Patrones de distribución de las grandes macroalgas en el golfo de México y el Caribe mexicano: una contribución a la biología de la conservación
Martha Isabel Vilchis a, b, *, Kurt M. Dreckmann a, Erick A. García-Trejo c,
Oscar E. Hernández a, b y Abel Sentíes a
a Departamento de Hidrobiología, Universidad Autónoma Metropolitana-Iztapalapa, Apartado postal 55-535, 09340 Ciudad de México, México
b Doctorado en Ciencias Biológicas y de la Salud, Universidad Autónoma Metropolitana-Iztapalapa, Apartado postal 55-535, 09340 Ciudad de México, México
c Unidad de Informática para la Biodiversidad, Departamento de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional Autónoma de México, Apartado postal 70-399, 04510 Ciudad de México, México
* Autor para correspondencia: miva.uam@gmail.com (M.I. Vilchis)
Resumen
Se realizó un análisis biogeográfico y de diversidad a partir de la distribución geográfica de 118 especies de grandes macroalgas (Chlorophyta, Ochrophyta y Rhodophyta) presentes en el golfo de México y Caribe mexicano, con el propósito de identificar patrones de distribución que puedan resultar en posibles áreas de conservación y robustecer a las previamente establecidas. El análisis de parsimonia de endemismos (PAE) resultó en 9 áreas definidas por sinapomorfías y 4 más, definidas por autapomorfías. El análisis de trazos permitió identificar 2 trazos generalizados y 1 nodo biogeográfico ubicado en el arrecife Alacranes, resultado de la complejidad geológica del área de estudio. Dicho nodo corresponde a una de las áreas con mayor complejidad taxonómica para el golfo de México, además de albergar el mayor número de especies y junto con el sur de Tamaulipas presentan los valores más altos de complementariedad. Con base en la integración de los análisis anteriores, se propone como área a conservar al arrecife Alacranes, que además coincide con uno de los sitios prioritarios marinos considerado de extrema importancia para la conservación. Este trabajo constituye la primera aproximación biogeográfica en el estudio de las macroalgas marinas bentónicas en México y representa una primera etapa bajo el enfoque de la biogeografía evolutiva.
Palabras clave:
Análisis de trazos; Conservación marina; Complementariedad; Macroalgas; PAE
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Distribution patterns of seaweeds in the Gulf of Mexico and Mexican Caribbean: a contribution to conservation biology
Abstract
A biogeographic and a species richness analysis were done using the geographic distribution of 118 species of seaweeds (Chlorophyta, Ochrophyta and Rhodophyta) in the Gulf of Mexico and Mexican Caribbean, in order to identify areas for conservation, and to enhance the previously established ones. The parsimony analysis of endemicity (PAE) revealed 9 groups defined by the presence of synapomorphies, and 4 quadrats were recognized based on autapomorphies. Track analysis allowed to define 2 generalized tracks and 1 panbiogeographic node located in the Alacranes Reef, as a result of the complex geological history of the region. This node corresponds to the area with the highest taxonomic complexity in the Gulf of Mexico, the highest species richness, and together with Tamaulipas southern quadrat, shows the highest values of species complementarity. As a product of the integration of the previous analyses, we propose the Alacranes Reef as a site of extreme marine importance for conservation, this been congruent with one of the priority sites already established there. This research constitutes the first biogeographic approach in the study of benthonic marine seaweeds in Mexico and also represents a first step within the framework of evolutionary biogeography.
Keywords:
Track analysis; Marine conservation; Complementarity; Seaweeds; PAE
El análisis de parsimonia de endemismos (PAE, por sus siglas en inglés) y el análisis de trazos, son 2 de los métodos más utilizados para identificar patrones de distribución con fines de conservación biológica. El PAE permite clasificar áreas en un cladograma a través de la solución más simple, basada en la presencia de taxones compartidos derivados (Nihei, 2006). Mediante este análisis se han podido proponer regionalizaciones que permiten categorizar a las áreas en términos de su biota (Escalante, 2009). Dichas áreas al presentar una combinación única de especies, resultan útiles en la planeación ambiental para priorizar sitios de conservación (Posadas y Miranda-Esquivel, 1999).
Por otra parte, un análisis panbiogeográfico parte de la construcción de trazos individuales de organismos filogenéticamente distantes, los cuales constituyen las coordenadas primarias de un taxón en el espacio, es decir, la unión de los puntos donde éste ha sido registrado (Crisci et al., 2000). A partir de la coincidencia de 2 o más trazos individuales, de taxones no relacionados filogenéticamente y con mecanismos de dispersión diferente, es posible visualizar patrones generales, conocidos como trazos generalizados o componentes bióticos (Humphries y Parenti, 1999), que representan biotas ancestrales fragmentadas por eventos geológicos o tectónicos (Craw, 1988). A todo este proceso se lo conoce como hipótesis de homología biogeográfica primaria (Morrone, 2001). La intersección de 2 o más de estos componentes da lugar a los nodos biogeográficos, áreas compuestas que permiten delinear límites biogeográficos (Craw, Grehan y Heads, 1999), derivadas de la convergencia de biotas con diferente origen espacial. Tal es el caso del nodo ubicado en América Central (Humphries y Parenti, 1999), y que constituye la zona de transición de las biotas neártica y neotropical. El análisis panbiogeográfico constituye una excelente herramienta para identificar áreas para su posible conservación (i.e., los nodos biogeográficos), al contener una alta riqueza de especies y una gran diversidad de relaciones filogenéticas y geográficas (Álvarez-Mondragón y Morrone, 2004).
Otros análisis que se incorporan en biogeografía para dar prioridad de conservación a las áreas son el análisis de complementariedad que permite identificar en un número reducido de áreas la máxima diversidad biológica posible y el análisis de diversidad filogenética que favorece el establecer prioridades entre los grupos de especies que se evalúan, de acuerdo con la máxima diversidad jerárquica representada, otorgando mayor importancia a los taxones con categorías superiores (Álvarez-Mondragón y Morrone, 2004; Torres-Miranda y Luna-Vega, 2006). Ambos criterios representan un requisito indispensable para la biología de la conservación.
Las macroalgas se dividen en 2 clases: macroalgas enanas (menores a 10 cm de longitud) y grandes macroalgas (mayores a 10 cm de longitud). Estas últimas, debido a su talla, dan lugar a hábitats biogénicos naturales y son refugio para una gran diversidad de organismos marinos (Connell, 2005). Además, representan materia prima sumamente importante en las industrias química (extracción de agar y carragenina), alimenticia, cosmética y papirogénica (Vilchis, 2015). A pesar de su importancia biológica y económica, dichos organismos no han sido objeto de estudios biogeográficos o de conservación en el país (Pedroche y Sentíes, 2003). Esto se debe en parte a que los criterios de conservación se han enfocado, principalmente, en la protección de áreas en las que se encuentran especies de vertebrados, invertebrados, pastos marinos y manglares; organismos conspicuos, de importancia económica y cultural, o por ser taxones catalogados en alguna categoría de riesgo (Escalante, 2003; Conabio-Conanp-TNC-Pronatura, 2007).
Dado lo anterior y tomando en cuenta que las divisiones que conforman al grupo de las macroalgas (Chlorophyta, Rhodophyta y Ochrophyta) no son un grupo natural, el objetivo del presente trabajo es identificar los patrones de distribución de las grandes macroalgas en el golfo de México y Caribe mexicano, a través de un análisis de parsimonia de endemismos (PAE) y uno de trazos, integrando un análisis de complementariedad y otro de diversidad filogenética, con la finalidad de encontrar áreas de importancia para la conservación de este grupo de organismos y que sean coincidentes con los sitios prioritarios para la conservación de la biodiversidad marina (SPCBM), propuestos por la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio) y la Comisión Nacional de Áreas Naturales Protegidas (Conanp), y de este modo, contribuir al conocimiento de este grupo de organismos y al entendimiento de su historia evolutiva.
Materiales y métodos
Para el PAE se trazó sobre un mapa del área de estudio una cuadrícula de 2° de latitud por 2° de longitud, utilizando el programa QGis v. 2.2.0 (Quantum GIS Development Team, 2014). En el mismo mapa se identificaron y se nombraron los cuadrantes que contenían registros de grandes macroalgas (fig. 1). Posteriormente, se elaboró una matriz de cuadrantes vs. especies, codificando con “1” la presencia y con “0” la ausencia de las especies en cada cuadrante, además se incorporó un área hipotética para el enraizamiento del árbol, consistente en ausencias para todos los taxones. Esta matriz se analizó con el algoritmo Nona (Goloboff, 1999) en la interface WinClada (Nixon, 2002) bajo una rutina de máxima parsimonia, usando la opción de búsqueda heurística y una estrategia TBR+TBR
Para el análisis de trazos se utilizaron las especies registradas en el área de estudio, exceptuando las que tuvieran una distribución amplia (i.e., que se distribuyan a lo largo de ambos litorales) o bien que están restringidas a un solo cuadrante. Se modificó la base de datos original utilizando OpenOffice 4 (hoja de cálculo), de tal manera que, sin eliminar información, solo quedara la entrada taxonómica y la georreferenciación. Esta información se exportó a formato dBaseIII, para posteriormente ser visualizadas en el sistema de información geográfica ArcView 3.2 (ESRI, 1999). Una vez obtenidas y cartografiadas las localidades puntuales de las 41 especies, se obtuvieron los trazos individuales, uniendo las localidades mediante el principio de distancia mínima, utilizando la extensión Trazos2004 (Rojas, 2004) en ArcView 3.2.; los archivos generados fueron transformados a formato Shapefile. Con la misma extensión, se construyeron los trazos generalizados, a partir de la congruencia de 2 o más trazos individuales, una vez delimitados se identificaron los nodos biogeo-
gráficos.
Se contabilizó el número total de especies contenidas en cada cuadrante del área de estudio, considerando también el número de órdenes, familias y géneros de grandes macroalgas (tabla 4), con la finalidad de conocer una primera aproximación de la diversidad de estos grupos en la región. Con base en la información anterior, se llevó a cabo un análisis de complementariedad, el cual permite encontrar pares de áreas que en conjunto alberguen el mayor número de especies posible, comparándolas unas con otras. Se utilizó el índice propuesto por Colwell y Coddington (1994), que cuantifica el grado de disimilitud en la composición de especies de las áreas que se comparan, mediante la siguiente formula:
ICC = (A+B-2j) / (A+B-j)
donde A: número de especies presentes en el área 1, B: número de especies presentes en el área 2, j: número de especies presentes en ambas áreas.
El valor del índice de complementariedad varía entre 0 y 1; si su valor es igual a 0, son áreas no complementarias, indicando que las áreas contienen las mismas especies. Cuando el valor es igual a 1 no tienen especies en común, es decir, son áreas que se complementarían entre sí por las especies que cada una presenta.
Con la intención de analizar si los esfuerzos actuales de conservación coinciden con la diversidad de grandes macroalgas, se superpusieron en un mapa los cuadrantes con mayor riqueza filogenética, los que presentaron especies exclusivas únicas, los nodos biogeográficos y las áreas con mayor complementariedad. Los cuadrantes donde 2 o más de estos criterios coincidieron, se compararon con el polígono de los sitios prioritarios para la conservación de la biodiversidad marina.
Resultados
El PAE produjo 2 cladogramas igualmente parsimoniosos, con una longitud de 309 pasos, un IC = 0.38 y un IR = 0.53. Se generó un consenso estricto, el cual tuvo una longitud de 311 pasos, un IC = 0.38 y IR = 0.53. En éste se reconocieron 9 grupos de áreas basados en la presencia de especies exclusivas (i.e., sinapomorfías) (fig. 2, tabla 1). Es posible observar un clado sustentado por 9 especies (clado III), el cual muestra una relación entre las siguientes áreas: centro de Campeche (Cam01), norte y sur de Tamaulipas (Tam01, Tam02), arrecife Alacranes (Yuc02), costa centro de Yucatán (Yuc03), norte de Veracruz (Ver01), Sistema Arrecifal Veracruzano (SAV), (Ver02, Ver03), norte y centro de Quintana Roo (QRoo01, QRoo02). De los 15 cuadrantes considerados en este análisis, 4 presentaron especies únicas restringidas a un solo cuadrante, es decir autapomorfías (tabla 2). Tal es el caso del clado VI que agrupa la región norte de la península de Yucatán (Yuc02, Yuc03) por la presencia de una sola especie (Sporochnus pedunculatus).
En el análisis de trazos se obtuvieron 40 trazos individuales, 1 por cada especie, cuya superposición resultó en 2 trazos generalizados, 1 para el golfo de México y otro para el Caribe mexicano (fig. 3a, b).
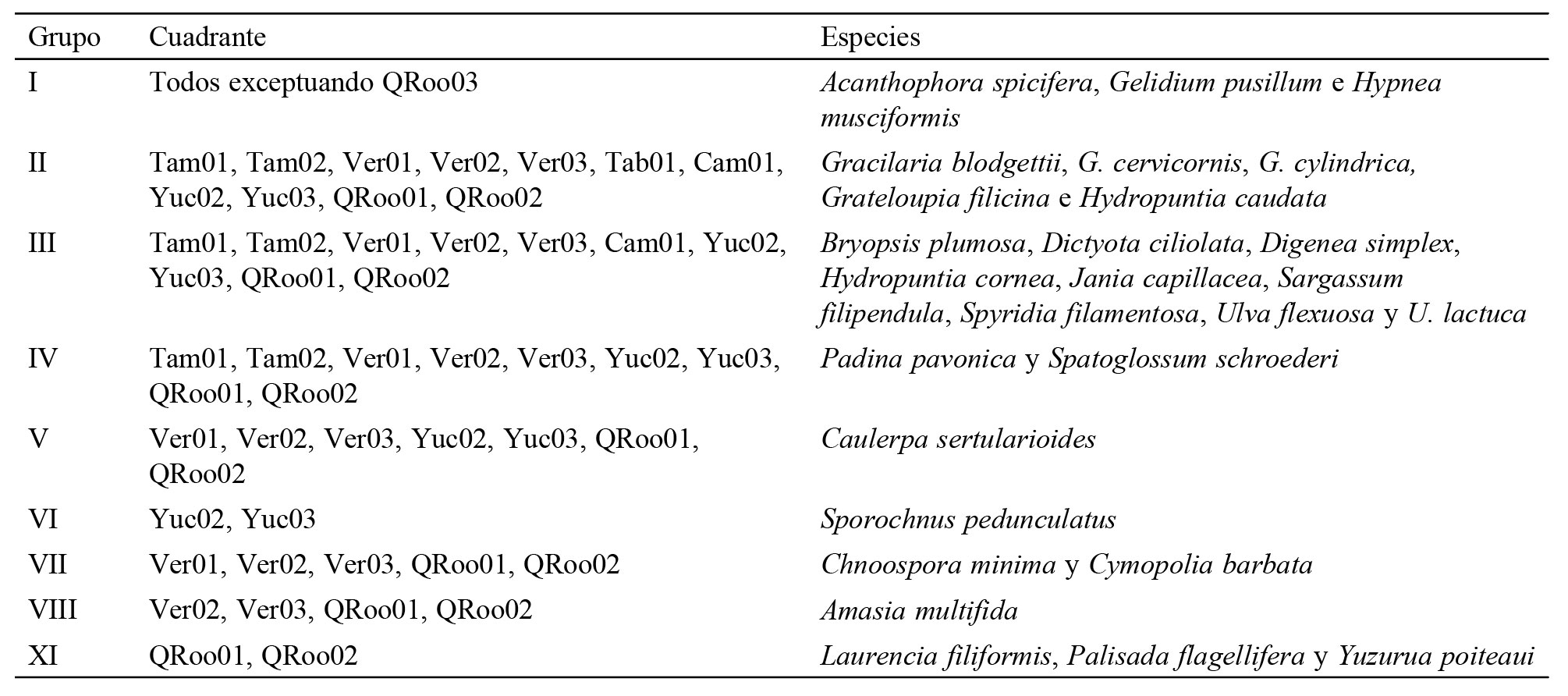
Trazo generalizado del golfo de México: abarca desde el norte de Tamaulipas al noroeste de la península de Yucatán. Construido con los trazos individuales de Caulerpa ashmeadii, Chondria atropurpurea, Cladophora laetevirens, Cymopolia barbata, Eucheuma isiforme, Gracilaria tikvahie, Hypnea valentiae, Osmundaria obtusiloba, Rosenvingea intricata, Solieria filiformis, Sporochnus pedunculatus, Ulva flexuosa y U. prolifera.
Trazo generalizado del Caribe mexicano: desde el noroeste de la península de Yucatán hasta el sur de Quintana Roo y está construido con los trazos individuales de Amphiroa tribulus, Caulerpa microphysa, Cladophora longicaulis, Codium decorticatum, Chondria atropurpurea, C. cnicophylla, Dictyopteris jamaicensis, D. justii, D. plagiogramma, D. polypodioides, Dictyota pinnatifida, Gracilaria cuneata, Halimeda incrassata, H. monile, H. simulans, Halymenia floresii, Hydropuntia crassissima, Laurencia filiformis, Palisada flagellifera, Penicillus dumetosus, Rhipocephalus oblongus, Sargassum acinarium, S. ramifolium, Udotea occidentalis, U. flabellum, U. dixonii y Yuzurua poiteaui.
Las especies de grandes macroalgas reportadas en este trabajo se encuentran distribuidas en 60 géneros, 30 familias y 17 órdenes. El cuadrante con mayor riqueza específica fue QRoo01, que corresponde al norte del estado de Quintana Roo, mientras que los que tuvieron menor número fueron QRoo03 y Yuc01 (tabla 4). De acuerdo con los porcentajes de las 4 jerarquías taxonómicas para cada cuadrante, los más altos corresponden al cuadrante QRoo01, seguido por Yuc02, QRoo02 y Ver02 (tabla 4).
El análisis de complementariedad identificó que los cuadrantes Tam02 y Yuc02 son los que mejor se complementan, al presentar un ICC de 0.81, lo que indica que comparten pocas especies, permitiendo reunir en este par de áreas un total de 85 especies de grandes macroalgas de las 118 presentes en el golfo de México y Caribe mexicano.

Prioridad de conservación de acuerdo a los 4 criterios establecidos en este trabajo y a las categorías de importancia de los sitios prioritarios para la conservación de la biodiversidad marina (SPCBM) propuestos por Conabio-Conanp-TNC-Pronatura, (2007).
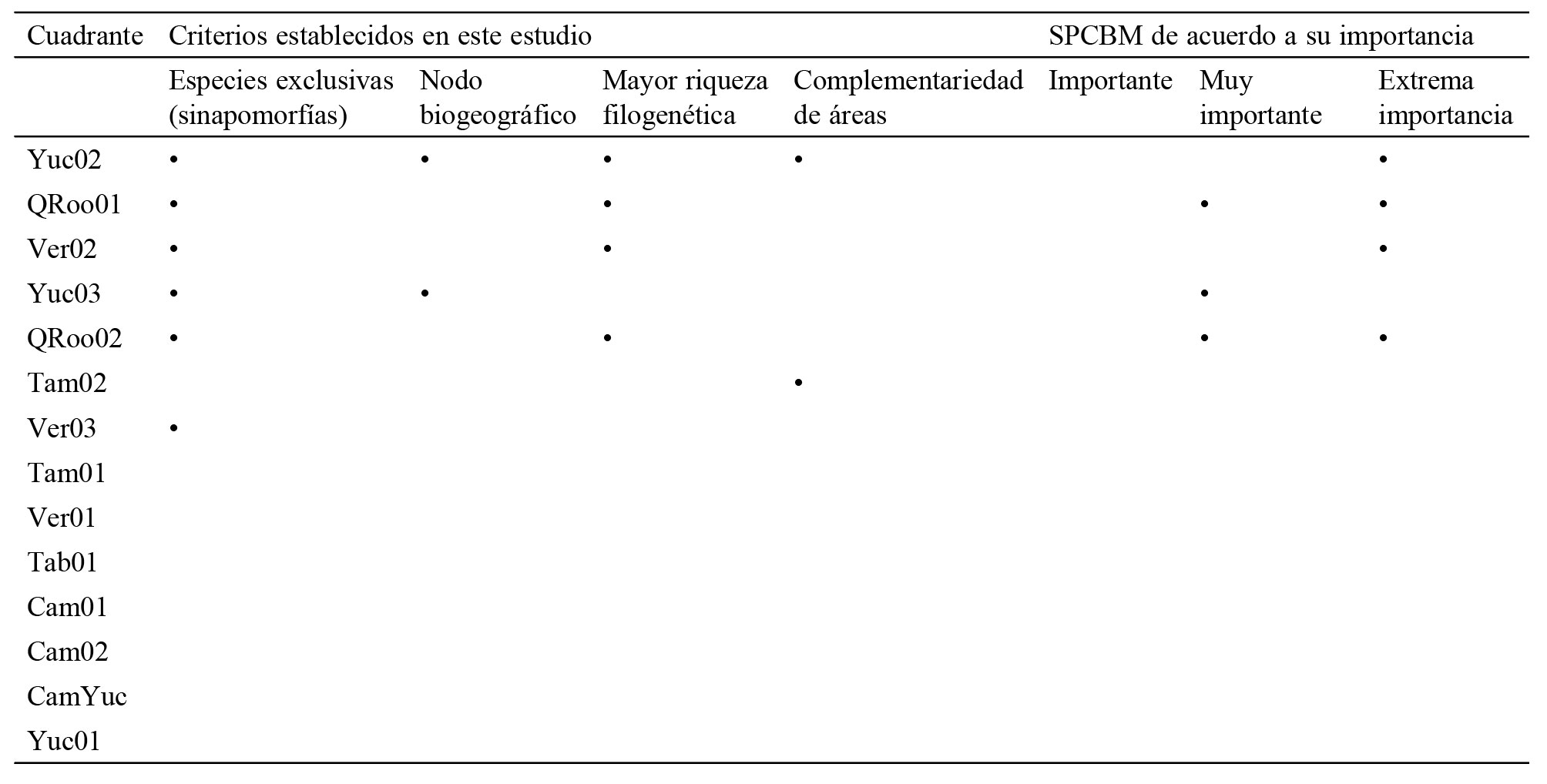
De acuerdo con las categorías de conservación propuestas por Conabio-Conanp-TNC-Pronatura (2007) para los sitios prioritarios para la conservación de la biodiversidad marina (SPCBM), arrecife Alacranes y el norte del SAV coinciden con 2 de “extrema importancia (EI)” (fig. 4). Mientras que el norte de Quintana Roo coincide con un sitio de “EI” y 3 de categoría “muy importante (MI)”, el centro de Quintana Roo con 2 de “EI” y uno “MI”. Finalmente, el centro de Yucatán coincide con un sitio catalogado como “MI” para la conservación marina (fig. 4).
Diversidad de cada uno de los cuadrantes analizados.
http://rev.mex.biodivers.unam.mx/wp-content/uploads/2018/vol-89/89-1-mar-2018/html/16-Fig4.tif
Figura 4. Coincidencia de los sitios prioritarios para la conservación de la biodiversidad marina (SPCBM) propuestos por Conabio-Conanp-TNC-Pronatura, (2007), con los cuadrantes que presentaron la sobreposición de 2 o más de los criterios establecidos en el presente estudio. A: norte del Sistema Arrecifal Veracruzano; B: arrecife Alacranes; C: centro de Yucatán, D: norte de Quintana Roo; E: centro de Quintana Roo. En un gradiente de colores se muestran los SPCBM de acuerdo a su importancia.
Discusión
Dichos patrones de distribución han permitido, junto con los análisis de complementariedad y diversidad filogenética, establecer 4 criterios para la identificación de áreas de conservación marinas considerando la distribución de las grandes macroalgas (tabla 3). A partir de éstos se señalan 5 áreas prioritarias para la conservación de estos grupos, principalmente el arrecife Alacranes en Yucatán, ya que representa un área de complejidad biótica y geológica, reflejada en la sobreposición de los 4 criterios. Aunque el análisis de complementariedad indique que el arrecife Alacranes y el sur de Tamaulipas son las áreas que en conjunto albergan el mayor número de especies (85), si a su vez se complementan con el norte del estado de Quintana Roo, se estarían conservando 110 de las 118 especies analizadas en este trabajo.
Aunque hasta el momento no hay información con respecto a especies de macroalgas en alguna categoría de riesgo nacional, se sabe que las grandes macroalgas constituyen un elemento fundamental en la estructura de las comunidades inframareales, al establecer ambientes biogénicos que contribuyen a la heterogeneidad de hábitats, permitiendo así el establecimiento, sobre ellas, de una biota rica en representantes de varios phyla, tanto procariontes como eucariontes (uni o multicelulares). Además, sirven como refugio para muchos animales (Cancino y Santelices, 1984; Connell, 2005), lo que permite la coexistencia de un número elevado de especies que da lugar a comunidades altamente diversas, con composiciones bióticas complejas. Esto se refleja de manera indirecta en la coincidencia de los resultados obtenidos en este trabajo, tanto con los SPCBM propuestos por Conabio-Conanp-TNC-Pronatura (2007), pero también con los resultados de riqueza y diversidad de otros organismos como los mencionados previamente.
Si bien es cierto que en este estudio no se consideró el intervalo de distribución completo de las especies de grandes macroalgas, es decir su distribución fuera de México, fue posible definir patrones de distribución y ofrecer algunas hipótesis sobre los procesos que los originaron, además de reconocer congruencias con los patrones de distribución de otros organismos. Sin embargo, dichas hipótesis deberán ser corroboradas continuando con el estudio de la distribución de las macroalgas en diversos contextos. En particular, es necesario llevar a cabo estudios histórico-evolutivos, filogenéticos y ecológicos con estos grupos para generar más información que permita confirmar hipótesis de homología biogeográfica primaria desde el punto de vista de la biogeografía evolutiva, entendiendo por homología biogeográfica primaria ese primer esbozo de relaciones históricas entre áreas, y que representa solo el primer paso dentro de un estudio de esta índole (Morrone, 2009).
Agradecimientos
Referencias
Álvarez-Mondragón, E. y Morrone, J. J. (2004). Propuesta de áreas para la conservación de aves de México, empleando herramientas panbiogeográficas e índices de complementariedad. Interciencia, 29, 112–120.
Arriaga-Cabrera, L., Vázquez-Domínguez, E., González-Cano, J., Jiménez-Rosenberg, R., Muñoz-López, E. y Aguilar-Sierra, V. (1998). Regiones marinas prioritarias de México. Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Bird, D. E., Burke, K., Hall, S. A. y Casey, J. F. (2011). Tectonic evolution of the Gulf of Mexico Basin. En J. W. Tunnell, D. L. Felder y S. A. Earle (Eds.), Gulf of Mexico. Origin, waters, and biota. Volumen 3, Geology. Texas: Texas A&M University Press.
Cancino, J. y Santelices, B. (1984). Importancia ecológica de los discos adhesivos de Lessonia nigrescens Bory (Phaeophyta) en Chile central. Revista Chilena de Historia Natural, 57, 23–33.
Castillo-Rodríguez, Z. G. (2014). Biodiversidad de moluscos marinos en México. Revista Mexicana de Biodiversidad, 85, 419–430.
Chávez-Hidalgo, A. (2009). Conectividad de los arrecifes coralinos del golfo de México y Caribe mexicano (Tesis de maestría). Centro Interdisciplinario de Ciencias Marinas, Instituto Politécnico Nacional, Ciudad de México.
Colwell, R. K. y Coddington, J. A. (1994). Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society of London (Series B), 345, 101–118.
Conabio-Conanp-TNC-Pronatura, (2007). Análisis de vacíos y omisiones en conservación de la biodiversidad marina de México: océanos, costas e islas. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Ciudad de México: Comisión Nacional de Áreas Naturales Protegidas, The Nature Conservancy-Programa México, Pronatura, A.C.
Connell, S. D. (2005). Assembly and maintenance of subtidal hábitat heterogeneity: synergistic effects of light penetration and sedimentation. Marine Ecology Progress Series, 289, 53–61.
Craw, R. C. (1988). Panbiogeography: methods and synthesis in biogeography. En A. A. Myers y P. S. Giller (Eds.), Analytical biogeography (pp. 405–435). Londres: Chapman and Hall.
Craw, R. C., Grehan, J. R. y Heads, M. J. (1999). Panbiogeography: tracking the history of life. New York: Oxford University Press.
Crisci, J. V., Katinas, L. y Posadas, P. (2000). Introducción a la teoría y práctica de la biogeografía histórica. Buenos Aires, Argentina: Sociedad Argentina de Botánica.
Dreckmann, K. M. (1998). Clasificación y nomenclatura de las macroalgas marinas bentónicas del Atlántico mexicano. Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Escalante, T. (2003). Determinación de prioridades en las áreas de conservación para los mamíferos terrestres de México, empleando criterios biogeográficos. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 74, 211–237.
Escalante, T. (2009). Un ensayo sobre regionalización biogeográfica. Revista Mexicana de Biodiversidad, 80, 551–560.
Escobar, E. (2004). Estado del conocimiento de las comunidades bentónicas en el golfo de México. En M. Caso, I. Pisanty y E. Ezcurra (Comps.), Diagnóstico ambiental del golfo de México, Vol. I (pp. 201–246). Ciudad de México: Instituto Nacional de Ecología.
ESRI (Environmental Systems Research Institute). (1999). ArcView 3.2 GIS. New York: Environmental Systems Research Institute, Inc.
Floeter, S. R., Rocha, L. A., Robertson, D. R., Joyeux, J. C., Smith-Vaniz, W. F., Wirtz, P. et al. (2008). Atlantic reef fish biogeography and evolution. Journal of Biogeography, 35, 22–47.
Goloboff, P. (1999). NONA, versión 2.0. San Miguel de Tucumán: Publicado por el autor.
González, M. A., Chávez, E. A., De la Cruz, G. y Torruco, D. (1991). Distribution patterns of gastropods and bivalves at the Yucatán Peninsula, Mexico. Ciencias Marinas, 17, 147–172.
Granados-Barba, A., Solís-Weiss, V., Tovar-Hernández, M. A. y Ochoa-Rivera, V. (2003). Distribution and diversity of the Syllidae (Annelida: Polychaeta) from the Mexican Gulf of Mexico and Caribbean. Hydrobiologia, 496, 337–345.
Guiry, M. D. y Guiry, G. M. (2016). AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. Recuperado el 10 Agosto, 2015 de: http://www.algaebase.org
Humphries, C. J. y Parenti, L. R. (1999). Cladistic biogeography: interpreting patterns of plant and animal distributions. Oxford: Oxford University Press.
Littler, D. S. y Littler, M. M. (2000). Caribbean reef plants. An identification guide to the reef plants of the Caribbean, Bahamas, Florida and Gulf of Mexico. Washington, D.C.: OffShore Graphics, Inc.
Morrone, J. J. (2001). Homology, biogeography and areas of endemism. Diversity and Distributions, 7, 297–300.
Morrone, J. J. (2004). Panbiogeografìa, componentes bióticos y zonas de transición. Revista Brasileira de Entomologia, 48, 149–162.
Morrone, J. J. (2005). Sistemática, biogeografía y evolución. Ciudad de México: Las Prensas de Ciencias, Facultad de Ciencias, UNAM.
Morrone, J. J. (2009). Evolutionary biogeography: an integrative approach with case studies. Nueva York: Columbia Uni-
versity Press.
Nihei, S. S. (2006). Misconceptions about parsimony analysis of endemicity. Journal of Biogeography, 33, 2099–2106.
Nixon, K. C. (2002). WinClada, ver 1.00.08. Ithaca. Recuperado el 05 mayo, 2014 de: www.cladistics.com/about_winc
Núñez, M. L., Zuccarello, G. C., Dreckmann, K. M. y Sentíes, A. (2016). Phylogeography of Hydropuntia cornea/ Hydropuntia usneoides complex (Gracilariales, Rhodophyta) in the Yucatán Peninsula. Phycologia, 56, 14–20.
Ortega, M. M., Godínez, J. L. y Garduño-Solórzano, G. (2001). Catálogos de algas bénticas de las costas mexicanas del golfo de México y mar Caribe. Ciudad de México: UNAM, Conabio.
Pedroche, F. F. y Sentíes, A. (2003). Ficología marina mexicana. Diversidad y problemática actual. Hidrobiológica, 13, 23–32.
Pindell, J. L., Cande, S. C., Pitman, W. C., Rowley, D. B., Dewey, J. F., Labrecque, J. et al. (1988). A plate-kinematic framework for models of Caribbean evolution. Tectonophysics, 155, 121–138.
Posadas, P. y Miranda-Esquivel, D. R. (1999). El PAE (Parsimony Analysis of Endemicity) como una herramienta en la evaluación de la biodiversidad. Revista Chilena de Historia Natural, 72, 539–546.
Quantum GIS Development Team. (2014). Quantum GIS Geographic Information System. Open Source Geospatial Foundation Project B.
Rojas, C. A. (2004). Automatización del método de la Panbiogeografía: identificación de centro de diversidad del Parque Nacional Iztacíhuatl, Popocatépetl y anexas (Tesis de Maestría). Universidad Nacional Autónoma de México, Ciudad de México.
Tapia-Silva, F. O., Hernández-Cervantes, O. E., Vilchis-Alfaro, M. I., Sentíes, A. y Dreckmann, K. M. (2015). Mapping of algae richness using spatial data interpolation. The International Archives of the Photogrammetry, Remote Sensing and Spatial Sciences, 40, 1005–1008.
Torres-Miranda, A. y Luna-Vega, I. (2006). Análisis de trazos para establecer áreas de conservación en la Faja Volcánica Transmexicana. Interciencia, 31, 849–855.
Vilchis, M. I. (2015). Biogeografía de las grandes macroalgas del golfo de México y Caribe mexicano (Tesis de Maestría). Universidad Autónoma Metropolitana, Ciudad de México.
Wynne, M. J. (2011). A checklist of benthic marine algae of the tropical and subtropical western Atlantic: third revision. Nova Hedwigia Beihefte, 140, 7–166.


