Huracán Jova: efecto de un fenómeno meteorológico severo sobre lagartijas insulares en el occidente de México
Andrés García a y Héctor Hugo Siliceo-Cantero b, *
a Estación de Biología Chamela, Instituto de Biología, Universidad Nacional Autónoma de México, Apartado postal 21, 48980 San Patricio, Melaque, la Huerta, Jalisco, México
b Instituto de Investigaciones en Ecosistemas y Sustentabilidad, Universidad Nacional Autónoma de México, Antigua Carretera a Pátzcuaro Núm. 8701, Ex-Hacienda de San José de la Huerta, 59180 Morelia, Michoacán, México
*Autor para correspondencia: hehusic@gmail.com (H.H. Siliceo-Cantero)
Resumen
El objetivo del presente estudio fue estimar el efecto del huracán Jova (12 de octubre de 2011) sobre el estrato arbóreo y los atributos poblacionales de 3 especies de lagartijas (Anolis nebulosus, Aspidoscelis lineattissimus y Urosaurus bicarinatus) en una pequeña isla de 3.3 ha (San Agustín), ubicada a 500 m de la costa de Jalisco, México. Para ello, se calculó el daño sufrido por los árboles en la isla y la relación con su altura y su diámetro. Además, se estimaron cambios en la densidad poblacional de las 3 especies de lagartijas, así como cambios en la supervivencia, talla, peso corporal y uso del hábitat de A. nebulosus y U. bicarinatus. Los resultados indicaron que Jova afectó a un tercio de los árboles muestreados, principalmente a los árboles más altos y delgados. La densidad de las 3 especies se incrementó tras el paso de Jova, así como la talla y el peso de A. nebulosus, pero no tuvo efecto sobre su supervivencia. Jova afectó el uso del hábitat de A. nebulosus pero no de U. bicarinatus. A pesar del efecto negativo sobre la vegetación y la elevada precipitación, Jova no afectó negativamente a las poblaciones de lagartijas.
Palabras clave: Anolis nebulosus; Aspidoscelis lineattissimus; Tamaño corporal; Uso del hábitat; Tasa de supervivencia en islas; Daño en árboles; Urosaurus bicarinatus
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Hurricane Jova: the effect of a severe meteorological phenomenon on insular lizards in western Mexico
Abstract
The focus of this study was to estimate the effect of hurricane Jova (12 October 2011) on the arboreal strata and population attributes of three lizard species (Anolis nebulosus, Aspidoscelis lineattissimus, and Urosaurus bicarinatus) of a small island of 3.3 ha (San Agustin), located 500 m from the coast of Jalisco, Mexico. To do this, the damage on trees and its relationship with diameter and height were calculated. Furthermore, we estimated population density changes in 3 lizard species, as well as the survivorship rate, size, body weight, and habitat use of A. nebulosus, and U. bicarinatus. The results indicated that the third part of recorded trees was damaged by Jova, principally the tallest and thinner ones. The density of the 3 lizard species increased after Jova, as well as the size and weight of A. nebulosus, however, the survivorship of this last species was not affected. The habitat use changed after Jova, but only in A. nebulosus. Hurricane Jova did not negatively affect the lizard populations despite the negative effects occurred in the vegetation and the amount of precipitation.
Keywords: Anolis nebulosus; Aspidoscelis lineattissimus; Body size; Habitat use; Survivor rates on islands; Tree damage; Urosaurus bicarinatus
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
En la actualidad, algunos estudios indican que la frecuencia, duración e intensidad de fenómenos naturales como los huracanes se ha incrementado en los últimos 35 años, siendo una de las causas el aumento de la temperatura de la superficie de los océanos (Hosseini et al., 2018; Webster et al., 2005; Zhao y Held, 2012). Con ello, diversas regiones continentales se han enfrentado inesperadamente a condiciones climáticas extremas como lluvias torrenciales y vientos intensos sostenidos (Folmer et al., 2015). Dichos vientos tienen efectos marcados sobre la vegetación que no está adaptada a estas condiciones, por lo que se reduce la altura del dosel, eliminando en gran medida a los individuos de mayor talla (Boutet y Weishampel, 2003).
Las islas son uno de los sistemas más afectados por los huracanes debido a que sufren el impacto de estos fenómenos meteorológicos antes que la zona continental; de hecho, algunas de ellas son reconocidas como “islas barrera” porque atenúan el impacto de los huracanes sobre la zona continental (Conery et al., 2018). La topografía de las islas influye en el efecto que tienen los huracanes sobre las mismas, una topografía con relieve complejo y pronunciado permite que algunas áreas no se expongan a los fuertes vientos, contrariamente, una topografía de relieve simple y plano permite que el impacto se extienda a una mayor superficie a través de la isla (Fickert, 2018; Walker, 1991). Así, la vegetación que se encuentra protegida por un relieve sufre menor daño que la vegetación que se encuentra directamente expuesta a los vientos (Walker, 1991). El daño que suele presentar la vegetación tras el paso de un huracán es la defoliación, rompimiento de ramas y troncos principales, así como desenraizamiento de algunos individuos, causando una mortalidad que puede oscilar entre 10% y 100% (Fickert, 2018; Ugarte et al., 2006; Walker, 1991). Adicionalmente, tras el paso de un huracán, la vegetación pude reducir su crecimiento, retrasando el restablecimiento de las condiciones bióticas del sistema (Funderburk et al., 2016).
El daño sobre la estructura vegetal tiene claras consecuencias sobre las poblaciones animales, debido a que la pérdida de follaje (Maass et al., 2017) y la mortalidad de algunos elementos vegetales pueden reducir la disponibilidad de microhábitats y alimento (Rittenhouse et al., 2010). Adicionalmente, los huracanes tienen efectos directos sobre las especies animales (Lugo, 2008; Nicholas y Walker, 1991). El más obvio y extremo es la eliminación de las poblaciones insulares (Schoener et al., 2004), la reducción de las poblaciones que puede conducir a una alta endogamia y consecuentemente poblaciones inviables (Pavelka et al., 2007), y el arrastre de individuos que puede aumentar la concentración de los mismos en ciertas zonas o llevarlos a establecer nuevos territorios, provocando la mezcla de poblaciones e incrementando con ello, interacciones riesgosas como la depredación (Johnson y Winker, 2010; Wiley y Wunderle, 1993).
En el caso particular del grupo de las lagartijas insulares, los efectos que los huracanes causan en sus poblaciones se han abordado en diversos trabajos (Donihue et al., 2018; Reagan, 1991; Schoener et al., 2004; Spiller y Schoener, 2007). Por ejemplo, 3 de 11 poblaciones insulares de Anolis sagrei fueron exterminadas por el huracán Floyd, y otras se redujeron en promedio casi 50%, siendo la topografía compleja un factor importante para la permanencia de las poblaciones (Schoener y Spiller, 2006; Schoener et al., 2001, 2004). Estos fenómenos naturales suelen arrastrar no solo a las especies de lagartijas fuera de las islas, sino también arrastran sus recursos alimenticios (Schoener y Spiller, 2006). Especies como Anolis stratulus y A. gundlachi modificaron el uso que hacen del hábitat debido a la reducción de la cobertura vegetal que provocó el huracán Hugo en Puerto Rico (Reagan, 1991). Un trabajo reciente indica que los huracanes pueden ser una causa de selección natural, ya que el tamaño de algunas estructuras corporales permite mayor sujeción ante los fuertes vientos, por lo que son los organismos que persisten tras el paso de un huracán (Donihue et al., 2018). Adicionalmente, la eliminación de ciertos fenotipos tras el paso de los huracanes en las islas puede afectar las interacciones bióticas como la polinización (Temeles y Bishop, 2019). Un ejemplo de la modificación de las interacciones bióticas por huracanes en lagartijas se presenta en Anolis sagrei, cuya reducción poblacional tras el paso de Floyd permitió el incremento de artrópodos defoliadores, los cuales a su vez impactaron severamente la vegetación insular (Spiller y Schoener, 2007).
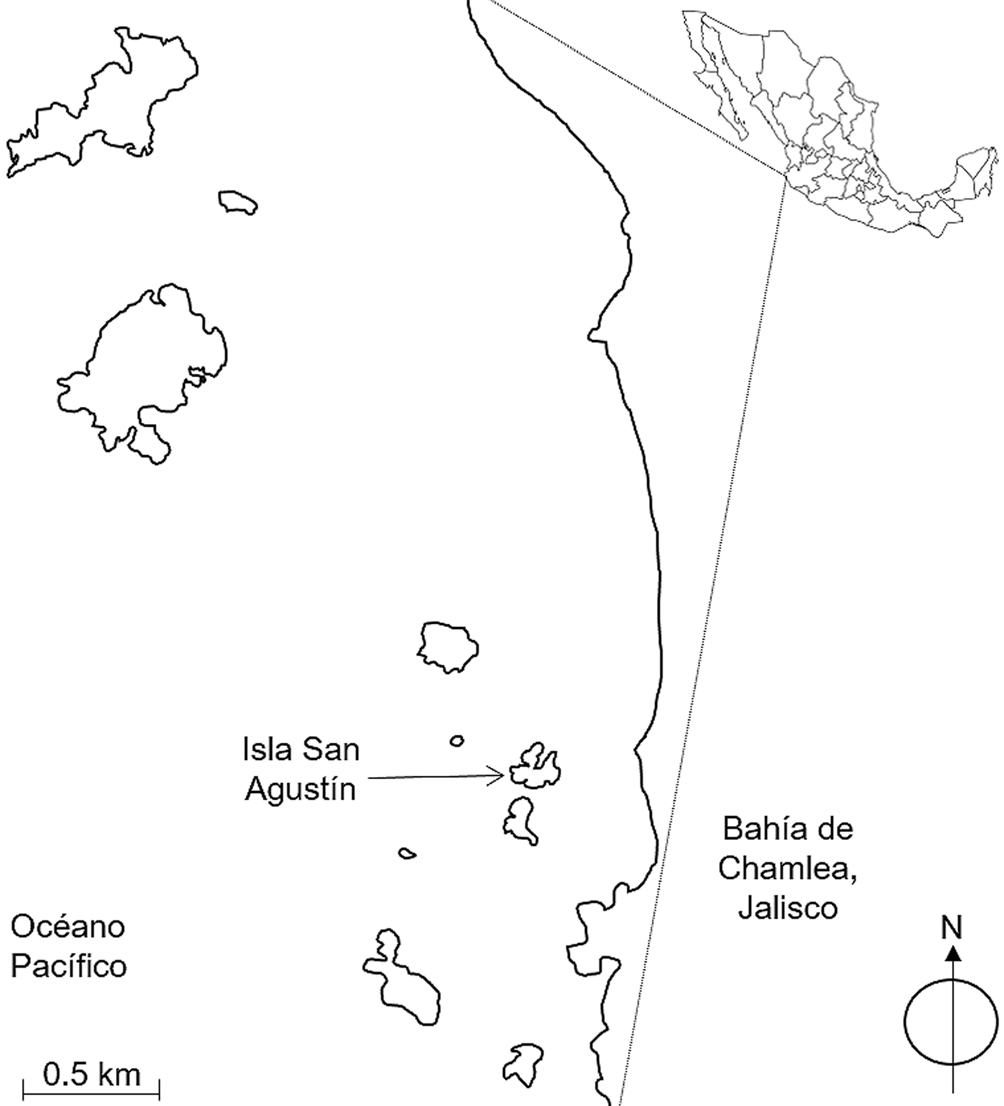
La ocurrencia de huracanes en el Pacífico mexicano es alta, registrando alrededor de 65 en un lapso de 50 años (de 1951 a 2000), de los cuales, 18% fueron de categoría 2, 9% de categoría 3, 12% de categoría 4 y 1.5% de categoría 5 (Jáuregui, 2003). Durante el mismo periodo, pero en el caso específico de la costa de Jalisco se registraron solo 10 huracanes (Jáuregui, 2003). Sin embargo, en años recientes la frecuencia e intensidad de los huracanes se ha incrementado en el Pacífico mexicano y en Jalisco en particular (Knutson et al., 2015; Maass et al., 2017). Un ejemplo de ello es el huracán Jova (categoría 2) que impactó cerca de Chamela, Jalisco, México, el 12 de octubre de 2011, con vientos de hasta 342 km/h y 299 mm de precipitación. Esto resultó en el daño de árboles y, consecuentemente, en una modificación de la estructura de la vegetación, tanto de la selva del continente como de las pequeñas islas de la región. Éste es el caso de la isla San Agustín (ISA) que cuenta con tan solo 3.3 ha de extensión, ubicada a 450 m de la costa en el suroeste de Jalisco.
Adicionalmente, en la región impera una marcada estacionalidad en la precipitación, con lluvias que se concentran en tan solo 4 meses al año, lo que resulta en un paisaje contrastante entre la estación seca y la estación de lluvias (Bullock, 1986; Maass et al., 2017). Esta estacionalidad ambiental también se refleja en la disponibilidad de recursos para las especies animales, entre los que se encuentran: sitios de termorregulación, refugios, alimento y agua, entre otros, los cuales se incrementan durante la temporada de lluvias (Ceballos y Miranda, 2000; García y Ceballos, 1994). En la región, son diversos los estudios ecológicos que incluyen a la estacionalidad como uno de los factores que influyen sobre las poblaciones de lagartijas (Gienger et al., 2002; Siliceo-Cantero y García, 2014; Valtierra-Azotla et al., 2010). Un estudio reciente (Maass et al., 2017) que analiza la dinámica hidrológica en la región de la costa de Jalisco, reporta un incremento reciente en la precipitación anual y en la duración de la estación de lluvias, así como mayor frecuencia en la formación de huracanes. Por ejemplo, el paso del huracán Jova significó un incremento en la precipitación anual de 24% con respecto a 2008, 118% con respecto a 2009 y 26% con respecto a 2010, según datos de la estación meteorológica de la Estación de Biología Chamela (UNAM). Dicho incremento podría influir sobre las condiciones del entorno a las que se enfrentan las poblaciones de lagartijas y consecuentemente, desarrollar un contraste marcado en su ecología entre la estación seca y la estación lluviosa.
Actualmente, no existe información publicada sobre los efectos del huracán Jova sobre las poblaciones de lagartijas en la región, a pesar del severo impacto de Jova y de la atención generada por ser considerado como un fenómeno atípico en la región por su gran precipitación (periodo de retorno de 75 años; Olguín-López et al., 2015). Sin embargo, la toma de datos ecológicos sobre 3 especies de lagartijas en ISA (Anolis nebulosus, Aspidoscelis lineattissimus y Urosaurus bicarinatus) previo y posterior a Jova, permitió determinar los efectos inmediatos de este huracán, principalmente sobre una población de lagartijas Anolis nebulosus en la costa de Chamela, Jalisco, México. Para ello, el presente estudio pretende responder las siguientes preguntas: ¿cuál fue el daño causado por el huracán Jova a la vegetación de ISA?, ¿qué atributos poblacionales de A. nebulosus (densidad poblacional, uso de la percha, tamaño corporal, peso corporal, y supervivencia) fueron los más afectados por Jova?, ¿cuál es el papel de la estacionalidad en A. nebulosus tras el paso de Jova?, ¿ el paso de Jova afectó hembras y machos de A. nebulosus por igual? y ¿cómo respondieron otras especies de lagartijas habitantes de ISA como la terrestre Aspidoscelis lineattissimus y la arborícola Urosaurus bicarinatus al paso del huracán Jova?
En el presente trabajo se presentan 3 hipótesis del impacto de Jova sobre las poblaciones insulares de lagartijas y su entorno: 1) debido a que ISA no presenta un relieve complejo que evite la exposición de la vegetación a los fuertes vientos y a la alta cantidad de precipitación (Fickert, 2018; Walker, 1991), Jova afectará ampliamente a la vegetación, desenraizando árboles, rompiendo troncos principales y ramas, lo que ocurrirá principalmente en los individuos de mayor talla que están más expuestos (Boutet y Weishampel, 2003). 2) Debido a los cambios en la estructura vegetal antes expuestos (reducción de la altura y pérdida del dosel), así como a la gran cantidad de lluvia, y fuerza y velocidad del viento propios de los huracanes (que suele lavar a lagartijas insulares; Schoener y Spiller, 2006; Schoener et al., 2001, 2004), se espera que las poblaciones de lagartijas en ISA se vean diezmadas en su densidad, que las lagartijas perchen a alturas más bajas debido a la pérdida de los árboles de mayor talla y disminución del dosel en general, y disminución en los atributos individuales como la longitud y peso de las lagartijas A. nebulosus debido a los efectos negativos del huracán sobre la disponibilidad de sus recursos alimenticios. 3) Finalmente, la marcada estacionalidad que propicia fluctuaciones generales asociadas con la disponibilidad de recursos y cambios en el microclima, que suelen causar cambios estacionales en el comportamiento de las lagartijas Anolis, como el uso de perchas más altas durante la estación lluviosa (Lister y García, 1992) y mejores condiciones corporales en la estación lluviosa (Siliceo-Cantero y García, 2014), se verán atenuados por factores provenientes del huracán como la reducción en la altura del dosel (Reagan, 1991), la mortalidad de lagartijas y el lavado o arrastre de recursos alimenticios (Schöner y Spiller, 2006).
Materiales y métodos
La isla San Agustín (ISA) se ubica a 0.45 km de la costa y a 1.9 km al noroeste del poblado de Chamela, municipio de La Huerta, Jalisco, México (19°32’05” N, 105°32’18” O; 15 m snm) (fig. 1). En ISA, el bosque tropical caducifolio (BTC) es la vegetación predominante, aunque está menos estructurada y es de menor tamaño que el BTC en la zona continental adyacente (Conanp, 2008). En la región, la temperatura promedio anual es 25 °C y la precipitación promedio anual es de 749 mm que se concentra de julio a octubre (Bullock, 1986; Maass et al., 2017). En ISA se muestrearon 1,875 m2 divididos en 3 parcelas de 25 × 25 m, es decir, 5% de los 33,000 m2 que comprenden su área total.
Entre agosto y septiembre de 2012, 9 meses después del paso del huracán Jova, se determinó el daño a los árboles en ISA en 3 parcelas de 25 × 25 m estudiadas previamente (Siliceo-Cantero y García, 2014). Para cada árbol dentro de las parcelas, se midió la altura y el diámetro a la altura del pecho (DAP) con la ayuda de una cinta métrica (solo se tomaron en cuenta árboles con un DAP mayor a 1 cm). Los árboles menores a 2 m de altura fueron medidos directamente con la cinta métrica, mientras que para árboles > 2 m de altura, se colocó la cinta métrica de 2 m verticalmente desde la base del árbol para posteriormente estimar visualmente su altura total. Los datos de diámetro y altura de la percha fueron categorizados, redondeando los números a las categorías establecidas, las cuales fueron para el diámetro: 5, 10, 20, 30, 40 y 50 cm; mientras que para la altura fueron: 200, 300, 400, 500, 600 y 700 cm. El daño a los árboles fue categorizado de la siguiente manera: 1) sin daño, árboles que no presentaron daño alguno en el tronco principal; 2) árbol roto, árboles cuyo tronco principal fue roto a una altura mayor o igual a 1 m y cuya parte superior se desprendió; 3) árbol doblado, árboles cuyo tronco principal presentó un doblez sin que se desprendiera la parte superior del mismo; y 4) árbol desenraizado, árboles cuyo tronco principal fue arrancado de raíz.
Cada parcela fue muestreada un total de 24 veces (48 h totales por parcela). Previo al paso del huracán Jova, se muestreó 6 veces cada parcela (2 h cada una) en la estación seca (febrero-abril, 2011) y 6 veces en la estación de lluvias (septiembre, 2011). Asimismo, posterior al paso de Jova, se muestreó 6 veces cada parcela en la estación seca (abril, 2012) y 6 veces en la estación lluviosa (agosto-septiembre, 2012). Para todas las lagartijas A. nebulosus observadas (315) se registró la altura sobre el suelo a la que se encontraba y el diámetro del árbol donde cada individuo estaba perchado. Del total de lagartijas A. nebulosus observadas, se capturaron, sexaron (presencia “macho” o ausencia “hembra” del abanico gular), midieron y pesaron 104 individuos. Para cada individuo A. nebulosus, se midió la longitud hocico-cloaca (LHC) usando un vernier digital (precisión 0.01 mm) y el peso corporal usando una báscula de resorte de 5 g (precisión 0.301 g). Al capturar un individuo A. nebulosus por primera vez, éste fue marcado mediante ectomización de falanges, cuidando de cortar solo la punta del dedo que está libre de lamelas (éstas permiten mayor sujeción a las superficies) y así evitar disminuir su desempeño en el hábitat. Estos datos fueron utilizados para estimar la probabilidad de supervivencia de A. nebulosus. Además, se registró la presencia de otras 2 especies de lagartijas presentes en la isla: Aspidoscelis lineattissimus (terrestre) y Urosaurus bicarinatus (arborícola) dentro de las mismas parcelas. Estas especies compiten potencialmente con A. nebulosus, además, para U. bicarinatus también se registraron datos sobre la altura de la percha. Se puede consultar información adicional sobre estas 3 especies de lagartijas de la región de Chamela en Navarro-García et al. (2008), Siliceo-Cantero y García (2013) y Renton et al. (2017).
Para determinar si existe una relación entre el tipo de daño a los árboles y las características físicas de los mismos, primeramente, se obtuvieron el porcentaje total de árboles con daño y el porcentaje de árboles por tipo de daño para cada una de las categorías de altura y diámetro. Posteriormente, se realizaron correlaciones de Spearman entre los porcentajes obtenidos y las categorías de diámetro y altura de los árboles.
Para estimar la densidad de lagartijas A. nebulosus (individuos/ha) por día, se multiplicó el número total de individuos observados (machos y hembras por separado) dentro de las 3 parcelas durante un día por 10,000 (1 ha) y se dividió el resultado entre el área total muestreada (1,875 m2). Finalmente, se realizó un ajuste de la densidad de lagartijas A. nebulosus por día, al dividirla por la probabilidad media mensual de recaptura (p). Este último dato fue obtenido a partir del análisis de captura-marca-recaptura (ver abajo). Para estimar posibles cambios en la densidad, la altura y el diámetro de la percha utilizada, y la longitud de las lagartijas A. nebulosus ocasionados por el paso de Jova, la estacionalidad (secas, lluvias), o el sexo, se aplicó un Anova de 3 vías mediante modelos lineales generalizados (MLG), utilizando una distribución Poisson, ya que los datos no se distribuyeron normalmente. El peso corporal presentó una distribución normal, por lo que se utilizó un Anova de 3 vías ordinario para estimar el efecto de Jova, la estacionalidad y el sexo. Todos los análisis se realizaron con el paquete estadístico Statistica 7 (STAT Soft, Tulsa, Oklahoma, USA). Para el caso de densidad, altura y diámetro de la percha se usaron datos del total de individuos observados, mientras para estimar talla y peso se usaron datos de los individuos capturados.
Se estimó la probabilidad de supervivencia (ϕ) y de captura (p) de las lagartijas, usando el procedimiento de máxima verosimilitud incluido en el programa Mark ® (Colorado State University, Colorado, USA; Lebreton et al., 1992; White y Burnham, 1999). Este programa utiliza el criterio de información de Akaike (AIC). El AIC ofrece el modelo más verosímil que mejor explica la variación de los datos, utilizando la menor cantidad de parámetros. El AIC se define como AIC = -2LN (£) + 2K, donde £ es el modelo de verosimilitud y K es el número de parámetros en el modelo. Los modelos se basan en historias de captura y recaptura de los individuos (Lebreton et al., 1992; White y Burnham, 1999). Se construyeron varios modelos para estimar diferentes fuentes de variación en ϕ y p, examinando el efecto del huracán Jova sobre la supervivencia y probabilidad de captura, es decir, dependiente del tiempo; además, se examinó el efecto del sexo sobre estos parámetros en la población de lagartijas A. nebulosus. Los modelos que proporcionaron resultados de ϕ y p por mes, permitieron contrastar la probabilidad de captura y de supervivencia tras el paso del huracán Jova (septiembre, 2011 – febrero, 2012) con respecto al promedio previo a Jova (octubre, 2010 – septiembre, 2011).
Para estimar la densidad de las otras especies (A. lineattissimus y U. bicarinatus), se realizó el mismo procedimiento descrito para A. nebulosus; sin embargo, para el ajuste de la densidad, ésta se dividió entre la probabilidad de detección (Armstrong et al., 2005), calculada mediante modelos de ocupación implementados en el programa PRESENCE (U.S. Geological Survey and Colorado State University, Colorado, USA; MacKenzie et al., 2017). Dichos modelos se basan en visitas repetidas a los sitios de observación que se usan para estimar la probabilidad de que se detecte a un individuo, dado que se encuentre presente en dicho sitio (MacKenzie et al., 2017). Para los datos de densidad ajustados, así como para la altura de la percha usada por U. bicarinatus, se aplicaron pruebas de Anova de 2 vías para estimar los efectos de Jova y de la estacionalidad, estas variables presentaron una distribución normal. Los datos del diámetro de la parcha utilizada por U. bicarinatus no presentaron una distribución normal, por lo que para estimar el efecto del huracán Jova y de la estacionalidad se aplicó una prueba de Anova de 2 vías usando un MLG con distribución Poisson. Del mismo modo que los análisis para A. nebulosus, todos estos análisis fueron llevados a cabo usando el programa Statistica 7 (STAT Soft, Tulsa, Oklahoma, USA). Los promedios reportados son acompañados por el error estándar.
Resultados
Alrededor de 36% de los 337 árboles registrados dentro de las parcelas fueron dañados por el huracán. Los resultados indicaron una correlación positiva de la altura de los árboles con el porcentaje de daño total y la categoría árbol roto, además de una correlación negativa con el tipo de daño árbol doblado (tabla 1). Por otro lado, el diámetro se correlacionó positivamente con el porcentaje de daño total y las categorías de daño árbol roto y árbol desenraizado (tabla 1).
Tabla 1
Correlaciones de Spearman entre el tipo de daño de árboles causado por el huracán Jova (octubre 2010) y la altura y diámetro de los árboles de la Isla San Agustín en la costa de Jalisco, México.
|
Tipo de daño |
Altura |
Diámetro |
|
Roto |
R = 0.82, p = 0.04 |
R = 0.94, p = 0.004 |
|
Doblado |
R = -0.89, p = 0.01 |
R = -0.14, p = 0.78 |
|
Desenraizado |
R = 0.31, p = 0.54 |
R = 0.82, p = 0.04 |
|
Total dañados |
R = 0.82, p = 0.04 |
R = 0.82, p = 0.04 |
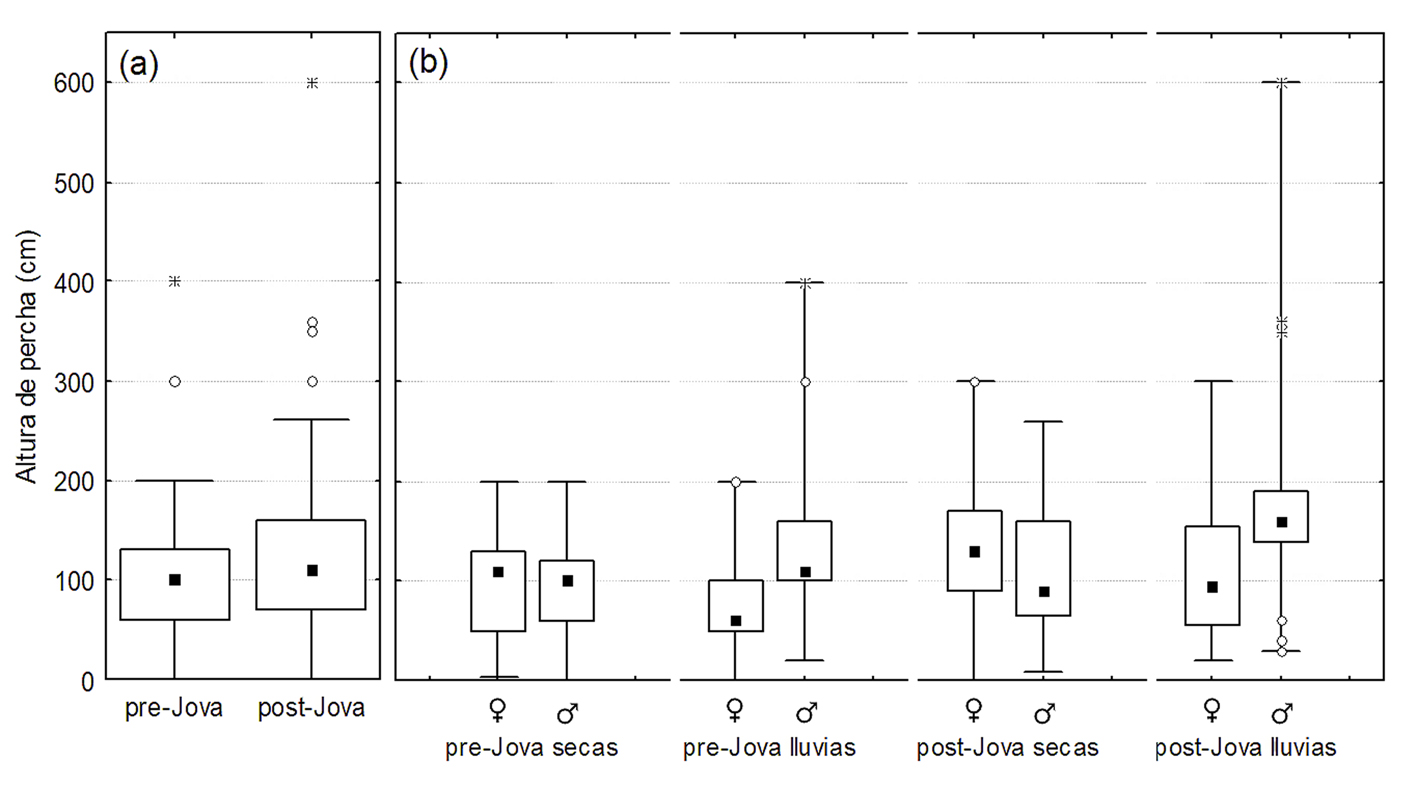
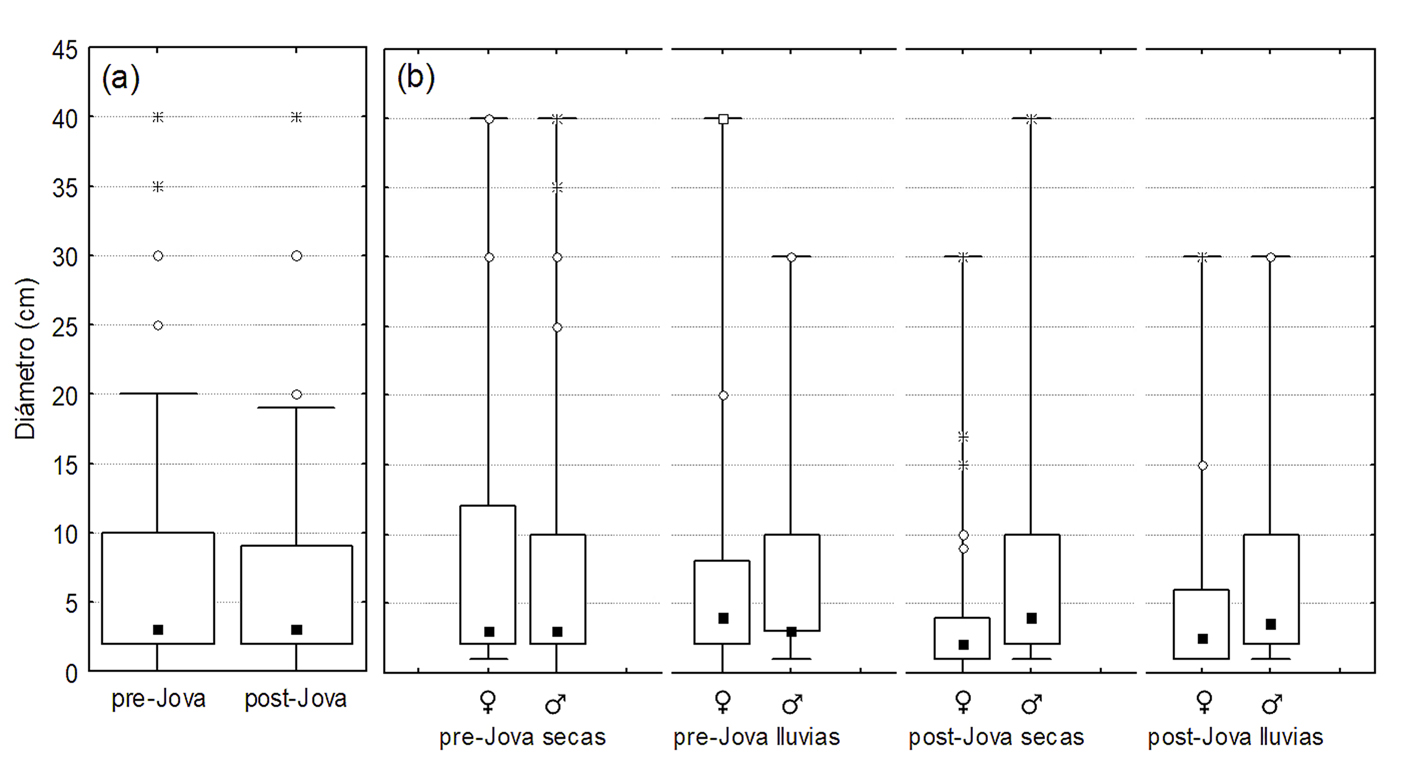
De manera general, se observaron 312 lagartijas A. nebulosus (171 machos y 141 hembras; 126 antes y 186 después del paso de Jova; 230 en la estación seca y 82 en la lluviosa). Los resultados indicaron un efecto significativo del huracán Jova sobre la densidad poblacional de A. nebulosus (χ2 = 404, df = 1, p < 0.001), así como sobre la altura (χ2 = 11.8, df = 1, p < 0.001) y diámetro (χ2 = 36.9, df = 1, p < 0.001) de la percha usados por las lagartijas. El efecto Jova-estacionalidad fue significativo para la densidad poblacional (χ2 = 73, df = 1, p < 0.001) y el diámetro de la percha usado (χ2 = 7.9, df = 1, p = 0.004), mientras que el efecto Jova-sexo solo fue significativo para el diámetro de percha usado (χ2 = 33.6, df <0 1, p < 0.001). La densidad poblacional de A. nebulosus se incrementó tras el paso de Jova (fig. 2a). Dicho incremento ocurrió entre estaciones secas y entre estaciones de lluvias (fig. 2b). Solo en la estación de lluvias previo a Jova se observó una marcada diferencia entre la densidad de hembras y machos, siendo mayor la de estos últimos (fig. 2b). En cuanto al uso del hábitat, la altura a la cual perchan las lagartijas A. nebulosus se incrementó tras el paso de Jova (fig. 3a), siendo relativamente mayor en lluvias que en secas previo y posterior a Jova, y relativamente mayor en machos que en hembras para ambas estaciones de lluvias (fig. 3b). Por otro lado, se redujo el diámetro de percha usado por las lagartijas tras el paso de Jova (fig. 4a), dicho decremento fue más marcado en hembras que en machos, sobretodo en la temporada de lluvias (fig. 4b).

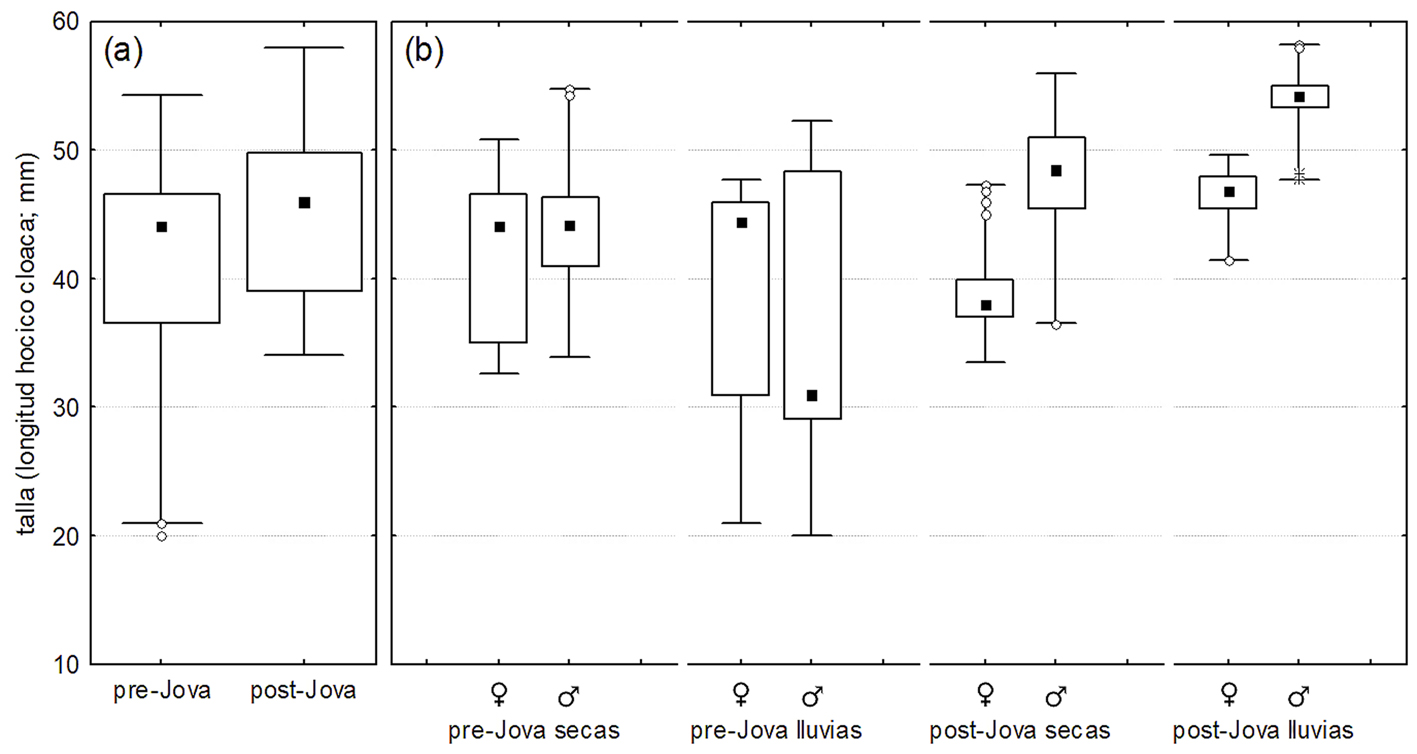
Se capturaron un total de 278 A. nebulosus (147 machos y 131 hembras; 126 antes y 152 después de Jova; 210 en la temporada de secas y 68 en la de lluvias) de los 312 observados. Los resultados indicaron un efecto significativo de Jova sobre la talla (χ2 = 16.4, df = 1, p < 0.001) y peso de A. nebulosus. El efecto Jova-estacionalidad fue significativo para la talla (χ2 = 32, df = 1, p < 0.001) y el peso (F = 13.9, df = 1, p < 0.001), también el efecto Jova-sexo fue significativo tanto para la talla (χ2 = 15.6, df = 1, p < 0.001) como para el peso (F = 28.5, df 0 1, p < 0.001). La talla de las lagartijas se incrementó posterior a Jova (fig. 5a), el incremento fue más marcado entre estaciones de lluvias que entre estaciones des secas, y más pronunciado en machos que en hembras (fig. 5b).
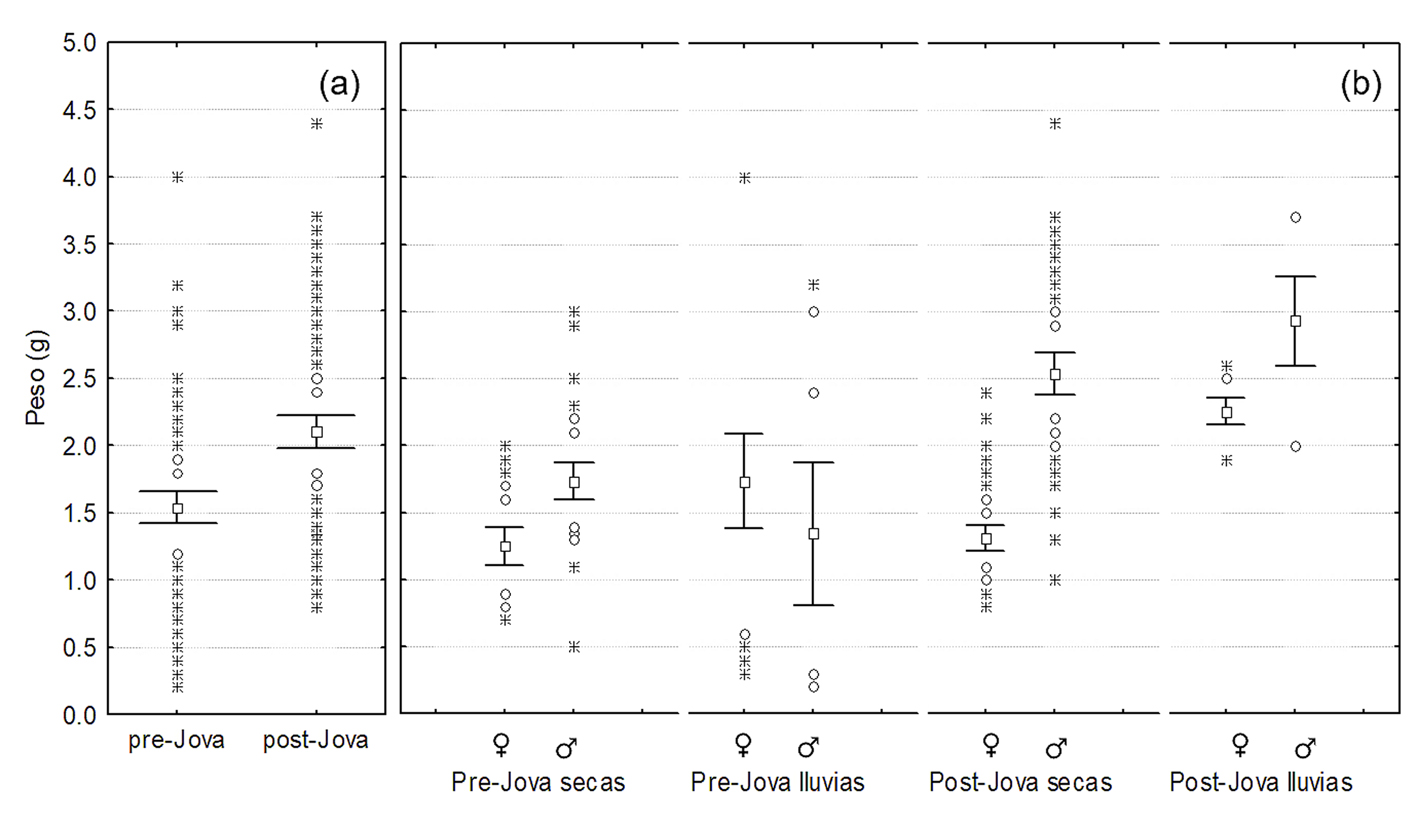
El mismo patrón de incremento tras el paso de Jova se registró en el peso (fig. 6a), siendo también más marcado en la estación lluviosa y también en machos (fig. 6b).
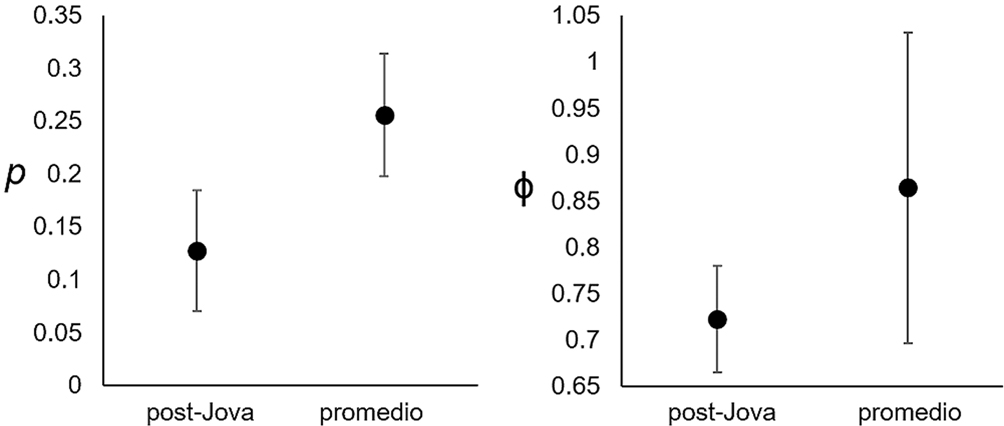
Se marcaron un total de 179 lagartijas A. nebulosus, 78 antes (45 adultos, 33 juveniles) y 101 después (88 adultos, 13 juveniles) de Jova. Tras el paso del huracán, sólo se recapturaron 8 (10.3%) de los 78 individuos marcados previamente. En general, los datos de captura-marcaje-recaptura proporcionaron poco soporte a los modelos, siendo 3 los modelos más apoyados: el modelo con mayor soporte (AICc = 834, ΔAICc = 0, AICc Weight = 0.41) fue ϕ “sexo” p “tiempo” que indicó una probabilidad de supervivencia mayor en hembras (0.877 ± 0.017) que en machos (0.841 ± 0.022), además de una probabilidad de captura significativamente baja tras el paso del huracán con respecto a la probabilidad de captura promedio (fig. 7); el segundo modelo con mayor soporte (AICc = 834, ΔAICc = 0.2, AICc Weight = 0.37) fue ϕ “continuo” p “tiempo” que es irrelevante para el propósito del estudio, ya que no permite identificar diferencias entre sexos, o diferencias en el tiempo; y el tercer modelo con mayor soporte (AICc = 836, ΔAICc = 2.1, AICc Weight = 0.14) fue ϕ “tiempo” p “tiempo”, que indicó menor probabilidad de supervivencia, similar tras el paso de Jova con respecto a la probabilidad de supervivencia promedio.
La densidad de U. bicarinatus aumentó significativamente tras el paso del huracán Jova (F = 8, df = 1, p = 0.01), siendo mayor después (11.4 ind/ha ± 2.5) que antes del huracán (4.6 ind/ha ± 0.9). La interacción Jova-estacionalidad sobre la densidad de U. bicarinatus fue marginal (F = 3.6, df = 1, p = 0.07) y se manifestó en un incremento mayor entre las temporadas de lluvias (de 4.5 ind/ha ± 1.4 a 16.6 ind/ha ± 3.6) que entre las de secas (de 4.7 ind/ha ± 1.2 a 7 ind/ha ± 2.6). El uso de la percha por parte de U. bicarinatus no cambió tras el paso de Jova, ni en altura (F = 0.02, df = 1, p = 0.88), ni en diámetro de la percha (χ2 = 0.02, df = 1, p = 0.63).
En el caso de A. lineattissimus, la densidad poblacional fue similar previo (8.7 ind/ha ± 2.5) y posterior (9.3 ind/ha ± 2.4) a Jova (F = 0.001, df = 1, p = 0.97); sin embargo, el efecto Jova-estacionalidad fue marginal (F = 3.2, df = 1, p = 0.08), aumentando entre las estaciones secas (de 6.3 ind/ha ± 1.1 a 12.5 ind/ha ± 3.7) y disminuyendo entre las estaciones de lluvias (de 11.5 ind/ha ± 5.4 a 5.5 ind/ha ± 2.4).
Discusión
Los resultados indicaron que el paso del huracán Jova (categoría 2, vientos de hasta 342 km/h) afectó a un tercio de los árboles muestreados en un área de 1,875 m2. La poca extensión del área y la topografía simple de ISA que alcanza una altura máxima de 21 m snm (Conanp, 2008) permitieron el alto impacto que tuvo Jova sobre su vegetación, ya que la isla no cuenta con una topografía pronunciada que proteja a la vegetación de los fuertes vientos provenientes de huracanes como en el caso de la isla de Guanaja, Honduras (Fickert, 2018).
Adicionalmente, las características de los árboles influyeron sobre el daño sufrido por los mismos. De manera general, los árboles más altos fueron los que sufrieron mayor daño tal como se esperaba en la hipótesis 1, el cual es un patrón que ocurre con frecuencia (Ugarte et al., 2006) y que claramente se debe a que los árboles más altos se encuentran más expuestos a los vientos (Boutet y Weishampel, 2003). En el caso de los árboles de ISA, la ruptura de los troncos fue el daño más relacionado con la altura, lo cual podría deberse a la estructura biomecánica que les proporciona mayor o menor resistencia (Méndez-Alonzo et al., 2012). Por otro lado, las correlaciones indicaron que los árboles con mayor DAP fueron los que sufrieron mayor daño, tanto por ruptura como por desenraizamiento. En el caso del daño por ruptura, el resultado concuerda parcialmente con lo reportado por Lewis y Bannar-Martin (2012) en el bosque seco de Madagascar, quienes indican que los árboles de diámetros intermedio son los que sufren más este tipo de daño tras el paso del ciclón Fanele en 2009. Asimismo, la correlación entre el desenraizamiento con el DAP concuerda con los resultados de Lewis y Bannar-Martin (2012), es decir, a mayor DAP hay mayor daño por desenraizamiento. El daño que causan los vientos provenientes de este tipo de fenómeno natural afecta a los árboles dependiendo del tipo de vegetación, ya que por ejemplo en los bosques templados son los árboles de menor DAP los que sufren mayor daño (Busing et al., 2009).
Contrario a la hipótesis 2, en la que esperábamos un decremento en la densidad de lagartijas, los resultados indicaron un incremento poblacional tanto de A. nebulosus como de U. bicarinatus, aunque no de A. lineattissimus tras el paso de Jova. Este patrón provocado por Jova parece generalizado, ya que en diversos sitios de la región continental se observó un incremento de lagartijas posterior al huracán, siendo A. nebulosus una de las especies con mayor incremento (Suazo-Ortuño et al., 2018). Este resultado sugiere que Jova no presentó la intensidad necesaria para afectar la densidad poblacional de las lagartijas en ISA, a diferencia de huracanes más intensos como Hugo (categoría 4) o Lili (categoría 3) que provocaron la disminución o desaparición de poblaciones de lagartijas insulares por arrastre de los individuos (Reagan, 1991; Spiller y Schoener, 2007; Spiller et al., 1998).
Los huracanes catalogados como intensos suelen lavar (arrastrar fuera del terreno o isla) las poblaciones de lagartijas, incluyendo sus puestas, por lo que inducen la extinción de poblaciones insulares (Schoener et al., 2001). Tras el paso de Jova, se registraron nuevos individuos adultos dentro de las parcelas de muestreo y se recapturaron pocos adultos marcados previo al huracán, por lo que se sugiere que la intensidad de Jova solo permitió el arrastre de individuos dentro y fuera de las parcelas, pero no fuera de la isla (al menos de forma importante), es decir, probablemente el huracán provocó una revoltura de la población de A. nebulosus en ISA, como ha ocurrido en aves (Johnson y Winker, 2010). Adicionalmente a la intensidad intermedia de Jova, la talla promedio de A. nebulosus (43 mm LHC; García y Ceballos, 1994), U. bicarinatus (45 mm LHC; García y Ceballos, 1994) y A. lineattissimus (112 mm LHC; García y Ceballos, 1994) podrían ser un factor que contribuyó a su permanencia en ISA, puesto que de acuerdo con Spiller et al. (1998), las lagartijas de tamaño intermedio (39 – 52 mm LHC) tienen una alta probabilidad de permanencia ante huracanes de intensidad moderada.
El incremento en la densidad poblacional de lagartijas en ISA indicado por nuestros resultados podría deberse a diversos factores, como son mayor productividad en el sistema, concentración de individuos en las perchas, o la ampliación del campo visual. El aumento en la precipitación provocado por Jova pudo reflejarse en el incremento de la productividad del BTC de ISA, ya que dicha productividad responde positivamente a la cantidad de lluvia (Maass et al., 2017). Esto solo tendría sentido si el huracán no presentó la intensidad necesaria para arrastrar los recursos alimenticios de las lagartijas, como se observó tras el paso del huracán Michelle que dañó severamente a los insectos defoliadores que podrían incluirse en la dieta de las lagartijas (Spiller y Schoener, 2007). Una alta productividad del sistema podría influir positivamente en diversos aspectos reproductivos de las lagartijas (Hall et al., 2018; Ramírez-Bautista y Vitt, 1997), induciendo un incremento de sus poblaciones. Por otro lado, la pérdida de árboles pudo conducir a las lagartijas A. nebulosus a usar los árboles que quedaron en pie, por lo que la detección de individuos se pudo incrementar y con ello la densidad estimada, dado que la pérdida de árboles puede incrementar el campo visual del observador. Al respecto, Reagan (1991) indicó que la modificación provocada en la vegetación por el huracán Hugo condujo a restricciones en el uso de la percha de lagartijas del género Anolis. Además del incremento en la densidad poblacional de lagartijas, Jova indujo el incremento de la talla y peso de A. nebulosus, lo que contrastó con la hipótesis 2. Los efectos positivos de Jova sobre las poblaciones de lagartijas insulares de nuestro estudio se deben muy probablemente a la intensidad intermedia del huracán, a diferencia de Floyd (categoría 4) que redujo la abundancia inmediata de artrópodos y de lagartijas tras el paso del huracán (Schöner y Spiller, 2006). El claro efecto negativo de Jova sobre la vegetación de ISA, conlleva una disminución de las perchas para las lagartijas arborícolas A. nebulosus y U. bicarinatus, es decir, más lagartijas por percha, lo que probablemente incrementó la competencia inter e intraespecífica por este recurso (Delaney y Warner, 2017; Zeng et al., 2014). Sin embargo, el incremento en la talla y peso de las lagartijas A. nebulosus podría sugerir suficiente viabilidad del recurso alimenticio o incluso, el incremento de dicho recurso como sugiere Reagan (1991) con los defoliadores. Asimismo, la pérdida de árboles permite la creación de claros que conducen a un incremento de temperatura y luz (Nicoletto, 2013; Turton y Siegenthaler, 2004), principalmente durante la temporada de lluvias que es cuando la vegetación presenta follaje, lo cual podría ser aprovechado por las lagartijas para termorregular y procesar de mejor manera los recursos alimenticios disponibles en su condición de ectotermos (Gienger et al., 2002; Schutle, 2015).
A pesar del incremento en la densidad poblacional indicada por nuestros resultados, la probabilidad de supervivencia de las lagartijas A. nebulosus no presentó una diferencia significativa tras el paso de Jova con respecto a la probabilidad de supervivencia promedio (0.72 posterior a Jova a 0.87 promedio; fig. 7). En este caso, se debe tomar en cuenta el poco soporte a los modelos obtenidos con los datos de captura-marcaje-recaptura; sin embargo, la tendencia en la reducción de la probabilidad de supervivencia contrastaría con el incremento de la densidad poblacional. Para explicar este patrón sugerimos 2 posibles escenarios. En el primero, los individuos capturados después de Jova pertenecen a una nueva generación que llegó pronto a la edad adulta debido a su alta tasa de crecimiento (Siliceo-Cantero y García, 2014), y cuyas puestas soportaron las condiciones de Jova incluyendo una poco probable inundación marina (Losos et al., 2003), ya que la altura de ISA es de 21 m snm (Conanp, 2008) y contrasta con la altura de 3 m snm de la isla, lo cual permitió su inundación según Schoener et al. (2001). En el segundo escenario, ocurre una revoltura de la población dentro de ISA debido al arrastre de individuos dentro y fuera de las parcelas como resultado de los fuertes vientos y la cantidad de precipitación, explicando el alto número de adultos sin marcaje capturados. Adicionalmente, la concentración de individuos en las perchas que se mantuvieron en pie tras el paso de Jova, y la mayor visibilidad por la pérdida de árboles, podrían estar jugando un papel importante en estos resultados.
Contrario a la hipótesis 2 sobre la reducción en la altura de la percha usada por A. nebulosus, esta lagartija perchó significativamente más alto. Este resultado también contrasta con lo observado en lagartijas del género Anolis tras el paso del huracán Hugo en Puerto Rico, las cuales disminuyen la altura de percha tras el achaparramiento del dosel (Reagan, 1991). Adicionalmente, A. nebulosus usó perchas significativamente más delgadas. Las modificaciones en ambos parámetros podrían estar relacionadas con la pérdida de un tercio de los elementos arbóreos en ISA tras Jova. La relación mostrada entre el daño a los árboles y los atributos de los mismos, indicaron mayor daño para los árboles más altos y de mayor DAP, por ello, las lagartijas A. nebulosus usaron perchas de menor diámetro. Sin embargo, la misma pérdida de árboles podría incrementar la competencia intraespecífica en una isla tan pequeña como lo es ISA, para la cual es de suma importancia el uso de perchas altas para mantener y vigilar el territorio (Kattan, 2007; Stamps, 1982).
Por otro lado, U. bicarinatus parece ser resiliente a los cambios en la estructura del hábitat. Esta especie se caracteriza por usar perchas altas (ramas con diámetros similares) y presentar picos de actividad durante las horas del día con mayor temperatura (García, 2008), por lo que es posible que la pérdida de árboles tras el paso de Jova no afectara la forma en la cual usa las perchas esta especie.
Los resultados indicaron que el huracán Jova afectó la respuesta de las especies de lagartijas a la estacionalidad, particularmente en cuanto a la densidad de lagartijas, la talla y el peso. Previo a Jova, hubo una reducción de secas a lluvias en las 3 variables, mientras que posterior a Jova sucede lo contrario. Este resultado podría sugerir que el aumento de la precipitación provocado por Jova fue benéfico para el sistema de manera inmediata, incrementando la cantidad del recurso alimenticio (e.g., artrópodos) para las lagartijas, lo que se reflejaría en su reproducción (mejores condiciones reproductivas a mayor productividad; Hall et al. [2018]) y condiciones corporales (a mayor productividad del sistema, mayor crecimiento; Brown et al. [2017]).
Concluyendo, solo la primera de las 3 hipótesis planteadas fue soportada por nuestros resultados. Esta se refiere a un daño considerable que recibiría la vegetación de la isla y que podría deberse a una topografía simple y al tamaño pequeño que presenta ISA. Así, nuestros resultados indicaron que aproximadamente 30% de los árboles presentaron algún daño tras el paso de Jova, incrementándose dicho daño en función de la altura y el DAP de los árboles.
Por otro lado, la hipótesis 2 no se cumplió, ya que contrario a lo esperado, se tuvo una respuesta inmediata positiva de las lagartijas en ISA ante un huracán de intensidad intermedia como Jova, a diferencia de la respuesta ante huracanes más intensos que causan la pérdida de poblaciones enteras, o cambios que pueden contribuir sobre el actuar de la selección natural (Donihue et al., 2018; Spiller y Schoener, 2007; Spiller et al., 1998). Esto nos resulta sorpresivo, puesto que Jova presentó fuertes vientos de hasta 342 km/h que causaron gran destrucción de la vegetación de ISA. A pesar de ello, las lagartijas respondieron a Jova incrementando su densidad poblacional, talla y peso corporal, tal vez como resultado de un incremento en la productividad del sistema inducido por la elevada precipitación. Es importante mencionar que las variables en nuestro estudio (densidad poblacional, probabilidad de supervivencia, altura y diámetro de la percha, y talla y peso corporal de las lagartijas) se midieron solo para A. nebulosus, mientras que para U. bicarinatus solo se midieron la densidad, altura y diámetro de la percha, y para A. lineattissimus solo la densidad. Por lo anterior, nuestras conclusiones podrían aplicarse solo a A. nebulosus. Sin embargo, también debe tomarse en cuenta que en las variables medidas para cada una de las especies se observó un efecto positivo de Jova, sugiriendo un patrón similar para las diversas especies de lagartijas en ISA.
La hipótesis 3 fue parcialmente cumplida, puesto que los resultados indicaron un cambio en el comportamiento de la transición estacional posterior a Jova, pero dicho cambio no fue una atenuación, sino un incremento de la diferencia entre secas y lluvias. Esto podría deberse principalmente a que Jova representó un incremento en la productividad de ISA, y no tuvo la intensidad necesaria como para arrastrar los recursos fuera de la isla como inicialmente pensamos.
Cabe mencionar que muestreos recientes en ISA sugieren una reducción dramática de la población de lagartijas A. nebulosus y un incremento considerable de la población de lagartijas U. bicarinatus, esto podría ser el resultado de 2 factores: 1) durante 2011 y 2012, Hernández-Salinas et al. (2016) extrajeron al menos 77 lagartijas A. nebulosus de ISA, lo que podría tener un impacto considerable debido al reducido tamaño de la isla; y 2) en Octubre de 2015, el huracán Patricia, considerado como el más intenso de la historia en el hemisferio occidental (categoría 5), impactó la región.
Agradecimientos
Al programa de Posgrado en Ciencias Biológicas del Instituto de Biología, de la Universidad Nacional Autónoma de México y a la estación de Biología Chamela de IB-UNAM, y a dos revisores anónimos por sus observaciones y comentarios que contribuyeron a mejorar sustancialmente el manuscrito.
Referencias
Armstrong, D. P., Davidson, R. S., Perrott, J. K., Roygard, J. y Buchanan, L. (2005). Density-dependent population growth in a reintroduced population of North Island saddlebacks. Journal of Animal Ecology, 74, 160–170. https://doi.org/10.1111/j.1365-2656.2004.00908.x
Boutet, J. C. y Weishampel, J. F. (2003). Spatial pattern analysis of pre-and post-hurricane forest canopy structure in North Carolina, USA. Landscape Ecology, 18, 553–559. https://doi.org/10.1093/jpe/rtm003
Brown, G. P., Madsen, T. R. y Shine, R. (2017). Resource availability and sexual size dimorphism: differential effects of prey abundance on the growth rates of tropical snakes. Functional Ecology, 31, 1592–1599. https://doi.org/10.1111/1365-2435.12877
Bullock, S. H. (1986). Climate of Chamela, Jalisco, and trends in the south coastal region of Mexico. Archives for Meteorology, Geophysics, and Bioclimatology, Series B, 36, 297–316. https://doi.org/10.1007/bf02263135
Busing, R. T., White, R. D., Harmon, R. D. y White, R. D. (2009). Hurricane disturbance in temperate deciduous forest: patch dynamics, tree mortality, and coarse woody detritus. Plant Ecology, 201, 351–363. https://doi.org/10.1007/978-90-481-2795-5_26
Ceballos, G. y Miranda, A. (2000). Guía de campo de los mamíferos de la costa de Jalisco, México. México D.F.: Fundación Ecológica de Cuixmala, A.C./ Universidad Nacional Autónoma de México.
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2008). Programa de conservación y manejo: Santuario Islas de la Bahía de Chamela. Mexico D.F.: Semarnat.
Conery, I., Walsh, J. P. y Corbett, D. R. (2018). Hurricane Overwash and Decadal-Scale Evolution of a Narrowing Barrier Island, Ocracoke Island, NC. Estuaries and Coasts, 41, 1626–1642. https://doi.org/10.1007/s12237-018-0374-y
Delaney, D. M. y Warner, D. A. (2017). Adult male density influences juvenile microhabitat use in a territorial lizard. Ethology, 123, 157–167. https://doi.org/10.1111/eth.12586
Donihue, C. M., Herrel, A., Fabre, A. C., Kamath, A., Geneva, A. J., Schoener, T. W. et al. (2018). Hurricane-induced selection on the morphology of an island lizard. Nature, 560, 88. https://doi.org/10.1038/s41586-018-0352-3
Fickert, T. (2018). Better resilient than resistant-regeneration dynamics of storm-disturbed mangrove forests on the Bay Island of Guanaja (Honduras) during the first two decades after Hurricane Mitch (October 1998). Diversity, 10, 8. https://doi.org/10.3390/d10010008
Folmer, M.J., DeMaria, M., Ferraro, R., Beven, J., Brennan, M., Daniels, J. et al. (2015). Satellite tools to monitor and predict Hurricane Sandy (2012): current and emerging products. Atmospheric Research, 166, 165–181. https://doi.org/10.1016/j.atmosres.2015.06.005
Funderburk, W. R., Carter, G. A. y Anderson, C. P. (2016). Evaluating the influence of elevation and impact of Hurricane Katrina on radial growth in slash pine (Pinus elliottii var. elliottii Engelm) on Cat Island, Mississippi, USA. Journal of Coastal Research, 32, 483–489. https://doi.org/10.2112/jcoastres-d-15-00038.1
García, A. (2008). The use of habitat and time by lizards in a tropical deciduous forest of western Mexico. Journal of Studies on Neotropical Fauna and Environment, 43, 107–115. https://doi.org/10.1080/01650520701735282
García, A. y Ceballos, G. (1994). Guía de campo de los reptiles y anfibios de la costa de Jalisco, México. México D.F.: Fundación Ecológica de Cuixmala, A.C./ Instituto de Biología, Universidad Nacional Autónoma de México.
Gienger, C. M., Beck, D. D., Sabari, N. C. y Stumbaugh, D. L. (2002). Dry season habitat use by lizards in a tropical deciduous forest of western Mexico. Journal of Herpetology, 36, 487–490. https://doi.org/10.1670/0022-1511(2002)036[0487:dshubl]2.0.co;2
Hall, J. M., Buckelew, A., Lovern, M., Secor, S. M. y Warner, D. A. (2018). Seasonal Shifts in Reproduction Depend on Prey Availability for an Income Breeder. Physiological and Biochemical Zoology, 91, 1129–1147. https://doi.org/10.1086/700341
Hernández-Salinas, U., Ramírez-Bautista, A. y Cruz-Elizalde, R. (2016). Variation in Feeding Habits of the Arboreal Lizard Anolis nebulosus (Squamata: Dactyloidae) from Island and Mainland Populations in Mexican Pacific. Copeia, 104, 831–837. https://doi.org/10.1643/ce-16-390
Hosseini, S. R., Scaioni, M. y Marani, M. (2018). On the influence of global warming on Atlantic hurricane frequency. In 2018 ISPRS TC III Mid-Term Symposium on Developments, Technologies and Applications in Remote Sensing. International Society for Photogrammetry and Remote Sensing, 42, 527–532. https://doi.org/10.5194/isprs-archives-xlii-3-527-2018
Jáuregui, E. (2003). Climatology of landfalling hurricanes and tropical storms in Mexico. Atmósfera, 16, 193–204.
Johnson, A. B. y Winker, K. (2010). Short-term hurricane impacts on a neotropical community of marked birds and implications for early-stage community resilience. Plos One, 5, e15109. https://doi.org/10.1371/journal.pone.0015109
Kattan, G. (2007). Sleeping perch selection in the lizard Anolis ventrimaculatus. Biotropica, 16, 328–329. https://doi.org/10.2307/2387945
Knutson, T. R., Sirutis, J. J., Zhao, M., Tuleya, R. E., Bender, M., Vecchi, G. A. et al. (2015). Global projections of intense tropical cyclone activity for the late twenty-first century from dynamical downscaling of CMIP5/RCP4. 5 scenarios. Journal of Climate, 28, 7203–7224. https://doi.org/10.1175/jcli-d-15-0129.1
Lebreton, J. D., Burnham, K. P., Clobert, J. y Anderson, D. R. (1992). Modeling survival and testing biological hypotheses using marked animals: A unified approach with case studies. Ecological Monographs, 62, 67–118. https://doi.org/10.2307/2937171
Lewis, R. J. y Bannar–Martin, K. H. (2012). The impact of cyclone Fanele on a tropical dry forest in Madagascar. Biotropica, 44, 135–140. https://doi.org/10.1111/j.1744-7429.2011.00799.x
Lister, B. C., y García, A. (1992). Seasonality, predation, and the behaviour of a tropical mainland anole. Journal of Animal Ecology, 61, 717–733. https://doi.org/10.2307/5626
Losos, J. B., Schoener, T. W. y Spiller, D. A. (2003). Effect of immersion in seawater on egg survival in the lizard Anolis sagrei. Oecologia, 137, 360–362. https://doi.org/10.1007/s00442-003-1363-8
Lugo, A. E. (2008). Visible and invisible effects of hurricanes on forest ecosystems: an international review. Austral Ecology, 33, 368–398. https://doi.org/10.1111/j.1442-9993.2008.01894.x
Maass, M., Ahedo-Hernández, R., Araiza, S., Verduzco, A., Martínez-Yrízar, A., Jaramillo, V. J. et al. (2017). Long-term (33 years) rainfall and runoff dynamics in a tropical dry forest ecosystem in western Mexico: management implications under extreme hydrometeorological events. Forest Ecology and Management, 426, 7–17. https://doi.org/10.1016/j.foreco.2017.09.040
MacKenzie, D. I., Nichols, J. D., Royle, J. A., Pollock, K. H., Bailey, L. y Hines, J. E. (2017). Occupancy estimation and modeling: inferring patterns and dynamics of species occurrence. Elsevier. https://doi.org/10.1016/b978-0-12-407197-1.00018-1
Méndez-Alonzo, R., Paz, H., Zuluaga, R. C., Rosell, J. A. y Olson, M. E. (2012). Coordinated evolution of leaf and stem economics in tropical dry forest trees. Ecology, 93, 2397–2406. https://doi.org/10.1890/11-1213.1
Navarro-García, J., García, A. y Mendéz-De la Cruz, R. (2008). Estacionalidad, eficiencia termoreguladora de Aspidocelis lineatissima (Sauria: Teiidae) y la calidad térmica del bosque tropical caducifolio en Chamela, Jalisco, México. Revista Mexicana de Biodiversidad, 79, 413–419. http://dx.doi.org/10.22201/ib.20078706e.2008.002.559
Nicholas, V. L. B. y Walker, L. R. (1991). Summary of the Effects of Caribbean Hurricanes on Vegetation. Biotropica, 23, 442–447. https://doi.org/10.2307/2388264
Nicoletto, P. F. (2013). Effects of hurricane Rita on the herpetofauna of Village Creek State Park, Hardin County, Texas. Southwestern Naturalist, 58, 64–69. https://doi.org/10.1894/0038-4909-58.1.64
Olguín-López, J. L., Guevara-Gutiérrez, R. D., Romero, J. M. R. y Rodríguez, M. R. A. (2015). Los efectos de “Jova” en el municipio de Autlán de Navarro, Jalisco, México: un caso histórico. Revista Iberoamericana para la Investigación y el Desarrollo Eductaivo, 2, 1–19. https://doi.org/10.23913/ride.v2i4.44
Pavelka, M. S., McGoogan, K. C. y Steffens, T. S. (2007). Population size and characteristics of Alouatta pigra before and after a major hurricane. International Journal of Primatology, 28, 919–929. https://doi.org/10.1007/s10764-007-9136-6
Ramírez-Bautista, A. y Vitt, L. J. (1997). Reproduction in the lizard Anolis nebulosus (Polychrotidae) from the Pacific coast of Mexico. Herpetologica, 53, 423–431.
Reagan, D. P. (1991). The response of Anolis lizards to hurricane-induced habitat changes in a Puerto Rican rain forest. Biotropica, 23, 468–474. https://doi.org/10.2307/2388268
Renton, K., Salinas-Melgoza, A., Rueda-Hernández, R. y Vázquez-Reyes, L. D. (2017). Differential resilience to extreme climate events of tree phenology and cavity resources in tropical dry forest: Cascading effects on a threatened species. Forest Ecology and Management, 426, 164–175. https://doi.org/10.1016/j.foreco.2017.10.012
Rittenhouse, C. D., Pidgeon, A. N., Albright, T. P., Culbert, T. P., Clayton, M. K., Flather, C. H. et al. (2010). Avifauna response to hurricanes: regional changes in community similarity. Global Change Biology, 16, 905–917. https://doi.org/10.1111/j.1365-2486.2009.02101.x
Schoener, T. W. y Spiller, D. A. (2006). Nonsynchronous recovery of community characteristics in island spiders after a catastrophic hurricane. Proceeding of the National Academy of Science, U.S.A., 103, 2220–2225. https://doi.org/10.1073/pnas.0510355103
Schoener, T. W., Spiller, D. A. y Losos, J. B. (2001). Natural restoration of the species-area relation for a lizard after a hurricane. Science, 294, 1525–1528. https://doi.org/10.1126/science.1064396
Schoener, T. W., Spiller, D. A. y Losos, J. B. (2004). Variable ecological effects of hurricanes: the importance of seasonal timing for survival of lizards on Bahamian islands. Proceeding of the National Academy of Science, U.S.A., 101, 177–81. https://doi.org/10.1073/pnas.0306887101
Siliceo-Cantero, H. H. y García, A. (2013). Anolis nebulosus (Clouded Anole). Predation. Herpetological Review, 44, 137.
Siliceo-Cantero, H. H. y García, A. (2014). Differences in growth rate, body condition, habitat use and food availability between island and mainland lizard populations of Anolis nebulosus in Jalisco, Mexico. Journal of Tropical Ecology, 30, 493–501. https://doi.org/10.1017/s0266467414000297
Spiller, D. A., Losos, J. B. y Schoener, T. W. (1998). Impact of a catastrophic hurricane on island populations. Science, 281, 695–697. https://doi.org/10.1126/science.281.5377.695
Spiller, D. A. y Schoener, T. W. (2007). Alteration of island food-web dynamics following major disturbance by hurricanes. Ecology, 88, 37–41. https://doi.org/10.1890/0012-9658(2007)88[37:aoifdf]2.0.co;2
Stamps, J. A. (1982). The Relationship between ontogenetic habitat shifts, competition and predator avoidance in a juvenile lizard (Anolis aeneus). Behavioral Ecology and Sociobiology, 12, 19–33. https://doi.org/10.1007/bf00296929
Suazo-Ortuño, I., Urbina-Cardona, J. N., Lara-Uribe, N., Marroquín-Páramo, J., Soto-Sandoval, Y., Rangel-Orozco, J. et al. (2018). Impact of a hurricane on the herpetofaunal assemblages of a successional chronosequence in a tropical dry forest. Biotropica, 50, 649–663. https://doi.org/10.1111/btp.12544
Temeles, E. J. y Bishop, G. A. (2019). A hurricane alters pollinator relationships and natural selection on an introduced island plant. Biotropica, 51, 129–138. https://doi.org/10.1111/btp.12634
Turton, S. M. y Siegenthaler, D. T. (2004). Immediate impacts of a severe tropical cyclone on the microclimate of a rain-forest canopy in north-east Australia. Journal of Tropical Ecology, 20, 583–586. https://doi.org/10.1017/s0266467404001622
Ugarte, C. A., Brandt, L. A., Melvin, S., Mazzotti, F. J. y Rice, K. G. (2006). Hurricane impacts to tree islands in Arthur – Marshall loxahatchee national wildlife refuge, Florida. Southwestern Naturalist, 5, 737–746. https://doi.org/10.1656/1528-7092(2006)5[737:hittii]2.0.co;2
Valtierra-Azotla, M., Lister, B. y García, A. (2010). Behavioral responses to seasonality by two Sceloporine lizard species from a tropical dry forest. Animal Biology, 60, 97–113. https://doi.org/10.1163/157075610×12610595764291
Walker, L. R. (1991). Summary of the effects of Caribbean hurricanes on vegetation. Biotropica, 23, 442–447. https://doi.org/10.2307/2388264
Webster, P. J., Holland, G. J., Curry, J. A. y Chang, H. R. (2005). Changes in tropical cyclone number, duration, and intensity in a warming environment. Science, 309, 1844–1846. https://doi.org/10.1126/science.1116448
White, G. C. y Burnham, K. P. (1999). Program MARK: Survival estimation from populations of marked animals. Bird Study Supplement, 46, 120–138. https://doi.org/10.1080/00063659909477239
Wiley, J. W. y Wunderle, J. M. (1993). The effects of hurricanes on birds, with special reference to Caribbean islands. Bird Conservation International, 3, 319–349. https://doi.org/10.1017/s0959270900002598
Zeng, Z. G., Bi, J. H., Li, S. R., Chen, S. Y., Pike, D. A., Gao, Y. et al. (2014). Effects of habitat alteration on lizard community and food web structure in a desert steppe ecosystem. Biological Conservation, 179, 86–92. https://doi.org/10.1016/j.biocon.2014.09.011
Zhao, M., y Held, I. M. (2012). TC-permitting GCM simulations of hurricane frequency response to sea surface temperature anomalies projected for the late-twenty-first century. Journal of Climate, 25, 2995–3009. https://doi.org/10.1175/jcli-d-11-00313.1


