Potencial de aprovechamiento de epífitas vasculares caídas en un bosque mesófilo de montaña de Los Tuxtlas, Veracruz, México
Esteban Francisco-Ventura a, Rebeca A. Menchaca-García a, Tarin Toledo-Aceves b y Thorsten Krömer a,*
a Centro de Investigaciones Tropicales, Universidad Veracruzana, José María Morelos Núm. 44 y 46, Zona Centro, 91000 Xalapa, Veracruz, México
b Red de Ecología Funcional, Instituto de Ecología, A.C., Apartado postal 63, 91070 Xalapa, Veracruz, México
*Autor para correspondencia: tkromer@uv.mx (T. Krömer)
Resumen
La abundancia de plantas epífitas vasculares caídas (EVC) por causas naturales en bosques húmedos tropicales indica un alto potencial para su manejo sostenible. Dado que las EVC están destinadas a morir, se pueden aprovechar sin afectar a las poblaciones que se encuentran en el dosel. Sin embargo, existen pocos estudios sobre el suministro de EVC. En este estudio se evaluó la riqueza, abundancia, diversidad y el potencial de uso de EVC en el bosque mesófilo de montaña (BMM) del volcán San Martín Tuxtla, Veracruz, México. Se establecieron 8 parcelas de 100 m2 y se muestrearon mensualmente durante 1 año. Se recolectaron y determinaron todas las especies de EVC, y se registró su estado de salud. En total se registraron 66 especies distribuidas en 34 géneros y 11 familias. Se estimó una alta abundancia de EVC (41,050 ± 1,905 plantas ha/año), de las cuales 72.9% fueron viables para el cultivo. Con base en las abundancias registradas de EVC, proporción de ejemplares viables y usos reportados u observados, se determinó que 5 especies tienen un alto potencial de aprovechamiento regional. Este estudio muestra que existe una gran riqueza y abundancia de EVC, las cuales pueden ser aprovechadas en el BMM de Los Tuxtlas.
Palabras clave:
Araceae; Biodiversidad; Bosque de niebla; Bromeliaceae; Orchidaceae; Productos forestales no maderables; Pteridofitas
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Potential harvesting of fallen vascular epiphytes in a tropical montane cloud forest at Los Tuxtlas, Veracruz, Mexico
Abstract
The abundance of epiphytic plants fallen by natural causes in humid tropical forests suggests their high potential for sustainable harvesting. Since fallen vascular epiphytes (FVE) are destined to die, their collection from the forest floor has no impact on the populations in the canopy. However, few studies have explored the supply of FVE. Here, we evaluated the richness, abundance, diversity and potential use of FVE in a tropical montane cloud forest on the volcano San Martín Tuxtla, Veracruz, Mexico. Eight plots each of 100 m2 were established and sampled monthly for 1 year. All species collected were identified and their health condition recorded. In total, 66 different species were found, belonging to 34 genera and 11 families. A high abundance was estimated (41,050 ± 1,905 plants ha/year), of which 72.9% plants were in a viable condition for cultivation. Based on the recorded abundances of FVE, proportion of viable specimens and reported or observed uses, we determined that 5 species have a high potential for regional use. This study shows that there is a high richness and abundance of FVE, which can be harvested from the cloud forest of Los Tuxtlas.
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Keywords:
Araceae; Biodiversity; Cloud forest; Bromeliaceae; Orchidaceae; Non-timber forest products; Pteridophytes
Introducción
El bosque mesófilo de montaña (BMM) es un ecosistema de alta prioridad para la conservación y el manejo sustentable debido a que alberga una extraordinaria biodiversidad, provee importantes servicios ecosistémicos y enfrenta importantes amenazas antrópicas por cambio de uso de suelo, fragmentación y tala selectiva ilegal (Conabio, 2010; Sánchez-Ramos y Dirzo, 2014; Scatena et al., 2010). Para México, el BMM es la formación vegetal con mayor riqueza de flora en relación al área que ocupa (~ 0.4% del territorio nacional) y Rzedowski (1996) estima que en este ecosistema existen ~ 2,500 especies de plantas de manera exclusiva, que representan aproximadamente el 10% de la flora vascular del país (Gual-Díaz y González-Medrano, 2014; Villaseñor, 2016).
El BMM también destaca por la abundancia y diversidad de epífitas, plantas que desarrollan todo su ciclo de vida en sus árboles hospederos (Gentry y Dodson, 1987; Krömer et al., 2007) y que son un componente importante de la riqueza y biomasa de la comunidad vegetal (Hofstede et al., 1993). Para los BMM de México se reportan más del 60% de las 1,650 especies de epífitas vasculares del país (Espejo-Serna, 2014). Además de la importancia ecológica de las epífitas vasculares por su participación en el ciclo de nutrientes, el mantenimiento de las condiciones microclimáticas del dosel y como hábitat y alimento para la fauna (Benzing, 1990; Hietz, 2010; Zotz y Andrade, 2002), numerosas especies son fuente de productos forestales no maderables (PFNM) (Acebey et al., 2010; Elliott y Ticktin, 2013; Hornung-Leoni, 2011; Krömer et al., 2018). Muchas epífitas se usan localmente en la medicina tradicional, como alimento, en la producción de artesanías o con fines ceremoniales (Acebey et al., 2007; Bennett, 1995, 2000; Hornung-Leoni, 2011). También, algunas especies son importantes en el mercado hortícola internacional (Peck y Christy, 2004); sin embargo, en México el comercio de epífitas silvestres es principalmente ilegal y su extracción se lleva a cabo sin planes de manejo (Cruz-García et al., 2015; Flores-Palacios y Valencia-Díaz, 2007; Toledo-Aceves, García-Franco et al., 2014).
Alrededor del 25% de la riqueza de epífitas en Veracruz está ligada al comercio ilegal, y la mayoría de las especies comercializadas regularmente son extraídas del BMM (Flores-Palacios y Valencia-Díaz, 2007). La extracción de epífitas del BMM sin una planificación que asegure su mantenimiento puede tener efectos negativos sobre sus poblaciones. Análisis demográficos previos reportan tasas de crecimiento poblacional (λ) menores a la unidad en bromelias epífitas aún sin extracción, y una disminución significativa importante en λ aún con niveles de extracción muy bajos (Mondragón y Ticktin, 2011; Toledo-Aceves, García-Franco et al., 2014). Al respecto, se sabe poco sobre el impacto de la recolección repetida de individuos sobre la dinámica de la población (Mondragón, 2009; Mondragón y Villa-Guzmán, 2008; Toledo-Aceves, Hernández-Apolinar et al., 2014). Sin embargo, se ha documentado que el aprovechamiento de las epífitas caídas por causas naturales, con base en un manejo planificado podría proporcionar un incentivo económico para el mantenimiento de la biodiversidad de los bosques (Mondragón y Ticktin, 2011; Toledo-Aceves, García-Franco et al., 2014).
Debido a que una causa importante de mortalidad en las epífitas es su caída de los árboles, pues una vez en el suelo están destinadas a morir (Matelson et al., 1993; Mondragón y Ticktin, 2011), la recolecta de ejemplares caídos para su posterior comercialización no afecta a las poblaciones (Mondragón y Ticktin, 2011, Toledo-Aceves, García-Franco et al., 2014). Las plantas epífitas se desprenden de los troncos y ramas de los árboles debido a la gran biomasa que pueden alcanzar, y a factores naturales como el desprendimiento de la corteza y caída de ramas que pueden estar asociados a diferentes eventos estocásticos como tormentas, huracanes y movimiento de animales (Matelson et al., 1993; Sarmento-Cabral et al., 2015; Winkler et al., 2007). Sin embargo, existe poca información sobre la tasa de caída de las epífitas, su aprovechamiento potencial a partir de ejemplares caídos (Mondragón y Ticktin, 2011; Toledo-Aceves, García-Franco et al., 2014) y sobre la implementación de viveros rústicos para el mantenimiento y la reproducción de epífitas (Damon, 2006; Gómez et al., 2015; Miranda-Jiménez et al., 2007). Con la idea de contribuir al desarrollo de estrategias para el aprovechamiento de PFNM que puedan fungir como incentivos para la conservación del bosque (Alexiades y Shanley, 2004), el objetivo del presente estudio fue determinar el potencial de aprovechamiento de las epífitas vasculares caídas (EVC) en un BMM de la región de Los Tuxtlas, Veracruz, México. Debido a que para conocer la disponibilidad del recurso a aprovechar como PFNM es necesario contar con información sobre la variación en la riqueza y abundancia de ejemplares disponibles por temporada, en este estudio se evaluó la riqueza, la abundancia y la condición de los ejemplares de EVC a lo largo de 1 año.
Materiales y métodos
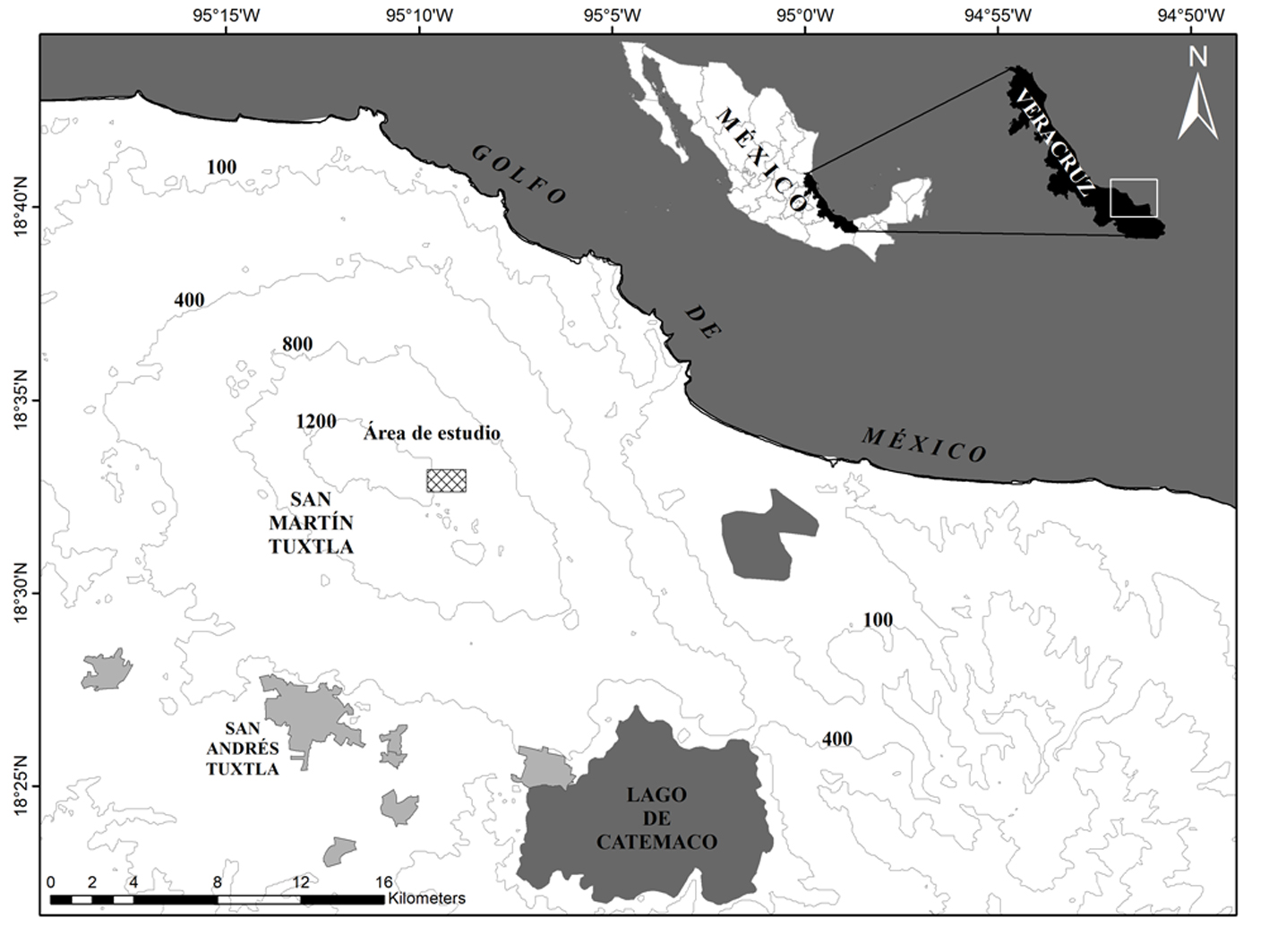
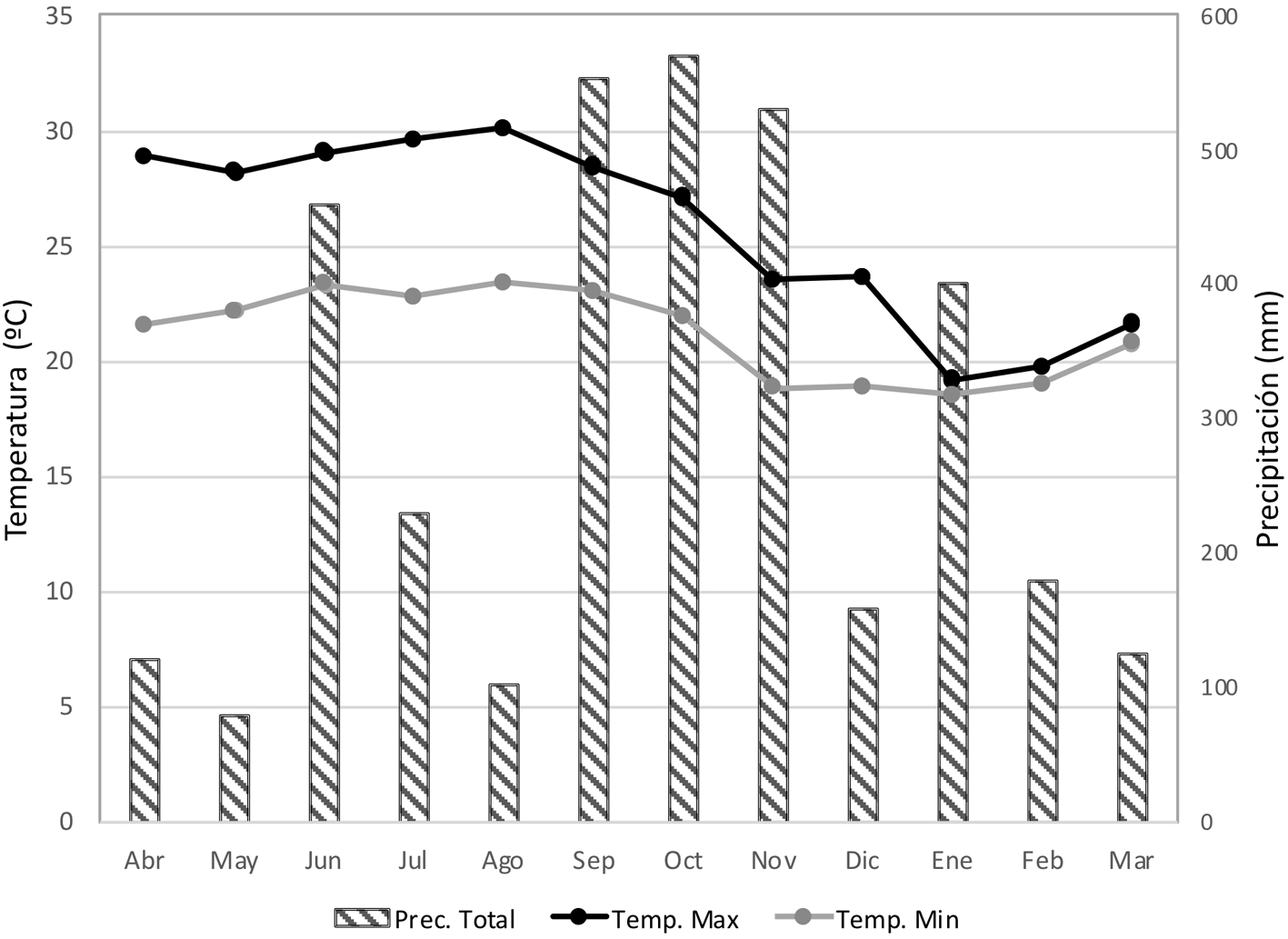
La sierra de Los Tuxtlas emerge sobre la llanura costera del golfo de México, ubicada al sureste del estado de Veracruz, México (Conabio, 2010). Esta región está constituida por 2 grandes macizos volcánicos: el volcán San Martín Tuxtla (1,680 m snm) y la sierra de Santa Marta (1,680 m snm) (Guevara et al., 2004). El estudio se llevó a cabo en el área de BMM conservado de la vertiente sureste del volcán San Martín Tuxtla, Municipio de San Andrés Tuxtla, que pertenece a la comunidad Adolfo Ruiz Cortines (fig. 1). Esta zona tiene un clima templado, con una temperatura media anual de 18 ºC, así como una precipitación media anual de 4,000 mm y por lo menos 40 mm de precipitación durante el mes más seco (Guevara et al., 2004). Los árboles del dosel oscilan entre 20 y 30 m de altura, aunque en algunas ocasiones superan 40 m, e incluyen especies de los géneros Carpinus, Clethra, Engelhardtia, Liquidambar, Quercus y Ulmus. Éstos son hospederos con alta riqueza y abundancia de epífitas vasculares, destacando los helechos y orquídeas (Acebey et al., 2015; Krömer et al., 2013). Debido a que no existe una estación meteorológica cercana a la zona de estudio, se obtuvieron datos meteorológicos (precipitación y temperatura) de abril 2014 a marzo 2015 de la “Estación de Biología Tropical Los Tuxtlas” de la UNAM, ubicada a 140 m snm al pie del volcán San Martín Tuxtla (apéndice 1).
Debido a que las epífitas caen constantemente, se realizaron muestreos mensuales durante un año (abril 2014 a marzo 2015) en un área de BMM entre 1,000 y 1,200 m snm. Se establecieron 8 parcelas de 10 × 10 m (área total = 800 m²) con una distancia mínima de 15 m entre parcelas. En cada parcela se recogieron todas las plantas completas (con hojas, raíz o rizoma) de epífitas vasculares que se encontraron en el suelo (fig. 2). Los fragmentos como hojas, tallos, pseudobulbos sin raíz o frondas de helechos no se tomaron en cuenta con el fin de no sobreestimar la cuantificación de las plantas. Para la familia Araceae solo se recogieron las plantas no enraizadas en el suelo. Posteriormente, se contaron y registraron todas las plantas de EVC.

Todas las plantas se asignaron a una de las siguientes 2 categorías de viabilidad: 1) viables para el cultivo: plantas sin daño o con daño físico parcial (hojas completas a parcialmente completas), coloración de la hoja típica de cada especie y sin manchas (que pueden indicar presencia de enfermedades o plagas) y 2) no viables para el cultivo: plantas muy dañadas, con coloración anómala, marchitas o podridas, con manchas en hojas y con plagas y/o enfermedades.
El potencial de las especies para su aprovechamiento se definió a partir de la combinación de los siguientes criterios: abundancia y proporción de ejemplares en la categoría 1) de viabilidad, así como su uso local observado durante el trabajo de campo en la comunidad Adolfo Ruiz Cortines y usos reportados en la literatura y otras fuentes para la región de Los Tuxtlas y diferentes estados de México (ver referencias en tabla 1). Para que una especie fuera considerada con alto potencial de aprovechamiento debía cumplir con 3 requisitos: alta abundancia, alta proporción de ejemplares viables y con usos reportados. En contraste una especie fue considerada con bajo potencial si presentó una baja proporción de ejemplares viables aun si presento alta abundancia. Las especies poco abundantes, si bien pueden ser atractivas, se consideraron también con menor potencial de aprovechamiento debido a la incertidumbre asociada en su suministro.
La identificación de las plantas vivas y ejemplares herborizados se realizó mediante la consulta de especialistas, así como con el uso de claves taxonómicas e información publicada de las diferentes familias (Araceae: Croat y Acebey, 2015; Bromeliaceae: Espejo-Serna, López-Ferrari y Ramírez-Morillo, 2005; Orchidaceae: Soto-Arenas et al., 2007; Piperaceae: Vergara-Rodríguez, 2009; Pteridophyta: Mickel y Smith, 2004). Los ejemplares de respaldo se depositaron en el herbario del Centro de Investigaciones Tropicales (CITRO) de la Universidad Veracruzana en Xalapa, Veracruz.
Tabla 1
Número de epífitas vasculares caídas (EVC) totales y viables por ha/año (promedio ± 1 desviación estándar) en un bosque mesófilo de montaña de Los Tuxtlas, Veracruz, México. Categorías de uso para la región de Los Tuxtlas y diferentes estados de México (reportado u observado; referencias), y de potencial (Pot.) de aprovechamiento regional (1 = alto, 2 = medio, 3 = bajo). Usos: Or = ornamental, Me = medicinal, Co = comestible, Ce = Cestería, Cr = ceremonial (* venta en mercado local, + venta en mercado internacional).
|
Tabla 1. Continuación. |
|||||||
|
Familia |
Especie |
Núm. de EVC ha/año |
Núm. de EVC viables ha/año |
Categoría de uso |
Estados de México |
Referencias |
Pot. |
|
Familia |
Especie |
Núm. de EVC ha/año |
Núm. de EVC viables ha/año |
Categoría de uso |
Estados de México |
Referencias |
Pot. |
|
Araceae |
Anthurium scandens (Aubl.) Engl. |
375±41.1 |
312.5±37.5 |
Or+, Co |
Puebla |
1,2 |
2 |
|
Monstera acuminata K. Koch |
25±7.2 |
25±4.9 |
Or+, Ce |
Los Tuxtlas, Veracruz; Oaxaca |
3,4,5,6,7 |
3 |
|
|
Monstera egregia Schott |
525±87.8 |
375±60.9 |
Or |
Los Tuxtlas, Veracruz |
8, Obs. pers. |
2 |
|
|
Philodendron tripartitum (Jacq.) Schott |
87.5±11.2 |
62.5±6.4 |
Or+, Co, Me, Ce |
Veracruz, Oaxaca, Puebla |
1,4,9,10,11 |
2 |
|
|
Syngonium sp. |
100±25.2 |
75±18.0 |
Or |
Los Tuxtlas, Veracruz |
Obs. pers. |
3 |
|
|
Bromeliaceae |
Catopsis sessiliflora (Ruiz y Pav.) Mez |
75±14.6 |
75±14.6 |
Or*, Cr |
Los Tuxtlas, Veracruz; Chiapas |
12,13,14, Obs. pers. |
2 |
|
Tillandsia punctulata Schltdl. y Cham. |
3,137.5±190.4 |
2,100±129.1 |
Or*+, Cr |
Los Tuxtlas, Veracruz; Estado de México; Oaxaca |
13,14,15,16,17,18,19,20, Obs. pers. |
1 |
|
|
Tillandsia viridiflora (Beer) Baker |
37.5±5.6 |
25±4.9 |
Or*, Cr |
Los Tuxtlas, Veracruz; Estado de México |
14,17,18,20, Obs. pers. |
3 |
|
|
Werauhia noctiflorens T. Krömer, Espejo, López-Ferr. y Acebey |
75±14.6 |
62.5±11.2 |
– |
– |
– |
3 |
|
|
Werauhia nutans (L.B. Sm.) J.R. Grant |
87.5±14.5 |
37.5±5.6 |
– |
– |
– |
3 |
|
|
Gesneriaceae |
Columnea purpusii Standl. |
50±9.7 |
25±7.2 |
– |
– |
– |
3 |
|
Orchidaceae |
Arpophyllum giganteum Hartw. ex Lindl. |
37.5±5.6 |
37.5±5.7 |
Or*+ |
Los Tuxtlas, Veracruz |
3,14,21,22 |
2 |
|
Coelia macrostachya Lindl. |
112.5±16.9 |
37.5±5.6 |
Or*+, Co |
Los Tuxtlas, Veracruz |
13,14,23, Obs. pers. |
2 |
|
|
Coelia triptera (Sm.) G. Don ex Steud. |
300±42.3 |
150±15.9 |
Or*+ |
Los Tuxtlas, Veracruz |
13,14, Obs. pers. |
1 |
|
|
Dichaea muricatoides Hamer y Garay |
412.5±34.6 |
287.5±28.9 |
Me, Or* |
Los Tuxtlas, Veracruz; Oaxaca; Puebla |
13,14,24,25, Obs. pers. |
2 |
|
|
Dichaea squarrosa Lindl. |
37.5±7.8 |
37.5±7.8 |
Or |
– |
14 |
3 |
|
|
Dichaea sp. 1 |
162±32.2 |
87.5±21.6 |
– |
– |
– |
3 |
|
|
Elleanthus capitatus (Poepp. y Endl.) Rchb. f. syn. Elleanthus cynarocephalus (Rchb. f.) Rchb. f. |
100±18.7 |
62.5±11.2 |
Or* |
Los Tuxtlas, Veracruz |
13,14,21,22 |
2 |
|
|
Epidendrum ramosum Jacq. |
387.5±25.3 |
225±19.6 |
Or* |
Veracruz |
13,14 |
2 |
|
|
Epidendrum veroscriptum Hágsater |
112.5±17.8 |
87.5±13.5 |
Or* |
Veracruz |
13,14,22 |
2 |
|
|
Gongora galeata (Lindl.) Rchb. f. |
87.5±9.9 |
62.5±6.4 |
Or*+ |
Los Tuxtlas, Veracruz; Estado de México |
13,14,17,21,22,26 |
2 |
|
|
Jacquiniella leucomelana (Rchb. f.) Schltr. |
400±20.9 |
312.5±18.1 |
Or* |
Los Tuxtlas, Veracruz |
13,14, Obs. pers. |
2 |
|
|
Jacquiniella teretifolia (Sw.) Britton y P. Wilson |
300±29.7 |
225±20.3 |
Or* |
Veracruz |
13,14 |
2 |
|
|
Maxillaria cucullata Lindl. |
137.5±14.6 |
87.5±12.4 |
Or*+ |
Los Tuxtlas, Veracruz; Estado de México; Puebla |
13,14,17,22,24, Obs. pers. |
2 |
|
|
Maxillaria densa Lindl. |
112.5±15.2 |
50±6.1 |
Or*+ |
Los Tuxtlas, Veracruz |
13,14,21,22,26 |
2 |
|
|
Maxillaria meleagris Lindl. |
175±19.1 |
162.5±13.5 |
Or*+ |
Los Tuxtlas, Veracruz; Oaxaca |
13,14,22,27, Obs. pers. |
1 |
|
|
Maxillaria variabilis Bateman ex Lindl. |
312.5±53.7 |
175±28.6 |
Or*+ |
Los Tuxtlas, Veracruz |
13,14,21,22,28 |
1 |
|
|
Nidema boothii (Lindl.) Schltr. |
175±20.5 |
37.5±5.6 |
Or*+ |
Los Tuxtlas, Veracruz; Oaxaca |
13,14,21,27 |
2 |
|
|
Ornithocephalus inflexus Lindl. |
200±16.3 |
200±16.3 |
Or |
– |
14 |
2 |
|
|
Prosthechea baculus (Rchb. f.) W.E. Higgins |
187.5±27.2 |
75±11.3 |
Or*+ |
Los Tuxtlas, Veracruz |
13,14,21,22 |
2 |
|
|
Prosthechea pseudopygmaea (Finet) W.E. Higgins |
12.5±3.6 |
12.5±3.6 |
Or* |
Veracruz |
13,14 |
3 |
|
|
Pleurothallis antonensis L.O. Williams |
150±13.1 |
125±11.7 |
Or+ |
– |
14 |
2 |
|
|
Pleurothallis cardiothallis Rchb. f. |
325±37.2 |
262.5±31.6 |
Or*+ |
Los Tuxtlas, Veracruz; Puebla |
13,14,24, Obs. pers. |
1 |
|
|
Sobralia sp. 1 |
25±4.9 |
25±4.9 |
Or |
Los Tuxtlas, Veracruz |
Obs. pers. |
3 |
|
|
Stelis aff. bidentata Schltr. |
912.5±64.1 |
650±48.7 |
– |
– |
– |
3 |
|
|
Stelis veracrucensis Solano |
1,900±155.9 |
1,450±127.5 |
Or |
– |
14 |
3 |
|
|
Stelis sp. 3 |
487.5±77.3 |
512±54.7 |
– |
– |
– |
3 |
|
|
Piperaceae |
Peperomia pseudoalpina Trel. |
337.5±40.6 |
200±19.5 |
Me |
Veracruz |
9 |
3 |
|
Peperomia sanjoseana C.DC |
100±15.4 |
37.5±8.4 |
Or |
Los Tuxtlas, Veracruz |
Obs. pers. |
3 |
|
|
Peperomia tenerrima Schltdl. y Cham. |
187.5±37.4 |
150±33.7 |
– |
– |
– |
3 |
|
|
Peperomia sp. |
150 ±26.1 |
112.5±14.2 |
– |
– |
– |
3 |
|
|
Pteridophyta |
|||||||
|
Aspleniaceae |
Asplenium auriculatum Sw. |
4,050±332.6 |
3,075±244.4 |
Or |
Los Tuxtlas, Veracruz |
14, Obs. pers. |
2 |
|
Asplenium cuspidatum Lam. |
1,837.5±169.8 |
1,450±126.9 |
Or |
Los Tuxtlas, Veracruz |
14, Obs. pers. |
2 |
|
|
Asplenium miradorense Liebm. |
137.5±17.2 |
75±14.6 |
– |
– |
– |
3 |
|
|
Hymenophyllaceae |
Hymenophyllum polyanthos (Sw.) Sw. |
4,175±252.9 |
3,137.5±198.5 |
– |
– |
– |
3 |
|
Hymenophyllum sp. 2 |
200±37.1 |
200±37.1 |
– |
– |
– |
3 |
|
|
Dryopteridaceae |
Elaphoglossum erinaceum (Fée) T. Moore |
100±11.1 |
50±6.1 |
Or |
Los Tuxtlas, Veracruz |
14, Obs. pers. |
2 |
|
Elaphoglossum peltatum (Sw.) Urb. |
4,537.5±228.70 |
3,850±181.9 |
Or+ |
Los Tuxtlas, Veracruz |
29,30, Obs. pers. |
2 |
|
|
Elaphoglossum petiolatum (Sw.) Urb. |
1,437.5±97.0 |
1,050±93.4 |
Or |
– |
14 |
2 |
|
|
Elaphoglossum sartorii (Liebm.) Mickel |
1,537.5±105.7 |
1,100±83.3 |
Or |
Los Tuxtlas, Veracruz |
Obs. pers. |
2 |
|
|
Elaphoglossum vestitum (Schltdl. y Cham.) T. |
500±44.1 |
212.5±16.4 |
Or |
Los Tuxtlas, Veracruz |
Obs. pers. |
2 |
|
|
Lycopodiaceae |
Phlegmariurus pithyoides (Schltdl. y Cham.) B. Øllg. |
25±7.2 |
25±4.9 |
Or* |
Veracruz |
31 |
3 |
|
Phlegmariurus taxifolium (Sw.) Á. Löve y D. Löve |
50±6.1 |
12.5±3.6 |
Or* |
Los Tuxtlas, Veracruz |
31, Obs. pers. |
3 |
|
|
Polypodiaceae |
Campyloneurum angustifolium (Sw.) Fée |
25±4.9 |
25±4.9 |
Me, Or+ |
14, 32 |
3 |
|
|
Campyloneurum xalapense Fée |
412.5±62.4 |
200±24.6 |
Or |
– |
14 |
2 |
|
|
Melpomene xiphopteroides (Liebm.) A.R. Sm. y R.C. Moran |
87.5±9.9 |
62.5±8.4 |
– |
– |
– |
3 |
|
|
Pecluma consimilis (Mett.) M.G. Price |
462.5±59.7 |
137.5±19.5 |
Or |
Los Tuxtlas, Veracruz |
Obs. pers. |
3 |
|
|
Pecluma sursumcurrens (Copel.) M.G. Price |
4,500±513.9 |
3,475±407.6 |
Or |
Los Tuxtlas, Veracruz |
Obs. pers. |
2 |
|
|
Pleopeltis angusta Humb. y Bonpl. Ex Willd. |
750±76.7 |
587.5±73.7 |
– |
– |
– |
3 |
|
|
Pleopeltis astrolepis (Liebm.) E. Fourn. |
187.5±36.2 |
125±23.1 |
– |
– |
– |
3 |
|
|
Pleopeltis crassinervata (Fée) T. Moore |
1,312.5±74.3 |
875±62.3 |
– |
– |
– |
3 |
|
|
Polypodium collinsii Maxon |
325±30.5 |
162.5±15.5 |
Or |
Los Tuxtlas, Veracruz |
Obs. pers. |
2 |
|
|
Polypodium longepinnulatum E. Fourn. |
325±31.4 |
225±18.8 |
Or |
– |
14 |
2 |
|
|
Polypodium plebeium Schltdl. y Cham |
912.5±63.3 |
500±30.3 |
Me, Or |
– |
14,33 |
2 |
|
|
Pteridaceae |
Scoliosorus ensiformis (Hook.) T. Moore |
187.5±19.3 |
150±16.8 |
Or |
Los Tuxtlas, Veracruz |
Obs. pers. |
3 |
|
Vittaria graminifolia Kaulf. |
75±12.5 |
62.5±11.2 |
– |
– |
– |
3 |
|
|
Total |
41,050±1,905.5 |
29,938±1,228.1 |
Referencias: 1 Martínez-Alfaro et al., 1995; 2 https://bens-jungle.com/Anthurium-scandens; 3 Ibarra-Manríquez et al., 1997; 4 Grupo Mesófilo, 2006; 5 https://bens-jungle.com/Monstera-acuminata-Philodendron-Terrarienpflanzen-Orchideen-Bromelien-Farne; 6 https://www.dutchrana.nl/de/shop/terrarienpflanzen-de/monstera-acuminata-2/; 7 https://orchideeen-shop.nl/monstera-acuminata.html; 8 Croat y Acebey, 2015; 9 Cano-Asseleih, 1997; 10 M. Leonti, 569 (XAL); 11 http://www.ecuagenera.com/Philodendron-tripartitum/en; 12 Bennett, 2000; 13 Flores-Palacios y Valencia-Díaz, 2007; 14 Red de Ornamentales, 2006; 15 Miranda-Jiménez et al., 2007; 16 Mondragón, 2008; 17 Munguía-Lino et al., 2010; 18 Hornung-Leoni, 2011; 19 Krömer et al., 2018; 20 Rauh, 1990; 21 Carmona, 1996; 22 Espejo-Serna, López-Ferrari et al., 2005; 23 Téllez-Velasco et al., 2012; 24 https://www.gob.mx/profepa/prensa/asegura-profepa-orquideas-y-bromelias-durante-operativo-en-huauchinango-puebla; 25 Avendaño-Montalvo, 2007; 26 Hágsater y Salazar, 1990; 27 Solano-Gómez et al., 2010; 28 Díaz-Toribio et al., 2013; 29 https://www.ebay.com/itm/Elaphoglossum-peltatum-Mexico-rare-terrarium-epiphytic-fern-frog-/321823470670/?_ul=AR; 30 https://www.glassboxtropicals.com/Elaphoglossum-peltatum-p/elaphpelt.htm; 31 Armenta-Montero et al., 2015; 32 Muñiz-Díaz de León et al., 2005; 33 http://tropical.theferns.info/viewtropical.php?id=Polypodium+plebeium
Para identificar posibles diferencias en la cantidad de EVC entre los 12 meses muestreados, se realizó un Anova de medidas repetidas, con el programa Past (V 3.10). Para conocer la suficiencia del esfuerzo de muestreo y determinar que fuera representativo de la riqueza de especies en el sitio, se realizó una curva de acumulación de especies utilizando el programa EstimateS (V 9.1.0). Para determinar la importancia de las especies en términos de su abundancia registrada durante los 12 meses de muestreo, se realizó una gráfica Log-Rank. En todos los análisis se utilizaron los números de plantas registradas en las parcelas de muestreo, y para la presentación de resultados se hicieron extrapolaciones del número de plantas caídas por hectárea en un año para facilitar comparaciones con otros estudios.
Resultados
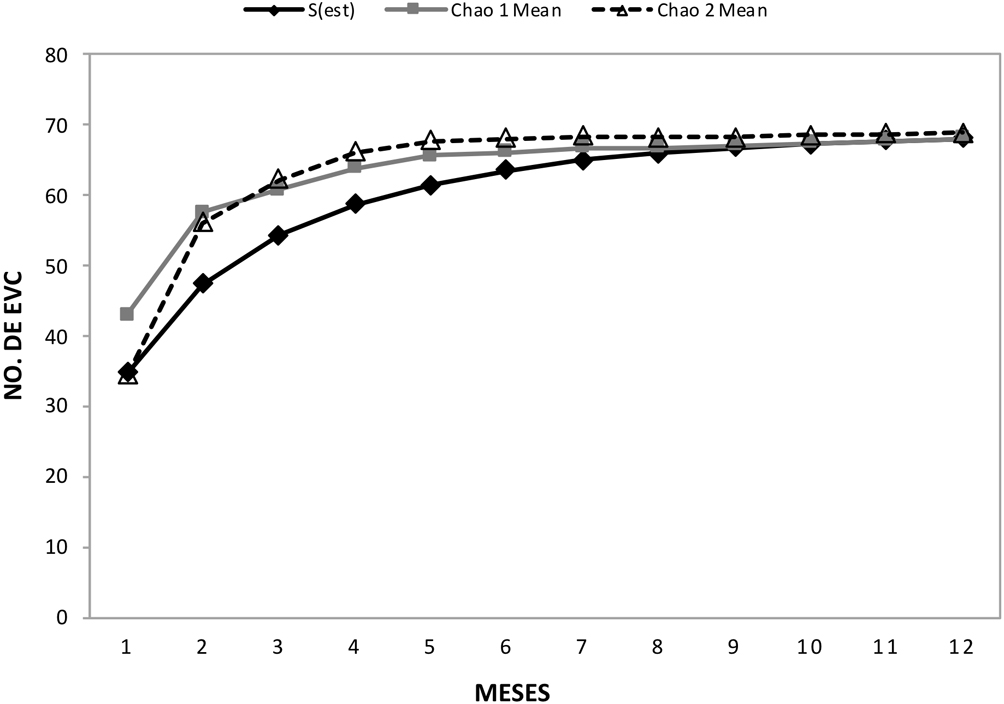
Se registraron, en total, 3,284 plantas de EVC en 0.08 ha (41,050 ± 1,906 plantas ha/año; tabla 1), las cuales representaron a 66 especies pertenecientes a 34 géneros y 11 familias. Las curvas de acumulación de especies observadas y estimadas indican que el esfuerzo de muestreo fue suficiente para alcanzar casi la totalidad de especies estimadas para el sitio. De acuerdo al estimador chao 1 se alcanzó el 97.1% de las 68 especies esperadas, mientras que de acuerdo con el estimador chao 2, se alcanzó 95.9% de las 68.8 especies esperadas (fig. 3).
Para el caso de las angiospermas, la familia Orchidaceae fue la más rica (26 spp.) seguida por Araceae y Bromeliaceae (5 cada una) y Piperaceae (4) (tabla 1). Dentro del grupo de las pteridofitas la familia Polypodiaceae fue la más rica en especies (11), seguida de Dryopteridaceae (5) y Aspleniaceae (3). En términos de abundancia, entre las pteridofitas las especies de Polypodiaceae fueron las más abundantes (9,312 ± 627 plantas ha/año) seguidas por Dryopteridaceae (8,112 ± 368 plantas ha/año) y para las angiospermas fueron las de la familia Orchidaceae (7,587 ± 356 plantas ha/año), seguida por Bromeliaceae (3,412 ± 208 plantas ha/año).
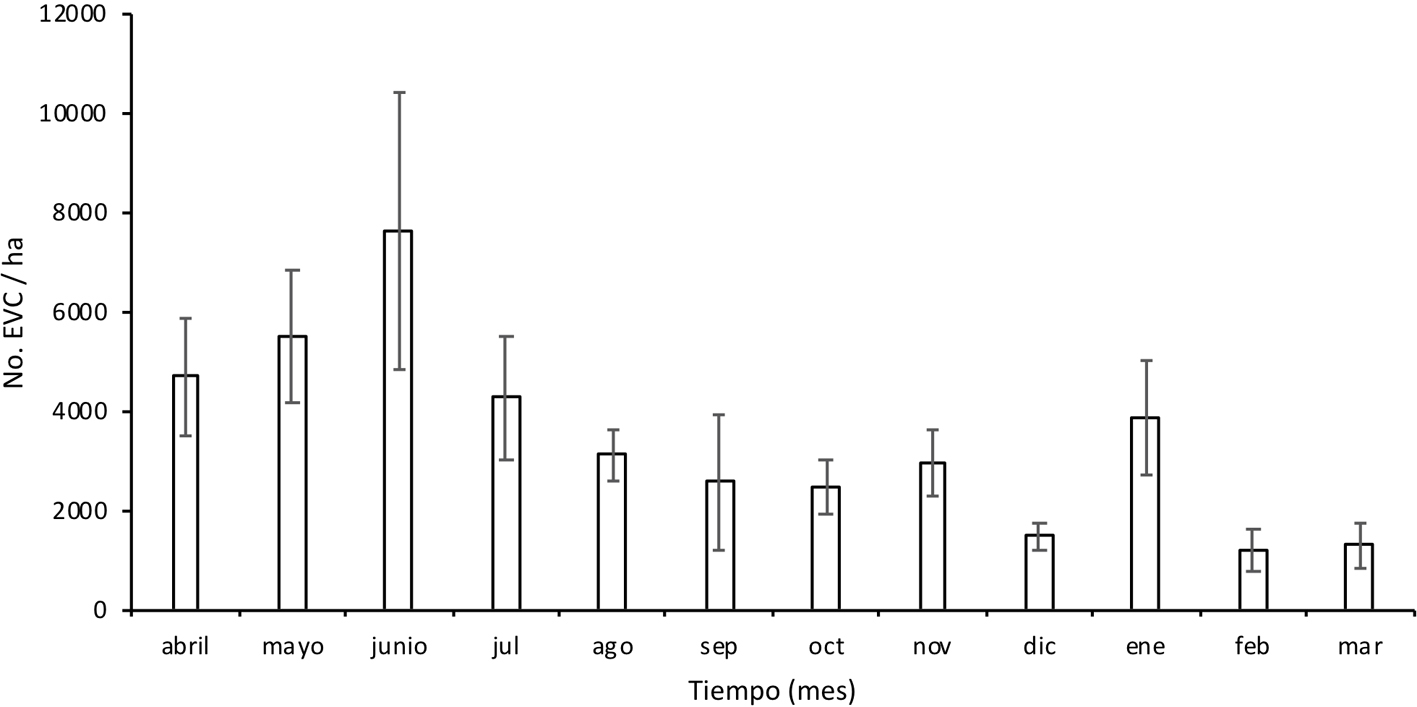
En la figura 4 se muestra la dinámica de la caída mensual de las epífitas vasculares a lo largo de un año, donde la abundancia de EVC difirió significativamente entre los meses del año de muestreo (gl = 11; F = 3.66; p = 0.01); junio presentó una mayor abundancia de EVC (7,638 ± 2,779 plantas/ha; promedio ± 1 error estándar) en comparación con el resto de los meses y el mes con menor abundancia fue febrero (1,187 ± 429 plantas/ha). A pesar de que junio fue el mes con la mayor abundancia de EVC no fue el que presentó mayor riqueza de especies (fig. 4).
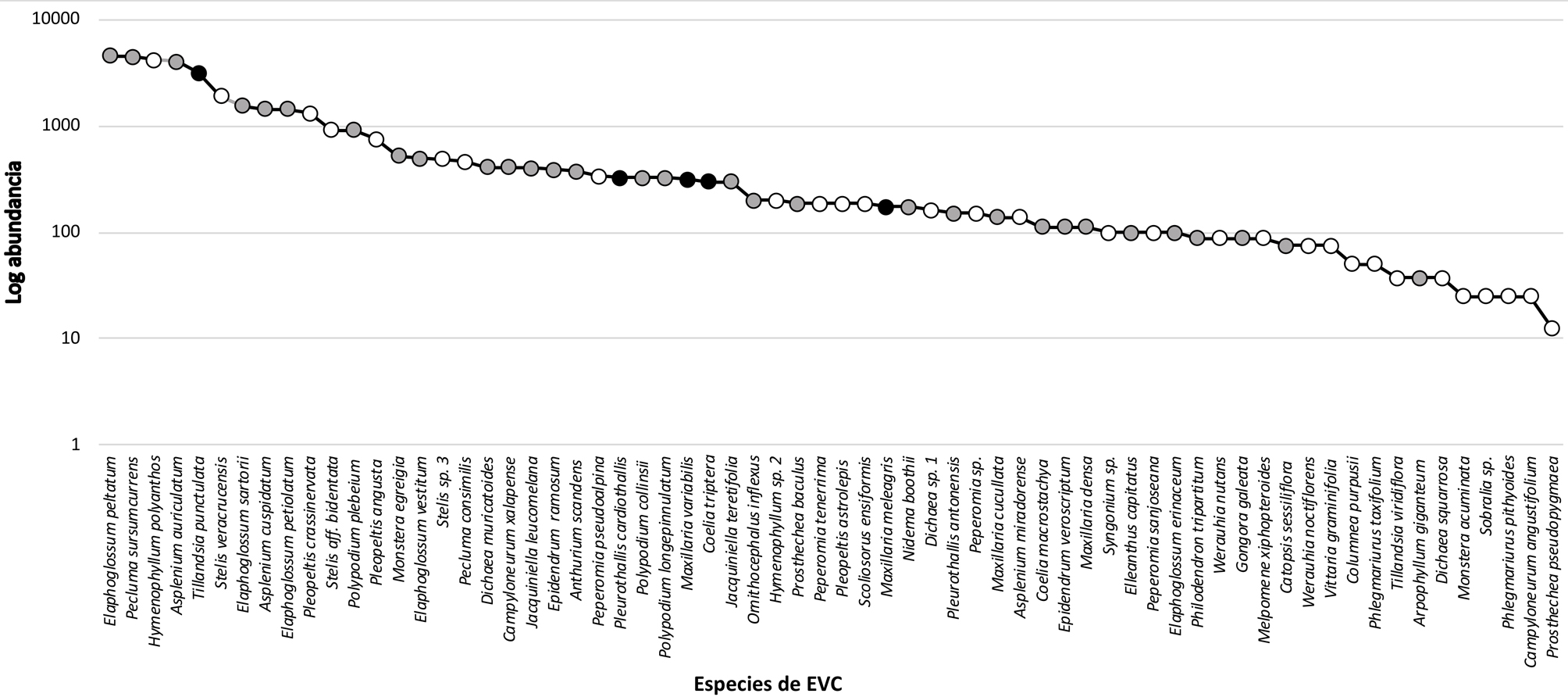
Entre las angiospermas las especies más abundantes son Tillandsia punctulata (Bromeliaceae), Stelis veracrucensis, S. aff. bidentata (Orchidaceae) y Monstera lechleriana (Araceae) (fig. 5, tabla 1). Las pteridofitas que presentaron mayor abundancia son Elaphoglossum peltatum (Dryopteridaceae), Pecluma sursumcurrens (Polypodiaceae), Hymenophyllum polyanthos (Hymenophyllaceae) y Asplenium auriculatum (Aspleniaceae). Tomando en cuenta solamente las EVC viables (tabla 1), para el caso de las angiospermas, Orchidaceae tuvo la mayor abundancia con 5,562 ± 244 plantas viables ha/año, de las cuales S. veracrucensis fue la más abundante con el 19.2% del total de los registros. Para Bromeliaceae, la especie que presentó mayor cantidad de plantas viables fue T. punctulata con el 61.5% del total del registro de esta familia. En Araceae la mayor abundancia de plantas viables fueron de Monstera egregia con el 25.2% del total de registros para esta familia. En el caso de Piperaceae las plantas de Peperomia pseudoalpina presentaron mayor porcentaje de plantas viables con el 25.8%.
Con base en la abundancia, la proporción de ejemplares viables y usos reportados u observados, se determinó que 5 especies (Tillandsia punctulata, Coelia triptera, Maxillaria meleagris, M. variabilis, Pleurothallis cardiothallis) tienen un alto potencial de aprovechamiento regional (fig. 2C-G), 31 especies tienen un potencial medio y otras 30 un potencial bajo (fig. 5, tabla 1). De las especies con potencial medio, la mayoría son de la familia Orchidaceae (15 spp.), seguidas por pteridofitas (12), Araceae (3) y Bromeliaceae (1). Entre las especies con potencial bajo, la mayoría son pteridofitas (13 spp.), seguidas por las de la familia Orchidaceae (7), Piperaceae (4), Bromeliaceae (3), Araceae (2) y Gesneriaceae (1).
Discusión
En un trabajo exhaustivo realizado en el dosel y sotobosque del BMM del volcán San Martín Tuxtla, entre 1,000 y 1,200 m se reportaron 103 especies de epífitas vasculares (Krömer et al., 2013). En el mismo sitio registramos 66 especies de EVC que corresponden al 64.1% del total reportado previamente, lo que representa un alto porcentaje de la riqueza registrada en los árboles. Además, los resultados indican que hay una semejanza entre la proporción de especies del dosel con las que se registraron en el suelo, por ejemplo, de las 103 especies registradas por Krömer et al. (2013), 36.9% pertenecen a pteridofitas, 32% a Orchidaceae, 8.7% a Piperaceae y 5.8% a Bromeliaceae, mientras que en el presente estudio estos grupos tienen proporciones parecidas de 37.9%, 39.4%, 6.1% y 7.6%, respectivamente. Solo las Araceae tuvieron un valor más alto en el dosel que en el suelo (14.6% y 7.6%, respectivamente), probablemente debido a que la mayoría de las especies son hemiepífitas, con hábitos de crecimiento en los troncos de los árboles y por lo tanto, son menos vulnerables a la caída. La relación entre epífitas del dosel y caídas concuerda con lo reportado previamente en otros BMM, en donde la diversidad y abundancia de bromelias epífitas observadas en el suelo del bosque estuvieron positivamente relacionadas con las encontradas en los árboles (Toledo-Aceves, García-Franco et al., 2014). De manera similar, Sarmento-Cabral et al. (2015) reportaron que las epífitas sobre las ramas tiradas tanto en una selva atlántica de Brasil como en una selva tropical de Panamá, representan el 80% de la riqueza de especies del dosel.
Nuestros resultados sobre la abundancia de las especies registradas en el suelo del bosque pueden dar una idea de la comunidad de epífitas del dosel en términos de estructura, composición y abundancia (Sarmento-Cabral et al., 2015; Toledo-Aceves, García-Franco et al., 2014). Sin embargo, también puede haber un sesgo porque en algunas especies la caída de plantas es mayor que en otras (y otras casi no caen, como ocurre con las aráceas hemiepífitas). Por ejemplo, las especies que acumulan mucha biomasa o tienen un crecimiento simpodial muy activo, podrían estar más propensas a que sus partes colgantes o no bien adheridas al sustrato se desprendan y caigan más fácilmente, lo que puede causar una sobreestimación de su abundancia real. Los estudios de epífitas que caen al suelo del bosque, además, pueden proporcionar información valiosa de las epífitas que ocupan las ramas delgadas (Chase, 1987) que conforman la zona exterior del dosel, la cual está sujeta a perturbaciones más frecuentes por efecto del viento y de la lluvia. Estas plantas son menos accesibles con técnicas de ascenso al dosel, por lo cual no siempre son registradas dentro de los muestreos (Sarmento-Cabral et al., 2015).
Los resultados obtenidos a nivel de familia también coinciden con lo reportado por Krömer et al. (2013); las familias más ricas fueron Polypodiaceae y Orchidaceae, aunque Araceae fue menos rica, posiblemente por su hábito de crecimiento mencionado anteriormente. En general, encontramos un bajo número de Bromeliaceae, sobre todo en comparación con el BMM del volcán Santa Marta, donde ocurren varias especies (p.ej. Catopsis subulata, Racinaea ghiesbreghtii, Tillandsia leiboldiana) que no fueron registradas en el San Martín Tuxtla (Krömer et al., 2013). Además, hay una ausencia de algunas bromelias xerotolerantes (Catopsis nitida, T. butzii, T. juncea, T. kirchhoffiana y T. multicaulis) que se encuentran en los BMM y vegetación derivada como acahuales y cafetales bajo sombra del centro de Veracruz, lo cual podría deberse a que muchas especies de bromelias tienden a dominar en ambientes más xéricos (García-Franco et al., 2008; Krömer et al., 2014; Susan-Tepetlan et al., 2015; Toledo-Aceves et al. 2012).
La mayor abundancia de bromelias epífitas caídas encontrada por Toledo-Aceves, García-Franco et al. (2014) en un BMM del centro de Veracruz (63,853 ± 7,789 plantas ha/año) y por Mondragón y Ticktin (2011) en bosque de pino-encino de Oaxaca (24,836 ± 9,710 plantas ha/año), en comparación al valor bajo de este grupo (3,412 ± 208 plantas ha/año) en nuestro estudio podría deberse a las diferencias entre los bosques, así como en la composición de especies. En los primeros existe una mayor riqueza de especies del género Tillandsia debido a la menor precipitación y humedad, además de una mayor luminosidad relacionada a la estructura de los bosques y un mayor grado de perturbación creando un ambiente más xérico. Varias especies de este género tienen adaptaciones ecomorfológicas que les permiten proliferar bajo condiciones secas, donde suelen ser más abundantes (Benzing, 2000; Flores-Palacios y García-Franco, 2004; Krömer et al., 2014).
En el BMM estudiado la caída constante de epífitas indica que se puede asegurar disponibilidad de plantas como recurso a aprovechar para su posible cultivo y posterior venta por parte de las comunidades locales a lo largo del año, sin embargo, es necesario evaluar su supervivencia en el proceso desde su recolección hasta su comercialización. La mayor abundancia de EVC registrada en el mes de junio podría estar relacionada con el patrón de precipitación y viento, ya que fue el primer mes del año estudiado con alta precipitación (459.3 mm; apéndice 1). Además, en este mes la tormenta tropical “Boris” afectó con fuertes lluvias y vientos (Conagua, 2014) que pudieron ocasionar la caída masiva de epífitas. El mes con menor abundancia de EVC fue febrero (1,200 ± 109 plantas ha/mes), lo que coincide con la baja precipitación en ese mes (178.9 mm). Cabe señalar que si bien mayo fue el mes que presentó menos precipitación (78.6 mm), y se registró la segunda abundancia más alta, ésta podría haber sido causada en parte por la caída de un árbol cuya copa cubierta de epífitas se encontró en una de las parcelas (fig. 2A). Lo anterior apoya que el flujo vertical (caída) de las epífitas vasculares no está determinado por un solo factor como la cantidad de precipitación sino por la interacción de varios factores como el viento, tormentas, sustrato inestable, animales e incluso por el peso mismo de las epífitas vasculares (Hietz, 1997; Matelson et al., 1993; Sarmento-Cabral et al., 2015; Strong, 1977; Zotz y Andrade, 2002).
Con base en las abundancias de EVC, proporción de ejemplares viables y usos observados o reportados, se determinó que solo T. punctulata, Coelia triptera, Maxillaria meleagris, M. variabilis y Pleurothallis cardiothallis tienen un alto potencial de aprovechamiento regional. Por ejemplo, para la bromelia T. punctulata se registraron 2,100 plantas viables ha/año y tiene uso ceremonial o de ornato (Hornung-Leoni, 2011; Miranda-Jiménez et al., 2007; Mondragón, 2008; Munguía-Lino et al., 2010) no solo en el estado de Veracruz sino también en el Estado de México y en Oaxaca. Además, esta bromelia tiene demanda en el mercado hortícola internacional como planta de ornato (Rauh, 1990). Regionalmente, es extraída de sus hábitats para decorar los arcos florales que adornan las iglesias en las fiestas religiosas, pero sin un manejo adecuado (Haeckel, 2008; Krömer et al., 2018). Así mismo, se ha registrado dentro del comercio ilegal en el mercado del centro de Veracruz (Flores-Palacios y Valencia-Díaz, 2007).
Las otras especies con alto potencial de aprovechamiento mencionadas anteriormente son orquídeas, para las cuales se registraron entre 150-262 plantas viables ha/año. Todas tienen un uso ornamental observado o reportado para la región de Los Tuxtlas y Veracruz (Carmona, 1996; Flores-Palacios y Valencia-Díaz, 2007), así como para México en general (Espejo-Serna, López-Ferrari et al., 2005; Red de Ornamentales, 2006). Además, estas 4 orquídeas se ofertan por un precio entre 12-24 USD por planta en el internet.
Entre las EVC con potencial medio de comercialización también destacan las orquídeas, siendo Arpophyllum giganteum, Gongora galeata, Maxillaria cucullata y Prosthechea baculus, especies con flores o inflorescencias grandes y vistosas, cuyo uso ornamental fue reportado para Los Tuxtlas y/o Veracruz (Carmona, 1996; Flores-Palacios y Valencia-Díaz, 2007; Ibarra-Manríquez et al., 1998), así como para el Estado de México o el país en general (Espejo-Serna, López-Ferrari, Jiménez-Machorro et al., 2005; Hágsater y Salazar, 1990; Munguía-Lino et al., 2010; Red de Ornamentales, 2006). Sin embargo, para estas se registraron valores bajos entre 37-87 plantas viables ha/año, por lo cual su cultivo y propagación en el vivero podría aumentar los costos de producción.
Las pteridofitas no incluyen ninguna especie con alto potencial, pero hay varias dentro de los géneros Asplenium, Elaphoglossum y Polypodium con follaje vistoso y forma de crecimiento atractiva en roseta o colgante, que tienen un potencial medio, entre ellas A. auriculatum, A. cuspidatum, E. erinaceum y P. longepinnulatum que fueron incluidas en la lista de las plantas ornamentales de México (Red de Ornamentales, 2006). Entre las 5 EVC de Elaphoglossum, resalta E. vestitum, con largas frondas cubiertas por escamas naranjas, para la cual se registró un valor de 212 plantas viables ha/año, a diferencia de otras menos atractivas (p. ej., E. petiolatum, E. sartorii) que alcanzaron valores de más de 1,000 plantas viables ha/año. Se ha reportado que especies del género Elaphoglossum son utilizadas en Estados Unidos de América como complemento de arreglos florales (Muñiz-Díaz de León et al., 2007), mientras que diferentes especies de Asplenium tienen usos ornamental y medicinal en México, Cuba y otras partes del mundo (Muñiz-Díaz de León et al., 2007; Núñez-Barrizonte et al., 2011; Rendón y Fernández-Nava, 2007). Por otro lado, se reporta a Phlegmariurus taxifolius con uso ceremonial y ornamental, la cual fue registrada con muy baja abundancia. Esta especie está en venta dentro del comercio ilegal en los mercados del centro de Veracruz (Flores-Palacios y Valencia-Díaz, 2007) y fue clasificada como vulnerable, según criterios de la Unión Internacional para la Conservación de la Naturaleza (UICN) a nivel regional (Armenta-Montero et al., 2015).
Para la familia Bromeliaceae solo existe una especie, C. sessiliflora, que tiene potencial medio, debido a una menor abundancia. Esta se ha propuesto para aprovechamiento ornamental (Díaz-Toribio et al., 2013) y fue reportada también dentro del comercio ilegal en Veracruz (Flores-Palacios y Valencia-Díaz, 2007). Para el caso de las Araceae caídas, su riqueza y abundancia generalmente es escasa. Las especies más abundantes y con potencial medio, Anthurium scandens y Monstera egregia, pueden ser utilizadas como plantas de ornato (Croat y Acebey, 2015; Martínez-Alfaro et al., 1995), al igual que la menos abundante Philodendron tripartitum, que además es una especie que tiene uso medicinal, comestible y para la cestería (Cano-Asseleih, 1997; Martínez-Alfaro et al., 1995).
Muchas EVC de diferentes familias, principalmente Orchidaceae (p.ej. Coelia macrostachya, Dichaea muricatoides, G. galeata, M. cucullata), Bromeliaceae (C. sessiliflora, T. punctulata) y del grupo de las pteridofitas (Pecluma sursumcurrens, Elaphoglossum peltatum, A. auriculatum), son encontradas en los traspatios de las casas de la gente de la comunidad Adolfo Ruiz Cortines (E. Francisco-Ventura, obs. pers.). La mayoría de estas especies son vistosas por su follaje, inflorescencias o flores de formas conspicuas y de colores, por lo cual tienen un alto valor ornamental (Díaz-Toribio et al., 2013; Flores-Palacios y Valencia-Díaz, 2007; Hornung-Leoni, 2011; Miranda-Jiménez et al., 2007; Núñez-Barrizonte et al., 2011; Solano-Gómez et al., 2010). Estos traspatios podrían ser objetos de estudio a largo plazo para observar la supervivencia de las especies y su adaptabilidad al cultivo, sentando algunas bases para una futura comercialización a nivel local. Cabe resaltar que el aprovechamiento y uso comercial de EVC como PFNM debe realizarse dentro del marco de la Ley General de Desarrollo Forestal Sustentable (si se trata de especies no incluidas en la NOM 059-Semarnat-2010) o de la Ley General de Vida Silvestre (si se trata de especies incluidas en la NOM-059-Semarnat-2010) y sus respectivos reglamentos. Sin embargo, como ha sido tratado en otros trabajos, los requerimientos para realizar el manejo de forma legal pueden desincentivar a los dueños del recurso (Krömer et al., 2018; Mondragón y Ticktin, 2011; Toledo-Aceves, García-Franco et al., 2014), por lo que se requiere una adecuación de la normatividad, así como de apoyos para el aprovechamiento de PFNM por parte de comunidades rurales y pequeños propietarios.
Las epífitas que caen al suelo generalmente mueren debido a factores como patógenos y herbívoros, disminución de fotosíntesis e intercambio de gases por la poca intensidad lumínica y gran humedad del suelo, y posteriormente se convierten en nutrientes que se mineralizan y son absorbidos por la vegetación terrestre (Matelson et al., 1993; Mondragón et al., 2004). Por lo tanto, la recolecta de EVC implica una reducción en el aporte de nutrientes en el ecosistema, donde la magnitud de este impacto dependería de la biomasa de cada especie viable para el cultivo que cae al suelo, pero también del tipo de hábitat. Sin embargo, Mondragón y Ticktin (2011) han reportado que en un bosque templado de Oaxaca las contribuciones relativas de bromelias caídas a la concentración de nutrientes del suelo son < 1%. Por otro lado, Toledo-Aceves, García-Franco et al. (2014) mencionan que el 60% de las bromelias epífitas caídas en un BMM del centro de Veracruz se encontraban en condiciones para cultivo o venta y que la extracción de estas representa una biomasa relativamente baja. No obstante, existe muy poca información sobre la contribución específica que tienen las epífitas de diferentes familias al ciclo de nutrientes. Para reducir el posible efecto en el ciclo de nutrientes del bosque, se podría implementar un esquema en el cual se distribuya la colecta de EVC en diferentes áreas.
En conclusión, el área de estudio muestra un potencial de aprovechamiento considerable de EVC fundamentado en los siguientes elementos: a) existe una alta riqueza de especies de epífitas vasculares (66 spp.) que caen al suelo del bosque; b) 72.9% de las EVC se encuentran en buen estado para su cultivo; c) la abundancia registrada de 36 especies con alto y medio potencial de aprovechamiento es suficiente como para evaluar su manejo y futura comercialización regional; d) los grupos con mayor riqueza y abundancia son orquídeas, pteridofitas y bromelias, cuyas especies son de las más valoradas en la comunidad local, además tienen diferentes usos en Veracruz y otros estados del país. Sin embargo, la comercialización aún debe ser evaluada dado que esta constituye el cuello de botella en la mayoría de los proyectos de aprovechamiento de PFNM (Marshall et al., 2006). Si bien no se abordó la organización de las comunidades locales en torno al manejo de las EVC como PFNM en este trabajo, es importante resaltar que su autoorganización es indispensable en los proyectos encaminados al manejo controlado de recursos del bosque (Ostrom, 2000; Stockdale, 2005).
Agradecimientos
Queremos agradecer a P. Díaz-Jiménez, P. Aguilar-Rodríguez, S. Castelán-Culebro y J. Y. S. Ruiz-Cruz, así como a los miembros del Grupo Ecoturístico “Los Clarines” en la comunidad de Ruiz Cortínez por su apoyo en el trabajo de campo. A C. I. Carvajal-Hernández, D. Vergara-Rodríguez y A. R. Acebey, por su apoyo con la identificación de ejemplares; a J. C. López-Acosta por su apoyo en el análisis estadístico; a S. Armenta-Montero por la elaboración del mapa; a Lilia Ruiz-Ruiz por preparar la lámina de fotos; a A. R. Acebey por sus valiosos comentarios y a Keith MacMillan por la revisión del abstract. La investigación fue financiada con una beca de Maestría del Conacyt (Núm. 364403) otorgada a EFV, así como un financiamiento del Sistema Nacional de Recursos Fitogenéticos para la Alimentación y la Agricultura (SINAREFI), red orquídeas, mediante el Proyecto integral de la Red Orquídeas para su conservación propagación y uso sustentable (Núm. ORN-ORQ-13-2) otorgada a RAMG.
Referencias
Acebey, A., Kessler, M. y Maass, B. (2007). Potencial de aprovechamiento de Araceae y Bromeliaceae como recursos no maderables en el bosque montano húmedo del Parque Nacional Cotapata, Bolivia. Ecología en Bolivia, 42, 4–22.
Acebey, A., Krömer, T., Maass, B. L. y Kessler, M. (2010). Ecoregional distribution of potentially useful species of Araceae and Bromeliaceae as non-timber forest products in Bolivia. Biodiversity and Conservation, 19, 2553–2564.
Acebey, A. R., Krömer, T., Vázquez-Torres, M. y Tejero-Díez, J. D. (2015). Helechos y licófitos de la Reserva de la Biosfera Los Tuxtlas, Veracruz, México. Botanical Sciences, 93, 1–32.
Alexiades, M. N. y Shanley, P. (2004). Productos forestales, medios de subsistencia y conservación: estudios de caso sobre sistemas de manejo de productos forestales no maderables, Volumen 3. América Latina. Bogor, Indonesia: CIFOR.
Armenta-Montero, S., Carvajal-Hernández, C. I., Ellis, E. A. y Krömer, T. (2015). Distribution and conservation status of Phlegmariurus (Lycopodiaceae) in the state of Veracruz, Mexico. Tropical Conservation Science, 8, 114–137.
Avendaño-Montalvo, O. (2007). Inventario de orquídeas en Chilchotla, Oaxaca (Tesis). Texcoco, Estado de México: Departamento de Suelos, Universidad Autónoma Chapingo.
Bennett, B. (1995). Ethnobotany and economic botany of epiphytes, lianas, and other host-dependent plants: an overview. En M. D. Lowman y N. K. Nadkarni (Eds.), Forest canopies (pp. 558–559). San Diego: Academic Press.
Bennett, B. (2000). Ethnobotany of Bromeliaceae. En D. H. Benzing (Eds.), Bromeliaceae. Profile of an adaptative radiation (pp. 587–608). Cambridge: Cambridge University Press.
Benzing, D. H. (1990). Vascular epiphytes. General biology and related biota. Cambridge: Cambridge University Press.
Benzing, D. H. (2000). Bromeliaceae. Profile of an adaptive radiation. Cambridge: Cambridge University Press.
Cano-Asseleih, L. M. (1997). Flora medicinal de Veracruz: inventario etnobotánico. Xalapa: Universidad Veracruzana.
Carmona, G. (1996). Las orquídeas del parque de Flora y Fauna Silvestre Tropical, Catemaco, Veracruz, Mexico (Tesis). Facultad de Biología, Universidad Veracruzana. Xalapa, Veracruz.
Chase, M. W. (1987). Obligate twig epiphytism in the Oncidiinae and other neotropical orchids. Selbyana, 10, 24–30.
Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad). (2010). El bosque mesófilo de montaña en México: amenazas y oportunidades para su conservación y manejo sostenible. México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Conagua (Comisión Nacional del Agua). (2014). Reseña de la tormenta tropical “Boris” del océano Pacífico. Recuperado el 17 septiembre, 2016 de: http://smn.cna.gob.mx/tools/DATA/Ciclones%20Tropicales/Ciclones/2014-Boris.pdf
Croat, T. B. y Acebey, A. R. (2015). Araceae. Flora de Veracruz: fascículo 164. Xalapa: Insituto de Ecología, A.C./ Centro de Investigaciones Tropicales, Universidad Veracruzana.
Cruz-García, G., Lagúnez-Rivera, L., Chávez-Angeles, M. G. y Solano-Gómez, R. (2015). The wild orchid trade in a Mexican local market: diversity and economics. Economic Botany, 69, 291–305.
Damon, B. A. (2006). Cultivo rústico y sustentable de orquídeas nativas del Soconusco. Tuxtla Gutiérrez: Fundación Produce Chiapas, A.C.
Díaz-Toribio, M. H., Toledo-Aceves, T., Mata-Rosas, M., Mehltreter, K., Hernández-Rojas, A. C., Mejía-Alemán, J. et al. (2013). Manual de cultivo de orquídeas, bromelias y helechos en cafetales de sombra. Xalapa: Instituto de Ecología, A.C.
Elliott, D. D. y Ticktin, T. (2013). Epiphytic plants as NTFPs from the forest canopies: priorities for management and conservation. En M. Lowman, S. Devy y T. Ganesh (Eds.), Treetops at risk. Challenges of global canopy ecology and conservation (pp. 435–444). New York: Springer.
Espejo-Serna, A. (2014). Las plantas vasculares de los bosques mesófilos de montaña en México. En M. Gual-Díaz y A. Rendón-Correa (Comps.), Bosques mesófilos de montaña de México: diversidad, ecología y manejo (pp. 189–195). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Espejo-Serna, A., López-Ferrari, A. R., Jiménez-Machorro, R. y Sánchez Saldaña, L. (2005). Las orquídeas de los cafetales en México: una opción para el uso sostenible de ecosistemas tropicales. Revista de Biología Tropical, 53, 73–84.
Espejo-Serna, A., López-Ferrari, A. R. y Ramírez- Morillo, I. (2005). Bromeliaceae. Flora de Veracruz, fascículo 136. Xalapa: Instituto de Ecología, A.C.
Flores-Palacios, A. y García-Franco, J. G. (2004). Effect of isolation on the structure and nutrient content of oak epiphyte communities. Plant Ecology, 173, 259–269.
Flores-Palacios, A. y Valencia-Díaz, S. (2007). Local illegal trade reveals unknown diversity and involves a high species richness of wild vascular epiphytes. Biological Conservation, 136, 372–387.
García-Franco, J. G., Castillo-Campos, G., Mehltreter, K., Martínez, M. L. y Vázquez, G. (2008). Composición florística de un bosque mesófilo del centro de Veracruz, México. Boletín de la Sociedad Botánica de México, 83, 37–52.
Gentry, A. H. y Dodson, C. H. (1987). Diversity and biogeography of neotropical vascular epiphytes. Annals of the Missouri Botanical Garden, 74, 205–233.
Gómez, L., Mondragón, D. y Méndez, E. (2015). Aprovechamiento sustentable de bromelias epífitas: Propuesta comercial del vivero comunitario “Las Bromelias”, Santa Catarina Ixtepeji, Oaxaca. Revista de Investigación Académica sin Frontera, 21, 1–20.
Grupo Mesófilo. (2006). Cestería. Productos Forestales no maderables de la Chinantla, Oaxaca. Boletín Vol. 1, Núm. 5. Oaxaca: Grupo Mesófilo, A.C.
Gual-Díaz, M. y González-Medrano, F. (2014). Los bosques mesófilos de montaña en México. En M. Gual-Díaz y A. Rendón-Correa (Eds.), Bosques mesófilos de montaña de México: diversidad, ecología y manejo (pp. 27–67). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Guevara, S., Laborde J. y Sánchez-Riíos, G. (Eds.). (2004). Los Tuxtlas. El paisaje de la sierra. Xalapa: Instituto de Ecología, A.C.
Haeckel, I. B. (2008). The “Arco Floral”: ethnobotany of Tillandsia and Dasylirion spp. in a Mexican religious adornment. Economic Botany, 62, 90–95.
Hágsater, E. y Salazar, G. (1990). Orchids of Mexico. Icones Orchidacearum. Fascículo 1, Parte 1. México D.F.: Asociación Mexicana de Orquideologia, A.C.
Hietz, P. (1997). Population dynamics of epiphytes in a mexican humid montane forest. Journal of Ecology, 6, 767–775.
Hietz, P. (2010). Ecology and ecophysiology of epiphytes in tropical montane cloud forests. En L. A. Bruijnzeel, F. N. Scatena y L. S. Hamilton (Eds.), Tropical montane cloud forests. Science for conservation and management (pp. 67–76). Cambridge: Cambridge University Press.
Hofstede, R. G. M., Wolf, G. D. H. y Benzing, D. H. (1993). Epiphytic biomass and nutrient status of a Colombian upper montane rain forest. Selbyana, 14, 37–45.
Hornung-Leoni, C. T. (2011). Avances sobre usos etnobotánicos de las Bromeliaceae en Latinoamérica. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas, 10, 297–314.
Ibarra-Manríquez, G., Ricker, M., Ángeles, G., Sinaca, C. S. y Sinaca, C. M. A. (1998). Useful plants of the Los Tuxtlas rainforest (Veracruz, México): considerations on their market potential. Economic Botany, 51, 362–376.
Krömer, T., Acebey, A., Kluge, J. y Kessler, M. (2013). Effects of altitude and climate in determining elevational plant species richness patterns: a case study from Los Tuxtlas, Mexico. Flora, 208, 197–210.
Krömer, T., Acebey, A. y Toledo-Aceves, T. (2018). Aprovechamiento de plantas epífitas: implicaciones para su conservación y manejo sustentable. En E. Silva-Rivera, V. Martínez-Valdéz, M. Lascurain y E. Rodríguez-Luna (Eds.), De la recolección a los agroecosistemas: soberanía alimentaria y conservación de la biodiversidad (pp. 175-196). Xalapa: Editorial de la Universidad Veracruzana.
Krömer, T., García-Franco, J. G. y Toledo-Aceves, T. (2014). Epífitas vasculares como bioindicadores de la calidad forestal: impacto antrópico sobre su diversidad y composición. En C. A. González-Zuarth, A. Vallarino, J. C. Pérez-Jiménez y A. M. Low-Pfeng (Eds.), Bioindicadores: guardianes de nuestro futuro ambiental (pp. 606–623). México D.F., Campeche: Instituto Nacional de Ecología y Cambio Climático/ El Colegio de la Frontera Sur.
Krömer, T., Kessler, M. y Gradstein, S. R. (2007). Vertical stratification of vascular epiphytes in submontane and montane forest of the Bolivian Andes: the importance of the understory. Plant Ecology, 189, 261–278.
Marshall, E., Schreckenberg, K. y Newton, A. C. (Eds). (2006). Comercialización de productos forestales no maderables: factores que influyen en el éxito. Conclusiones del estudio de México y Bolivia e implicancias políticas para los tomadores de decisión. Cambridge, RU: Centro Mundial de Vigilancia de la Conservación del PNUMA.
Martínez-Alfaro, M. A., Evangelista-Oliva, V., Mendoza-Cruz, M., Morales-García, G., Toledo-Olazcoaga, G. y Wong-León, A. (1995). Catálogo de plantas útiles de la sierra Norte de Puebla, México. Cuadernos del Instituto de Biología 27. México D.F.: Instituto de Biología, Universidad Nacional Autónoma de México.
Matelson, T. J., Nadkarni, N. M. y Longino, J. T. (1993). Longevity of fallen epiphytes in a neotropical montane forest. Ecology, 74, 265–269.
Mickel, J. y Smith, A. R. (2004). The pteridophytes of Mexico. Memoirs of the New York Botanical Garden, 88, 1–1054.
Miranda-Jiménez, M. E., Arellano-Mijangos, J. J., Salazar-Acevedo, B. S. Hernández-Martínez, F., Quero-Cruz R. y Pérez-Santiago, L. (2007). Bases para el manejo comunitario de bromelias ornamentales. Colección manejo campesino de recursos naturales. Oaxaca: GAIA, RAISES.
Mondragón, D. (2008). La comercialización navideña de bromelias epífitas en la Ciudad de Oaxaca, México. Etnobiología, 6, 24–28.
Mondragón, D. (2009). Population viability analysis for an endangered epiphytic orchid: Guarianthe (Cattleya) aurantiaca (Bateman ex Lindley) Dressler and W. E. Higgins. Plant Species Biology, 24, 35–41.
Mondragón, D., Calvo-Irabien, L. M. y Benzing, D. H. (2004). The basis for obligate epiphytism in Tillandsia brachycaulos (Bromeliacaea) in Mexican tropical dry forest. Journal of Tropical Ecology, 20, 97–104.
Mondragón, D. y Ticktin, T. (2011). Demographic effects of harvesting epiphytic bromeliads and an alternative approach to collection. Conservation Biology, 25, 797–807.
Mondragón, D. y Villa-Guzmán, D. M. (2008). Estudio etnobotánico de las bromelias epífitas en la comunidad de Santa Catarina Ixtepeji. Polibotánica, 26, 175–191.
Munguía-Lino, G., Vázquez-García, L. M. y López-Sandoval, J. A. (2010). Plantas silvestres ornamentales comercializadas en los mercados de la Flor de Tenancingo y Jamaica, México. Polibotánica, 29, 281–308.
Muñiz-Díaz de León, M. E., Mendoza-Ruíz, A. y Pérez-García, B. (2007). Usos de los helechos y plantas afines. Etnobiología, 5, 117–125.
Núñez-Barrizonte, C. A., Fernández-Castillo, L. B. y Vento-Rivero, I. (2011). Contribución al estudio de las epífitas como producto forestal no maderable de valor cultural. Revista Forestal Baracoa, 30, 37–42.
Ostrom, E. (2000). Social capital: a fad or a fundamental concept. Social capital: A Multifaceted Perspective, 172, 195–98.
Peck, J. L. E. y Christy, J. A. (2004). Putting the stewardship concept into practice: commercial moss harvest in Northwestern Oregon, USA. Forest Ecology and Management, 225, 225–233.
Rauh, W. (1990). Bromelien: Tillandsien und andere kulturwürdige Bromelien. Stuttgart: Ulmer.
Red de Ornamentales. (2006). Plantas ornamentales de México. Consultado 17 junio 2017. http://bva.colech.edu.mx/xmlui/bitstream/handle/123456789/HASH0137425108dc2a555962892c/doc.pdf?sequence=2
Rendón, C. A. y Fernández-Nava, R. (2007). Plantas con potencial uso ornamental del Estado de Morelos, México. Polibotánica, 23, 121–164.
Rzedowski, J. (1996). Análisis preliminar de la flora vascular de los bosques mesófilos de montaña de México. Acta Botanica Mexicana, 35, 25–44.
Sánchez-Ramos, G. y Dirzo, R. (2014). El bosque mesófilo de montaña: un ecosistema prioritario amenazado. En M. Gual-Díaz y A. Rendón-Correa (Eds.), Bosques mesófilos de montaña de México: diversidad, ecología y manejo (pp. 109–140). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Sarmento-Cabral J., Petter, G., Mendieta-Leiva, G., Wagner, K., Zotz, G. y Kreft, H. (2015). Branchfall as a demographic filter for epiphyte communities: lessons from forest floor-based sampling. Plos One, 10, e0128019.
Scatena, F. N., Bruijnzeel, L. A., Bubb, P. y Das, S. (2010). Setting the stage. En L. A. Bruijnzeel, F. N. Scatena y L. S. Hamilton (Eds.), Tropical montane cloud forests science for conservation and management (pp. 38–63). Cambridge: Cambridge University Press.
Solano-Gómez, G. R., Cruz-Lustre, G., Martínez-Feria, A. y Lagunez-Rivera, L. (2010). Plantas utilizadas en la celebración de la Semana Santa en Zaachila, Oaxaca, México. Polibotánica, 29, 263–279.
Soto-Arenas, M. A., Hágsater, E., Jiménez, R., Salazar, G. A., Solano-Gómez, R., Flores, R. et al. (2007). Las orquídeas de México. Catálogo digital. México D.F: Instituto Chinoín, A.C.
Stockdale, M. (2005). Steps to sustainable and community-based NTFP management: a manual written with special reference to South and Southeast Asia. Desa Putera, Philippines: SMT Grafika.
Strong, D. R. (1977). Epiphyte loads, tree falls, and perennial forest disruption: a mechanism for maintaining higher tree species richness in the tropics without animals. Journal of Biogeography, 4, 215–218.
Susan-Tepetlan, T. M., Velázquez-Rosas, N. y Krömer, T. (2015). Cambios en las características funcionales de epífitas vasculares de bosque mesofilo de montaña y vegetación secundaria en la región central de Veracruz, México. Botanical Sciences, 93, 153–163.
Téllez-Velasco, M. A. (2012). La importancia de la conservación ex situ, un ejemplo: la Colección de Orquídeas del Jardín Botánico del Instituto de Biología de la Universidad Nacional Autonoma de Mexico. En M. A. Téllez-Velasco (Eds.), Conservación de orquídeas en México (pp. 79–87). México D.F.: Universidad Nacional Autónoma de México.
Toledo-Aceves, T., García-Franco, J. G., Hernández-Rojas, A. y MacMillan, K. (2012). Recolonization of vascular epiphytes in a shaded coffee agroecosystem. Applied Vegetation Science, 15, 99–107.
Toledo-Aceves, T., García-Franco, J. G. y López-Barrera, F. (2014). Bromeliad rain: an opportunity for cloud forest management. Forest Ecology and Management, 329, 129–136.
Toledo-Aceves, T., Hernández-Apolinar, M. y Valverde, T. (2014). Potential impact of harvesting on the population dynamics of two epiphytic bromeliads. Acta Oecologica, 59, 52–61.
Vergara-Rodríguez, D. (2009). Las especies del género Peperomia (Piperaceae) del estado de Veracruz, un estudio preliminar (Tesis). Facultad de Biología, Universidad Veracruzana. Xalapa, Veracruz.
Villaseñor, J. L. (2016). Checklist of the native vascular plants of Mexico. Revista Mexicana de Biodiversidad, 87, 559–902.
Winkler, M., Hülber, K. y Hietz, P. (2007). Population dynamics of epiphytic bromeliads: life strategies and the role of host branches. Basic and Applied Ecology, 8, 183–196.
Zotz, G. y Andrade, J. L. (2002). La ecología y la fisiología de las epífitas y las hemiepífitas. En M. R. Guariguata y G. H. Kattan (Eds.), Ecología y conservación de bosques neotropicales (pp. 271–296). San José: Libro Universitario Regional del Instituto Tecnológico de Costa Rica.


