por altitud, latitud y longitud.
|
Taxón |
Elevación |
Latitud |
Longitud |
|
L. acapulcensis |
0-2,900 |
9-20 |
105-84 |
|
L. ciliolata |
0-3,100 |
14-22 |
100-90 |
|
L. dejecta |
1,700-3,000 |
18-24 |
104-97 |
|
L. hintonii |
1,450-1,550 |
23 |
99 |
|
L. moziniana |
700-3,200 |
16-24 |
105-95 |
|
L. peduncularis |
800-2,800 |
16-20 |
103-95 |
|
L. rzedowskii |
1,100-2,700 |
18-19 |
101-99 |
|
L. starbuckii |
1,600-1,900 |
18 |
100 |
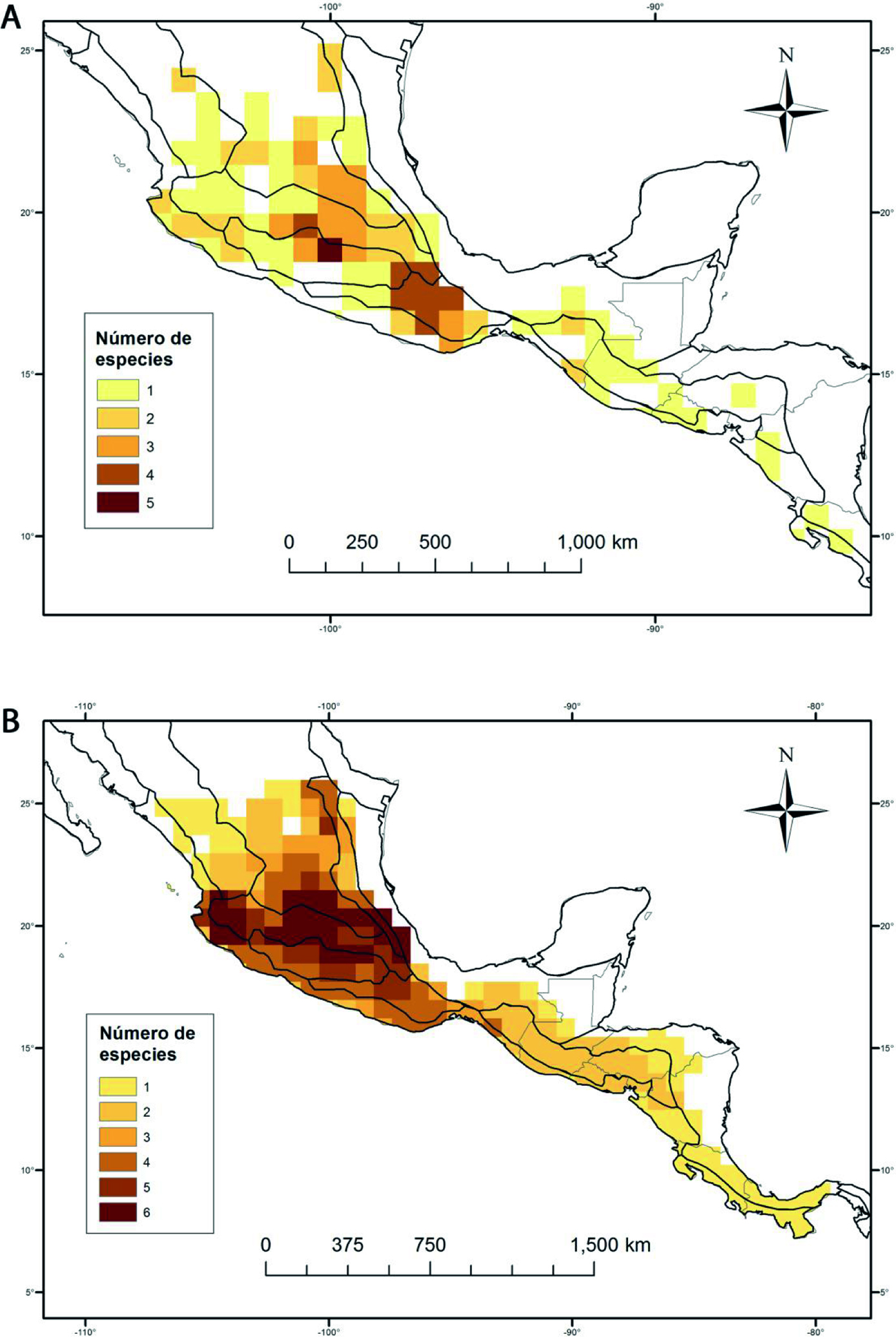
El análisis de Lycianthes serie Meizonodontae empleando celdas de 83 × 83 km, permitió identificar áreas de alta riqueza. El área de estudio se dividió en 609 celdas, de las cuales solo 114 registraron presencia de los taxones estudiados (18.7% de la superficie analizada). El número de especies por celda varía de 1 a 5. Solo 1 celda contuvo 5 especies, en tanto que 74 celdas registraron solamente 1 especie. La celda más rica se ubica al suroeste del Estado de México, en sus límites con Michoacán y Guerrero e incluye 23 municipios del Estado de México (Almoloya de Alquisiras, Amanalco, Amatepec, Calimaya, Coatepec Harinas, Ixtapan de la Sal, Ixtapan del Oro, Otzoloapan, San Simón de Guerrero, Santo Tomás, Sultepec, Tejupilco, Temascaltepec, Tenango de Valle, Texcaltitlán, Tlatlaya, Toluca, Tonatico, Valle de Bravo, Villa Guerrero, Zacazonapan, Zacualpan y Zinacantepec), 8 de Guerrero (General Canuto A. Neri, Ixcateopan de Cuauhtémoc, Pedro Ascencio Alquisiras, Pilcaya, Taxco de Alarcón, Tetipac y Tlalchapa) y 1 de Michoacán (Susupuato). En esta zona también convergen la FVT y la CB (fig. 3A). En esta celda más rica se encuentran L. acapulcensis, L. dejecta, L. moziniana, L. rzedowskii y L. starbuckii, que representan 62.5% de la diversidad de la serie (fig. 3A). Seis de las 8 especies que componen la serie se registraron para menos de 30 celdas. Lycianthes moziniana y L. acapulcensis son las mejor representadas, con 56 y 51 celdas, respectivamente. A diferencia de estas, las especies presentes en el menor número de celdas fueron, L. starbuckii en 2 y L. hintonii en 1 celda.
Los 3 primeros componentes del ACP explicaron el 85% de la variación total observada en las variables utilizadas. De dichos componentes se eligieron las 5 variables con los valores absolutos más altos. En total, 15 variables se utilizaron para modelar la distribución (tabla 3). Los modelos se evaluaron mediante los valores de área bajo la curva (AUC), considerando buenos modelos aquellos valores AUC superiores a 0.83 (tabla 4). La prueba de Jackknife mostró que la estacionalidad de la temperatura (Bio4) y la precipitación anual (Bio12) contribuyeron con más frecuencia en los modelos (tabla 5). Los modelos de distribución potencial permitieron predecir la riqueza, a partir de las preferencias ambientales de los taxones. De esta manera, el número de taxones varió de 1 a 6 por celda (fig. 3B).
Lycianthes serie Meizonodonte se distribuye desde el noreste de México, en el estado de Nuevo León, hasta el centro de Costa Rica. Se encuentran desde las regiones semiáridas de la SMOr y SMOc, desplazándose por el AM, la FVT, la SMS y la CPM. Lycianthes dejecta tiene la distribución más septentrional. En contraste, el límite más meridional del grupo lo mostraron L. acapulcensis y L. ciliolata. Así mismo, estas 2 últimas, poseen la mayor distancia entre 2 puntos. En contraste, L. hintonii y L. starbuckii registran los intervalos de distancia más pequeños (tabla 6, fig. 4).
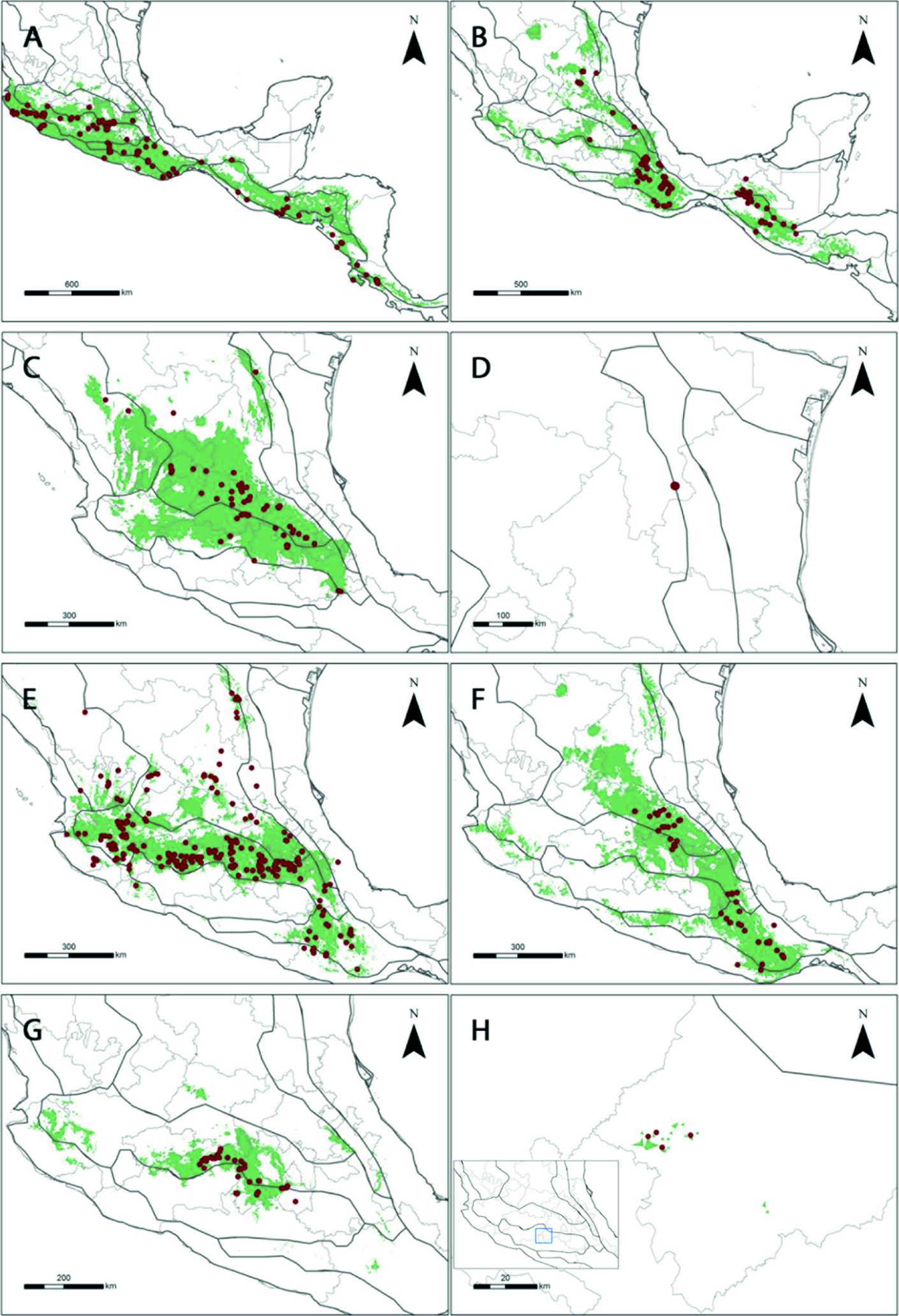
Tabla 3
Variables climáticas seleccionadas mediante el análisis de componentes principales (ACP) para ser usados en los modelos de distribución potencial.
|
Variables climáticas |
Valor absoluto |
Componente |
Varianza acumulada |
|
Temperatura mínima promedio del periodo más frío (Bio6) |
0.288 |
ACP1 |
52 % |
|
Precipitación anual (Bio12) |
0.281 |
||
|
Oscilación anual de la temperatura (Bio7) |
0.276 |
||
|
Temperatura promedio del cuatrimestre más frío (Bio11) |
0.275 |
||
|
Oscilación diurna de la temperatura (Bio2) |
0.265 |
||
|
Temperatura máxima promedio del periodo más cálido (Bio5) |
0.438 |
ACP2 |
75 % |
|
Temperatura promedio del cuatrimestre más cálido (Bio10) |
0.430 |
||
|
Temperatura promedio del cuatrimestre más lluvioso (Bio8) |
0.374 |
||
|
Elevación |
0.341 |
||
|
Temperatura promedio anual (Bio1) |
0.291 |
||
|
Estacionalidad de la precipitación (Bio15) |
0.454 |
ACP3 |
85 % |
|
Precipitación del cuatrimestre más seco (Bio17) |
0.410 |
||
|
Precipitación del periodo más seco (Bio14) |
0.408 |
||
|
Isotermalidad (Bio3) |
0.363 |
||
|
Estacionalidad de la temperatura (Bio4) |
0.318 |
En general, las estimaciones de EOO y AOO mostraron que la distribución de las especies de Lycianthes serie Meizonodontae es amplia. Los valores de EOO se mantuvieron por arriba de los 20,000 km2 en L. acapulcensis, L. ciliolata, L. dejecta, L. moziniana y L. peduncularis, más de 5,000 km2 en L. rzedowskii y por debajo de los 100 km2 en L. starbuckii. Lycianthes hintonii no pudo ser evaluado por falta de registros. Por otra parte, con excepción de L. hintonii y L. starbuckii, las demás especies obtuvieron valores de AOO altos, todos mayores a 2,000 km2. Lycianthes hintonii y L. starbuckii registraron valores por debajo de los 10 km2 (tabla 6).
De acuerdo con el criterio B2 (AOO) y su uso para definir las categorías de riesgo, Lycianthes hintonii y L. starbuckii están en peligro crítico de extinción (CR) y 6 especies se catalogan en preocupación menor (LC) (tabla 6).
Tabla 4
Resumen de los atributos de los modelos de distribución potencial de Lycianthes serie Meizonodontae. MTP, Minimum training presence; BTO, balance training omission; FCV5, fixed cumulative value 5; MTSPS, maximum test sensitivity plus specificity; EETOD, equate entropy of threshold and original distribution.
|
Taxón |
AUC |
Umbral de corte |
Porcentaje de omisión |
|
|
Entrenamiento |
Prueba |
|||
|
L. acapulcensis |
0.832 |
MTP |
0 |
0 |
|
L. ciliolata |
0.952 |
BTO |
1.3 |
15.4 |
|
L. dejecta |
0.917 |
FCV5 |
6.4 |
6.7 |
|
L. moziniana |
0.940 |
MTSPS |
9.5 |
7.2 |
|
L. peduncularis |
0.942 |
EETOD |
2.3 |
7.1 |
|
L. rzedowskii |
0.967 |
MTSPS |
2.9 |
0 |
|
L. starbuckii |
1 |
— |
— |
— |
Tabla 5
Variables climáticas informativas en la generación de los modelos de distribución potencial con base en la prueba Jackknife de Maxent. Las celdas sombreadas indican las variables que en conjunto contribuyeron con 85% o más en los modelos. La X indica la variable que obtuvo la mayor contribución por taxón.
|
Taxón |
Altitud |
Bio1 |
Bio2 |
Bio3 |
Bio4 |
Bio5 |
Bio6 |
Bio7 |
Bio8 |
Bio10 |
Bio11 |
Bio12 |
Bio14 |
Bio15 |
Bio17 |
|
L. acapulcensis |
X |
||||||||||||||
|
L. ciliolata |
X |
||||||||||||||
|
L. dejecta |
X |
||||||||||||||
|
L. moziniana |
X |
||||||||||||||
|
L. peduncularis |
X |
||||||||||||||
|
L. rzedowskii |
X |
||||||||||||||
|
L. starbuckii |
X |
Discusión
México y Centroamérica son un importante centro de diversificación de Solanaceae (tabla 7). Prueba de ello es la alta diversidad y endemismo de Brunfelsia L., Cestrum L., Datura L., Lycianthes Dunal (Hassl.), Nectouxia Kunth, Physalis L., Plowmania Hunz. & Subils, Solanum L., Tzeltalia E. Estrada & M. Martínez y Witheringia L. Hér. (Barboza et al., 2016; Gentry y D’Arcy, 1986; Hijmans y Spooner, 2001; Knapp et al., 2005; Martínez et al., 2017). En México crecen 387 especies agrupadas en 33 géneros, de las cuales 133 son endémicas al país (34%). La riqueza se concentra en los estados de Oaxaca, Chiapas, Veracruz y Jalisco (Cuevas-Arias et al., 2008; Martínez et al., 2017).
En México, Lycianthes muestra un patrón similar a Solanaceae. Treinta y nueve especies (20% del total del género) crecen en su territorio y 14 son endémicas del país (35% del total). Los estados más ricos en especies son Oaxaca, Chiapas y Veracruz con 23, 22 y 14, respectivamente. Sin embargo, Lycianthes serie Meizonodontae muestra un patrón diferente, pues sus 8 especies crecen en México, 6 de ellas endémicas (75%). Estado de México y Puebla poseen el mayor número de especies de la serie, con 7 y 5 cada uno. En contraste, Oaxaca y Chiapas acumulan 4 y 2 especies, respectivamente. En Veracruz solo se encuentra a L. moziniana mientras que L. acapulcensis, L. dejecta y L. moziniana se reportan en Jalisco.
Biogeográficamente, México fue dividido en 2 regiones (Neártica y Neotropical), la Zona de Transición Mexicana y 14 provincias (Morrone, 2001, 2010). Aunque Lycianthes es un elemento neotropical, Lycianthes serie Meizonodontae diversificó en México colonizando todo su territorio con la excepción de las penínsulas de Baja California y de Yucatán. Las provincias biogeográficas con mayor número de especies son CB (7) y la FVT (6); ambas provincias contienen el 88% de la diversidad de la serie. Lycianthes hintonii crece en la SMOr y es endémica de Nuevo León. Por su parte, L. starbuckii es endémica de la sierra de Nanchititla, en el Estado de México, en la parte alta de la CB.
Tabla 6
Resumen de la distribución geográfica de Lycianthes serie Meizonodontae. El tamaño de celda es de 83 × 83 km. El asterisco indica a las especies endémicas de México. NR, número de registros; NC, número de celdas; MaxD (km), máxima distancia entre 2 puntos más distantes; EOO (km2), extensión del área estimada de la presencia de un taxón; AOO (km2), área de ocupación de un taxón; ADP (km2), área de distribución potencial; CR, categoría de riesgo; LC, preocupación menor; CR, en peligro crítico de extinción.
|
Taxón |
NR |
NC |
MaxD |
EOO |
AOO |
ADP |
CR |
|
L. acapulcensis |
120 |
51 |
255.81 |
689,191.49 |
1’308,764.9 |
456,103.09 |
LC |
|
L. ciliolata |
111 |
28 |
137 |
356,250 |
281,543 |
235,219.44 |
LC |
|
L. dejecta* |
67 |
23 |
98.67 |
263,375 |
175,259 |
216,900.58 |
LC |
|
L. hintonii* |
2 |
1 |
0.15 |
0 |
0.04 |
— |
CR |
|
L. moziniana* |
296 |
56 |
122.54 |
513,957 |
435,447 |
169,051.07 |
LC |
|
L. peduncularis* |
61 |
15 |
68.7 |
86,884.60 |
66,077 |
163,626.58 |
LC |
|
L. rzedowskii* |
50 |
7 |
24.84 |
13,622.40 |
11,104 |
33,546.06 |
LC |
|
L. starbuckii* |
4 |
2 |
1.28 |
30.98 |
6.54 |
23.58 |
CR |
Con base en el número de especies y ejemplares de herbario analizados, Lycianthes serie Meizonodontae prospera preferentemente en un intervalo altitudinal entre 1,500 y 2,500 m. La cota de 1,500 m en gran parte de México representa el límite superior de la distribución del bosque tropical caducifolio y el límite inferior del bosque de pino-encino. La vegetación predominante de la FVT es el bosque de pino-encino. En contraste, en la CB predomina el bosque tropical caducifolio. La serie está bien representada en estas dos comunidades vegetales y en las mencionadas provincias (tabla 1). A pesar de esto, el alto número de registros y especies presentes en el bosque de pino-encino, sugiere que la serie prefiere los climas templados.
La formación de la FVT inició en el Mioceno temprano y finalizó en el Plioceno tardío (Ferrari et al., 2012). Durante este proceso se generaron fluctuaciones climáticas y de topografía que propiciaron la aparición de nuevos nichos y con ello la diversificación biológica. Los modelos de distribución potencial confirman que las condiciones ambientales idóneas para que se desarrolle la serie, se localizan a lo largo de la FVT y en las partes altas de la CB, donde dominan el bosque de pino-encino y el bosque tropical caducifolio (fig. 3B).
El análisis Jackknife indicó que ninguna de las 15 variables utilizadas para la modelación mostró una importante contribución para todas las especies. En cambio, cada modelo tuvo una combinación particular de variables. Sin embargo, la estacionalidad de la temperatura (Bio4) y la precipitación anual (Bio12) explicaron el 85% de la variación de los modelos (tabla 5). Lycianthes serie Meizonodontae agrupa a hierbas perennes que durante el periodo de estiaje la parte aérea muere y durante el periodo de lluvia emerge, se desarrolla, florece y fructifica. La estacionalidad de la temperatura explicaría su definido ciclo de vida y su adaptación a la variación estacional. La precipitación anual sugiere la necesidad de la época lluviosa para completar su ciclo.
La riqueza potencial predijo 24 celdas con un máximo de 6 especies, mientras que la riqueza conocida reportó solamente 1 celda con un máximo de 5 especies (fig. 3). Esta última, ubicada al sur del Estado de México, se conservó como una de las celdas de mayor riqueza predichas. Las celdas de alta riqueza potencial se ubicaron a lo largo de la FVT, en sitios divididos en 2 áreas. La primera, con 5 celdas, se localizó al sur y centro de Jalisco. Otros grupos que presentan un patrón similar son Quercus sección Lobatae Loudon (Torres-Miranda et al., 2011), Cosmos Cav. (Vargas-Amado et al., 2013), la subtribu Physalinae de Solanaceae (Miers) Hunz. y Physalis L. (Zamora-Tavares, 2016). La segunda área, con 19 celdas, se localiza en el centro de México, en el Estado de México, Guanajuato, Hidalgo, Michoacán, Morelos, Puebla, Querétaro, Tlaxcala, Veracruz y Ciudad de México. También, se ha documentado en esta zona una alta riqueza de especies de Solanum sección Petota Dumort. (Hijmans y Spooner, 2001) y de la tribu Tigridieae B. M. Kittel (Munguía-Lino et al., 2015).
Con base en sus características geológicas, la FVT fue dividida en 3 subregiones, este, centro y oeste (Ferrari et al., 2012). Por otro lado, a diferentes escalas se proponen 2, 4 y 9 regiones climáticas. (Suárez-Mota et al., 2014). La propuesta de regionalización dentro de la FVT fue evaluada con mamíferos (Escalante et al., 2007) y angiospermas (Suárez-Mota et al., 2013) obteniendo patrones similares. Las áreas potenciales de riqueza para Lycianthes serie Meizonodontae muestran resultados similares (fig. 3B). El centro y el oeste de la FVT fueron divididos por la región del Bajío. En otras palabras, la presencia del bosque tropical caducifolio del Bajío limita la ocurrencia de la serie. A pesar de que, en general, la serie está bien representada en el bosque tropical caducifolio, su baja ocurrencia quizá se vea influenciada por la alta actividad ganadera y agrícola de la región. Sus especies necesitan de espacios abiertos para prosperar y la vegetación dominada por árboles de baja altura en esta región evita su establecimiento. También es importante destacar que en esta parte del país son comunes los malpaíses con escasa vegetación.
La SMOr, en el estado de Nuevo León, contiene 1 celda con 5 taxones (L. ciliolata, L. dejecta, L. hintonii, L. moziniana y L. peduncularis) (fig. 3B). Cuatro de estas especies son endémicas de México. Se han reconocido 61 especies endémicas en el matorral rosetófilo del noreste en Nuevo León, por lo que se considera como un área importante de endemismo de la flora de México (Sosa y De Nova, 2012). También, allí se ha identificado la mayor diversidad florística y heterogeneidad climática de la SMOr (Suárez-Mota et al., 2017). Por otro lado, L. hintonii crece en el bosque de pino-encino mientras que las otras especies prosperan tanto en el bosque de pino-encino como en el matorral xerófilo; esto sugiere que la adaptación a condiciones xerófilas ha propiciado, en parte, la diversificación de la serie. Un fenómeno similar presenta la subtribu Physalinae (Zamora-Tavares, 2016).
Lycianthes acapulcensis mostró el área de distribución más amplia, Lyciantahes moziniana ocupó un segundo lugar. Estas 2 especies también obtuvieron los valores más altos de AOO. Sin embargo, L. moziniana se registró para una mayor cantidad de celdas, lo que la convierte en la especie más abundante (tabla 6). En contraste, L. hintonii y L. starbuckii son las especies más raras (tabla 6), debido a su restringida distribución y su escaso número de registros. Por último, aunque los valores de AOO y EOO sugieren que la distribución de la serie es amplia, en 65% del área estudiada solo se registró 1 especie. Ninguna de las especies se encontró en más de la mitad de las celdas analizadas. El reducido número de celdas con alta riqueza y el alto porcentaje de celdas con 1 taxón posicionan a Lycianthes serie Meizonodontae como un grupo de distribución restringida.
Los patrones de distribución por latitud y longitud muestran que la riqueza incrementa de sur a norte y confirman a México como el principal centro de diversificación de la serie. Aunque Lycianthes serie Meizonodontae crece desde los 0 hasta 3,200 m snm, el intervalo altitudinal favorable para sus especies es entre 1,500 y 2,000 m. En este intervalo se encuentran todas sus especies, el cual coincide con las escalas propuestas por Rzedowski (1978) para el bosque de encino, bosque de pino y matorral xerófilo, la FVT (Ferrusquia-Villafranca, 2007) y la ZTM (Morrone, 2010).
Lycianthes ciliolata tuvo una distribución geográfica discontinua (fig. 4B). Sus poblaciones fueron divididas por el Istmo de Tehuantepec. El modelo de distribución potencial confirma esta discontinuidad. La delimitación taxonómica de L. ciliolata es compleja, algo común en especies de reciente especiación. Estudios filogeográficos posteriores confirmarían la identidad taxonómica de las poblaciones disyuntas, así como su posible origen. Lycianthes peduncularis representa un caso similar; su distribución se observa al norte y sur de la FVT (fig. 4F). Sin embargo, el modelo de distribución potencial sugiere que esta disyunción es por falta de registros, por lo que es posible documentar en el futuro su presencia en la FVT.
Existen otros 2 casos contrastantes, uno es Lycianthes rzedowskii, endémica del Estado de México, Michoacán y Morelos, donde su distribución potencial sugiere que el sur de Jalisco y pequeñas áreas de Guanajuato, Guerrero, Hidalgo, Oaxaca, Puebla, Querétaro y Veracruz poseen las condiciones ambientales para que ésta prospere (fig. 4G). Por otro lado, la distribución potencial de L. starbuckii señala que sus condiciones idóneas solo se encuentran en la sierra Nanchititla, de donde es endémica (fig. 4H). La evaluación de los modelos es complicada, por ello, se requiere dar seguimiento a estas predicciones con trabajo de campo.
La distribución de las especies y su filogenia pueden sugerir hipótesis sobre su especiación. Lycianthes acapulcensis y L. ciliolata tienen una relación de grupos hermanos (Särkinen et al., 2013). La primera se distribuye desde el occidente de México hasta Costa Rica a lo largo de la CPM, FVT y SMS. Habita en la ecotonía entre el bosque de pino-encino y el bosque tropical caducifolio (fig. 4A). En contraste, L. ciliolata habita el bosque de pino-encino y el matorral xerófilo a lo largo del AM, Chis, FVT y la SMOr hasta Guatemala (fig. 4B). Por otro lado, sus intervalos altitudinales son muy similares. Lycianthes acapulcensis habita desde los 0 hasta los 2,900 m, mientras que L. ciliolata crece de los 0 hasta los 3,100 m (tabla 2). Ambas viven en simpatría en el sur de México y Guatemala. Sus patrones de distribución y su relación de grupos hermanos, apoyan la hipótesis del conservadurismo de nicho. Vargas-Amado (2013) identificó que la especiación simpátrica es el mecanismo más común en Cosmos sección Mesinenia Sherff. Así mismo, discutió las similitudes ecológicas entre taxones hermanos e indicó que algunos integrantes de la sección han iniciado un proceso de diversificación ecológica.
El clado hermano de L. acapulcensis más L. ciliolata está compuesto por L. moziniana, L. rzedowskii, L. dejecta y L. peduncularis (Särkinen et al., 2013). A su vez, L. dejecta y L. peduncularis son especies hermanas. Ambas crecen sobre suelos calcáreos en el matorral xerófilo, pero L. dejecta se encuentra hacia el norte de la FVT, mientras que L. peduncularis al sur del mismo sistema (fig. 4C, F). Podría especularse que el ancestro común de ambas tuvo una distribución geográfica más amplia y continua y que la formación de la FVT separó sus poblaciones. Sin embargo, se identificó a L. peduncularis al norte de la FVT y a L. dejecta al sur de la misma provincia. Esto apoyaría la hipótesis del conservadurismo de nicho. El género Dyscritothamnus B. L. Rob. (Asteraceae) tiene una distribución similar a L. dejecta y L. peduncularis. Además, esas 2 especies poseen alta similitud entre sus nichos y se sugiere que apoyan el conservadurismo de nicho, por lo que podrían estar iniciando su proceso de diversificación (Suárez-Mota et al., 2015).
Frecuentemente, la información necesaria para evaluar el riesgo de extinción de una especie es escasa. La mejor información disponible proviene de los ejemplares de herbario. Los valores de AOO y EOO son útiles para evaluar la categoría de riesgo, según el criterio B de la Lista Roja (UICN, 2012a; Maes et al., 2015). Plectranthus L’Hér. (Willis et al., 2003) y Polianthes L. (Feria-Arroyo et al., 2010) han sido evaluados por este criterio. Por su área de distribución restringida y valor de AOO, Lycianthes hintonii y L. starbuckii se deben catalogar en peligro crítico de extinción (CR) (tabla 6). Con el mismo criterio, L. acapulcensis, L. ciliolata, L. dejecta, L. moziniana, L. peduncularis y L. rzedowskii deben ser catalogadas en preocupación menor (LC) (tabla 6). Estas últimas habitan el bosque de pino-encino y matorral xerófilo, comunidades altamente amenazadas por el constante aprovechamiento de sus recursos y el excesivo cambio de uso de suelo (Challenger, 2003; Rzedowski, 1978). Por último, el uso excesivo de herbicidas podría afectar seriamente a las poblaciones de L. acapulcensis, L. ciliolata, L. dejecta, L. moziniana y L. peduncularis, especies que también crecen cerca de campos agrícolas.
El monitoreo constante de sitios previamente analizados y el uso de bases de datos, permitirían obtener información necesaria para evaluar la pertinencia de una especie a una categoría de amenaza. Por ejemplo, L. moziniana fue reportada por última vez para el matorral xerófilo de la Reserva Ecológica del Pedregal de San Ángel en 1951.
Tabla 7
Diversidad y endemismo de Solanaceae en México y Centroamérica (Knapp et al., 2006; Martínez et al., 2017).
|
País |
Especies |
Endémicas |
|
Belice |
74 |
9 |
|
Costa Rica |
162 |
41 |
|
Guatemala |
182 |
46 |
|
El Salvador |
80 |
5 |
|
Honduras |
125 |
12 |
|
México |
387 |
133 |
|
Nicaragua |
122 |
19 |
|
Panamá |
161 |
43 |
Sin embargo, los estudios florísticos posteriores omiten su presencia (Cano-Santana et al., 2008; Castillo-Agüero et al., 2004; González-Hidalgo et al., 2001; Rzedowski, 1954). Esto evidencia el constante riesgo de las comunidades antes mencionadas y la necesidad de programas eficientes de monitoreo para las ANP.
Con excepción de Lycianthes dejecta, los frutos de las otras especies son consumidos y comercializados localmente, lo que hace necesario complementar la evaluación del peligro de extinción por los criterios A, C, D y E de la Lista Roja. La recolecta, comercialización y consumo de los frutos de Lycianthes serie Meizonodontae puede favorecer o actuar en contra de su conservación. Por un lado, su utilidad como alimento fomenta su cuidado. Por otro, la excesiva recolección de los frutos disminuye su capacidad de propagación. Los modelos de nicho ecológico podrían ayudar a identificar áreas idóneas para su cultivo y aprovechamiento adecuado.
Finalmente, emplear los SIG y los algoritmos de modelación, en conjunto con las colecciones biológicas, ofrece un amplio panorama sobre el estado actual de la biodiversidad. México es el centro de diversificación de Lycianthes serie Meizonodontae. En él, se identificaron 3 áreas de alta riqueza, el oeste y centro de la FVT y el noroeste de Nuevo León en la SMOr. Todas ellas coinciden con la presencia del bosque de pino-encino, hábitat idóneo para que se desarrolle la serie. Esta información es fundamental para la toma de decisiones en el manejo y conservación de Lycianthes serie Meizonodontae y los grupos con patrones similares. Con excepción de L. hintonii y L. starbuckii, el riesgo de extinción de la serie es bajo, de acuerdo con el criterio de distribución geográfico analizado en el presente estudio. Sin embargo, su uso alimenticio y el uso potencial de L. moziniana como nuevo cultivo, hace necesario reevaluar el estado de conservación aquí presentado. Obtener información ecológica de las poblaciones silvestres ayudaría a conocer su estado real de amenaza. Por otro lado, programas de manejo y mejoramiento propiciarían el aprovechamiento adecuado de L. moziniana en su área de distribución potencial.
Agradecimientos
Al personal de los herbarios ANSM, CICY, CIIDIR, HUAA, IBUG, IEB, INEGI, MEXU, SLPM, UNL y ZEA por las facilidades brindadas durante la consulta de sus colecciones.
Referencias
Aubriot, X., Singh, P. y Knapp, S. (2016). Tropical Asian species show that the Old World clade of “spiny solanums” (Solanum subgenus Leptostemonum pro parte: Solanaceae) is not monophyletic. Botanical Journal of the Linnean Society, 181, 199–223.
Baldwin, R. A. (2009). Use of maximum entropy modeling in wildlife research. Entropy, 11, 854–866.
Barboza, G. E., Hunziker, A. T., Bernardello, G., Cocucci, A. A., Moscone, A. E., Carrizo-García, C. et al. (2016). Solanaceae. En J. W. Kadereit y V. Bittrich (Eds.), Flowering plants. Eudicots (pp. 295–358). Switzerland: Springer International publishing.
Bitter, G. (1919). Die Gattung Lycianthes. Abhandlungen Naturwissenschaftlichn Vereine Zu Bremen, 24, 292–520.
Bohs, L. y Olmsteand, R. G. (1997). Phylogenetic relationships in Solanum (Solanaceae) based on ndhF sequences. Systematic Botany, 22, 5–17.
Cano-Santana, Z., Castillo-Argüero, S., Martínez-Orea, Y. y Juárez-Orozco, S. (2008). Análisis de la riqueza vegetal y el valor de la conservación de tres áreas incorporadas a la Reserva Ecológica del Pedregal de San Ángel, Distrito Federal (México). Boletín de la Sociedad Botánica de México, 82, 1–14.
Castillo-Argüero, S., Montes-Cartas, G., Romero-Romero, M. A., Martínez-Orea, Y., Guadarrama-Chávez, P., Sánchez-Gallen, I. et al. (2004). Dinámica y conservación de la flora del matorral xerófilo de la Reserva Ecológica del Pedregal de San Ángel, Distrito Federal (México). Boletín de la Sociedad Botánica de México, 74, 51–75.
Cuevas-Arias, C. T., Vargas, O. y Rodríguez, A. (2008). Solanaceae diversity in the state of Jalisco, Mexico. Revista Mexicana de Biodiversidad, 79, 67–79.
Challenger, A. (2003). Conceptos generales acerca de los ecosistemas templados de montaña de México y su estado de conservación. En O. Sánchez, E. Vega, E. Peters y O. Monroy-Vilchis (Eds.), Conservación de ecosistemas templados de montaña en México. Diplomado de conservación, manejo y aprovechamiento de vida silvestre (pp. 17–44). México D.F.: Instituto Nacional de Ecología/ Semarnat.
D’Amen, M., Dubis, A., Fernandes, R. F., Pottier, J., Pellissier, L. y Guisan, A. (2015). Using species richness and functional traits predictions to constrain assemblages predictions from stacked species distribution models. Journal of Biogeography, 42, 1255–1266.
D’Arcy, W. G. (1986). The calyx in Lycianthes and some other genera. Annals of Missouri Botanical Garden, 73, 117–127.
D’Arcy, W. G. (1991). The Solanaceae since 1976, with a review of its biogeography. En J. G. Hawkes, R. N. Lester, M. Nee y N. Estrada (Eds.), Solanaceae III: taxonomy, chemistry and evolution (pp. 75–137). London: The Royal Botanical Garden/ Kew & Linnean Society.
Dean, E. (1994). Lycianthes starbuckii and Lycianthes rzedowskii (Solanaceae), two new species of perennial herbs from México. Novon, 4, 324–329.
Dean, E. (1995). Systematics and ethnobotany of Lycianthes series Meizonodontae (Tesis doctoral). Berkeley: University of California.
Dean, E. (2004). A taxonomic revision of Lycianthes series Meizonodontae (Solanaceae). Botanical Journal of the Linnean Society, 145, 385–424.
Escalante, T., Rodríguez, G., Gámez, N., León-Paniagua, L., Barrera, O. y Sánchez-Cordero, V. (2007). Biogeografía y conservación de los mamíferos. En J. J. Morrone y D. Espinosa (Eds.), Biodiversidad de la Faja Volcánica Transmexicana (pp. 485–502). México D.F.: Universidad Nacional Autónoma de México.
ESRI (Environmental Systems Research Institute). (1992). ArcView 3.3. Environmental Systems Research Institute, Redlands, CA.
ESRI (Environmental Systems Research Institute). (2011). ArcGIS Desktop: Release 10. Environmental Systems Research Institute, Redlands, CA.
Feria-Arroyo, T. P., Solano, E. y García-Mendoza, A. (2010). Reevaluación del riesgo de extinción de cinco especies del género Polianthes L. (Agavaceae). Acta Botanica Mexicana, 92, 11–28.
Ferrari, L., Orozco-Esquivel, T., Manea, V. y Manea, M. (2012). The dynamic history of the Trans-Mexican Volcanic Belt and the Mexico subduction zone. Tectonophysics, 522–523, 122–149.
Ferrusquía-Villafranca, I. (2007). Ensayo sobre la caracterización y significación biológica. En I. Luna, J. J. Morrone y D. Espinosa (Eds.), Biodiversidad de la Faja Volcánica Transmexicana (pp. 7–23). México D.F.: Universidad Autónoma Nacional de México.
Gentry, J. L. y D’Arcy, W. G. (1986). Solanaceae of Mesoamérica. En W. G. D’Arcy (Ed.), Solanaceae biology and systematics (pp. 15–26). New York: Columbia University Press.
González-Hidalgo, B., Orozco-Segovia, A. y Diego-Pérez, N. (2001). La vegetación de la Reserva Ecológica Lomas de Seminario, Ajusco, México. Boletín de la Sociedad Botánica de México, 69, 77–99.
Hassler, E. (1917). Solanaceae, Austro-Americanae. Annuaire Conservatoire et Jardin Botaniques de Geneve, 20, 173–183.
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G. y Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, 1965–1978.
Hijmans, R. J. y Spooner, D. M. (2001). Geographic distribution of wild potato species. American Journal of Botany, 88, 2101–2112.
Knapp, S., Stafford, M. y Martínez, M. (2006). A checklist of the Solanaceae of Guatemala. En E. B. Cano (Ed.), Biodiversidad de Guatemala, Vol. 1 (pp. 259–282). Guatemala: Universidad del Valle de Guatemala.
Knapp, S., Stafford, M., Sousa-Peña, M. y Martínez, M. (2005). A preliminary names list for the Solanaceae of Mesoamerica. En R. C. Keating, V. C. Hollowell y T. B. Croat (Eds.), A festschrift form William G. D’Arcy the legacy of a taxonomist (pp. 71–116). St. Louis, Missouri: Missouri Botanical Garden Press.
Lira, J. (2008). Sistemas de información geográfica y análisis espaciales: un método combinado para realizar estudios panbiogeográficos. Revista Mexicana de Biodiversidad, 79, 281–284.
Maes, D., Isaac, N. J. B., Harrower, C. A., Collen, B., Van Strien, A. J. y Roy, D. B. (2015). The use of opportunistic data for IUCN Red List assessment. Biological of the Linnean Society, 115, 690–706.
Martínez, M., Vargas-Ponce, O., Rodríguez, A., Chiang, F. y Ocegueda, S. (2017). La familia Solanaceae en México. Botanical Sciences, 95, 131–145.
Missouri Botanical Garden. (2015). Tropicos.org. Saint Louis, Missouri. http://www.tropicos.org
Moat, J. (2007). Conservation assessment tools, extension for ArcView 3.x, version 1.2. GIS Unit. Kew: Royal Botanic Gardens.
Morrone, J. J. (2001). Biogeografía de América Latina y el Caribe, Volumen 3. Manuales y Tesis de la Sociedad Entomológica Aragonesa. Zaragoza.
Morrone, J. J. (2010). Fundamental biogeographic patterns across the Mexican Transition Zone: an evolutionary approach. Ecography, 33, 355–361.
Munguía-Lino, G., Vargas-Amado, G., Vázquez-García, L. M. y Rodríguez, A. (2015). Riqueza y distribución geográfica de la tribu Tigridieae (Iridaceae) en Norteamérica. Revista Mexicana de Biodiversidad, 86, 80–98.
Nee, M. (1986). Solanaceae I. Flora de Veracruz, 49, 1–191.
Olmstead, R. G. (2013). Phylogeny and biogeography in Solanaceae, Verbenaceae and Bignoniaceae: a comparison of continental and intercontinental diversification patterns. Botanical Journal of the Linnean Society, 171, 80–102.
Olmstead, R. G., Bohs, L., Migid, H. A., Santiago-Valentín, E., García, V. F. y Collier, S. M. (2008). A molecular phylogeny of the Solanaceae. Taxon, 4, 1159–1181.
Olmstead, R. G. y Palmer, J. D. (1997). Implications for the phylogeny, classification, and biogeography of Solanum from cpDNA restriction site variation. Systematic Botany, 22, 19–29.
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C. et al. (2001). Terrestrial ecoregions of the world: a new map of life on Earth. BioScience, 51, 933–938.
Phillips, S. J., Anderson, R. P. y Schapire, R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecological Modelling, 190, 231–259.
Powney, G. D. e Issac, N. J. (2015). Beyond maps: a review of the applications of biological records. Biological Journal of the Linnean Society, 115, 532–542.
QGIS. (2014). Quantum GIS development team, version 2.2.0. Recuperado de http://www.qgis.org
R Development Core Team. (2015). R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
Rzedowski, J. (1954). Vegetación del Pedregal de San Ángel (Distrito Federal, México). Anales de la Escuela Nacional de Ciencias Biológicas. Anales de la Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, 8, 59–129.
Rzedowski, J. (1978). Vegetación de México. México D.F.: Limusa.
Särkinen, T., Bohs, L., Olmstead, R. G. y Knapp, S. (2013). A phylogenetic framework for evolutionary study of the nightshades (Solanaceae): a dated 1000-tip tree. BMC Evolutionary Biology, 13, 214.
SEINet. (2015). Southwest Environmental Information Network. http//:swbiodiversity.org/seinet/index.php
Sierra-Muñoz, J. C., Siqueiros-Delgado, M. E., Flores-Ancira, E., Moreno-Rico, O. y Arredondo-Figueroa, J. L. (2015). Riqueza y distribución de la familia Solanaceae en el estado de Aguascalientes, México. Botanical Sciences, 93, 97–117.
Soberón, J. y Peterson, A. T. (2005). Interpretation of models of fundamental ecological niches and species distributional area. Biodiversity Informatics, 2, 1–10.
Sosa, V. y De Nova, A. (2012). Endemic angiosperm lineages in Mexico: hotspots for conservation. Acta Botanica Mexicana, 100, 293–315.
Suárez-Mota, M. E., Téllez-Valdés, O., Lira-Saade, R. y Villaseñor, J. L. (2013). Una regionalización de la Faja Volcánica Transmexicana con base en su riqueza florística. Botanical Science, 91, 93–105.
Suárez-Mota, M. E., Téllez-Valdés, O. y Martínez Meyer, E. (2014). Dominios climáticos de las áreas protegidas del Eje Volcánico Transversal de México. GeoFocus, 14, 120–143.
Suárez-Mota, M. E., Villaseñor, J. L. y López-Mata, L. (2015). Ecological niche similarity between congeneric Mexican plant species. Plant Ecology and Evolution, 148, 318–328.
Suárez-Mota, M. E., Villaseñor, J. L. y López-Mata, L. (2017). Dominios climáticos de la Sierra Madre Oriental y su relación con la diversidad florística. Revista Mexicana de Biodiversidad, 88, 224–233.
Thiers, B. (2016). Index herbariorum: a global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. Recuperado en noviembre de http://sweetgum.nybg.org/science/ih/
Torres-Miranda, A., Luna-Vega, I. y Oyama, K. (2011). Conservation biogeography of red oaks (Quercus section Lobatae) in Mexico and Central America. American Journal of Botany, 98, 290–305.
UICN (Unión Internacional para la Conservación de la Naturaleza). (2012a). IUCN Red list categories and criteria: Version 3.1. Gland, Switzerland and Cambridge, UK: IUCN Species Survival Commission, IUCN.
UICN (Unión Internacional para la Conservación de la Naturaleza). (2012b). IUCN guidelines for application of IUCN red list criteria at regional and national levels: Version 4.0. Gland, Switzerland and Cambridge, UK: IUCN Species Survival Commission, IUCN.
van Proosdij, A. S., Sosef, M. S. M., Wieringa, J. J. y Raes, N. (2015). Minimum required number of specimen records to develop accurate species distribution models. Ecography, 38, 001–011.
Vargas-Amado, G. (2013). Biogeografía del género Cosmos (Coreopsideae, Asteraceae) (Tesis doctoral). Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara. Jalisco, México.
Vargas-Amado, G., Castro-Castro, A., Harker, M., Villaseñor, J. L., Ortiz, E. y Rodríguez, A. (2013). Distribución geográfica y riqueza del género Cosmos (Asteraceae: Coreopsidae). Revista Mexicana de Biodiversidad, 84, 536–555.
Villaseñor, J. L. (2016). Checklist of the native vascular plants of Mexico. Revista Mexicana de Biodiversidad, 87, 559–
902.
Wilf, P., Carvalho, M. R., Gandolfo, M. A. y Cúneo, N. R. (2017). Eocene lantern fruits from Gondwanan Patagonia and the early origins of Solanaceae. Science, 355, 71–75.
Williams, D. E. (1993). Lycianthes moziniana (Solanaceae): an underutilized Mexican food plant with “new” crop potential. Economic Botany, 47, 387–400.
Willis, F., Moat, J. y Paton, A. (2003). Defining a role for herbarium data in Red List assessment: a case study of Plectranthus from eastern and southern tropical Africa. Biodiversity and Conservation, 12, 1537–1552.
Zamora-Tavares, M. P. (2016). Relaciones filogéneticas de la subtribu Physalinae (Solanaceae) con énfasis en las especies americanas (Tesis doctoral). Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, México, Jalisco.


