Helmintos del pez anual Millerichthys robustus (Teleostei: Rivulidae), una especie endémica de México
Miguel Ángel Mosqueda-Cabrera a, *, José Ángel Labastida-Valerio a,
Andrés Moctezuma Sotelo-Viveros a, Rosa Esmeralda Becerra-García a
y María Isabel Jiménez-García b
a Departamento El Hombre y su Ambiente, Universidad Autónoma Metropolitana Unidad Xochimilco, Calzada del Hueso Núm. 1100, Col. Villa Quietud, Delegación Coyoacán, 04960 Ciudad de México, México
b Laboratorio de Parasitología y Sanidad Acuícola, División de Estudios de Posgrado e Investigación, Instituto Tecnológico de Boca del Río,
Km. 12 carretera Veracruz-Córdoba, 94290 Boca del Río, Veracruz, México
*Autor para correspondencia: zitzitl@correo.xoc.uam.mx (M.A. Mosqueda-Cabrera)
Resumen
Los peces anuales habitan cuerpos de agua temporales que se forman durante la estación de lluvias, que cuando se secan conducen a la muerte de toda la población. En México, Millerichthys robustus es la única especie con historia de vida anual y con escasa información sobre sus metazoarios parásitos. La helmintofauna de M. robustus está formada por las fases larvarias de las especies alogénicas Valipora minuta, V. campylancristrota, Clinostomum marginatum, Contracaecum sp., Serpinema trispinosum, Eustrongylides sp., Gnathostoma lamothei y Polymorphus sp. Los parámetros de la infección se relacionan con la temporalidad del hábitat; estos valores, en la mayoría de las especies, son superiores a los registrados en peces de cuerpos de agua permanentes. La metacercaria de C. marginatum, con prevalencias de 16.7% en testículo y 4.1% en ovario, puede impactar la reproducción y esperanza de vida del hospedero. El número de especies encontradas (8) y los valores altos de los parámetros de infección, junto con las características físicas del hábitat, juegan un papel importante que asegura la transmisión de la infección hacia los hospederos definitivos. Estos hábitats pueden ser un laboratorio natural para el estudio del sistema parásito-hospedero, por ejemplo, en lo referente a patrones de colonización y desarrollo larvario.
Palabras clave:
Metazoarios parásitos; Peces anuales; Cuerpos de agua temporales
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Helminth of the annual fish Millerichthys robustus (Teleostei: Rivulidae), an endemic species to Mexico
Abstract
Annual fishes inhabit temporary water bodies that form during the rainy season, when they dry up, lead to death the entire population. In Mexico, Millerichthys robustus is the only species with an annual life history, with scarce information about its metazoan parasites. Helminth fauna of M. robustus is constituted by the larval stages of allogenic species such as Valipora minuta, V. campylancristrota, Clinostomum marginatum, Contracaecum sp., Serpinema trispinosum, Eustrongylides sp., Gnathostoma lamothei and Polymorphus sp. The parameters of infection are related with temporality of habitat, and the values for most of species are higher than those registered in fish of permanent waters. The metacercaria of C. marginatum, with prevalences of 16.7% in testicle and 4.1% in ovary, can impact the reproduction and life expectancy of host. The number of species found (8) and the high values in infection parameters, together with the physical characteristics of habitat, play an important role that ensures the transmission of infection toward the definitive hosts. These habitats can be a natural laboratory for the study of the parasite-host system, for example, in relation to patterns of colonization and larval development.
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Keywords:
Metazoan parasites; Annual fishes; Temporary water bodies
Introducción
Algunas especies de parásitos, en particular los que requieren el paso por las cadenas tróficas, han sido mencionadas como bioingenieros o constructores de las relaciones tróficas; sus ciclos de vida complejos poseen un gran valor ecológico para comprender el funcionamiento de un ecosistema (Goater et al., 2013; Marcogliese y Cone, 1997). En México, el conocimiento acerca de la helmintofauna de peces dulceacuícolas es amplio, se ha incluido en temas de conservación (Falcón-Ordaz et al., 2015; Monks et al., 2013; Pulido-Flores et al., 2005), biogeografía (Aguilar-Aguilar et al., 2005, 2008; Salgado-Maldonado y Quiroz-Martínez, 2013), y un gran número de listados de especies (Aguilar-Aguilar et al., 2004, 2014; Aguilar-Aguilar y Salgado-Maldonado, 2006; Caspeta-Mandujano, 2005, 2010; Pérez-Ponce de León et al., 2009, 2010; Salgado-Maldonado, 2006; Salgado-Maldonado, Cabañas-Carranza, Caspeta-Mandujano Salgado-Maldonado et al., 2001; Cabañas-Carranza, Soto-Galera et al., 2001; Salgado-Maldonado, Cabañas-Carranza et al., 2004; Salgado-Maldonado, Mercado-Silva et al., 2004; Salgado-Maldonado et al., 2005, 2011, 2016; Scholz et al., 1996; Vidal-Martínez et al., 2001). A pesar del considerable esfuerzo de muestreo en estudios taxonómicos sobre helmintos de peces dulceacuícolas en el país, a la fecha, los peces anuales no han sido estudiados. En Latinoamérica, pocos son los estudios sobre helmintos en peces anuales, con registros en Uruguay (Delgado, 2014; Delgado y García, 2015), Venezuela (Nico y Thomerson, 1989) y México (Mosqueda-Cabrera y Ocampo-Jaimes, 2009). Los peces anuales habitan cuerpos de agua temporales que se inundan por semanas o meses durante la estación de lluvias, siguiendo los ciclos estacionales y transitando obligadamente por una fase seca (Myers, 1952; Wourms, 1972a, b). Durante el hidroperíodo (tiempo en que el hábitat presenta agua) los peces tienen un acelerado crecimiento y reproducción, y toda la población muere al secarse el estanque. En la siguiente estación de lluvias una nueva población tiene lugar a partir de los huevos que resisten la sequía mediante la detención del desarrollo conocido como diapausas embrionarias (Dolfi et al., 2014; Furness, 2016; Podrabsky et al., 2010). En México el conocimiento de peces con historia de vida anual es reciente (Domínguez-Castañedo et al., 2013). Millerichthys robustus es una especie endémica de las cuencas de los ríos Coatzacoalcos y Papaloapan, este último catalogado como uno de los ríos con mayor riqueza y endemismos de helmintos en el país (Salgado-Maldonado et al., 2005). La información sobre los helmintos ha permitido conocer la manera en que la asociación parásito-hospedero se establece en cuerpos de agua permanentes (Pérez-Ponce de León et al., 2010); no obstante, a pesar de que en cuerpos de agua temporales los procesos de especiación y coevolución entre las especies y su fauna parasitaria se potencializan (Delgado y García, 2015), existe poca información acerca de las relaciones parásito-hospedero en estos hábitats. Esta investigación presenta la helmintofauna de la única especie de pez anual distribuida en México (M. robustus) y discute brevemente el riesgo que representa sobre sus poblaciones en cuerpos de agua temporales de la parte baja de la cuenca del río Papaloapan.
Materiales y métodos
La recolecta de peces se realizó durante 2 momentos de un hidroperíodo (al inicio, en octubre de 2015 y al final, en febrero de 2016) en 2 cuerpos de agua temporales asociados a la cuenca del río Papaloapan, en las inmediaciones de Playa Salinas (PS-1) y Tlacotalpan (TL-1). Los peces fueron capturados durante la fase de reproducción utilizando redes de cuchara con luz de malla de 1/8”. Al finalizar la recolecta, inmediatamente fueron sacrificados, fijados en formaldehído al 10% durante 10 días y posteriormente conservados en alcohol etílico al 70% hasta su estudio. Cuando fue posible, la búsqueda de helmintos se realizó en peces recién sacrificados y disectados bajo un microscopio estereoscópico. Los helmintos fueron mantenidos en solución salina al 0.65% hasta que fueron fijados de acuerdo a cada grupo taxonómico: metacercarias y metacéstodos con formaldehído al 4% caliente; para la observación de los ganchos de estos últimos, se utilizó solución GAP (glicerina y ácido pícrico). Las metacercarias y cistacantos fueron teñidos con paracarmín de Mayer y montadas en resina sintética; larvas de nemátodos con alcohol etílico al 70% caliente y transparentados con lactofenol de Amman. La recolecta de peces fue amparada bajo los permisos de colecta científica (SPGA/DGVS/04147/15, 05529/16) y se identificaron de acuerdo con Costa (1995) y Miller y Hubbs (1974).
Los descriptores cuantitativos de la infección fueron calculados de acuerdo con Bush et al. (1997) y el intervalo de confianza (95%), de acuerdo con el método exacto de Fisher. Los parásitos fueron identificados de acuerdo con Caffara et al. (2011), García-Prieto et al. (2010), Gaspar-Navarro et al. (2013), Moravec (1998), Scholz y Salgado-Maldonado (2001). Los ejemplares fueron primeramente observados con un microscopio estereoscópico y fotografiados con una cámara digital adaptada a un microscopio compuesto. Algunos peces fueron depositados en la Colección Nacional de Peces, Instituto de Biología, Universidad Nacional Autónoma de México (22560, 22561, 22562 y 22563) y los helmintos de referencia, en la Colección Nacional de Helmintos (CNHE) del Instituto de Biología, UNAM.
Resultados
La disección de 48 peces procedentes de 2 hábitats (tabla 1) resultó en el hallazgo de 8 especies de helmintos; 7 en peces de PS-1 y 4 de TL-1. Al inicio del hidroperíodo, solo 3 especies de helmintos se encontraron en ambos hábitats: Valipora minuta, Clinostomum marginatum y Serpinema trispinosum; mientras que al final, a las especies anteriores, se sumaron Eustrongylides sp., Gnathostoma lamothei, Contracaecum sp., Valipora campylancristrota y Polymorphus sp. El sitio de infección de todas las especies fue el mismo a lo largo del hidroperíodo, excepto para las larvas de S. trispinosum; al inicio del hidroperíodo se encontraron libres a lo largo del intestino mientras que al final se localizaron enquistados sobre la serosa externa del recto. Por otra parte, las metacercarias de C. marginatum, además de encontrarse en el celoma, también infectaron las gónadas (fig. 1) con una prevalencia de 20.8% (16.7% en testículo y 4.1% en ovario) al final del hidroperíodo, únicamente en peces de PS-1 (tabla 2).
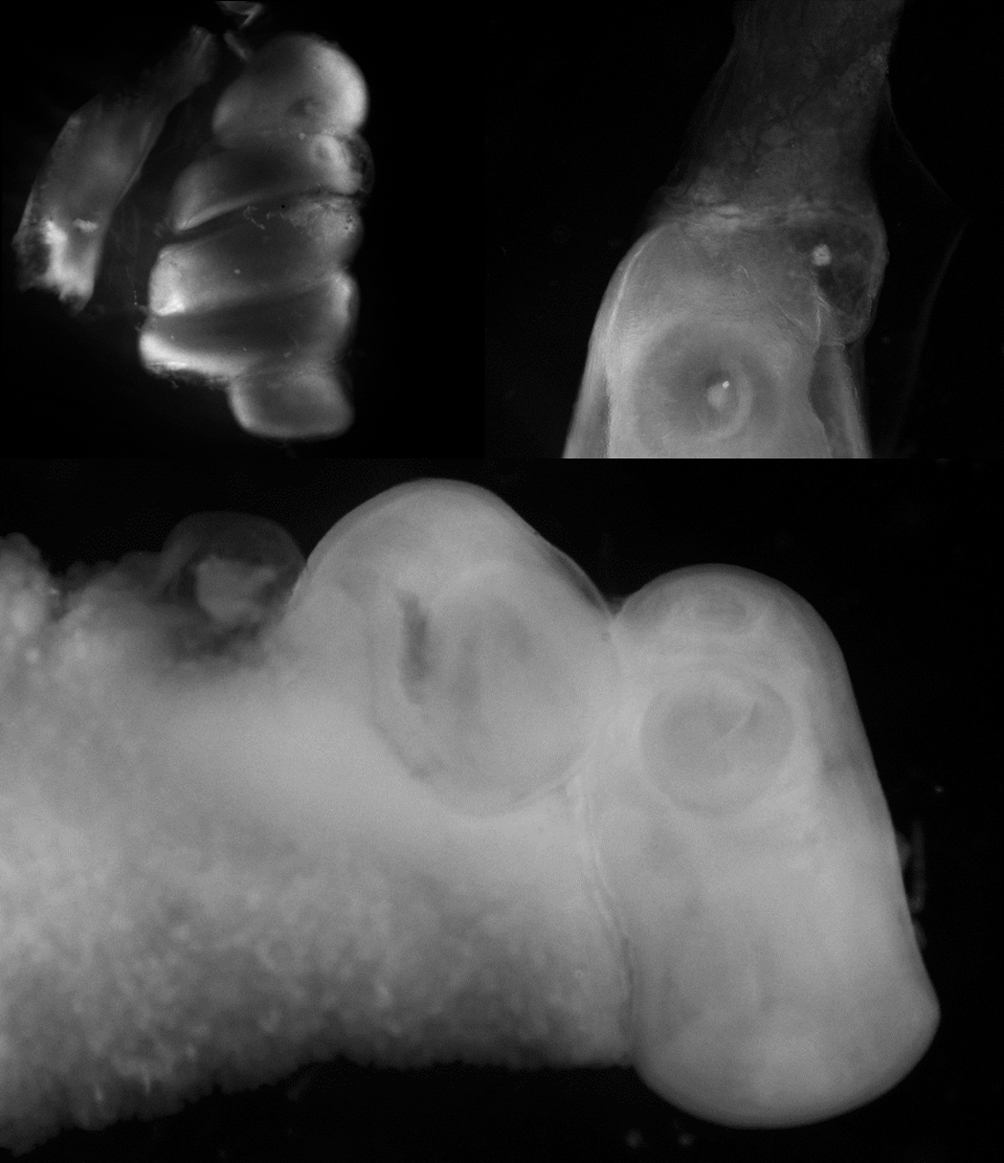
Tabla 1
Total de peces recolectados en 2 cuerpos de agua temporales durante 2 momentos del hidroperíodo. He = Hospederos examinados, M = macho, H = hembra, n = total de peces examinados.
|
Hábitat |
Hidroperíodo |
n |
|
|
Inicio He (M, H) |
Final He (M, H) |
||
|
PS-1 (18°54’48” N, 95°56’56” O) |
9 (9, 0) |
15 (4, 11) |
24 |
|
TL-1 (18°37’06” N, 95°38’08” O) |
10 (10, 0) |
14 (0, 14) |
24 |
|
Total |
19 (19, 0) |
29 (4, 25) |
48 |
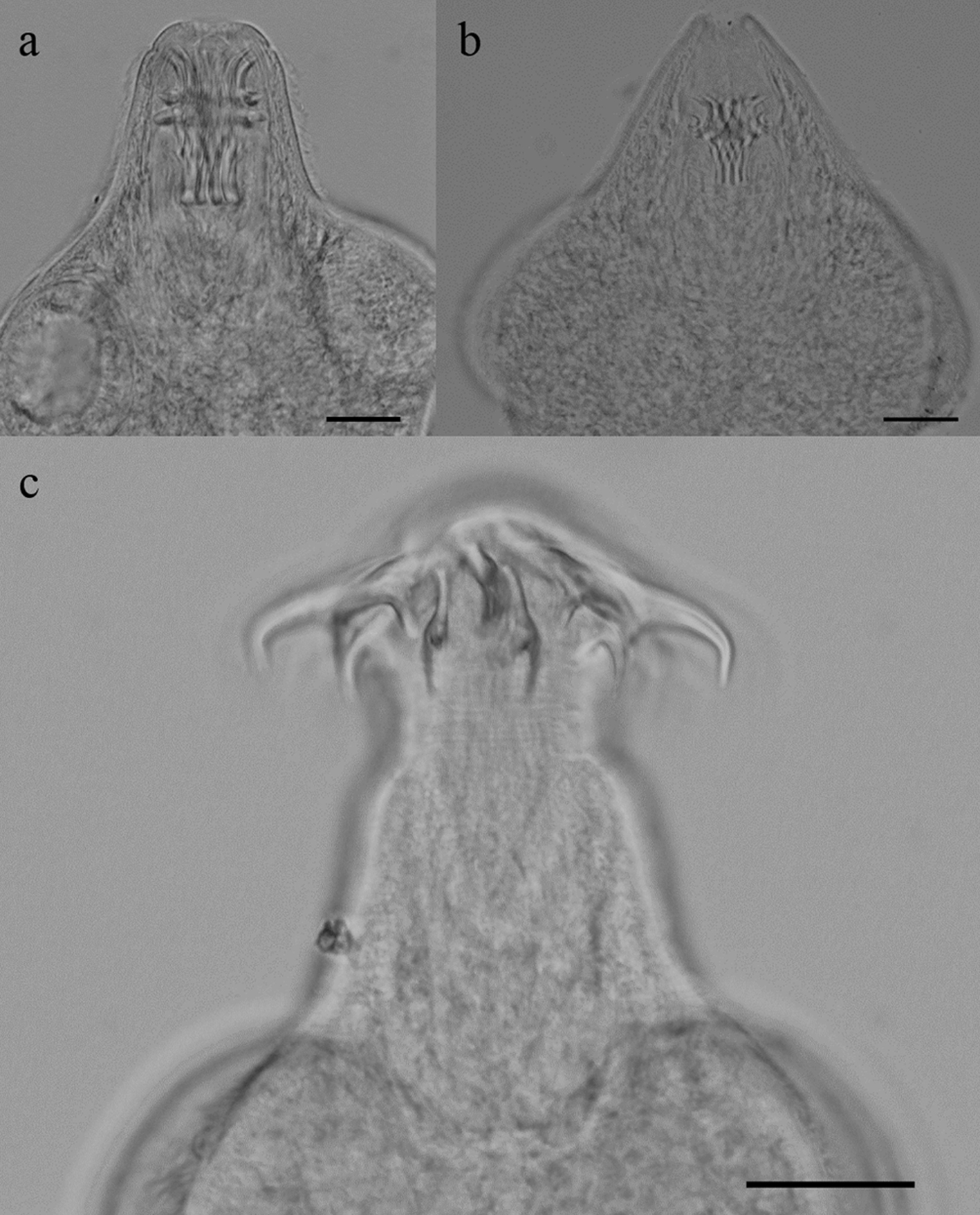
Todas las especies de helmintos fueron encontradas en sus fases larvarias; los nemátodos conformaron el grupo mejor representado, con 4 especies Contracaecum sp., G. lamothei, S. trispinosum (fig. 2) y Eustrongylides sp.; excepto para Eustrongylides sp. y Polymorphus sp., todas las especies de helmintos presentaron prevalencias mayores de 20%. El metacéstodo V. campylancristrota fue registrado solo al final del hidroperíodo en PS-1 (fig. 3) y 3 peces presentaron infección mixta con V. minuta. Esta última especie presentó las prevalencias más altas y junto con S. trispinosum, sus valores se incrementaron al final del hidroperíodo en peces de TL-1 (tabla 2).

Tabla 2
Parámetros cuantitativos de la infección de 8 especies de helmintos aislados de Millerichthys robustus recolectado en 2 cuerpos de agua temporales. S = Sitio de infección, Vb = vesícula biliar, Ce = celoma, G = gónadas, H = hígado, I = intestino, M = musculatura, P = prevalencia, i = inicio del hidroperíodo, f = final del hidroperíodo, IC = intervalo de confianza, I = intensidad promedio, A = abundancia promedio
|
Especie de helminto |
S |
Hábitat (Fase del hidroperíodo) |
P (IC) 95% |
I |
A |
|
Metacestodos |
|||||
|
Gryporhynchidae |
|||||
|
Valipora minuta |
Vb Vb Vb Vb |
PS-1(i) PS-1(f) TL-1(i) TL-1(f) |
44.44 (13.70-78.80) 20.00 (04.33-48.09) 60.00 (26.24-87.84) 92.85 (66.13-99.82) |
1.0 1.0 1.8 2.7 |
0.4 0.2 1.1 2.5 |
|
V. campylancristrota |
Vb |
PS-1(f) |
26.67 (07.79-55.10) |
1.5 |
0.4 |
|
Metacercaria Clinostomidae |
|||||
|
Clinostomum marginatum |
Ce Ce, G Ce |
PS-1(i) PS-1(f) TL-1(i) |
33.33 (07.48-70.07) 33.33 (11.82-61.62) 20.00 (02.52-55.61) |
3.3 1.4 1.5 |
1.1 0.5 0.3 |
|
Larvas de nemátodos |
|||||
|
Anisakidae |
|||||
|
Contracaecum sp. |
H, Ce |
PS-1(f) |
40.00 (16.34-67.71) |
1.7 |
0.7 |
|
Camallanidae |
|||||
|
Serpinema trispinosum |
I I Ce |
PS-1(i) TL-1(i) TL-1(f) |
33.33 (7.49-70.07) 10.00 (0.25-44.50) 28.57 (08.40-58.10) |
1.0 2.0 1.0 |
0.3 0.2 0.3 |
|
Dioctophymatidae |
|||||
|
Eustrongylides sp. |
Ce |
PS-1(f) |
6.67 (0.17-31.95) |
6.0 |
0.4 |
|
Gnathostomatidae Gnathostoma lamothei |
M |
TL-1(f) |
26.67 (7.79-55.10) |
0.8 |
0.2 |
|
Cistacanto |
|||||
|
Polymorphidae |
|||||
|
Polymorphus sp. |
Ce |
PS-1(f) |
6.67 (0.17-31.95) |
1.0 |
0.07 |
Todas las especies de helmintos presentan ciclos de vida heteroxénicos y utilizan a M. robustus como hospedero intermediario. El hecho de haber encontrado los ovarios infectados con metacercarias de C. marginatum, en algunos casos ocupando la totalidad del órgano, sin la observación de ovocitos maduros, supone un efecto negativo sobre su reproducción y/o sobrevivencia (fig. 1). En el caso de los machos, el impacto puede estar reflejado al final del hidroperíodo, en el que la cantidad de machos fue menor con relación a las hembras (tabla 1).
Discusión
México es uno de los países con mayor número de estudios dirigidos al conocimiento de la helmintofauna de peces dulceacuícolas en Latinoamérica (Luque y Poulin, 2007). Este grupo de hospederos presenta una capacidad limitada en su distribución, resultado de la historia geológica de sus hábitats; no obstante, muchas especies de helmintos cumplen sus ciclos de vida en peces (Salgado-Maldonado y Quiroz-Martínez, 2013). En contraste, todas las especies de helmintos aquí documentadas constituyen especies alogénicas (Esch et al., 1988), es decir, durante su transmisión, M. robustus funge como hospedero intermediario hacia reptiles (S. trispinosum), aves (V. campylancristrota, V. minuta, C. marginatum, Eustrongylides sp., Contracaecum sp.) y mamíferos (G. lamothei), sus hospederos definitivos.
La ausencia de helmintos adultos en M. robustus está asociada principalmente a la efimeridad del hábitat. En términos generales, los peces anuales presentan una esperanza de vida corta (Blažek et al., 2013; Furness, 2016) e inevitablemente la muerte de todas las formas de vida acuática ocurre durante la sequía. En este escenario, los ciclos de vida de las especies de helmintos solo pueden ser completados en hospederos externos al cuerpo de agua, ya que una hipotética población de formas adultas de helmintos infectando a M. robustus, las cuales liberarían huevos y larvas que parasitarían a microcrustáceos y caracoles (primeros hospederos intermediarios) también moriría al final del hidroperíodo. No obstante, la presencia de helmintos en su fase adulta en M. robustus podría ocurrir bajo el siguiente supuesto. Durante las lluvias torrenciales, es posible que peces de cuerpos de agua permanentes infectados por un parásito adulto (p. ej. Bothriocephalus) lleguen a cuerpos de agua temporales a través de las escorrentías. De esta manera, M. robustus podría adquirir la infección al alimentarse de copépodos infectados y actuar como hospedero definitivo. No obstante esta situación dependería exclusivamente de este escenario.
En comparación con los cuerpos de aguas permanentes (p. ej. ríos, lagos y lagunas), los cuerpos de agua temporales, hábitat típico de peces anuales, presentan una fase seca obligatoria en un ciclo anual (Furness, 2016). Durante el hidroperíodo, los cuerpos de agua temporales son el hábitat común de crustáceos, insectos, moluscos y peces, entre otros; también de anfibios, reptiles y aves que llegan por razones de alimentación y reproducción. Sin embargo, solo las especies que presentan formas de resistencia a la sequía del hábitat darán lugar a nuevas poblaciones en el siguiente hidroperíodo (Williams, 2006). Las poblaciones de M. robustus mueren al final del hidroperíodo, cuando sus estanques se secan por completo; una población nueva tiene lugar en el siguiente hidroperíodo, al inicio de la estación de lluvias con la eclosión de huevos fertilizados por la generación anterior, que resistieron la temporada seca mediante diapausas embrionarias. De tal manera que cuando los comparamos con sus contrapartes — peces de ríos, lagos y lagunas—, los peces de cuerpos de agua temporales se encuentran aislados y presentan una vagilidad muy reducida. En este contexto, al inicio del hidroperíodo, el proceso de infección entre parásitos y los hospederos intermediarios depende de la visita recurrente de hospederos definitivos infectados (reptiles, aves y mamíferos) en busca de alimento, momento en que liberan los huevos junto con las heces. Estos hábitats proporcionan una abundante fuente de alimento durante el hidroperíodo, lo que atrae a muchas aves residentes y migratorias y a medida que el nivel del agua desciende se presenta una fuerte depredación (Bonetto, 1975; Williams, 2006).
En algunas especies anuales del género Nothobranchius, el tamaño de sus poblaciones llega a ser de 1,750-1,956 individuos en un estanque de 875 m2 (2 peces/m2) (Berois et al., 2016). Los hábitats aquí estudiados presentan un área reducida y poca profundidad (180 m2 × 80 cm), aun cuando en algunos casos observamos poblaciones grandes (110 peces) en hábitats con dimensiones de 60 m2 × 7 cm (1.8 peces/m2). Esta condición del hábitat podría incrementar la frecuencia de encuentro parásito-hospedero y por tanto, las tasas de infección se incrementan de un hospedero a otro durante su ciclo de vida. En relación con lo anterior, los valores de los parámetros de la infección que presentan la mayoría de las especies de helmintos encontrados en M. robustus son mayores a los registrados en peces de cuerpos de agua permanentes asociados a la cuenca del río Papaloapan y otros sistemas hidrológicos (tabla 3).
Por otra parte, las especies del género Clinostomum utilizan caracoles de la familia Planorbidae como hospederos intermediarios (Choudhury et al., 2016; Lane y Morris, 2010; Rosser et al., 2017). Se conoce que los esporocistos de varias especies de tremátodos no esquistosómicos causan muerte prematura, alteraciones del crecimiento y castración en los caracoles y bivalvos (Bohes et al., 2010; Esteban et al., 2011); sin embargo, nada se ha documentado sobre estas afectaciones por C. marginatum en sus primeros y segundos hospederos intermediarios. En M. robustus, una cantidad considerable de metacercarias de C. marginatum se encontraron en los testículos y ovarios, un caso extremo con 5 metacercarias ocupando por completo el ovario (fig. 2). Por lo tanto, un efecto negativo sobre la capacidad reproductiva de M. robustus por C. marginatum es posible. En otras especies de peces anuales como Rachovia maculpinnis y R. transilis, se ha documentado la presencia de parásitos en gónadas causando la castración total o parcial de los organismos (Nico y Thomerson, 1989), un factor que podría ser determinante en la tasa de fecundidad y supervivencia de los peces. A menudo, la proporción de sexos en peces anuales está dominada por las hembras (Passos et al., 2014; Reichard et al., 2014). Algunos factores que actúan sobre los machos llegan a representar presiones importantes en sus poblaciones, p. ej., la depredación por aves que se intensifica en especies que presentan coloraciones más llamativas (Reichard et al., 2009) y las altas temperaturas que afectan la esperanza de vida de los machos de algunas especies (Berois et al., 2016). A este escenario se suma el posible efecto negativo de las metacercarias de C. marginatum; por lo tanto, el conocimiento sobre la interacción de M. robustus y su helmintofauna en términos de su reproducción y supervivencia, es pertinente, y comprobar dicho efecto podría dar la pauta para trabajar en potenciales programas de conservación de la especie.
Esta investigación da cuenta del primer estudio sobre la helmintofauna de M. robustus; todas las especies alojadas por este pez anual constituyen nuevos registros. El número relativamente alto de especies de helmintos encontrados en el hospedero y los valores altos en los parámetros de infección, junto con las características físicas del hábitat (área reducida, poca profundidad y permanencia), juegan un rol importante que asegura la transmisión de la infección desde M. robustus hacia los hospederos definitivos.
Finalmente, los cuerpos de agua temporales constituyen hábitats que transitan una fase seca y un hidroperíodo con base en un ciclo anual. El inicio de la estación de lluvias en México está bien definido y consecuentemente el inicio del hidroperíodo y de las poblaciones de M. robustus; por lo tanto, en estos hábitats es posible conocer con exactitud el periodo de desarrollo larvario de las especies de helmintos en sus hospederos intermediarios, aspecto que se complica en cuerpos de agua permanentes. Los estudios sobre patrones de colonización de las especies de helmintos a lo largo del hidroperíodo, podrían dar cuenta de la sincronización de los ciclos de vida de las especies parásitas con el ambiente y con sus hospederos intermediarios y definitivos.
Tabla 3
Comparación de los parámetros cuantitativos de la infección de helmintos de peces de la cuenca del río Papaloapan con relación a los registrados en M. robustus de cuerpos de agua temporales. N = Número de hospederos examinados, P/I = prevalencia e intensidad promedio, (*) = datos de la infección en otros sistemas hidrológicos, – sin datos.
|
Especie de helminto |
Hospedero |
Localidad |
N |
P/I |
Estudio |
|
Valipora campylancristrota |
Rhamdia guatemalensis |
Ixin-há cenote |
131 |
1.5/- |
Scholz et al. (1996)* |
|
Millerichthys robustus |
Playa Salinas |
15 |
26.7/1.5 |
Presente estudio |
|
|
V. minuta |
R. guatemalensis |
Ixin-há cenote |
131 |
3.05/- |
Scholz et al. (1996)* |
|
Poecilia sphenops |
Presa Tepecoacuilco |
6 |
33.3/6.0 |
Scholz y Salgado-Maldonado (2001)* |
|
|
M. robustus |
Tlacotalpan Playa Salinas |
14 9 |
92.9/2.7 44.4/1.0 |
Presente estudio |
|
|
Clinostomum complanatum |
Dormitator maculatus |
Tlacotalpan |
184 |
82.3/49.7 |
Montoya-Mendoza et al. (2004) |
|
36 |
86.1/39.6 |
Salgado-Maldonado et al. (2005) |
|||
|
Cichlasoma urophthalmus |
Tlacotalpan |
10 |
20.0/2.0 |
Salgado-Maldonado et al. (2005) |
|
|
Xipophorus helleri |
Arrollo Balzapote |
8 |
12.5/1.0 |
||
|
C. marginatum |
M. robustus |
Playa Salinas |
15 |
33.3/1.4 |
Presente estudio |
|
Contracaecum sp. |
C. urophthalmus D. maculatus |
Tlacotalpan |
10 11 |
100/1.8 18.2/2.0 |
Salgado-Maldonado et al. (2005) |
|
Ophisternon aenigmaticum |
Lago de Catemaco Tlacotalpan |
27 10 |
11.1/1.0 80.0/3.6 |
||
|
M. robustus |
Playa Salinas |
15 |
40.0/1.7 |
Presente estudio |
|
|
Serpinema trispinosum |
D. maculatus O. aenigmaticum |
Tlacotalpan |
47 10 |
2.0/1.0 10.0/1.0 |
Salgado-Maldonado et al. (2005) |
|
M. robustus |
Tlacotalpan Playa Salinas |
24 9 |
21.0/1.2 33.3/1.0 |
Presente estudio |
|
|
Eustrongylides sp. |
D. maculatus |
Tlacotalpan |
47 |
2.0/1.0 |
Salgado-Maldonado et al. (2005) |
|
M. robustus |
Playa Salinas |
15 |
6.7/6.0 |
Presente estudio |
|
|
Gnathostoma binucleatum |
Gobiomorus dormitor |
Tlacotalpan |
14 |
36.0/2.6 |
Mosqueda y Ocampo (2009) |
|
G. lamothei |
M. robustus |
15 |
26.7/0.8 |
Presente estudio |
|
|
Polymorphus sp. |
R. guatemalensis |
Lago de Catemaco |
4 |
25.0/1.0 |
Salgado-Maldonado et al. (2005) |
|
M. robustus |
Playa Salinas |
15 |
6.7/1.0 |
Presente estudio |
Agradecimientos
A la Semarnat por otorgar la Licencia de Colecta Científica por Proyecto sobre especies o poblaciones en riesgo o hábitat crítico. Nuestra gratitud a Víctor Rosales Pérez por la asistencia en campo durante la búsqueda y recolecta de los peces.
Referencias
Aguilar-Aguilar, R., Contreras-Medina, R., Martínez-Aquino, A., Salgado-Maldonado, G. y González-Zamora, A. (2005). Aplicación del análisis de parsimonia de endemismos (PAE) en los sistemas hidrológicos de México: un ejemplo con helmintos parásitos de peces dulceacuícolas. En J. Llorente-Bousquets y J. J. Morrone (Eds.), Regionalización biogeográfica en Iberoamérica y tópicos afines (pp. 227–239). México D. F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Universidad Nacional Autónoma de México.
Aguilar-Aguilar, R., Martínez-Aquino, A., Espinosa-Pérez, H. y Pérez-Ponce de León, G. (2014). Helminth parasites of freshwater fishes from Cuatro Ciénegas, Coahuila, in the Chihuahuan Desert of Mexico: inventory and biogeographical implications. Integrative Zoology, 9, 328.
Aguilar-Aguilar, R. y Salgado-Maldonado, G. (2006). Diversidad de helmintos parásitos de peces dulceacuícolas en dos cuencas hidrológicas de México: los helmintos y la hipótesis del México betadiverso. Interciencia, 31, 484–490.
Aguilar-Aguilar, R., Salgado-Maldonado, G., Contreras-Medina, R. y Martínez-Aquino, A. (2008). Richness and endemism of helminth parasites of freshwater fishes in Mexico. Biological Journal of the Linnean Society, 94, 435–444.
Aguilar-Aguilar, R., Salgado-Maldonado, G., Moreno-Navarrete, R. G. y Cabañas-Carranza, G. (2004). Helmintos parásitos de peces dulceacuícolas. En I. Luna, J. J. Morrone y D. Espinosa (Eds.), Biodiversidad de la Sierra Madre Oriental (pp. 261–269). México D. F.: Las Prensas de Ciencias, Facultad de Ciencias, Universidad Nacional Autónoma de México.
Berois, N., García, G. y De Sá, R. (2016). Annual fishes: life history strategy, diversity and evolution, Boca Raton: CRC Press.
Blažek, R., Polačik, M. y Reichard, M. (2013). Rapid growth, early maturation and short generation time in African annual fishes. EvoDevo, 4, 24.
Boehs, G., Villalba, A., Ceuta, L. O. y Luz, J. R. (2010). Parasites of three commercially exploited bivalve mollusc species of the estuarine region of the Cachoeira River (Ilhéus, Bahia, Brazil). Journal of Invertebrate Pathology, 103, 43–47.
Bonetto, A. A. (1975). Hydraulic regime of the Parana River and its influence on ecosystems. Ecological Studies, 10, 175–97.
Bush, A. O., Lafferty, K. D., Lotz, J. M. y Shostak, A. W. (1997). Parasitology meets ecology on its own terms: Margolis et al. revisited. Journal of Parasitology, 83, 575–583.
Caffara, M., Locke, S. A., Gustinelli, A., Marcogliese, D. J. y Fioravanti, M. L. (2011). Morphological and molecular differentiation of Clinostomum complanatum and Clinostomum marginatum (Digenea: Clinostomidae) metacercariae and adults. Journal of Parasitology, 97, 884–891.
Caspeta-Mandujano, J. M. (2005). Nematode parasites of freshwater fish in Mexico: key to species, descriptions and distribution. Morelos, Cuernavaca: Universidad Autónoma del Estado de Morelos.
Caspeta-Mandujano, J. M. (2010) Nematode parasites of freshwater fish in Mexico. Mexico D.F.: AGT Editor.
Choudhury, A., Aguirre-Macedo, M. L., Curran, S. S., De Núñez, M. O., Overstreet, R. M., Pérez-Ponce de León, G. et al. (2016). Trematode diversity in freshwater fishes of the Globe II:‘New World’. Systematic Parasitology, 93, 271–282.
Costa, W. J. E. M. (1995). Pearl killifishes: the Cynolebiatinae: systematics and biogeography of the Neotropical annual fish subfamily (Cyprinodontiformes: Rivulidae). Neptune City: TFH Publications.
Delgado, C. (2014). Patrones de diferenciación en el complejo parásito-hospedador integrado por nematodos del género Contracaecum (Nematoda, Anisakidae) y peces anuales (Cyprinodontiformes, Rivulidae) de Uruguay (Tesis de Maestría). Facultad de Ciencias UDELAR, Uruguay.
Delgado, C. y García, G. (2015). Coevolution between Contracaecum (Nematoda, Anisakidae) and Austrolebias (Cyprinodontiformes, Rivulidae) host-parasite complex from SW Atlantic coastal basins. Parasitology Research, 14, 913–927.
Dolfi, L., Ripa, R. y Cellerino A. (2014). Transition to annual life history coincides with reduction in cell cycle speed during early cleavage in three independent clades of annual killifish. EvoDevo, 5, 32.
Domínguez-Castanedo, O., Mosqueda-Cabrera, M. Á. y Valdesalici, S. (2013). First observations of annualism in Millerichthys robustus (Cyprinodontiformes: Rivulidae). Ichthyological Exploration of Freshwaters, 24, 15–20.
Esch, G. W., Kennedy, C. R., Bush, A. O. y Aho, J. M. (1988). Patterns in helminth communities in freshwater fish in Great Britain: alternative strategies for colonization. Parasitology, 96, 519–532.
Esteban, J. G., Muñoz-Antoli, C., Trelis, M. y Toledo, R. (2011). Effects of nonschistosome larval trematodes on Biomphalaria snails. En R. Toledo y B. Fried (Eds.), Biomphalaria snails and larval trematodes (pp. 127–157). New York: Springer.
Falcón-Ordaz, J., Monks, S. y Pulido-Flores, G. (2015). Revisión del uso de céstodos como indicadores de contaminación por metales pesados en mamíferos de medios terrestres. En G. Pulido-Flores, S. Monks y M. López-Herrera (Eds), Estudios en biodiversidad (pp. 78–95). Lincoln, Nebraska: Zea Books.
Furness, A. I. (2016). The evolution of an annual life cycle in killifish: adaptation to ephemeral aquatic environments through embryonic diapause. Biological Reviews, 1, 1–18.
García-Prieto, L., García-Varela, M., Mendoza-Garfias, B. y Pérez-Ponce de León, G. (2010). Checklist of the Acanthocephala in wildlife vertebrates of Mexico. Zootaxa, 2419, 1–50.
Gaspar-Navarro, J., Almeyda-Artigas, R. J., Sánchez-Miranda, E., Carranza-Calderón, L. y Mosqueda-Cabrera, M. Á. (2013). Description of advanced third-stage larvae of Gnathostoma lamothei Bertoni-Ruiz et al. 2005 (Nematoda: Gnathostomatidae) from experimental hosts and contributions to its life cycle. Parasitology Research, 112, 169–175.
Goater, T. M., Goater, C. P. y Esch, G. W. (2013). Parasitism: the diversity and ecology of animal parasites. New York: Cambridge University Press.
Lane, R. L. y Morris, J. E. (2010). Biology, prevention, and effects of common grubs (Digenetic trematodes) in freshwater fish. Ames, Iowa: North Central Regional Aquaculture Center, Technical Bulletins. 14.
Luque, J. L. y Poulin, R. (2007). Metazoan parasite species richness in Neotropical fishes: hotspots and the geography of biodiversity. Parasitology, 134, 865–878.
Marcogliese, D. J. y Cone, D. K. (1997). Food webs: a plea for parasites. Trends in Ecology and Evolution, 12, 320–325.
Miller, R. R. y Hubbs, C. L. (1974). Rivulus robustus, a new cypronodontid fish from southeastern México. Copeia, 865–869.
Monks, S., Pulido-Flores, G., Bautista-Hernández, C. E., Alemán-García, B., Falcón-Ordaz, J. y Gaytán-Oyarzún, J. C. (2013). El uso de helmintos parásitos como bioindicadores en la evaluación de la calidad del agua: lago de Tecocomulco vs. laguna de Metztitlán, Hidalgo, México. En G. Pulido-Flores y S. Monks (Eds.), Estudios científicos en el estado de Hidalgo y zonas aledañas, Vol. II. (pp. 25–34). Lincoln, Nebraska: Zea Books.
Montoya-Mendoza, J., Chávez-López, R. y Franco-López, J. (2004). Helminths from Dormitator maculatus (Pisces: Eleotridae) in Alvarado Lagoon, Veracruz, Mexico, and supplemental data for Clinostomum complanatum Rudolphi, 1814 from Egretta caerulea (Aves: Ardeidae). Gulf and Caribbean Research, 16, 115–127.
Moravec, F. (1998). Nematodes of freshwater fishes of the Neotropical Region. Praga: Publishing House of the Academy of Sciences of the Czech Republic.
Mosqueda-Cabrera, M. Á. y Ocampo-Jaimes, A. (2009). Abnormalities on cephalic hooklets of advanced third-stage larvae from Gnathostoma Owen, 1836 (Nematoda: Gnathostomidae) collected from Mexican rivulus Millerichthys robustus Costa, 1995 (Cyprinodontiformes: Rivulidae) in Tlacotalpan, Veracruz, Mexico. Parasitology Research, 105, 1637.
Myers, G.S. (1952). Annual fishes. Aquarium Journal, 23, 125–141.
Nico, L. G. y Thomerson, J. E. (1989). Ecology, food habits and spatial interactions of Orinoco Basin annual killifish. Acta Biológica de Venezuela, 12, 106–120.
Passos, C., Tassino, B., Reyes, F. y Rosenthal, G. G. (2014). Seasonal variation in female mate choice and operational sex ratio in wild populations of an annual fish, Austrolebias reicherti. Plos One, 9, e101649.
Pérez-Ponce de León, G., Rosas-Valdez, R., Aguilar-Aguilar, R., Mendoza-Garfias, B., Mendoza-Palmero C., García-Prieto, L. et al. (2010). Helminth parasites of freshwater fishes, Nazas River basin, northern Mexico. Check List, 6, 26–35.
Pérez-Ponce de León, G., Rosas-Valdez, R., Mendoza-Garfias, B., Aguilar-Aguilar, R. y Falcón-Ordaz, J. (2009). Survey of the endohelminth parasites of freshwater fishes in the upper Mezquital River Basin, Durango State, Mexico. Zootaxa, 2164, 1–20.
Podrabsky, J. E., A. Tingaud-Sequeira y Cerdà, J. (2010). Metabolic dormancy and responses to environmental desiccation in fish embryos. En E. Lubzens, J. Cerdà y M. Clark (Eds.), Topics in current genetics. Dormancy and resistance in harsh environments (pp. 203–226). Berlín, Heidelberg: Springer.
Pulido-Flores, G., Monks, S. y Gordillo-Martínez, A. J. (2005). Monitoreo de bajo costo en la evaluación de la calidad ambiental. Revista Internacional de Ciencias Ambientales, 21, 578–583.
Reichard, M., Polačik, M., Blažek, R. y Vrtílek, M. (2014). Female bias in the adult sex ratio of African annual fishes: interspecific differences, seasonal trends and environmental predictors. Evolutionary Ecology, 28, 1105–1120.
Reichard, M., Polačik, M. y Sedláček, O. (2009). Distribution, colour polymorphism and habitat use of the African killifish Nothobranchius furzeri, the vertebrate with the shortest life span. Journal of Fish Biology, 74, 198–212.
Rosser, T. G., Alberson, N. R., Woodyard, E. T., Cunningham, F. L., Pote, L. M. y Griffin, M. J. (2017). Clinostomum album n. sp. and Clinostomum marginatum (Rudolphi, 1819), parasites of the great egret Ardea alba L. from Mississippi, USA. Systematic Parasitology, 94, 35–49.
Salgado-Maldonado, G. (2006). Checklist of helminth parasites of freshwater fishes of Mexico. Zootaxa, 1324, 1–357.
Salgado-Maldonado, G., Aguilar-Aguilar, R., Cabañas-Carranza, G., Soto-Galera, E. y Mendoza-Palmero, C. (2005). Helminth parasites in freshwater fish from the Papaloapan river basin, México. Parasitology Research, 96, 69–89.
Salgado-Maldonado, G., Cabañas-Carranza, G., Caspeta-Mandujano, J. M., Soto-Galera, E., Mayén-Peña, E., Brailovsky, D. y Báez-Vale, R. (2001). Helminth parasites of freshwater fishes of the Balsas River drainage basin of southwestern Mexico. Comparative Parasitology, 68, 196–203.
Salgado-Maldonado, G., Cabañas-Carranza, G., Soto-Galera, E., Caspeta-Mandujano, J. M., Moreno-Navarrete, R. G., Sánchez-Nava, P. et al. (2001). A checklist of helminth parasites of freshwater fishes from the Lerma Santiago river basin, Mexico. Comparative Parasitology, 68, 204–217.
Salgado-Maldonado, G., Cabañas-Carranza, G., Soto-Galera, E., Pineda-López, R., Caspeta-Mandujano, J. M., Aguilar-Castellanos, E. et al. (2004). Helminth parasites of freshwater fishes of the Pánuco River Basin, east Central Mexico. Comparative Parasitology, 71, 190–202.
Salgado-Maldonado, G., Caspeta-Mandujano, J. M., Moravec, F., Soto-Galera, E., Rodiles-Hernández, R., Cabañas-Carranza, G. et al. (2011). Helminth parasites of freshwater fish in Chiapas, Mexico. Parasitology Research, 108, 31–59.
Salgado-Maldonado, G., Mercado-Silva, N., Cabañas-Carranza, G., Caspeta-Mandujano, J. M., Aguilar-Aguilar, R. e Íñiguez-Dávalos, L. I. (2004). Helminth parasites of freshwater fishes of the Ayuquila river, Sierra de Manantlán Biosphere Reserve, west-central Mexico. Comparative Parasitology, 71, 67–72.
Salgado-Maldonado, G., Novelo-Turcotte, M. T., Caspeta-Mandujano, J. M., Vázquez-Hurtado, G., Quiroz-Martínez, B., Mercado-Silva, N. et al. (2016). Host specificity and the structure of helminth parasite communities of fishes in a Neotropical river in Mexico. Parasite, 23, 61.
Salgado-Maldonado, G. y Quiroz-Martínez, B. (2013). Taxonomic composition and endemism of the helminth fauna of freshwater fishes of Mexico. Parasitology Research, 112, 1–18.
Scholz, T. y Salgado-Maldonado, G. (2001). Metacestodes of the family Dilepididae (Cestoda: Cyclophyllidea) parasitizing fishes in Mexico. Systematic Parasitology, 49, 23–39.
Scholz, T., Vargas-Vázquez, J., Moravec, F., Vivas-Rodríguez, C. y Mendoza-Franco, E. (1996). Cestoda and Acanthocephala of fishes from cenotes (sinkholes) of Yucatan, Mexico. Folia Parasitologica, 43, 141–152.
Vidal-Martínez, V. M., Aguirre-Macedo, M. L., Scholz, T., González-Solís, D. y Mendoza-Franco, E. F. (2001). Atlas of the helminth parasites of cichlid fish of Mexico. Praga, Czech Republic: Academia.
Williams, D. D. (2006). The biology of temporary waters. Londres: Oxford University Press.
Wourms, J. P. (1972a). Developmental biology of annual fishes. I. Stages in the normal development of Austrofundulus myersi Dahl. Journal of Experimental Biology, 182, 143–168.
Wourms, J. P. (1972b). Developmental biology of annual fishes. II. Naturally occurring dispersion and reaggregation of blastomeres during the development of annual fish eggs. Journal of Experimental Biology, 182, 169–200.


