1580 años de impacto humano y cambio climático en la dinámica del bosque de Pinus-Quercus-Abies en el centro-occidente de México
Ana Patricia del Castillo-Batista a, Blanca Lorena Figueroa-Rangel b, *, Socorro Lozano-García c, Miguel Olvera-Vargas b y Ramón Cuevas-Guzmán b
a Posgrado en Biosistemática, Ecología y Manejo de Recursos Naturales y Agrícolas, Universidad de Guadalajara, Centro Universitario de la Costa Sur, Av. Independencia Nacional Núm. 151, 48900 Autlán de Navarro, Jalisco, México
b Universidad de Guadalajara, Centro Universitario de la Costa Sur, Av. Independencia Nacional Núm. 151, 48900 Autlán de Navarro, Jalisco, México
c Departamento de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México, Circuito Escolar Universitario, Cd. Universitaria, Delegación Coyoacán, 04510 Ciudad de México, México
*Autor para correspondencia: bfrangel@cucsur.udg.mx (B.L. Figueroa-Rangel)
Resumen
Se presentan los resultados de la reconstrucción de la vegetación y el ambiente de los últimos 1,580 años en un bosque dominado por Pinus-Quercus-Abies en el centro-occidente de México. Se utilizaron técnicas paleoecológicas que incluyen el análisis polínico, partículas de microcarbón fósil y elementos geoquímicos, se documentan los cambios ocurridos en la composición taxonómica de la vegetación, su relación con actividades antropogénicas y el cambio climático. La cronología se determinó con base en el análisis de 14C y un modelo de edad-profundidad por interpolación lineal. Se realizó un análisis de redundancia (RDA) para identificar patrones de ordenación en la composición de los taxones y las variables ambientales y un análisis de correlación cruzada entre los taxones y las partículas de microcarbón fósil para determinar su sincronía. Los resultados del registro palinológico indican que Pinus ha sido el taxón dominante en gran parte de la secuencia, acompañado de taxones codominantes como Quercus, Abies y Alnus. El análisis de correlación cruzada indicó que existe una correlación positiva con Pinus y Ternstroemia, ambos en sincronía con las partículas de microcarbón fósil. El RDA muestra que Pinus, Asteraceae, Poaceae, Phaseolus y Zea se encuentran asociados a incendios forestales. También fueron identificados 2 eventos de cambio climático como la Anomalía Climática Medieval (ACM) (~950-1400 d.C.) y la Pequeña Edad de Hielo (PEH) (~1350-1850 d.C.). Las principales respuestas de la vegetación fueron la expansión del bosque de pino en la primera parte y su disminución, al final de la ACM. Durante la PEH la vegetación fue fluctuante en la composición de taxones. La evidencia del registro polínico indica la existencia de impacto humano por la presencia de taxones culturales como Amaranthaceae, Asteraceae, Phaseolus y Zea a partir de ~370 d.C. al presente. La evidencia señala patrones complejos en las respuestas de la vegetación durante los periodos de cambio ambiental, por lo que pueden ofrecer nuevos escenarios para evaluar los impactos del cambio climático a nivel local.
Palabras clave:
Anomalía Climática Medieval; Pequeña Edad de Hielo; Paleoecología; Macropartículas de carbón fósil; Polen fósil
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
1580 years of human impact and climate change on the dynamics of a Pinus-Quercus-Abies forest in west-central Mexico
Abstract
The present research involved vegetation and environmental reconstruction of a Pinus-Quercus-Abies dominated forest in west-central Mexico over the last 1,580 years. Paleoecological techniques such as the analysis of pollen, micro-fossil charcoal particles, and geochemical elements were used to document changes in the taxonomic composition of vegetation and its relationship with anthropogenic activities and climate change. Chronology was determined using 14C dating and the age-depth model was constructed by linear interpolation. A redundancy analysis (RDA) was performed to identify patterns in composition of taxa and environmental variables and a cross-correlation analysis between taxa and micro fossil charcoal particles to determine synchrony. Results of the palynological record indicate that Pinus has been the dominant taxon throughout the sequence, together with codominant taxa such as Quercus, Abies and Alnus. Cross-correlation analysis indicated a positive correlation with Pinus and Ternstroemia, both in synchrony with micro-fossil charcoal. RDA showed that Pinus, Asteraceae, Poaceae, Phaseolus and Zea are associated with forest fires. Two events of climate change were identified: the Medieval Climatic Anomaly (MCA) (~ 950-1400 AD) and the Little Ice Age (LIA) (~ 1350-1850 AD). Main vegetation responses were the expansion of pine forest in the beginning of the ACM and a decrease at the end of this period; throughout the PEH, vegetation fluctuated in the composition of taxa. Pollen of cultural taxa such as Amaranthaceae, Asteraceae, Phaseolus, and Zea suggests the existence of human impact from ~1,580 AD to the present. The evidence points to complex patterns in response of the vegetation during periods of environmental change, which may offer new scenarios for assessing the impacts of local
climate change.
Keywords:
Medieval Climatic Anomaly; Little Ice Age; Paleoecology; Charcoal particles; Fossil pollen
Estudios paleoecológicos (e.g., Lozano-García et al., 2007; Ortega-Rosas et al., 2008) señalan que, durante el Holoceno temprano, en respuesta al calentamiento que caracterizó este periodo, se presentó un aumento en la expansión de los bosques de Pinus en diversas regiones de México. Sin embargo, durante el Holoceno tardío se presentaron condiciones variables en el clima que condujeron a 2 principales anomalías: la Anomalía Climática Medieval (ACM) (950-1400 d.C.), que fue un periodo más cálido al promedio normal y la Pequeña Edad de Hielo (PEH) (1400-1900 d.C.), un periodo frío en general pero de mayor variabilidad sobre todo en latitudes medias y altas del Hemisferio Norte (Díaz et al., 2011). Los registros de diversos sitios en México señalan que la respuesta ante la variabilidad climática es compleja y la duración en cada lugar puede ser diferente (Caballero et al., 2006; Lozano-García et al., 2007; Metcalfe et al., 2010; Rodríguez-Ramírez et al., 2015; Sosa-Nájera et al., 2010). Estos cambios ambientales se atribuyen a la variación en el balance de diferentes forzamientos globales, tal como la actividad solar (Bond et al., 2001), así como con los cambios en la posición media latitudinal de la zona intertropical de convergencia (ZITC) y El Niño-Oscilación del Sur (ENOS) (Metcalfe et al., 2000). También se ha documentado que algunos de los bosques de Pinus se han generado como consecuencia de impacto humano, derivados de las actividades del desmonte de tierras para la explotación forestal y la agricultura (Almeida-Leñero et al., 2005; Ludlow-Wiechers et al., 2005). Estudios arqueológicos aledaños al sitio de estudio en la Sierra de Cacoma del estado de Jalisco, indican que han sido ocupados desde ~1200 d.C., por poblaciones indígenas (Jardel, 1994; Jiménez, 1995; Kelly, 1945; Weigand, 1996). La evidencia arqueológica sugiere que la actividad humana ha estado presente en las partes bajas del sitio de estudio, y archivos históricos indican la existencia de diferentes localidades indígenas antes de la llegada de los españoles (Acuña, 1988). Excavaciones arqueológicas en Autlán, una localidad cercana ubicada al suroeste del sitio de la sierra de Cacoma, señalan la presencia de asentamientos humanos por lo menos desde 3,400 ± 200 años antes del presente (Kelly, 1980). Sin embargo, la cerámica prehispánica encontrada en dicha zona ubica la presencia de asentamientos humanos entre 800 y 500 años antes del presente (Kelly, 1945). Algunos documentos históricos del siglo XVI de la Nueva Galicia mencionan la llegada de los españoles en la región en 1524 d.C. y registran 22 asentamientos humanos en 1579 d.C. en tierras bajas alrededor del sitio de estudio (Acuña, 1988). Las poblaciones indígenas estaban comprendidas principalmente por grupos nahuas y otomíes asentadas en aldeas dispersas; se dedicaban a cultivar la tierra bajo la modalidad de roza, tumba y quema, producían maíz, frijol y calabaza, principalmente (Tetreault et al., 2011). Posterior a la llegada de los españoles se modificaron las actividades culturales de los indígenas nativos, se intensifica la explotación forestal, se introduce ganado bovino y se abren áreas por el aprovechamiento minero, la cual se convirtió en una actividad importante en la región durante esa época, por lo que las poblaciones humanas se establecieron cercanas a las minas (Jiménez,
1995).
Los bosques de Pinus-Quercus-Abies de alta montaña en el occidente de México, representan una fuente importante de recursos maderables y de servicios ambientales. Sin embargo, la sierra de Cacoma es una región que ha sido poco explorada y el conocimiento sobre la dinámica de la vegetación y la influencia de las actividades humanas a largo plazo es desconocido. Por lo tanto, es de interés conocer la historia en la distribución del bosque de Pinus-Quercus-Abies del sitio de estudio, la incidencia de las actividades humanas en el área, además de entender si la distribución actual del bosque de Pinus-Quercus-Abies es consecuencia de los procesos naturales como el cambio climático y los incendios forestales, e inclusive del incremento de los periodos de aridez del último milenio. Por lo que, el objetivo del trabajo es responder las preguntas: ¿Cuál ha sido la dinámica del bosque de Pinus-Quercus-Abies durante los últimos 1,580 años? ¿Cuál ha sido la influencia del cambio climático en su dinámica? ¿Es posible explicar la permanencia y expansión del bosque de Pinus-Quercus-Abies a partir de actividades antropogénicas durante los últimos 1,580 años?
Materiales y métodos
Como indicador de cambio paleoambiental se utilizó la susceptibilidad magnética, ésta mide el aporte de minerales magnéticos en el sedimento (Thompson y Oldfield, 1986) y se considera como un indicador de procesos de erosión. El análisis de susceptibilidad magnética se realizó cada centímetro a lo largo del núcleo de sedimento de 62 cm utilizando un dispositivo Bartington MS2 (Bartington Instruments, Witney Oxford, Reino Unido). Se determinó la pérdida progresiva por ignición (LOI) con el fin de estimar el contenido de materia orgánica y carbonatos del núcleo de sedimento utilizando las relaciones lineales entre los valores LOI y el contenido de materia orgánica y los carbonatos expresados en porcentaje (Dean, 1974). Este análisis se realizó a cada centímetro en todo el núcleo de sedimento, tomando muestras de 0.5 cm3 y mediante la pérdida de peso en las muestras al ser sometidos a altas temperaturas secuenciales (550 °C y 910 °C). La interpretación en los cambios de porcentaje de LOI está relacionada con eventos de mayor productividad y estabilización de la vegetación. El análisis de la composición de elementos químicos (e.g., As, Ca, Fe, Mo, Pb, Rb, Sr, Ti, Zn y Zr) se realizó por medio de fluorescencia de rayos X (FRX) a cada 3 centímetros en todo el núcleo de sedimento con un espectrómetro manual Thermo Scientific Niton XL3t. Los resultados del análisis de elementos se expresan en la concentración de partes por millón (ppm) los cuales se utilizan como indicadores de cambios ambientales. En particular el titanio es considerado indicador del registro de aporte de humedad (registro clástico), mientras que el Ca puede indicar momentos de menor sedimentación, por lo que valores altos de Ti y bajos de Ca indican un incremento en el aporte de sedimentos terrígenos
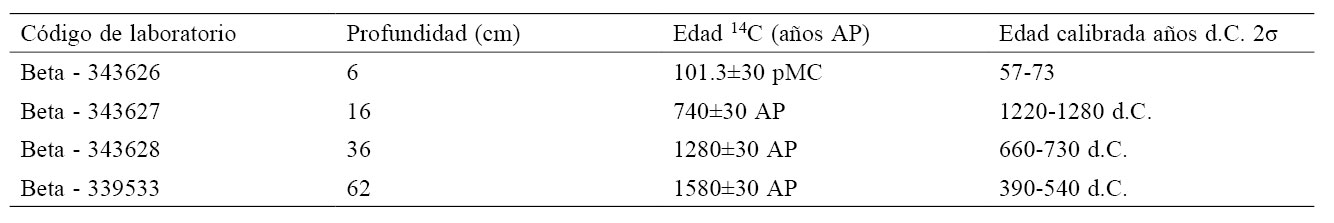
El modelo cronológico de edad-profundidad fue calculado por interpolación lineal basado en 4 dataciones radiométricas (14C; Beta Analytic de Florida, USA) a lo largo del núcleo de sedimento mediante la técnica de espectrometría de aceleración de masas (AMS, por sus siglas en inglés). Las fechas de radiocarbono fueron calibradas con el programa CALIB v5.02 (Stuiver y Reimer, 1993) e INTCAL09.14 (Reimer et al., 2009). Las zonas palinológicas se determinaron por el método de conglomerados CONISS (Constrained Cluster Analysis by Sum of Squares) (Grimm, 1987). Se aplicó un análisis de rarefacción [E(Tn)] para estimar la riqueza palinológica en cada muestra, esta técnica permite estimar la riqueza polínica que se esperaría obtener si los conteos polínicos tuvieran el mismo tamaño de muestra (Birks et al., 2012). Así mismo, se realizó el análisis de la tasa de cambio para identificar los cambios sincrónicos de la vegetación entre las muestras de polen, este análisis mide la disimilitud entre pares de muestras adyacentes y la diferencia temporal entre las muestras (Bennett, 2005). Todos los análisis se efectuaron con el programa PSIMPOLL 4.25 (Bennett, 2005). Se realizó un análisis de correlación cruzada entre 6 taxones leñosos (Pinus, Alnus, Parathesis, Quercus, Ternstroemia y Cupressaceae) y las partículas de microcarbón fósil, así como una correlación cruzada entre las partículas de microcarbón fósil y 2 taxones herbáceos (Cyperaceae y Asteraceae) con el programa estadístico R (R Core Team, 2013) para estimar el grado de correlación entre 2 muestras (taxón/microcarbón fósil) y determinar su sincronía.
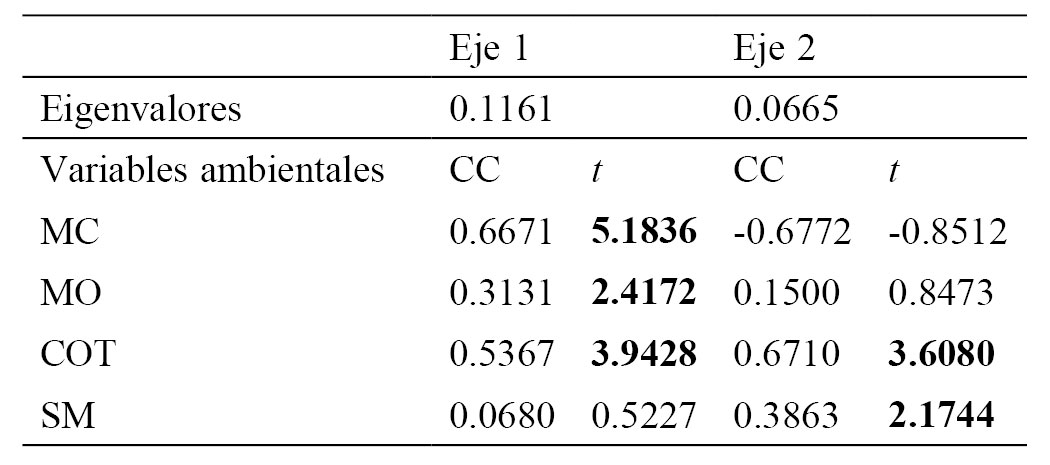
Se realizó un análisis de redundancia (RDA) con el fin de identificar las variables ambientales que mejor explican los patrones de ordenación observados en la composición de los taxones (Ter Braak, 1986). El RDA es un método directo de ordenación apropiado cuando se asume una relación monotónica entre los datos de ambas matrices (Ter Braak, 1986). Las variables ambientales incluidas en el análisis RDA fueron la susceptibilidad magnética, el contenido de materia orgánica, carbonatos y las partículas de microcarbón fósil. El análisis de ordenación se realizó con el programa CANOCO 5.0 (Ter Braak y Smilauer, 2002).
Resultados
El registro palinológico del bosque de Pinus-Quercus-Abies reveló 67 taxones con 3 zonas palinológicas de acuerdo al análisis de conglomerados (fig. 4). Pinus se presentó en toda la secuencia estratigráfica dominando el estrato arbóreo, principalmente durante la ACM y la PEH, con fluctuaciones constantes en su porcentaje a lo largo de toda la secuencia. En el estrato herbáceo las familias Asteraceae, Cyperaceae, Poaceae, Polypodiaceae, Aspleniaceae y Adiantaceae fueron los taxones más dominantes y al igual que Pinus estas familias estuvieron presentes en toda la secuencia estratigráfica. En el grupo de pteridofitas sensu lato Hymenophyllaceae fue la familia
Zona C-1 (62-59 cm, 370 d.C. – 405 d.C.). Los cambios más evidentes para esta zona se establecen con la dominancia de taxones arbóreos como Alnus, Pinus y Quercus con valores entre 10-25% (fig. 4); otros taxones arbóreos presentes en esta zona, aunque con bajos porcentajes son Abies, Fraxinus, Ostrya-Carpinus, Parathesis y Xylosma, así como las epífitas representadas por las familias Hymenophyllaceae y Bromeliaceae, además de Sphagnum, un tipo de musgo de hábitats húmedos como las turberas. Los taxones herbáceos son poco abundantes, con porcentajes menores al 10%, representado principalmente por las familias Poaceae, Cyperaceae, Asteraceae, Lamiaceae, Aspleniaceae, Polypodiaceae y Adiantaceae (fig. 4). Otros taxones herbáceos con porcentajes menores al 5% son Chenopodiaceae, Apiaceae, Onagraceae y Plantago. Por su parte, la riqueza palinológica registra valores altos para esta zona con el mayor pico a los 393 d.C., al igual que la tasa de cambio (fig. 5).
Zona C-2 (59-17 cm, 405-1183 d.C.). Al inicio de esta zona Pinus presenta porcentajes bajos (10%), con un aumento considerable a los 1048 d.C. y valores de hasta 35%. Por el contrario, Alnus se presenta como el taxón más dominante del estrato arbóreo al inicio de esta zona, pero rápidamente disminuye con menos del 10% alrededor de 520 d.C. Quercus se observa en toda la secuencia con valores fluctuantes menores al 10%, presentándose como codominante con Pinus y Alnus (fig. 4). Otros taxones arbóreos presentes en esta zona con porcentajes bajos son Abies, Cyathea, Fraxinus, Ostrya-Carpinus, Parathesis, Salix, Ternstroemia y Xylosma. Las familias presentes en esta zona son Cupressaceae, Lauraceae, Leguminosae, Myrtaceae y Rosaceae. En el estrato herbáceo las familias más abundantes son Asteraceae, Cyperaceae y Poaceae con valores de 10 a 25%. Zea y Phaseolus aparecen por primera vez en el registro estratigráfico en esta zona (fig. 4).
Fechas de radiocarbono del núcleo de sedimento CUM-N2 calibradas en edades antes del presente (AP) y en años calendáricos (d.C.).
Zona C-3 (17-0 cm, 1183 d.C.-Presente). Esta zona está dividida en 2 subzonas, la primera de ellas (C-3a) ocurre entre ~1183-1280 d.C., representa la transición entre la ACM y la PEH los taxones leñosos y las epífitas presentan porcentajes más elevados, 45% y 30% respectivamente, en comparación con la vegetación herbácea con 20%. Pinus tiene el mayor porcentaje de los taxones leñosos con el 25%, seguido de Alnus con el 15%, después se encuentra Ternstroemia, Xylosma y Quercus con el 5%, mientras que Hymenophyllaceae presenta 15% seguida de Sphagnum con 10% y Polypodiaceae con 5% (fig. 4). Con respecto a los taxones herbáceos en esta zona, el polen de la familia Asteraceae se mantiene abundante con el 15% en comparación con Cyperaceae y Poaceae que presentan menos del 5%. Por su parte, el registro palinológico de la zona C-3b muestra los porcentajes de Pinus como los más altos con fluctuaciones constantes a lo largo de la secuencia; Alnus y Quercus se mantienen en el estrato arbóreo con porcentajes variables. Otros taxones con bajos porcentajes (menores de 5%) son Abies, Fraxinus, Parathesis, Salix y Xylosma. La comunidad de herbáceas está representada principalmente por Asteraceae, Cyperaceae y Poaceae, aunque con porcentajes irregulares, Zea y Phaseolus continúan su registro en esta zona hasta el presente (fig. 4). En el conjunto de las pteridofitas, las esporas de Hymenophyllaceae son abundantes al inicio de la zona, decreciendo sus porcentajes al final de la secuencia, los helechos como Polypodiaceae, Adiantaceae y Aspleniaceae presentan porcentajes muy bajos. En cambio, Sphagnum presenta un ligero aumento al inicio de esta zona y disminuye al final de la misma. Esta zona muestra un incremento de la riqueza palinológica a los ~1350 d.C., registrando una disminución a los ~1491 d.C. con los valores más bajos de toda la secuencia, manteniendo fluctuaciones hasta el presente.
El registro de partículas de microcarbón fósil indica la ocurrencia de incendios durante los ca. 1,580 años que comprende la secuencia sedimentaria (fig. 5). Sin embargo, los incendios más intensos se registran en los periodos de ~647, 967, 1048, 1210 y 1491 d.C., a mediados de la zona C-2 y en la zona C-3a y C-3b. Los resultados de la susceptibilidad magnética indican valores altos en la base de la secuencia, pero se caracteriza por presentar grandes variaciones dentro de la misma, al igual que los valores de la materia orgánica, que registra valores bajos en la base de la secuencia en la zona C-1, con aumentos irregulares hasta el final de la secuencia en la zona C-3b (fig. 6).
Análisis de Redundancia (RDA) para las profundidades de núcleo de sedimento CUM-N2. Coeficientes de correlación (CC) de variables ambientales y valores de t (t); los números en negritas representan el valor crítico de (df ≥ α = 0.05) = 2.1.
Discusión
1993).
Las evidencias encontradas en el registro polínico y los resultados de la ordenación directa RDA, muestran relación entre los taxones asociados a los incendios forestales através de las partículas de microcarbón fósil y la susceptibilidad magnética. Estos taxones son Pinus, Asteraceae, Poaceae, Phaseolus y Zea. En cambio, otros taxones como Abies y el grupo de pteridofitas sensu lato y epífitas requieren condiciones de mayor humedad y menor radiación solar (Martínez-Salas y Ramos, 2014) y se encuentran relacionadas con la presencia de mayor concentración de materia orgánica. En este sentido, la concentración de las partículas de microcarbón fósil que son indicadoras de incendios forestales parecen incidir en la abundancia de Pinus. Por otro lado, es evidente que los incendios ocurridos en el área de estudio han sido recurrentes durante los últimos 1,580 años. Sin embargo, cuando se presentan periodos de mayor humedad, Pinus es reemplazado por Alnus y en menor medida por Quercus, Abies, Parathesis, Fraxinus, Cupressaseae, Rosaceae y Myrtaceae indicando la presencia de un bosque mixto con algunos elementos arbóreos de bosque mesófilo de montaña. Resultados similares fueron registrados en el centro de México (Almeida-Leñero et al., 2005) donde algunos intervalos de mayor humedad favorecieron el florecimiento de la vegetación mesófila. También en la Sierra de Manantlán los periodos de mayor humedad registrados en esta región favorecieron la reducción del bosque de pino (Figueroa-Rangel et al., 2008), pero al retornar los periodos de aridez el bosque de pino se recuperó. Asimismo, Cerano-Paredes et al. (2015) encontraron una relación significativa en este mismo sitio entre las sequías y los periodos de incendios, por lo que el fuego se considera un factor clave para la dominancia del bosque de Pinus, relacionado con los intervalos de aridez.
Los resultados de este estudio indican que la abundancia de Pinus ha fluctuado durante los últimos ca. 1,580 años en el sitio de estudio como respuesta a los periodos de sequía y frecuencia de incendios. A través del tiempo Pinus ha sido el taxón dominante en gran parte de la secuencia, acompañado de taxones codominantes como Quercus, Abies y Alnus. Durante el periodo de ~370-405 d.C. (Zona C-1) se muestran condiciones relativamente secas inferido con base en el registro de Ti, carbonatos, Ca y susceptibilidad magnética. Estos indicadores señalan que el bosque transitaba por un recambio de taxones impulsado principalmente como respuesta a los cambios ambientales.
El periodo de ~405-1183 d.C. (Zona C-2) correspondió con la ACM que, de acuerdo con la mayoría de las evidencias provenientes de diversos registros paleoclimáticos, este periodo fue generalmente más cálido que durante la PEH (Díaz et al., 2011; PAGES-2K-Consortium, 2013). Por ejemplo, la secuencia sedimentaria de la Laguna de Juanacatlán (Metcalfe et al., 2010) y el Lago de Santa María del Oro (Sosa-Nájera et al., 2010) ambos al occidente de México, muestran un patrón de condiciones secas durante la ACM con alternancia de condiciones húmedas al final de la misma. Este patrón también está presente en el bosque del área de estudio. El registro de Ca muestra condiciones secas en el periodo de ~800 a ~1075 d.C., que coincide con la primera parte de la ACM, donde se observa la expansión del bosque de pino. Este periodo de sequía ha sido identificado en varios registros de México (Hodell et al., 2001, 2005; Lozano-García et al., 2010) al final del periodo Clásico (850 a 950 d.C.), con implicaciones en el colapso de la cultura Maya (Gill et al., 2007). También hay evidencia de que las diferentes culturas mesoamericanas podrían haber enfrentado diferentes periodos de cambios abruptos en el clima (Bhattacharya et al., 2015; Rodríguez-Ramírez et al., 2015), siendo en la mayoría de ellas los intervalos de sequías el factor más importante en las principales transiciones culturales. En esta región del occidente de México los patrones normales del clima pueden ser modificados por el desplazamiento estacional de la ZITC. En el verano del Hemisferio Norte se desplaza hacia el norte, este movimiento de sur a norte puede resultar en periodos de intensas lluvias o sequías extremas (Magaña et al., 2003; Sosa-Nájera et al., 2010). Otros mecanismos que podrían crear condiciones más secas o húmedas en esta región son los cambios en la frecuencia e intensidad de ENOS (Magaña et al., 2003; Metcalfe et al., 2010). La frecuencia de los huracanes y tormentas tropicales del Pacífico también son condiciones que pueden modificar el equilibrio de humedad en esta área (Vargas-Rodríguez y Platt, 2012). En algunos registros, las sequías muestran un patrón recurrente o cíclico (Figueroa-Rangel et al., 2008) lo que podría explicar los valores fluctuantes de los registros geoquímicos en el sitio de estudio. Estas condiciones parecen incidir en la vegetación de forma diferencial en Pinus, Quercus y Abies.
La correlación positiva entre Pinus y Ternstroemia con las partículas de microcarbón fósil, que cambian en sincronía sugiere que los incendios forestales en el área de estudio están relacionados con algunos periodos de expansión en el bosque. Estudios actuales sobre la dinámica del bosque de Pinus (González-Espinosa et al., 1991; Richardson y Bond, 1991) indican que este tipo de vegetación se establece en claros abiertos posteriores a los incendios forestales de baja intensidad, desmontes agrícolas o la explotación forestal. Mientras que Alnus y Parathesis se establecieron 100 años después de un aumento en la concentración de las partículas de microcarbón fósil, lo que implica que son elementos arbóreos de etapas tardías en la sucesión en bosque de alta montaña.
Es posible que los cambios ambientales estén relacionados con la dinámica del bosque de Pinus-Quercus-Abies. En nuestro registro Abies presenta los mayores porcentajes de la secuencia durante ~1048-1075 d.C. Posteriormente, se infieren condiciones de mayor humedad en el periodo de ~1102 a ~1156, el cual se asemejan con las condiciones en el registro de Ti del lago de Juanacatlán y el lago de Santa María del Oro, ambos sitios ubicados al occidente de México (Metcalfe et al., 2010; Rodríguez-Ramírez et al., 2015; Sosa-Nájera et al., 2010). Con la disminución de Pinus ocurrió un aumento de otros taxones leñosos como Parathesis, Ostrya-Carpinus, Xylosma y las familias Cupressaceae y Leguminosae, mientras que los taxones herbáceos como Cyperaceae también aumentaron, sobre todo Polypodiaceae y Aspleniaceae, grupos taxonómicos característicos del bosque mesófilo de montaña (Rzedowski, 1996).
El útimo periodo de ~1183 al presente, la zona C-3a refleja un periodo de transición de la ACM a la PEH con mayor variabilidad ambiental y recambio de taxones con la disminución en el componente herbáceo y el incremento en algunos taxones arbóreos como Pinus, Alnus, Ternstroemia y Xylosma. Sin embargo, muestra un patrón con 2 principales eventos de enfriamiento en la zona C-3b (~1421-1561 y ~1701-1842 d.C.) que se infieren por el incremento en los porcentajes de Pinus y Abies, así como la disminución de Quercus. Sin embargo, las condiciones ambientales son distintas en ambos periodos. En nuestro registro muestra condiciones más húmedas al principio de la PEH asociado con la reducción en los valores de Ca y el aumento en las concentraciones de Ti. En cambio, al final de la PEH muestra condiciones más secas y frías entre ~1701-1842 d.C. inferido con base en los mismos indicadores. Algunos registros históricos de México señalan condiciones extremas de sequías entre los siglos XVI y XIX, teniendo consecuencias severas en la agricultura, con la pérdida de cosechas, crisis sociales y la propagación de epidemias mortales en la población del centro de México (Contreras-Servín; 2005; Florescano et al., 1980; Therrell et al., 2004). Esta variabilidad ambiental puede estar relacionada con los ciclos de actividad solar (Spörer y Maunder) (Haug et al., 2003; Hodell et al., 2001). Los registros de Lago Verde en los Tuxtlas sugieren que los niveles del lago y la vegetación respondieron a este forzamiento solar y que además se puede tratar de un elemento importante que controla la variabilidad climática a escala de décadas y centurias en la región tropical del Atlántico norte (Lozano-García et al., 2007).
De acuerdo con nuestros datos, a partir de secuencias de sedimentos dentro del bosque, se proporcionan evidencias de la respuesta local de los bosques, por lo que nuestro trabajo sugiere que los distintos tipos de vegetación no responden unidireccionalmente ante el cambio global incluso durante los mismos periodos de tiempo. En su lugar, la evidencia señala patrones complejos en las respuestas de la vegetación durante los periodos de cambio ambiental, por lo que pueden ofrecer nuevos escenarios para evaluar los impactos del cambio climático a nivel local.
La evidencia palinológica como el registro de taxones indicadores de actividades culturales (Amaranthaceae y Asteraceae), así como taxones específicos de cultivares como Zea a partir de ~451 d.C. y Phaseolus a los ~1021 d.C., indica la existencia de impacto humano en el área de estudio. Los datos arqueológicos de la época prehispánica de esta región señalan la existencia de numerosos asentamientos indígenas dispersos en las partes bajas del área de estudio. Hacia el noroeste del valle de Mascota, Jalisco, se tienen indicios de poblaciones indígenas posiblemente Huicholes desde aproximadamente 100 a.C. (Cupul-Magaña y Mounjoy, 2012; Kelly, 1980), hacia el sureste en Colima aproximadamente entre 1200-800 a.C. (Mounjoy, 1994). Además, se destaca el hallazgo de Zea mexicana subsp. mexicana (teocintle de Chalco) y Zea mexicana subsp. parviglumis (teocintle del Balsas) cerca del volcán de Colima y la cuenca del Balsas, que posiblemente fue una zona de domesticación del maíz del occidente de México (Benz y Benz, 1990; Pollard, 1997). También hay evidencia de erosión del suelo por el aumento y marcada inestabilidad de la susceptibilidad magnética, además de la disminución en la materia orgánica, así como un régimen de incendios forestales frecuentes. Estos indicadores sugieren cambios importantes en la vegetación, probablemente asociados con disturbios humanos por el aclareo en los bosques para la agricultura (Noble y Dirzo, 1997), por lo que se infiere que la ocupación humana en el sitio de estudio ha estado presente durante algunos intervalos de tiempo, principalmente cuando las condiciones ambientales eran favorables.
La disminución de Pinus ocurrida al final del periodo ~1100-1200 d.C. registrado en nuestra secuencia, así como el aumento de otros taxones característicos de la vegetación herbácea de terrenos abiertos como Poaceae, Asteraceae, Cyperaceae y la presencia de polen de Zea, coincide con valores elevados de Ca y susceptibilidad magnética los cuales son considerados como indicadores de periodos secos y erosión respectivamente. Tales indicadores probablemente sugieren la presencia de actividades antropogénicas llevadas a cabo en el área de estudio. La historia de las diferentes culturas registradas para el occidente de México indica que las poblaciones indígenas eran afectadas por diversos procesos sociales que operaban en el centro de México, por lo que sus sociedades eran dinámicas con patrones complejos de interacción entre las poblaciones de la costa, la sierra del occidente y las poblaciones de las tierras altas del centro de México (Flannery y Marcus, 1994). Tal vez la complejidad de estos patrones se vean reflejados en los diferentes procesos ecológicos que muestra el registro palinológico del sitio de La Cumbre de Guadalupe. Sin embargo, el componente ambiental debió ser decisivo en la movilidad de las poblaciones de las partes altas de la sierra hacia las partes bajas cuando las condiciones del clima se tornaban desfavorables. Por ejemplo, la transición del ACM a la PEH pudo ser un momento difícil para los pobladores de las partes altas de la sierra debido a que en este periodo se presenta un aumento de algunos taxones leñosos
y pteridofitas, lo que indica la recuperación del bosque.
Durante algunos periodos de la última zona de nuestro registro palinológico, está presente el polen de Phaseolus y Zea, por lo que sugiere el establecimiento de los pobladores hacia las partes altas en el área. Seguramente enfrentaban condiciones de sequía en las partes bajas de la sierra, debido a que éstas fueron generalizadas en diferentes zonas de nuestro país (Endfield y O´Hara, 1997; Florescano et al., 1980; Stahle et al., 2011).
Con la llegada de los españoles y la actividad minera, que caracterizó la actividad económica de las localidades aledañas al sitio de estudio en los últimos 2 siglos (Jiménez, 1995), el desarrollo de las actividades agrícolas en las partes altas de la sierra debieron implicar el aumento en el desmonte de las masas forestales; la evidencia de ello se observa en el registro palinológico con la disminución en la riqueza palinológica a partir de ~1927 d.C. y la alta concentración de micropartículas de carbón en ~1912 d.C. Posteriormente, se observa el incremento de los elementos como Ti, Fe, Zn y Pb, además del aumento en la susceptibilidad magnética que pueden estar asociados a mayores tasas de erosión y probablemente relacionadas con estas actividades.
Agradecimientos
Pacheco Contreras por la elaboración del mapa y trabajo de campo. A los estudiantes de IRNA y Agronomía del CU Costa SUR-Universidad de Guadalajara por su apoyo en la extracción de los núcleos de sedimento. A la Dra. Susana Sosa Nájera y Dra. Esperanza Torres Rodríguez por su asesoría en la extracción e identificación de palinomorfos. A los revisores anónimos por sus observaciones y sugerencias. Este trabajo fue realizado con apoyo financiero de los proyectos: Conacyt CB-2008-106435, y COECYTJAL 5-2010-875, y gracias a la beca otorgada por el Conacyt a la primera autora para la realización de sus estudios de posgrado.
Referencias
Agee, J. K. (2000). Fire and pine ecosystems. En D. M. Richardson (Eds.), Ecology and biogeography of Pinus (pp. 193–213). Cambridge: University Press.
Almeida-Leñero, L., Hooghiemstra, H., Cleef, A. M. y van-Geel, B. (2005). Holocene climatic and environmental change from pollen records of lakes Zempoala and Quila, central Mexican highlands. Review of Palaeobotany and Palynology, 136, 63–92.
Bennett, K. D. (2005). Documentation for psimpoll 4.25 and pscomb 1.03: C programs for plotting pollen diagrams and analyzing pollen data. Uppsala, Sweden: Uppsala Universitet.
Benz, K. L. y Benz, B. F. (1990). Resumen del proyecto estudios arqueológicos dentro de la Reserva de la Biosfera Sierra de Manantlán. Consejo de Arqueología. México D.F.: Instituto Nacional de Antropología e Historia.
Bhattacharya, T., Byrne, R., Böhnel, H., Wogau, K., Kienel, U., Ingram, B. L. et al. (2015). Cultural implications of late Holocene climate change in the Cuenca Oriental, Mexico. Proceedings of the National Academy of Sciences, 112, 1693–1698.
Birks, H. J. B., Lotter, A. F., Juggins, S. y Smol, J. P. (2012). Data handling and numerical techniques. En J. P. Smol, H. J. B. Birks, y W. M. Last (Eds.), Tracking environmental change using lake sediments. Dordrecht, The Netherlands: Kluwer Academic Publishers.
Bond, G., Kromer, B., Beer, J., Muscheler, R., Evans, M. N., Showers, W.et al. (2001). Persistent solar influence on North Atlantic climate during the Holocene. Science, 294, 2130–2136.
Caballero, M., Vázquez, G., Lozano-García, S., Rodríguez, A., Sosa-Nájera, S., Ruiz-Fernández, A. C. et al. (2006). Present limnological conditions and recent (ca. 340 yr) palaeolimnology of a tropical lake in the Sierra de Los Tuxtlas, Eastern Mexico. Journal of Paleolimnology, 35, 83–97.
Calcote, R. (1995). Pollen source area and pollen productivity: evidence from forest hollows. Journal of Ecology, 83, 591–602.
Calcote, R. (1998). Identifying forest stand types using pollen from forest hollows. The Holocene, 8, 423–432.
Caldararo, N. (2002). Human ecological intervention and the role of forest fires in human ecology. Science of the Total Environment, 292, 141–165.
Cayuela, L., Golicher, D. J., Benayas, J., González, E. R. M. y Ramírez, M. N. (2006). Fragmentation, disturbance and tree diversity conservation in tropical montane forests. Journal of Applied Ecology, 43, 1172–1181.
Cerano-Paredes, J., Villanueva-Díaz, J., Cervantes-Martínez, R., Fulé, P., Yocom, L., Esquivel-Arriaga, G. et al.(2015). Historia de incendios en un bosque de pino de la sierra de Manantlán, Jalisco, México. Bosque, 36, 41–52.
Clark, R. L. (1982). Point count estimation of charcoal in pollen preparations and thin sections of sediments. Pollen et Spores, 24, 523–535.
Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad). (2010). El bosque mesófilo de montaña en México: amenazas y oportunidades para su conservación y manejo sostenible. México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Contreras- Servín, C. (2005). Las sequías en México durante el siglo XIX. Investigaciones Geográficas, 56, 118–133.
Cuevas-Guzmán, R., Cisneros-Lepe, E. A., Jardel-Peláez E. J., Sánchez-Rodríguez, E. V., Guzmán-Hernández, L., Núñez-López, N. M. et al. (2011). Análisis estructural y de diversidad en los bosques de Abies de Jalisco, México. Revista Mexicana de Biodiversidad, 82, 1219–1233.
Cupul-Magaña, F. G. y Mountjoy, J. B. (2012). Tachemys ornata (Gray, 1831) (Testudines: Emydidae) en un yacimiento arqueológico del occidente de México. Acta Zoológica Mexicana, 28, 222–226.
Dean, W. E. (1974). Determination of carbonate and organic matter in calcareous sediments and sedimentary rocks by loss on ignition: comparison with other methods. Journal of Sedimentary Petrology, 44, 242–248.
del Castillo-Batista, A. P. (2017). Historia de la vegetación y cambio climático en bosques templados de alta montaña del centro-occidente de México durante el Holoceno tardío (Tesis doctoral en Ciencias en Biosistemática, Ecología y Manejo de Recursos Naturales y Agrícolas). Universidad de Guadalajara. Autlán de Navarro, Jalisco, México.
Díaz, H. F., Trigo, R., Hughes, M. K., Mann, M. E., Xoplaki, E. y Barriopedro, D. (2011). Spatial and temporal characteristics of climate in medieval times revisited. Bulletin of the American Meteorological Society, 92, 1487–1500.
Encina-Domínguez, J. A., Encina-Domínguez, F. J., Mata-Rocha, E. y Valdés, J. (2008). Aspectos estructurales, composición florística y caracterización ecológica del bosque de oyamel de la Sierra de Zapalinamé, Coahuila, México. Boletín de la Sociedad Botánica de México, 83, 13–24.
Endfield, G. H. y O’Hara, S. L. (1997). Conflicts over water in “The Little Drought Age” in central Mexico. Environment and History, 3, 255–272.
Faegri, K. y Iversen, J. (1989). Textbook of pollen analysis. Chichester, Great Britain: John Wiley & Sons .
Figueroa-Rangel, B. L., Willis, K. J. y Olvera-Vargas, M. (2008). 4200 years of pine-dominated upland forest dynamics in west-central Mexico: human or natural legacy. Ecology, 89, 1893–1907.
Flannery, K. V. y Marcus, J. (1994). Early formative pottery of the valley of Oaxaca, Mexico. Memoirs of the Museum of Anthropology, 27. University of Michigan Museum of Anthropology.
Florescano, E., Sancho, J. y Pérez, D. (1980). Las sequías en México: historia, características y efectos. Comercio Exterior, 30, 747–757.
Galindo-Jaimes, L., González-Espinosa, M., Quintana-Ascencio, P. y García-Barrios, L. (2002). Tree composition and structure in disturbed stands with varying dominance by Pinus spp. in the highlands of Chiapas, Mexico. Plant Ecology, 162, 259–272.
Gernandt, D. S. y Pérez-de la Rosa, J. A. (2014). Biodiversidad de Pinophyta (coníferas) en México. Revista Mexicana de Biodiversidad, 85 (Suplem.), S126–S133.
Gill, R. B., Mayewski, P. A., Nyberg, J., Haug, G. H. y Peterson, L. C. (2007). Drought and the Maya collapse. Ancient Mesoamerica, 18, 283–302.
González-Espinosa, M., Quintana-Ascencio, P. F., Ramírez-Marcial, N. y Gaytán-Guzmán, P. (1991). Secondary succession in disturbed Pinus-Quercus forests in the highlands of Chiapas, Mexico. Journal of Vegetation Science, 2, 351–360.
Grimm, E. C. (1987). CONISS a Fortan program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Geosciences, 13, 13–35.
Guerrero-Hernández, R., González-Gallegos, J. G. y Castro-Castro, A. (2014). Análisis florístico de un bosque de Abies y el bosque mesófilo de montaña adyacente en Juanacatlán, Mascota, Jalisco, México. Botanical Sciences, 92, 541–562.
Haug, G. H., Günther, D., Peterson, L. C., Sigman, D. M., Hughen, K. A. y Aeschlimann, B. (2003). Climate and the collapse of Maya civilization. Science, 299, 1731–1735.
Hodell, D. A., Brenner, M., Curtis, J. H. y Guilderson, T. (2001). Solar forcing of drought frequency in the Maya lowlands. Science, 292, 1367–1370.
Hodell, D. A., Brenner, M., Curtis, J. H., Medina-Gonzalez, R., Can, E. I. C., Albornaz-Pat, A. et al. (2005). Climate change on the Yucatán Peninsula during the Little Ice Age. Quaternary Research, 63, 109–121.
Inegi (Instituto Nacional de Estadística y Geografía). (1976). Carta edafológica de Llano Grande F13D81, escala 1:50,000: México. D.F.: Instituto Nacional de Estadística, Geografía e Informática.
Inegi (Instituto Nacional de Estadística y Geografía). (2015). Síntesis de información geográfica del Estado de Jalisco, México. Recuperado el 05 de septiembre, 2015 de http://cuentame.inegi.org.mx/monografias/informacion/jal/territorio/relieve.aspx?tema=me.
Jardel, P. E. (1994). Diversidad ecológica y transformaciones del paisaje en el occidente de México. En R. Ávila, (Ed.) Transformaciones mayores en el occidente de México (pp. 13–39). Guadalajara: Universidad de Guadalajara.
Jiménez, P. A. (1995). Santa Rosalía y Ayutla: vida cotidiana y economía. México: Colegio de Jalisco.
Keeley, J. E. (2012). Ecology and evolution of pine life histories. Annals of Forest Science, 69, 445–453.
Kelly, I. (1945). The Archeology of the Autlán-Tuxcacuesco, area of Jalisco. I: The Autlán Zone. Ibero-Americana, 26, 1–93
Kelly, I. (1980). Ceramic sequence in Colima: Capacha, an early phase. Anthropological Papers of the University of Arizona. Tucson: University of Arizona Press.
Lara-González, R., Sánchez-Velásquez, L. R. y Corral-Aguirre, J. (2009). Regeneración de Abies religiosa en claros del dosel versus sotobosque, Parque Nacional Cofre de Perote, México. Agrociencia, 43, 739–747.
Lozano-García, S., Caballero, M., Ortega, B., Rodríguez, A. y Sosa, S. (2007). Tracing the effects of the Little Ice Age in the tropical lowlands of eastern Mesoamerica. Proceedings of the National Academy of Sciences, 104, 16200–16203.
Lozano-García, S., Caballero, M., Ortega, B., Sosa, S., Rodríguez, A. y Schaaf, P. (2010). Late Holocene palaeoecology of Lago Verde: evidence of human impact and climate change in the northern limit of the neotropics during the late formative and classic periods. Vegetation History and Archaeobotany, 19, 177–190.
Ludlow-Wiechers, B., Almeida-Lenero, L. y Islebe, G. (2005). Paleoecological and climatic changes of the Upper Lerma Basin, Central Mexico during the Holocene. Quaternary Research, 64, 318–332.
Magaña, V. O., Vázquez, J. L., Pérez, J. L. y Pérez, J. B. (2003). Impact of El Niño on precipitation in Mexico. Geofísica Internacional, 42, 313–330.
Manzanilla, H. (1974). Investigaciones epidométricas y silvícolas en bosques mexicanos de Abies religiosa. México D.F.: Dirección General de Información y Relaciones Públicas de la Secretaría de Agricultura y Ganadería.
Martínez-Salas, E. y Ramos, C. H. (2014). Biodiversidad de Pteridophyta en México. Revista Mexicana de Biodiversidad, 85, 110–113.
Metcalfe, S. E., Jones, M. D., Davies, S. J., Noren, A. y MacKenzie, A. (2010). Climate variability over the last two millennia in the North American Monsoon region, recorded in laminated lake sediments from Laguna de Juanacatlán, Mexico. The Holocene, 20, 1195–1206.
Metcalfe, S. E., O’Hara, S. L., Caballero, M. y Davies, S. J. (2000). Records of Late Pleistocene-Holocene climatic change in Mexico-a review. Quaternary Science Reviews, 19, 699–721.
Mountjoy, J. B. (1994). Capacha: una cultura enigmática del occidente de México. Arqueología Mexicana, 2, 39–42.
Nelson, D. M., Hu, F. S., Grimm, E. C., Curry, B. B. y Slate, J. E. (2006). The influence of aridity and fire on Holocene prairie communities in the eastern Prairie Peninsula. Ecology, 87, 2523–2536.
Noble, I. R. y Dirzo, R. (1997). Forests as human-dominated ecosystems. Science, 277, 522–525.
Ortega-Rosas, C. I., Peñalba, M. C. y Guiot, J. (2008). Holocene altitudinal shifts in vegetation belts and environmental changes in the Sierra Madre Occidental, Northwestern Mexico, based on modern and fossil pollen data. Review of Palaeobotany and Palynology, 151, 1–20.
PAGES-2k-Consortium. (2013). Continental-scale temperature variability during the past two millennia. Nature Geoscience, 6, 339–346.
Perry, J. P., Graham, A. y Richardson, D. M. (2000). The history of pines in Mexico and Central America. En D. M. Richardson, (Ed.), Ecology and biogeography of Pinus (pp.137–147), Cambridge University Press.
Pollard, H. P. (1997). Recent research in west Mexican archaeology. Journal of Archaeological Research, 5, 345–384.
Quintana-Ascencio, P. F. y González-Espinosa, M. (1993). Afinidad fitogeográfica y papel sucesional de la flora leñosa de los bosques de pino-encino de los Altos de Chiapas, México. Acta Botanica Mexicana, 21, 43–57.
R Development Core Team (2013). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Recuperado el 07 septiembre 2015 de: http://www.R-project.org
Reimer, P. J., Baillie, M. G., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G. et al. (2009). IntCal09 and Marine09 radiocarbon age calibration curves, 0-50,000 years cal BP. Radiocarbon, 51, 1111–1150.
Richardson, D. M. y Bond, W. J. (1991). Determinants of plant distribution: evidence from pine invasions. American Naturalist, 137, 639–668.
Richardson, D. M. y Rundel, P. W. (1998). Ecology and biogeography of Pinus: an introduction. En D. M. Richardson, (Ed.), Ecology and biogeography of Pinus (pp. 3–46). Cambridge: Cambridge University Press.
Rodríguez-Ramírez, A., Caballero, M., Roy, P., Ortega, B., Vázquez-Castro, G. y Lozano-García, S. (2015). Climatic variability and human impact during the last 2000 years in western Mesoamerica: evidence of late Classic (AD 600-900) and Little Ice Age drought events. Climate of the Past, 11, 1239.
Rodríguez-Trejo, D. A. y Fulé, P. Z. (2003). Fire ecology of Mexican pines and a fire management proposal. International Journal of Wildland Fire, 12, 23–37.
Rzedowski, J. (1996). Análisis preliminar de la flora vascular de los bosques mesófilos de montaña de México. Acta Botanica Mexicana, 35, 25–44.
Rzedowski, J. (2006). Vegetación de México. México, D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Rzedowski, J. y McVaugh, R. (1966). La vegetación de Nueva Galicia. Contributions from the University of Michigan Herbarium, 9, 1–123.
Sosa-Nájera, S., Lozano-García, S., Roy, P. D. y Caballero, M. (2010). Registro de sequías históricas en el occidente de México con base en el análisis elemental de sedimentos lacustres: el caso del lago de Santa María del Oro. Boletín de la Sociedad Geológica Mexicana, 62, 437–451.
Stahle, D. W., Villanueva-Díaz, J., Burnette, D. J., Cerano-Paredes, J., Heim, R. R., Fye, F. K. et al. (2011). Major Mesoamerican droughts of the past millennium. Geophysical Research Letters (38). doi:10.1029/2010GL046472, 2011
Stockmarr, J. (1971). Tablets with spores used in absolute pollen analysis. Pollen et Spores, 13, 615–621.
Stuiver, M. y Reimer, J. P. (1993). Extended 14C data base and revised CALIB 3.014 C age calibration program. Radiocarbon, 35, 215–230.
Ter Braak, C. J. (1986). Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology, 67, 1167–1179.
Ter Braak, C. T. y Smilauer, P. (2002). CANOCO reference manual and canodraw for windows, users guide. Software for Canonical Community Ordination (version 4.5), Ithaca, Microcomputer Power.
Tetreault, D. V., López, L. y Federico, C. (2011). Diversidad biocultural en el estado de Jalisco: pueblos indígenas y regiones de alto valor biológico. Espiral, 18, 165–199.
Therrell, M. D., Stahle, D. W. y Soto, R. A. (2004). Aztec drought and the “Curse of one rabbit”. Bulletin of the American Meteorological Society, 85, 1263–1272.
Thompson, R. y Oldfield, F. (1986). Environmental magnetism. Londres: Allen y Unwin.
Vargas-Rodríguez, Y. L. y Platt, W. J. (2012). Remnant sugar maple (Acer saccharum subsp. skutchii) populations at their range edge: characteristics, environmental constraints and conservation implications in tropical America. Biological Conservation, 150, 111–120.
Weber, P., Bugmann, H., Fonti, P. y Rigling, A. (2008). Using a retrospective dynamic competition index to reconstruct forest succession. Forest Ecology and Management, 254, 96–106.
Weigand, P. C. (1996). La evolución y ocaso de un núcleo de civilización: la tradición Teuchitlán y la arqueología de Jalisco. En E. Williams y P. C. Weigand (Eds.), Las cuencas del occidente de México: época prehispánica, (pp. 185–246). México D.F.: El Colegio de Michoacán.
Whitlock, C. y Larsen, C. (2001). Charcoal as a fire proxy. En J. P. Smol, H. J. B. Birks y W. M. Last (Eds.), Tracking environmental change using lake sediments (pp. 75–97) Dordrecht, The Netherlands: Kluwer Academic Publishers.


