
Sebastián Larrea-Meza a, Pablo Pinto b y Álvaro Zúñiga-Reinoso c, *
a Escuela de Biología Marina, Facultad de Ciencias del Mar y de Recursos Naturales, Universidad de Valparaíso, Avenida Borgoño 16344, Reñaca, Viña del Mar, Chile
b Escuela de Salud, Campus Rodelillo, Universidad de Viña del Mar, Agua Santa 7055, Sector Rodelillo, Viña del Mar, Chile
c Instituto de Zoología, Facultad de Matemáticas y Ciencias Naturales, Universidad de Colonia, Zülpicher Str. 47b, 50674, Colonia, Alemania
*Autor para correspondencia: alzure@gmail.com (A. Zúñiga-Reinoso)
Recibido: 14 enero 2020; aceptado: 13 abril 2020
http://zoobank.org/urn:lsid:zoobank.org:pub:C574BB39-1E92-4D7E-958E-55540C29E0B0
Resumen
Callyntra Solier, 1836 es un género endémico de Chile y Argentina actualmente compuesto por 24 especies descritas. El reciente análisis filogenético de este taxón develó un linaje hermano de C. cantillana, el cual estaba oculto bajo este nombre debido a su sorprendente similitud morfológica. En este trabajo se diagnostica a esta especie nueva realizando un estudio comparativo entre linajes hermanos, utilizando la morfología externa y genitalia de ambos sexos. Adicionalmente, utilizando el gen COI de 7 individuos, se realizó un análisis filogenético para probar la monofilia de la especie. Esta especie nueva es endémica del cordón Cantillana, en la cordillera de la Costa de Chile central. Se discute acerca de su variabilidad morfológica, la cripsis con otras especies y las relaciones filogenéticas dentro de Callyntra.
Palabras clave: Cordón Cantillana; Filogenia; Nycteliini; Pimeliinae; Taxonomía
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
A new cryptic species of Callyntra (Coleoptera: Tenebrionidae) from Central Chile: morphological and genetic evidence
Abstract
Callyntra Solier, 1836 is an endemic genus of Chile and Argentina which contains 24 described species. The last phylogenetic analysis of this group suggested a sister lineage of C. cantillana which was hidden under this name, because of their morphological similarity. In this work we describe this new species making a comparative study between sister lineages, using the external morphology and both sex genitalia. Moreover, using the COI gene from 7 specimens we performed a phylogenetic analysis to test the species monophyly. This new species is endemic to the Cantillana mountains, in the Coastal cordillera of Central Chile. Its morphological variability, the crypsis with other species and the relationship with other Callyntra species are discussed.
Keywords: Cantillana mountain range; Phylogeny; Nycteliini; Pimeliinae; Taxonomy
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
El género Callyntra Solier, 1836 incluye un conjunto de especies endémicas de Chile (Flores y Vidal, 2000a), compartiendo solo una especie con Argentina (Roig-Juñent et al., 2004). La distribución en Chile abarca toda la zona de Chile central, donde las especies habitan en ambas cordilleras, de los Andes y de la Costa, y en terrazas costeras (Flores y Vidal, 2000a, b; Zúñiga-Reinoso y Méndez, 2018). Posterior a la validación y revisión del género, el análisis filogenético basado en morfología daba cuenta de 17 especies para este grupo (Flores, 2000; Flores y Vidal, 2000a, b). Trabajos posteriores han descrito 8 especies nuevas (Aceituno y Guerrero, 2014; Guerrero y Vidal, 2020; Zúñiga-Reinoso, 2016; Zúñiga-Reinoso y Cid-Arcos, 2013, 2015; Zúñiga-Reinoso et al., 2017, 2018), elevando a 24 el total. Sin embargo, el reciente análisis filogenético molecular y de delimitación de especies, sugirió al menos 5 especies crípticas, de las cuales 1 fue descrita recientemente (Zúñiga-Reinoso et al., 2018).
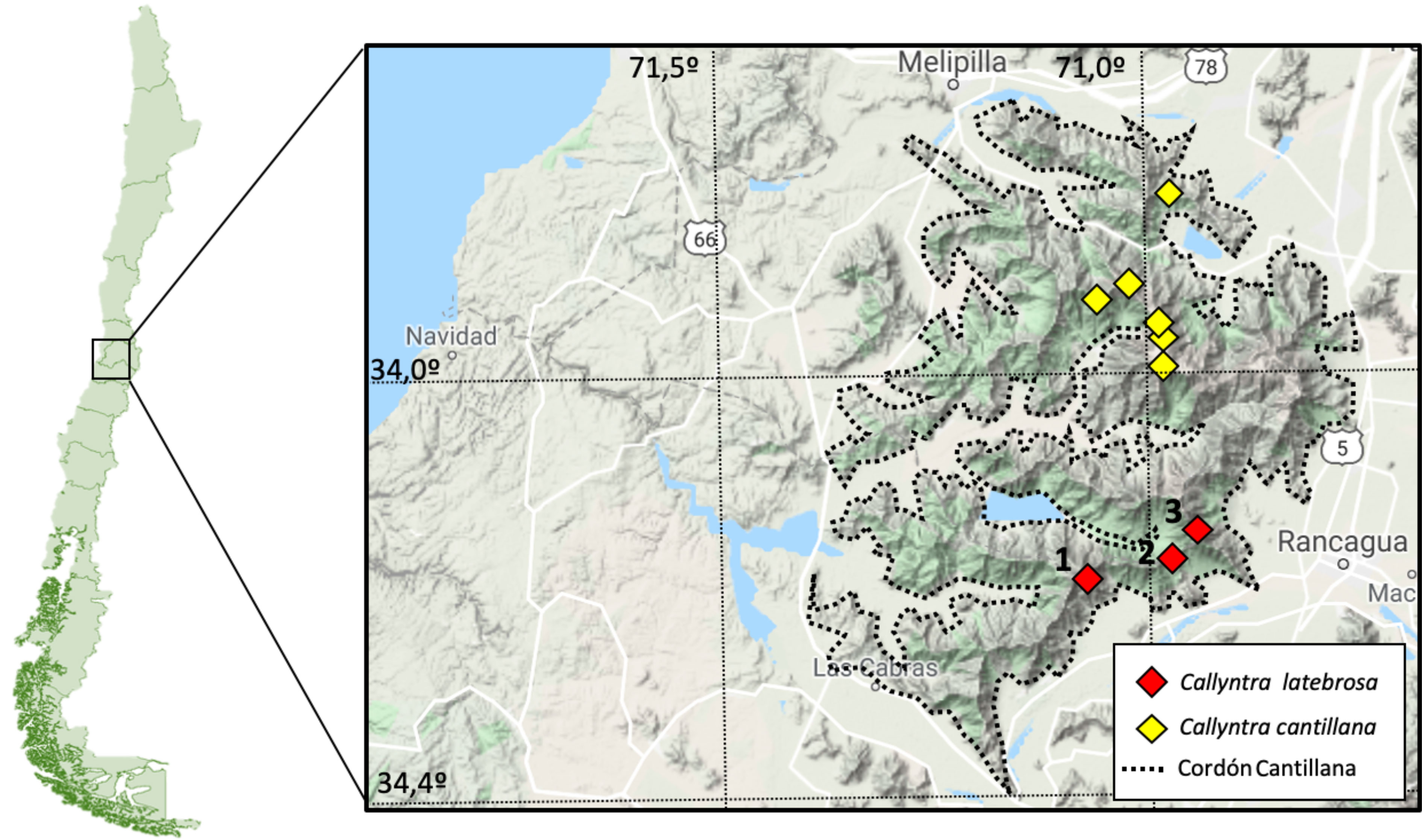
La cripsis en Callyntra es aparentemente un fenómeno común, debido a las respuestas adaptativas a las diferentes altitudes donde habitan las especies (Zúñiga-Reinoso y Méndez, 2018). Un ejemplo de ello es el trabajo de Flores y Vidal (2000b), donde se describe la especie C. cantillana que estuvo oculta bajo C. macrocosta (Guérin-Méneville, 1834) por muchos años debido a la similitud morfológica entre ambas. Posteriormente, Zúñiga-Reinoso y Méndez (2018) validaron filogenéticamente a ambas especies y sugirieron un nuevo linaje que estaba bajo el nombre de C. cantillana. Los individuos de este nuevo linaje provienen del cerro Poqui (en el límite sur del cordón Cantillana, región de O´Higgins; fig. 1), localidad que incluso corresponde a algunos paratipos de C. cantillana (Flores y Vidal, 2000b). Nuestro objetivo fue describir este nuevo linaje, adicionando más individuos en el análisis genético para tener una mayor representación genética y morfológica.
Materiales y métodos
El material fue recolectado de forma manual en los cerros Poqui (1), Chivato (2) y Llivi-Llivi (3) en la región de O´Higgins, Chile, entre los 800-1,500 m snm, todos los cerros pertenecientes al cordón costero de Cantillana (fig. 1). El material recolectado fue conservado en etanol absoluto al 99.8% para la conservación del ADN.
La descripción de la morfología externa y de los edeagos se realizó siguiendo a Flores y Vidal (2000a), mientras que para la descripción del ovopositor se siguió a Zúñiga et al. (2018). El material fue revisado bajo lupa estereoscópica y microscopio electrónico Hitachi SU3500. Para establecer las diferencias entre las especies, se compararon los genitales masculinos y femeninos del nuevo taxón con la especie filogenéticamente más cercana, C. cantillana Vidal y Flores (2000). Los ejemplares fueron reblandecidos en agua caliente por 5 minutos para la extracción de los genitales. La limpieza de los genitales fue efectuada en una solución de KOH a 70 °C durante 10 minutos para los genitalia masculinos y 5 para los femeninos.
Los ejemplares examinados y asignados a la serie tipo se encuentran depositados en las siguientes colecciones: Museo Nacional de Historia Natural, Santiago, Chile (MNNC); Instituto Argentino de Investigaciones de Zonas Áridas, Mendoza, Argentina (IADIZA); Museo de Zoología, Universidad de Concepción, Concepción, Chile (MZUC); Colección Particular Sebastián Larrea, Valparaíso, Chile (CPSL); Colección Particular Pablo Pinto, Valparaíso, Chile (CPPP); Colección Particular Andrés Ramírez, Santiago, Chile (CPAR); Colección Particular Marcelo Guerrero, Santiago, Chile (CPMG); Colección Particular Manuel Diéguez, Santiago, Chile (CPMD); y Colección Particular Mauricio Cid, Talca, Chile (CPMC).
Para el análisis molecular se analizaron fragmentos del gen mitocondrial citocromo oxidasa I (COI) de 7 paratipos de Callyntra sp. n. recolectados en el cerro Poqui. Cuatro de las secuencias fueron descargadas de Genebank (ver códigos en material examinado), las cuales fueron originalmente designadas por Zúñiga-Reinoso y Méndez (2018) como C. cantillana. Para los otros 3 ejemplares, se extrajo el ADN de los músculos torácicos usando el kit EZNA® Insect DNA Kit (Omega Bio-Tek, Inc.). Para el protocolo de amplificación del fragmento del gen COI, se siguió a Zúñiga-Reinoso y Méndez (2018). Las secuencias de la nueva especie se compararon con los fragmentos de COI disponibles en Genbank de las especies filogenéticamente cercanas perteneciente al clado “i” de Callyntra (Zúñiga-Reinoso y Méndez, 2018). Los haplotipos de cada individuo se editaron y alinearon en BIOEDIT versión 7.0.5.3 (Hall, 1999) y posteriormente, se verificaron manualmente en busca de inconsistencias. La distancia genética promedio entre especies relacionadas de Callyntra en estudio se estimó utilizando MEGA 7 (Tamura et al., 2013). Finalmente, se realizó un análisis de inferencia bayesiana utilizando MrBayes v. 3.1.2 (Ronquist y Huelsenbeck, 2003); con 4 corridas independientes, 4 cadenas y comenzando con 1 árbol aleatorio. El análisis se realizó durante 10 millones de generaciones, muestreando cada 1,000 árboles, utilizando a Callyntra tortuosa como grupo externo. El 25% inicial de los árboles resultantes se descartó. Los resultados de las corridas se combinaron para obtener un total de 30,004 árboles. Construimos un árbol de consenso por la regla de mayoría de 50%. La consistencia de los nodos fue determinada por la probabilidad posterior.
Descripción
Callyntra latebrosa Larrea-Meza y Zúñiga-Reinoso, sp. n. (figs. 2, 3)
http://zoobank.org/urn:lsid:zoobank.org:act:90975E70-
D113-4424-9B63-58B123D45A4C
Holotipo macho. Longitud total 20.6 mm. Cuerpo de color negro brillante, patas y antenas de color anaranjado. Cabeza: labro con puntuaciones, escotado, con sedas largas y gruesas en los bordes, que disminuyen en tamaño y grosor hacia la zona media, quedando éste libre de sedas. En el ápice, por la zona ventral, posee gruesas sedas a modo de cepillo. Clípeo con arrugas longitudinales y depresiones laterales, con puntuación y sedas umbilicadas cortas y escasas sobre la superficie. Sutura clipeal no visible. Mejillas levemente convexas, con sedas umbilicadas cortas que apuntan hacia adelante. Margen anterior de los ojos con pocas sedas erectas que apuntan hacia adelante. Frente realzada, con carena central longitudinal que la divide en 2. Puntuación y sedas similares a las del clípeo. Mentón con puntuación gruesa y abundantes sedas largas umbilicadas. Antenas de coloración anaranjada con abundantes sedas en toda su superficie, teniendo en los últimos 3 antenitos abundante pilosidad de color blanco, que les da apariencia aterciopelada. Extensión de las antenas llega hasta 3/4 del pronoto. Tercer antenito más largo que el primero y segundo; antenitos cuarto al sexto de proporciones semejantes; antenitos séptimo y octavo más pequeño que los anteriores y de medida semejante; noveno y décimo antenitos redondeados y más anchos que los precedentes; antenito undécimo terminado en punta. Tórax: pronoto transverso, en declive hacia la zona anterior, presenta pequeñas sedas erectas muy espaciadas. Alcanza su ancho pronotal máximo en el centro de la estructura debido a proyecciones laterales redondeadas que se estrechan suavemente hacia la base del pronoto. Ángulos anteriores redondeados. Margen anterior con puntuación y abundante pilosidad corta que nace bajo este en dirección hacia la cabeza, delimitado completamente por un surco profundo con pequeñas arrugas perpendiculares al margen. Este surco se hace menos profundo hacia la zona media, en donde el margen se ensancha levemente. Margen lateral levantado, rugoso, con sedas cortas que apuntan hacia atrás, siendo más abundantes y largas en la mitad anterior. Ángulos posteriores en punta proyectados hacia atrás quedando encima de los húmeros. Margen posterior levemente rugoso con puntuación poco profunda, escasa y espaciada; con abundante pilosidad corta que nace bajo este y en dirección hacia los élitros. Disco del pronoto con 2 carenas gruesas bien levantadas, y puntuación poco marcada; espaciada y escasa. Marcada rugosidad desordenada entre las carenas. Abundantes arrugas desordenadas entre las carenas que alcanzan hasta el margen lateral (fig. 2c). Proepisterno con estrías longitudinales paralelas poco notorias y escasas sedas umbilicadas. Prosterno convexo, arrugado, con sedas largas y margen anterior levantado, delgado, rugoso y con sedas umbilicadas. Apófisis proesternal ovalada y con reborde grueso arrugado; levantada en la parte inicial y en declive hacia el ápice. Con puntuación leve y sedas umbilicadas largas esparcidas. Mesoventrito con forma de V y con reborde delimitado por surco profundo con arrugas, con el ápice ensanchado y algunas sedas largas. Metaventrito subrugoso, con arrugas longitudinales y con sedas esparcidas. Élitros: rugosos con presencia de abundantes sedas amarillentas cortas, siendo más densas en la mitad posterior de los élitros. Con costilla prominente y gruesa, que toma aspecto almenado desde la mitad posterior hasta el término de la costilla. Espacio entre las costillas principales subcóncavo. Sutura elitral almenada, levantada y levemente ensanchada, alcanzando la mitad de la altura de la costilla principal (fig. 2d). Borde elitral levantado y almenado. Espacio entre la costilla y el borde elitral con rugosidad de apariencia paralela que en el centro se levanta formando una costilla secundaría interrumpida por la rugosidad. Espacio entre la sutura elitral y la costilla principal con rugosidad de apariencia oblicua a desordenada. Pseudopleura subrugosa, glabra los 2/3 anteriores y con puntuación y sedas umbilicadas en el último 1/3. Epipleura subrugosa, glabra los 3/4 anteriores y con pocas sedas cortas el último 1/4. Margen anterior se ensancha hasta tocar los húmeros y los ángulos posteriores del pronoto. Abdomen: esternitos de coloración general negra brillante con estrías longitudinales y presencia de microvellosidades hacía los bordes. Último esternito con abundantes sedas. Patas: todas las patas de color naranja. Coxas arrugadas con puntuación y varias sedas cortas. Trocánter con abundante pilosidad. Fémures con abundante pilosidad de color amarilla en su faz inferior, en el resto de la estructura con sedas más esparcidas que en su faz superior. Tibias con pilosidad corta y gruesa. Tarsos con abundantes sedas. Cinco tarsitos en las patas protorácicas, los 4 primeros de dimensiones semejantes, el quinto más largo, de tamaño 1:3. Cinco tarsitos en las patas mesotorácicas, el primero más largo que los tarsitos segundo, tercero y cuarto, tarsito quinto más largo que el primero. Cuatro tarsitos en patas metatorácicas, primero y cuarto más largos que segundo y tercero. Uñas simples en todas las patas. Genitalia: edeago con estilos laterales del tegmen curvados y subparalelos, estrechándose abruptamente cerca del ápice. Margen proximal ventralmente bisinuado y más ancho en la base. Ventralmente incompleto formando un espacio ovoide poco esclerosado. Mitad apical con una fila de pilosidad corta esparcida y perforaciones. Lámina basal del tegmen con base subredonda más ancha en el tercio apical. Lóbulo medio tubular, de la mitad del ancho de los estilos laterales, ápice redondo y distalmente ensanchado (fig. 3a).
Hembras. En general son muy similares a los machos, pero existe dimorfismo sexual; se presentan entre paréntesis los caracteres de los machos. Élitros anchos y fuertemente convexos (angostos y levemente convexos). Sutura elitral ancha y levantada (sutura elitral angosta). Antenas, fémures, tibias y tarsos más cortos que en los machos. Largo promedio de las hembras 21.8 mm (DE ± 1.24; n = 9, rango 20.3-23.4); en los machos 18.8 mm (DE ± 1.07; n = 7, rango: 17.6-20.6). Genitalia: en vista ventral, los primeros lóbulos de los coxitos (i.e., cl 1) son trapezoides, jorobados en la base y glabros. Más estrechos hacia el margen lateral, siendo este margen bisinuado. Superficie perforada y con hendidura laterobasal. Segundos lóbulos de los coxitos (i.e., cl 2+3+4) semitriangulares, superficie perforada y pilosidad corta muy escasa. Un mechón de pelos cortos y escasos en la base de los gonostilos. Gonostilos fuertemente esclerosados, paralelos y levemente más estrechos en el ápice. Ápice redondeado (fig. 3b). En vista lateral, ambos lóbulos del coxito y gonostilo en ángulo o subrectos. Margen lateral del segundo lóbulo del coxito levantado formando una carena, existiendo una perforación notoria bajo esta carena marginal (fig. 3b). Huevos: presentan coloración general amarillenta, forma elongada y levemente arqueada, 4.5 mm de largo (DE ± 0.27 mm, rango: 4.1-4.9) por 1.8 mm de ancho (DE ± 0.16 mm, rango: 1.4-2.0) (n = 10), polos redondeados, siendo levemente más anchos hacia la base. Capa externa del corion de color blanquecino hialino, de apariencia lisa (fig. 3e).
Variación intraespecífica. Existe gran variación en las arrugas del disco del pronoto y en las arrugas entre las carenas y el margen del pronoto, siendo prácticamente únicas para cada ejemplar. También hubo variación en la pilosidad de los élitros, observándose ejemplares con muy poca pilosidad, de apariencia brillante, y otros con abundante pilosidad, de apariencia opaca. También se observó variación en la abundancia de las sedas del pronoto, del clípeo y labro. Adicionalmente, el largo de los gonostilos parece ser variable en esta especie, apareciendo algunas pocas hembras con gonostilos muy cortos. Finalmente, los individuos de la Reserva Nacional Roblería del Cobre de Loncha, región de O´Higgins, presentan patas más oscuras de tonalidad rojo castaño y gonostilos levemente más largos.
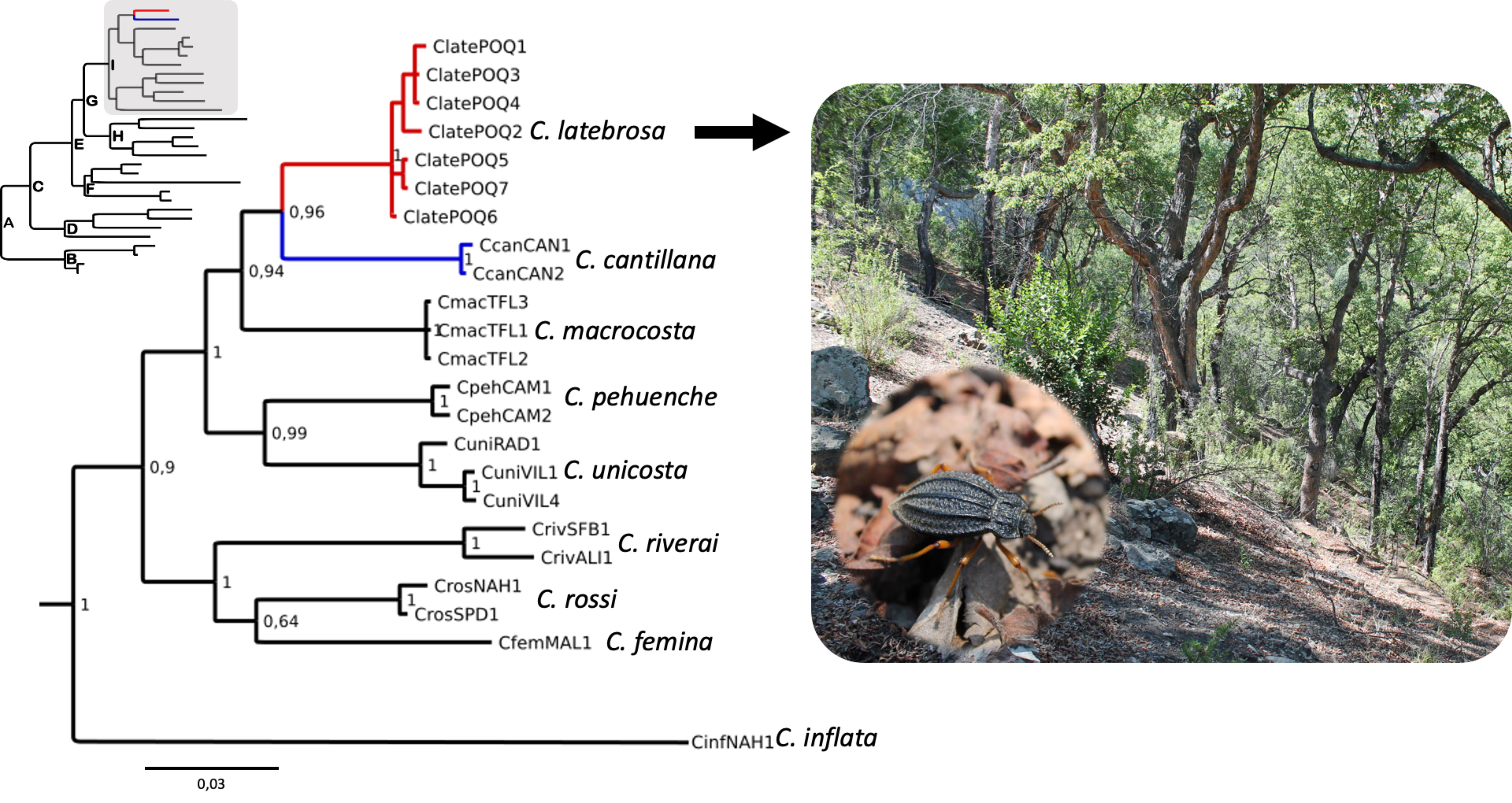
Filogenia y distancia genética. El árbol bayesiano mostró 3 clados bien sustentados, con valores de probabilidad posterior por encima de 0.9 (fig. 4). Callyntra latebrosa resultó ser la especie hermana de C. cantillana, y ambas hermanas de C. macrocosta. Éstas 3, junto con C. unicosta Solier y C. pehuenche Zúñiga-Reinoso et Cid-Arcos, son el clado hermano de C. riverai (Kulzer), C. rossi (Kulzer) y C. femina Zúñiga-Reinoso, Pinto et Collado. La distancia genética interespecífica para el gen COI entre C. latebrosa y el resto de las Callyntra analizadas fue mayor a 4%, pero intraespecíficamente fue menor a 0.5%. Callyntra latebrosa es genéticamente más cercana a C. cantillana con 4.3% de diferenciación.


Resumen taxonómico
Material examinado. Holotipo: Chile, Región de O´Higgins, Provincia de Cachapoal, Coltauco, Cerro Poqui. 01. XI. 2013. col. S. Larrea. 1♂ (MNNC). Paratipos: Chile, Región de O´Higgins, Provincia de Cachapoal, Coltauco, Cerro Poqui. 17. XI. 2012. col. P. Pinto 2♂♂/2♀♀ (1♂ IADIZA, 1♂ MZUC y 2♀ MNNC). 08. XII. 2012. col. R. Barrera 1♂(CPSL). Cerro Poqui, Coltauco VI Reg. 1300 m.s.n.m. 29. XII. 2012. col. A. Ramírez (reverso etiqueta: caminando en bosque de Nothofagus glauca) 1♂/4♀♀ (1♂ CPAR, 1♀ CPAR, 1♀ CPPP, 1♀ CPSL, 1♀ CPMC). Cerro Poqui 03. I. 2013. VI región Chile. Leg: Mauricio Cid 2♂♂ (1♂ en CPMC, 1♂ en MNNC). 01. XI. 2013. col. S. Larrea. 1♀ (CPSL). 27. I. 2013. col. A. Zúñiga. 2♂♂/2♀♀en etanol (2♂♂/2♀♀en MZUC, código Genebank: MH282465, MH260095, MH260096, MH260097). 01. XI. 2013. col. P. Pinto. 1♂/1♀ (1♂ en CPPP y 1♀ en IADIZA). 20. XII. 2015. col. M. Ríos. 2♂♂/2♀♀ (2♂ en CPSL, 1♀ en MNNC y 1♀ en MZUC). 06. XII. 2018. col. D. Mardones. 2♂♂/1♀ en etanol (2♂♂/1♀en MNNC, código Genebank: SUB6834816). 05. X. 2019. col. P. Pinto. 3♂♂/4♀♀ (1♂/1♀ en MNNC, 1♂ en MZUC, 1♂/ 2♀♀ en CPSL, 1♀ en IADIZA). Morro del Chivato, R. N. Roblería del Cobre de Loncha 14. IV. 2013. col. S. Larrea. 2♂♂/1♀ (1♂ en MNNC, 1♂/1♀ en CPSL). R. N. Roblería de Loncha, VI Región 19. IX. 2014. Leg: Mauricio Cid. 1♀ (1♀ en CPMC). Cerro Llivi-Llivi 12. X. 2012. Leg. M. Guerrero 1♂/1♀ (1♂ en CPMG, 1♀ en MNNC) 02. XI. 2002. Leg. A. Pérez 2♂♂/1♀ (1♂/1♀ en CPMG, 1♂en MNNC). Robleria, Llivi-Llivi. 27. XI. 2003 “Robleria”. Leg. C. Lobos 2♂♂/1♀ (1♂/1♀ en CPMD, 1♂en MNNC)
Distribución. Chile, Región de O’Higgins, Provincia de Cachapoal. Corresponde a la región entomofaunística Cordillera de la Costa Central (Peña, 1966) y al área de endemismo homónima (Flores y Vidal, 2000b).
Preferencia de hábitat y biología. Los ejemplares fueron encontrados durante el día en claros al interior de bosques de Nothofagus (fig. 4). Esta especie prefiere sitios semiabiertos con presencia de hojarasca, además es posible encontrarla refugiada bajo piedras y palos en los cuales se oculta en las horas de mayor calor y durante la noche (fig. 4). En laboratorio, las hembras entierran los huevos entre 5 y 20 mm de profundidad, de forma individual o en grupos de hasta 3 huevos, unidos lateralmente y depositados verticalmente. El huevo queda recubierto por una película de arcilla adherida al corion debido a un mucílago producido al momento de la puesta (fig. 3e).
Etimología. El nombre específico proviene del latín latebrosus, que quiere decir que se oculta y no se deja conocer, haciendo referencia a que esta especie es críptica y que fue desapercibida como especie diferente de C. cantillana en la descripción de Flores y Vidal (2000b).
Comentarios taxonómicos
Externamente, C. latebrosa sp. n. es una especie muy similar a C. cantillana pero se diferencia de esta última porque las arrugas del disco del pronoto alcanzan hasta el margen lateral y por presentar la sutura elitral notoriamente más ensanchada, realzada, glabra y con crenulación uniforme que no se mezcla con las arrugas del disco elitral. Los genitalia masculinos difieren levemente siendo la de C. latebrosa sp. n. más recta, mientras que la de C. cantillana aguzada. Los genitalia femeninos de C. latebrosa, c1 es jorobada y hendida, cl 2 + 3 + 4 con escasos pelos cortos, margen del cl 2 + 3 + 4 con carena estrecha, margen dorsal del cl 2 + 3 + 4 no quillado. Mechón de pelos en la base del gonostilo corto y escaso. Mientras que en C. cantillana (fig. 3d) la c1 es plana y pareja, cl 2 + 3 + 4 posee pilosidad abundante, con un margen fuertemente quillado y el mechón de pelos en la base del gonostilo largo y abundante
Discusión
En este estudio proponemos a C. latebrosa sp. n. como una especie nueva basado en morfología externa, los genitalia de machos y hembras, y la evidencia genética. Adicionalmente, a pesar de leves variaciones morfológicas, se establece bajo este nombre el material proveniente de la Reserva Nacional Roblería del Cobre de Loncha (morro Chivato y cerro Llivi-Llivi), el cual no ha sido estudiado genéticamente, pero comparte varios de los caracteres morfológicos diagnósticos más importantes. Con la revisión solo de la morfología externa, fue difícil diferenciar a C. labrosa de C. cantillana y solo 2 caracteres fueron realmente útiles para diferenciarlas: arrugas del disco del pronoto que alcanzan hasta el margen lateral y sutura elitral realzada con crenulación uniforme. La extrema similitud externa de ambas especies podría deberse a la reciente historia de divergencia de los linajes y a la similitud de su hábitat (Struck et al., 2018; Zúñiga-Reinoso y Méndez, 2018). Por lo tanto, es esperable que estas especies hermanas hayan “retenido” rasgos morfológicos conllevando a la cripsis. Sin embargo, la comparación de los genitalia fue decisiva para la diferenciación de ambas especies. Como se describió anteriormente para otros Nycteliini por Zúñiga-Reinoso et al. (2018, 2019), los genitalia femeninos fueron particularmente útiles para diferenciar especies crípticas, incluso más que los masculinos, debido a que estos órganos poseen una estructura más compleja que la de los machos. Por lo tanto, este trabajo vuelve a corroborar la importancia de los genitalia femeninos para trabajos taxonómicos en Nycteliini. Finalmente, el uso de herramientas genéticas ha sido muy útil para descubrir y/o corroborar la hipótesis de especies crípticas de insectos en Chile (Alfaro et al., 2018; Anguita et al., 2017; Zúñiga-Reinoso y Predel 2019; Zúñiga-Reinoso et al., 2018, 2019). Los resultados filogenéticos obtenidos en este estudio refuerzan la hipótesis de Zúñiga-Reinoso y Méndez (2018), que sugería a C. latebrosa como entidad independiente y que se describe, formalmente en este trabajo, con un mayor número de individuos analizados.
Se aborda por segunda vez la morfología de los huevos en Callyntra, siendo los de C. latebrosa sp. n. muy similares a los de C. tortuosa (Zúñiga-Reinoso et al., 2017), pero se pueden diferenciar de esta última por la apariencia lisa del exocorion, por su forma arqueada y por ser de mayor tamaño (1.03×). Esto contrasta con lo expuesto por Pizarro-Araya et al. (2005), que no encontraron diferencias interespecíficas en la estructura coriónica del huevo de 8 especies de Gyriosomus Guérin-Méneville analizados. Probablemente, las diferencias encontradas en la estructura coriónica se deban a una respuesta en función del sustrato donde se depositan los huevos. Las especies de Gyriosomus estudiadas por Pizarro-Araya et al. (2005), comparten hábitat con sustratos similares en la costa, mientras que C. tortuosa y C. latebrosa ocupan hábitats con sustratos muy disímiles, como las terrazas costeras y el bosque de la cordillera de la costa, respectivamente.
El área de distribución de C. latebrosa se encuentra dentro del sitio prioritario de conservación “La Roblería/Cordillera de la Costa Norte y Cocalán”, siendo el cerro Poqui declarado recientemente como Santuario de la Naturaleza (https://www.monumentos.gob.cl/monumentos/santuarios-de-la-naturaleza/cerro-poqui). Este cordón montañoso alberga especies de coleópteros que han sido recientemente redescubiertos, como Apterodorcus tristis y Sclerostomulus nitidus (Lucanidae; Paulsen, 2010) o especies endémicas de este cerro y que han sido descritos recientemente como Callyntra hibrida (Zúñiga-Reinoso y Cid-Arcos, 2013) y Licinoma poqui (Tenebrionidae; Guerrero y Vidal, 2019). Esta nueva especie aumenta la endemicidad de insectos del cordón montañoso costero de Cantillana. Este cordón fue mencionado como una zona importante para la conservación de insectos desde un punto de vista evolutivo (Jerez et al., 2015) y con estos resultados se realza como un importante “hotspot” de insectos de Chile central.
Agradecimientos
A Andrés Ramírez Cuadros y Mauricio Cid por disponer de ejemplares para este estudio. Un especial agradecimiento a Gonzalo Collado (Universidad del Bío Bío) por proveer las imágenes SEM y a Fabián Larrea por proveer las fotografías del hábito y genitalia. Finalmente agradecemos a los revisores anónimos del manuscrito.
Referencias
Aceituno, G. y Guerrero, M. (2014). Nuevas especies de Callyntra (Coleoptera: Tenebrionidae) de Chile central. Revista Chilena de Entomología, 39, 5–11.
Alfaro, F. M., Zúñiga-Reinoso, A., Muñoz-Ramírez, C., Trewick, S. A. y Méndez, M. A. (2018). Phylogeography of Chilean red cricket Cratomelus armatus (Orthoptera, Anostostomatidae) reveals high cryptic diversity in Central Chile. Biological Journal of the Linnean Society, 123, 712–727. https://doi.org/10.1093/biolinnean/bly019.
Anguita-Salinas, S., Barahona-Segovia, R., Poulin, E. y Zúñiga-Reinoso, A. (2017). Genetic and morphological evidence for a new cryptic species of Ectinogonia (Coleoptera: Buprestidae) from central Chile. Zootaxa, 4303, 284–292. https://doi.org/10.11646/zootaxa.4303.2.8
Flores, G. E. (2000). Cladistic analysis of the Neotropical tribe Nycteliini (Coleoptera: Tenebrionidae). Journal of the New York Entomological Society, 108, 13–25. https://doi.org/10.1664/0028-7199(2000)108[0013:CAOTNT]2.0.CO;2
Flores, G. E. y Vidal, P. (2000a). Revalidation and systematic revision of the Chilean genus Callyntra Solier (Coleoptera: Tenebrionidae). Annals of the Entomological Society of America, 93, 1052–1075. https://doi.org/10.1603/0013-8746(2000)093[1052:RASROT]2.0.CO;2
Flores, G. E. y Vidal, P. (2000b). Cladistic analysis of the Chilean genus Callyntra Solier (Coleoptera: Tenebrionidae), with description of a new species. Journal of the New York Entomological Society, 108, 187–204. https://doi.org/10.1664/0028-7199(2000)108[0187:CAOTCG]2.0.CO;2
Guerrero, M. y Vidal, P. (2019). Nueva especie del género Licinoma Pascoe (Tenebrionidae: Adeliini) de Chile central. Revista Chilena de Entomología, 45, 633–638.
Guerrero, M. y Vidal, P. (2020). Nueva especie de Callyntra Solier (Coleoptera: Tenebrionidae) de Chile central. Revista Chilena de Entomología, 46, 113–120.
Hall, T. A. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41, 95–98.
Jerez, V., Zúñiga-Reinoso, A., Muñoz-Escobar, C. y Pizarro-Araya, J. (2015). Acciones y avances sobre la conservación de insectos en Chile, Gayana, 79, 1–3. http://dx.doi.org/10.4067/S0717-65382015000100001
Paulsen, M. J. (2010). The stag beetles of southern South America. Bulletin of The University of Nebraska State Museum, 24, 1–124.
Peña, L. E. (1966). A preliminary attempt to divide Chile into entomofaunal regions, based on the Tenebrionidae (Coleoptera), Postilla, 97, 1–17.
Pizarro-Araya, J., Jerez, V. y Cepeda-Pizarro, J. (2005). Descripción de huevos y larvas de primer estadio del género Gyriosomus Guérin-Méneville, 1834 (Coleoptera, Tenebrionidae, Nycteliini), Gayana, 69, 277–284. http://dx.doi.org/10.4067/S0717-65382005000200008
Roig-Juñent, S., Flores, G. E., Ocampo, F. C. y Smith, A. B. (2004). Nuevas citas de Coleoptera para la Argentina (Carabidae, Lucanidae, Scarabaeidae y Tenebrionidae). Revista de la Sociedad Entomológica Argentina, 63, 45–48.
Ronquist, F. y Huelsenbeck, J. P. (2003). MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics, 19, 1572–1574. https://doi.org/10.1093/bioinformatics/btg180
Struck, T. H., Feder, J. L., Bendiksby, M., Birkeland, S., Cerca, J., Gusarov, V. I. et al. (2018). Finding evolutionary processes hidden in cryptic species. Trends in Ecology and Evolution, 33, 153–163. https://doi.org/10.1016/j.tree.2017.11.007
Tamura, K., Stecher, G., Peterson, D., Filipski, A. y Kumar, S. (2013). MEGA6: molecular evolutionary genetics analysis, version 6.0. Molecular Biology and Evolution, 30, 2725–2729. https://doi.org/10.1093/molbev/mst197
Zúñiga-Reinoso, A. (2016). Callyntra inconsiderata sp. nov (Coleoptera: Tenebrionidae), una nueva especie para la costa de Chile Central, Gayana, 80, 175–180. http://dx.doi.org/10.4067/S0717-65382016000200175
Zúñiga-Reinoso, A. y Cid-Arcos, M. (2013). Callyntra hibrida sp. n.: un nuevo tenebriónido de Chile Central (Coleoptera: Tenebrionidae), Gayana, 77, 132–135. http://dx.doi.org/10.4067/S0717-65382013000200007
Zúñiga-Reinoso, A. y Cid-Arcos, M. (2015). A new species of Callyntra (Coleoptera: Tenebrionidae) from Central Chile. Zootaxa, 4000, 294–298. https://doi.org/10.11646/zootaxa.4000.2.8
Zúñiga-Reinoso, A. y Méndez, M. A. (2018). Hidden and cryptic species reflect parallel and correlated evolution in the phylogeny of the genus Callyntra (Coleoptera: Tenebrionidae) of Central Chile. Molecular Phylogenetic and Evolution, 127, 405-415. https://doi.org/10.1016/j.ympev.2018.04.022
Zúñiga-Reinoso, A., Pinto, P. y Collado, G. (2018). No more machismo in Callyntra (Coleoptera: Tenebrionidae): Callyntra femina, a new species discovered based on female genitalia and genetic evidence. Zootaxa, 4461, 429–437. https://doi.org/10.11646/zootaxa.4464.3.8
Zúñiga-Reinoso, A., Pinto, P. y Larrea, S. (2017). Una nueva especie de Callyntra (Coleoptera: Tenebrionidae) para la costa de Chile Central, Revista Mexicana de Biodiversidad, 88, 307–311. https://doi.org/10.1016/j.rmb.2017.03.024
Zúñiga-Reinoso, A., Pinto, P. y Predel, P. (2019). A new a species of Gyriosomus Guérin-Méneville, 1834 (Coleoptera: Tenebrionidae) from the Chilean Atacama desert. Annales Zoologici, 69, 105–112. https://doi.org/10.3161/00034541ANZ2019.69.1.006
Zúñiga-Reinoso, A. y Predel, P. (2019). Past climatic changes and their effects on the phylogenetic pattern of the Gondwanan relict Maindronia (Insecta: Zygentoma) in the Chilean Atacama Desert. Global and Planetary Change, 182, 103007. https://doi.org/10.1016/j.gloplacha.2019.103007


