Mamíferos medianos y grandes en sitios de tala de impacto reducido y de conservación en la sierra Juárez, Oaxaca
Enrique Hernández-Rodríguez a, Luis Escalera-Vázquez b, Jaime M. Calderón-Patrón c y Eduardo Mendoza a, *
a Instituto de Investigaciones sobre los Recursos Naturales, Universidad Michoacana de San Nicolás de Hidalgo, Av. San Juanito Itzícuaro s/n, Col. Nueva Esperanza, 58330, Morelia, Michoacán, México
b Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo, calle Gral. Francisco J. Múgica s/n A-1, Felícitas de Río, 58030 Morelia, Michoacán, México
c Centro de Investigaciones Biológicas, Instituto de Ciencias Básicas e Ingeniería, Universidad Autónoma del Estado de Hidalgo, carretera Pachuca-Tulancingo Km 4.5, Colonia Carboneras, 42184 Mineral de la Reforma, Hidalgo, México
*Autor para correspondencia: emramirez@umich.mx (E. Mendoza)
Resumen
El sistema de tala de impacto reducido busca afectar mínimamente la biodiversidad. Sin embargo, su efecto sobre las poblaciones de mamíferos medianos y grandes ha sido escasamente explorado. En la sierra Juárez de Oaxaca, que presenta una alta diversidad de mamíferos medianos y grandes, se ha implementado ese sistema de manejo, lo que permite evaluar los impactos de esta técnica forestal sobre este tipo de fauna. Se estudió la diversidad y estructura del ensamble de mamíferos medianos y grandes presentes en áreas con tala de impacto reducido (n = 3) y áreas de conservación (n = 3). Los resultados indican que este sistema de manejo forestal permite mantener la estructura del ensamble de mamíferos de talla media y grande. Entre los factores que parecen favorecer esta situación son el tamaño pequeño de las áreas de manejo y la alta cobertura forestal circundante. La implementación de esta técnica en otros ecosistemas forestales puede ayudar a reducir el fuerte impacto que hay sobre las poblaciones de mamíferos silvestres en áreas bajo explotación.
Palabras clave: Diversidad α; Diversidad β; Estrategias de conservación; Manejo forestal
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Medium and large mammals in reduced-impact logging and conservation sites in Sierra Juárez, Oaxaca
Abstract
The reduced-impact logging system aims to minimally affect biodiversity. However, its effects on medium and large mammals’ populations have been scarcely explored. In the Sierra Juárez of Oaxaca, which has a high diversity of medium and large mammals, this management system has been implemented, which allows evaluating the impacts of this forest management on this fauna group. We studied the diversity and structure of medium and large mammals ensembles ocurring in areas with reduce-impact logging (n = 3) and conservation areas (n = 3). The results revealed that reduced-impact logging system is compatible with maintaining the structure of the ensembles of medium and large mammals. Among the factors that seem to favor this situation are the small size of the management areas and the abundant surrounding forest cover. The implementation of this system in other forest ecosystems can help to reduce the strong impact on wild mammal’s populations in areas under exploitation.
Keywords: α diversity; β diversity; Conservation strategies; Forest management
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
La principal causa de pérdida de biodiversidad en los países tropicales es la modificación del hábitat por la conversión de la cobertura forestal a distintos usos de suelo (FAO, 2015; Grillo y Gianfranco, 2011; Günter et al., 2011). Una de las causas de esta modificación es la explotación forestal que puede producir fuertes cambios en la estructura de los ecosistemas (Chaudhary et al., 2016; Putz et al., 2000). De acuerdo con Chaudhary et al. (2016), a nivel global se identifican 6 regímenes comunes de manejo forestal que se distinguen por su intensidad y la planificación con la que se realiza la cosecha de madera. Estos regímenes van desde los que tienen impactos drásticos sobre la estructura del bosque (e.g., tala rasa) hasta aquellos en los que la estructura del bosque permanece muy similar a la de sitios conservados (e.g., tala selectiva). Estos últimos se han propuesto como un medio para contribuir a una gestión más sostenible de los recursos naturales en donde se mitiguen los efectos negativos sobre la biodiversidad (Günter et al., 2011).
En particular, la tala de impacto reducido o RIL (por sus siglas en inglés) involucra la realización de inventarios forestales, previos a la explotación, para definir los procedimientos de extracción que llevan a cabo trabajadores capacitados, así como la elaboración de planes de cosecha enfocados a minimizar el impacto sobre la biodiversidad y los procesos ecológicos del bosque (ITTO, 2018; Miller et al., 2011; Putz et al., 2008).
Uno de los componentes de la biodiversidad que puede ser afectado por el manejo forestal es la fauna de mamíferos (Chaudhary et al., 2016; Putz et al., 2012). Se han realizado numerosos estudios para evaluar el efecto del manejo forestal sobre este grupo de vertebrados, sin embargo, las revisiones hechas por Chaudhary et al. (2016), Gibson et al. (2011) y Putz et al.(2012) muestran que la mayoría de estos estudios se han centrado en mamíferos pequeños, siendo menos explorados los efectos sobre los mamíferos de mayor talla.
Los mamíferos medianos y grandes incluyen especies terrestres y arborícolas no voladoras, normalmente identificables sin la necesidad de ser capturados, con un peso generalmente superior a 1 kg para los de talla mediana y superior a los 20 kg para los de talla grande (Benchimol, 2016; Morrison et al., 2007; Rumiz et al., 1998). Esta fauna es de gran importancia ya que su actividad tiene efectos directos e indirectos sobre la vegetación y sobre otras especies animales a través de la herbivoría, dispersión de semillas y el consumo de presas (Dirzo y Miranda, 1990; Miller et al., 2001; Ripple et al., 2014).
La fauna de mayor tamaño, generalmente, es la más afectada por las actividades humanas, por lo tanto presenta un mayor riesgo de extinción (Dirzo et al., 2014). Este tipo de fauna silvestre incluye especies que son muy atractivas para ser cazadas o capturadas, ya sea porque proporcionan alimento, por sus pieles o porque son usadas como mascotas (Ojasti, 2000). Asimismo, algunas de estas especies son perseguidas por considerarlas una amenaza para los humanos o para sus animales domésticos (Inskip y Zimmermann, 2009). Por otra parte, la fauna de mayor talla comúnmente presenta tasas reproductivas bajas y ámbitos hogareños amplios (Hernández-Huerta, 1992; Powell, 2012), lo que hace que la deforestación y fragmentación del hábitat las afecte con mayor intensidad (Galetti y Dirzo, 2013). Las áreas naturales protegidas ayudan a mitigar los impactos negativos sobre la fauna, pero son insuficientes ante las distintas amenazas existentes. Por ello es altamente deseable que los sistemas de manejo forestal favorezcan la permanencia de la fauna silvestre de talla mediana y grande (Chaudhary et al., 2016).
Este estudio analiza el impacto del manejo forestal por tala de impacto reducido sobre la diversidad y composición del ensamble de mamíferos medianos y grandes presentes en el bosque de pino encino en la sierra Juárez, Oaxaca. Esta región destaca por albergar un alto porcentaje de endemismos y una alta riqueza de mamíferos que incluye alrededor de 30 especies medianas y grandes (Briones-Salas et al., 2015). Por otra parte, estos bosques son importantes para la producción forestal ya que forman parte de las 57,000 ha bajo manejo de la región (Sistema Estatal de Información Forestal, 2015) que proporcionan una de las principales fuentes de ingreso económico para las comunidades locales (Álvarez y Rubio, 2013). En particular, se contrastó la diversidad α y la β del ensamble de mamíferos presentes en 3 sitios bajo manejo y 3 sitios en los que no se realizan actividades de explotación y que son denominadas localmente como sitios de conservación. Nuestra predicción es que en sitios bajo manejo forestal por tala de impacto reducido, la diversidad del ensamble de mamíferos medianos y grandes será menor que la de los ensambles registrados en los sitios de conservación.
Materiales y métodos
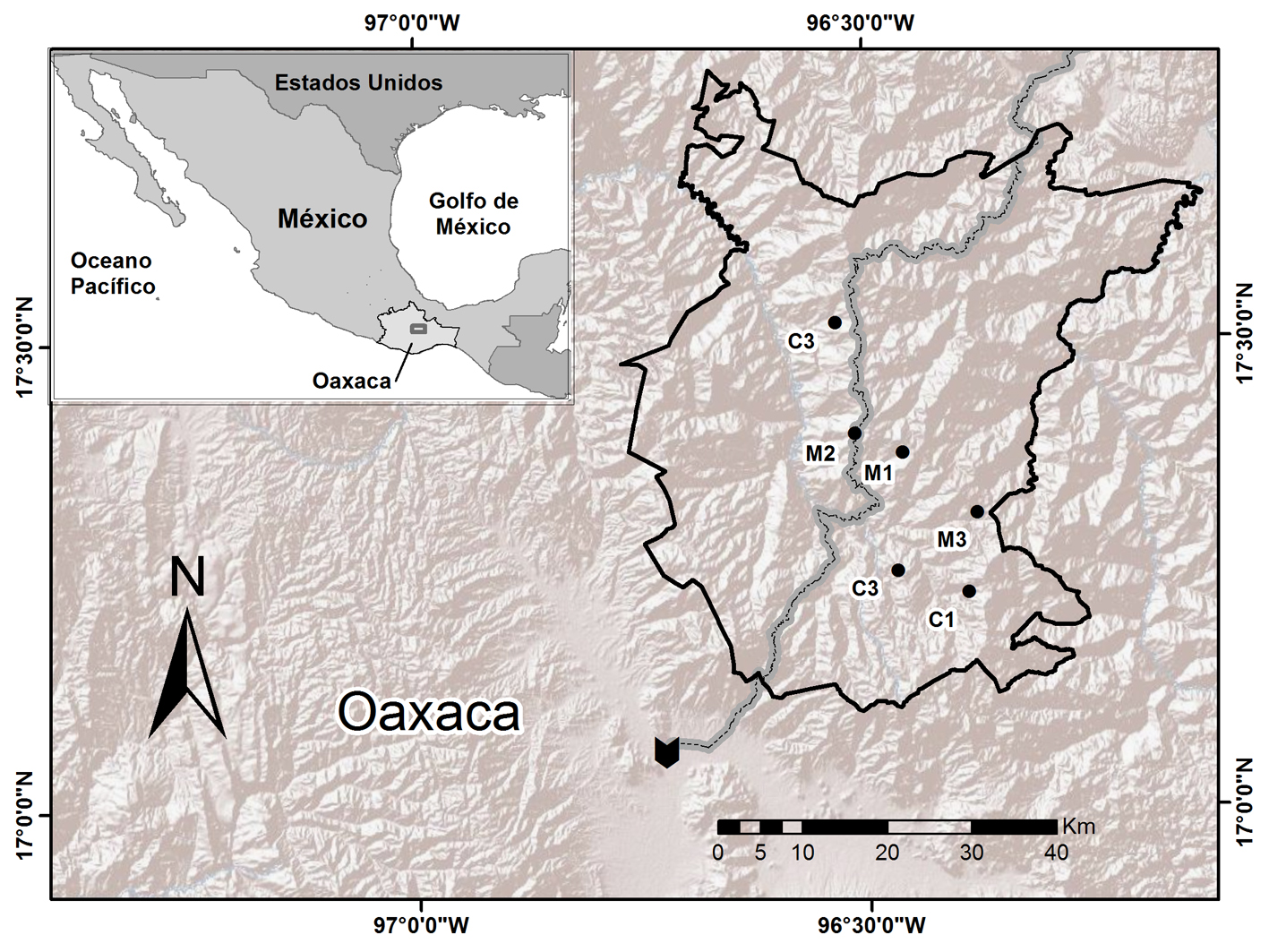
La sierra Juárez se localiza en la región norte del estado de Oaxaca entre las coordenadas 17° N, 96° O (Ortiz-Pérez et al., 2004). El clima es templado subhúmedo (C(w)) con una temperatura media anual de 16 °C. En la zona se distinguen varios tipos de vegetación, siendo los bosques de pino-encino donde se realizan las actividades de manejo. Entre las especies arbóreas representativas están: Pinus ayacahuite, P. leiophylla, P. patula, P. pseudostrobus, Quercus crassifolia, Q. elliptica y Q. laeta (Torres-Colín, 2004). Las localidades que corresponden a las áreas de conservación designadas son: Santa María Yavesía (C1; 7,134 ha), Santa Catarina Lachatao (C2; 12,936 ha) y San Juan Luvina (C3; 230), destinadas a este propósito. Las que constituyen áreas para el aprovechamiento forestal son: Ixtlán de Juárez (M1), San Juan Evangelista Analco (M2) y Capulálpam de Méndez (M3) con 107, 13.5 y 10 hectáreas, respectivamente, para manejo forestal de impacto reducido (fig. 1).
En cada una de las localidades se colocaron 4 cámaras trampa por un lapso de 58 días (del 22 de febrero al 21 abril de 2017). Las cámaras fueron programadas para tomar 3 fotografías por detección y fueron ubicadas en sitios donde la visibilidad no estaba obstruida, así como en sitios de acceso de los animales. El espacio mínimo entre cámaras fue de 500 m. Cada cámara fue georreferenciada con un GPS Garmin GPSMAP64s. Para obtener valores de la frecuencia de registros de cada especie se cuantificaron las imágenes respectivas agrupadas en periodos de 24 horas. En el caso de fotografías con más de un individuo, se registró el total de individuos capturados (Rowcliffe et al., 2008). Para la identificación de las especies se utilizaron los trabajos de Ceballos y Oliva (2005) y Reid (2009). La nomenclatura utilizada corresponde a Ramírez-Pulido et al. (2014).
La completitud del inventario en cada condición se evalúo mediante el cálculo de la “cobertura de la muestra”, que mide la proporción que representan los individuos de cada especie en la muestra, con respecto al número total de individuos. Este método estandariza comunidades desigualmente muestreadas, de manera que aprovecha la mayor parte de la información obtenida en campo, y se propone como un mejor método evaluar la riqueza de especies que la rarefacción tradicional (Chao y Jost, 2012). Para el análisis de la rarefacción por cobertura de la muestra se utilizó el procedimiento de Chao y Jost (2012). Los análisis se realizaron con 100 aleatorizaciones y extrapolando al doble del número de individuos de la comunidad con la menor cobertura de muestra (Chao y Jost, 2012). De este modo se realizaron curvas de rarefacción/extrapolación (R/E) con intervalos de confianza de 95% para comparar la riqueza observada y estimada entre cada condición (Chao et al., 2014).
Para evaluar el recambio de especies (diversidad β) entre y dentro de bosques de conservación o bajo manejo (condiciones), se utilizó el método propuesto por Carvalho et al. (2012) el cual considera al reemplazo de especies (βrepl) y a las diferencias en riqueza (βrich), como procesos que explican las diferencias en el recambio de especies total (βtotal).
Para comparar la estructura de los ensambles de mamíferos en cada localidad se realizaron curvas de rango-abundancia (Magurran, 2004) utilizando las frecuencias de registro de las especies. Posteriormente, para evaluar el grado de similitud de los ensambles registrados en cada condición se realizó un dendrograma de agrupación jerárquica mediante el método de aglomeración simple. Finalmente, para evaluar si existía una relación entre la composición de los ensambles, la distancia geográfica de las localidades de estudio y la condición de manejado/conservado, se realizó un análisis de Mantel parcial (Legendre y Legendre, 2012).
Los análisis se realizaron en el programa R versión 3.5.0 (R Core Team, 2018) usando diferentes paquetes. Las curvas de rarefacción/extrapolación y la estimación de la diversidad α se realizaron con iNEXT 2.0.14 (Hsieh et al., 2016), mientras que el análisis de diversidad β se desarrolló con el paquete BAT (Cardoso et al., 2017). Para el análisis de conglomerados se utilizó Constant (R Core Team, 2018) y para el análisis de Mantel parcial el paquete Vegan 2.4-4 (Oksanen et al., 2018).
Tabla 1
Listado de especies registradas mediante fototrampeo en sitios conservación (C) y manejo (M) forestal en la sierra de Juárez.
|
Ordenes, familias y especies |
Conservación |
Manejo |
||||
|
C1 |
C2 |
C3 |
M1 |
M2 |
M3 |
|
|
Artiodactyla |
||||||
|
Cervidae |
||||||
|
Odocoileus virginianus |
* |
* |
* |
* |
* |
* |
|
Tayassuidae |
||||||
|
Dicotyles angulatus |
* |
|||||
|
Carnivora |
||||||
|
Canidae |
||||||
|
Urocyon cinereoargenteus |
* |
* |
||||
|
Felidae |
||||||
|
*Leopardu spardalis |
* |
* |
* |
|||
|
Lynx rufus |
* |
|||||
|
Puma concolor |
* |
* |
||||
|
Mephitidae |
||||||
|
Conepatus leuconotus |
* |
* |
* |
|||
|
Spilogale angustifrons |
* |
* |
||||
|
Procyonidae |
||||||
|
*Bassariscus astutus |
* |
|||||
|
*Nasua narica |
* |
* |
||||
|
Lagomorpha |
||||||
|
Leporidae |
||||||
|
Sylvilagus sp. |
* |
* Especies incluidas en la NOM-059- Semarnat 2010.
Resultados
Durante el esfuerzo de muestreo de 1,320 días cámara trampa (672 para sitios conservados y 648 para manejados), se registraron para todos los sitios un total de 11 especies de mamíferos medianos y grandes (7 en sitios de manejo y 9 en sitios conservados) correspondientes a 7 familias y 3 órdenes. De estas especies, 3 se encuentran en la NOM-059-Semarnat-2010. El orden con mayor número de especies fue Carnívora (tabla 1). A excepción de la comunidad de Santa María Yavesía (C1), donde se registraron 6 especies, las localidades restantes tuvieron de 3 a 4 especies de mamíferos. Al comparar con la estimación de la riqueza, se registró un promedio de 88% y 97% de la riqueza estimada de mamíferos en sitios conservados y manejados respectivamente (tabla 2). Aunque una de las comunidades registró una mayor riqueza (C1), los resultados de la curva R/E indican que no hay diferencias significativas entre las 2 condiciones de estudio. Los patrones de riqueza se mantienen al extrapolar las estimaciones a un mayor número de días cámara trampa (fig. 2).
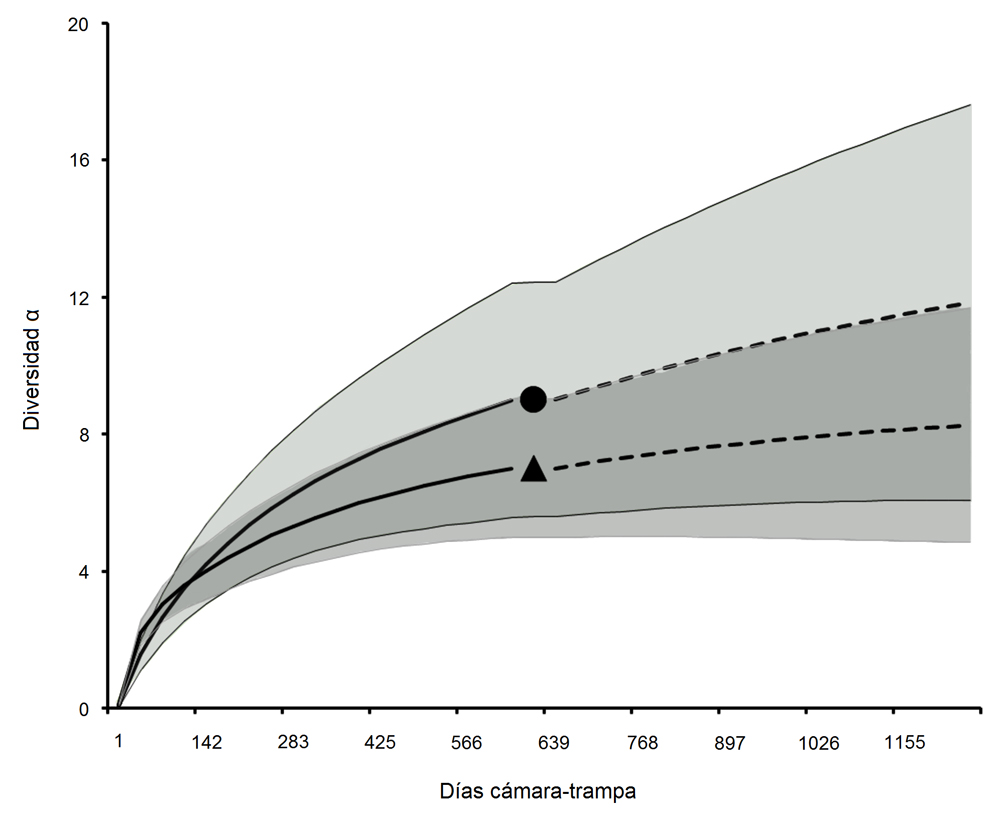
Tabla 2
Riqueza de mamíferos medianos y grandes registrada, estimada y su porcentaje de registro mediante el método por cobertura de la muestra para sitios de conservación y manejo.
|
Condición |
Riqueza registrada |
Riqueza estimada |
% de registro de especies |
|
Conservación |
9 |
12 |
88 |
|
Manejo |
7 |
8 |
97 |
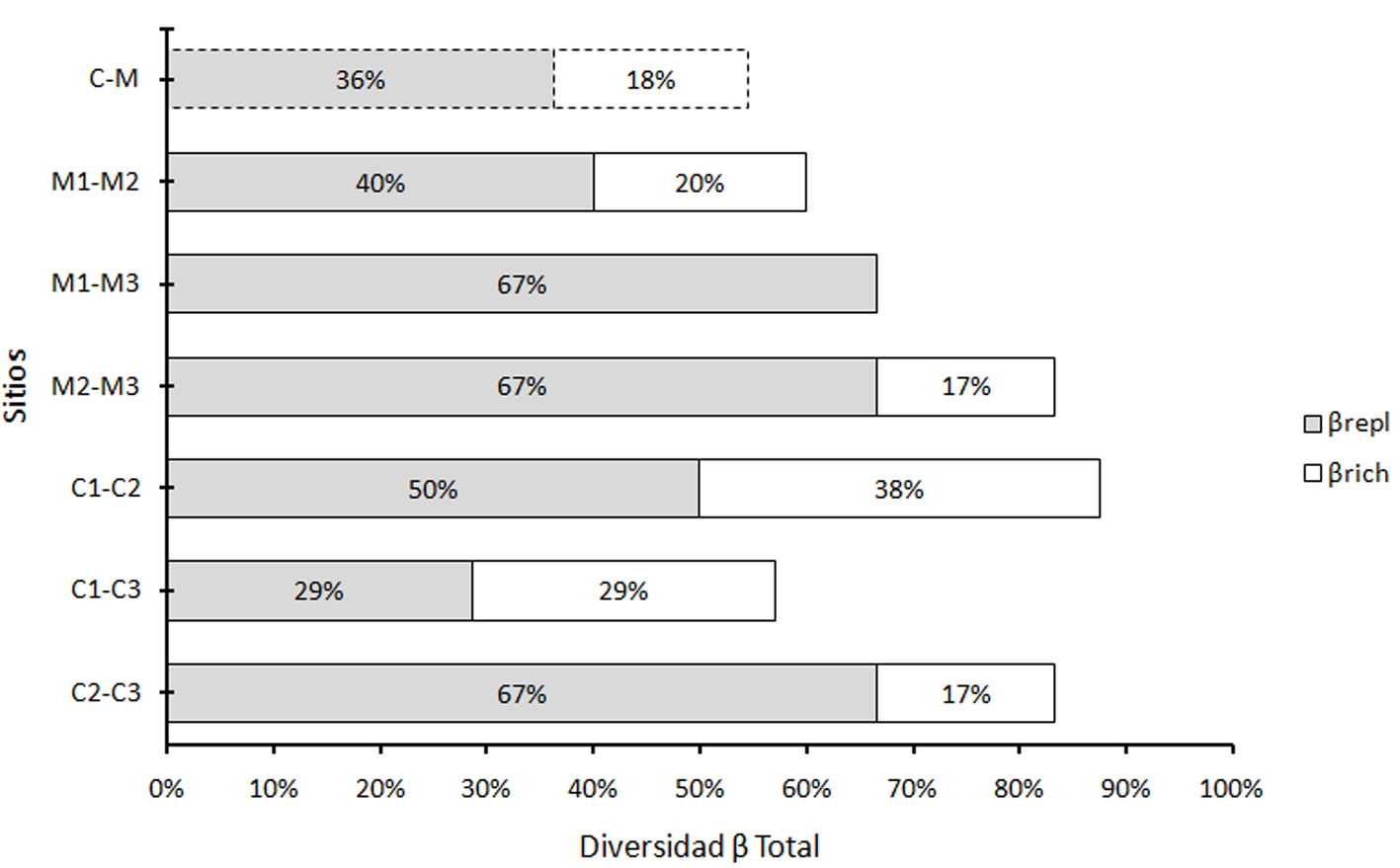
El análisis de diversidad β dentro de sitios de conservación, muestra en promedio un 76% de βtotal. De esta diversidad, en promedio 48% de esta variación es explicada por el reemplazo de especies (βrepl) y 28% por diferencias en riqueza (βrich). Dentro de sitios manejados la diversidad β es en promedio del 70%, donde 58% es explicado por reemplazo (βrepl) y 12% por diferencias en riqueza (βrich). En sitios conservados la βtotal, sólo en un caso (C1-C2), es mayor que en sitios manejados. En el caso de la diversidad β entre condiciones, la βtotal es de 54%, donde 36% es explicado por reemplazo (βrepl) y 18% por diferencias en riqueza (βrich). De esta manera la βtotal es mayor dentro de condiciones que entre condiciones (fig. 3). Además, el reemplazo siempre fue mayor que las diferencias en riqueza, dentro y entre condiciones.
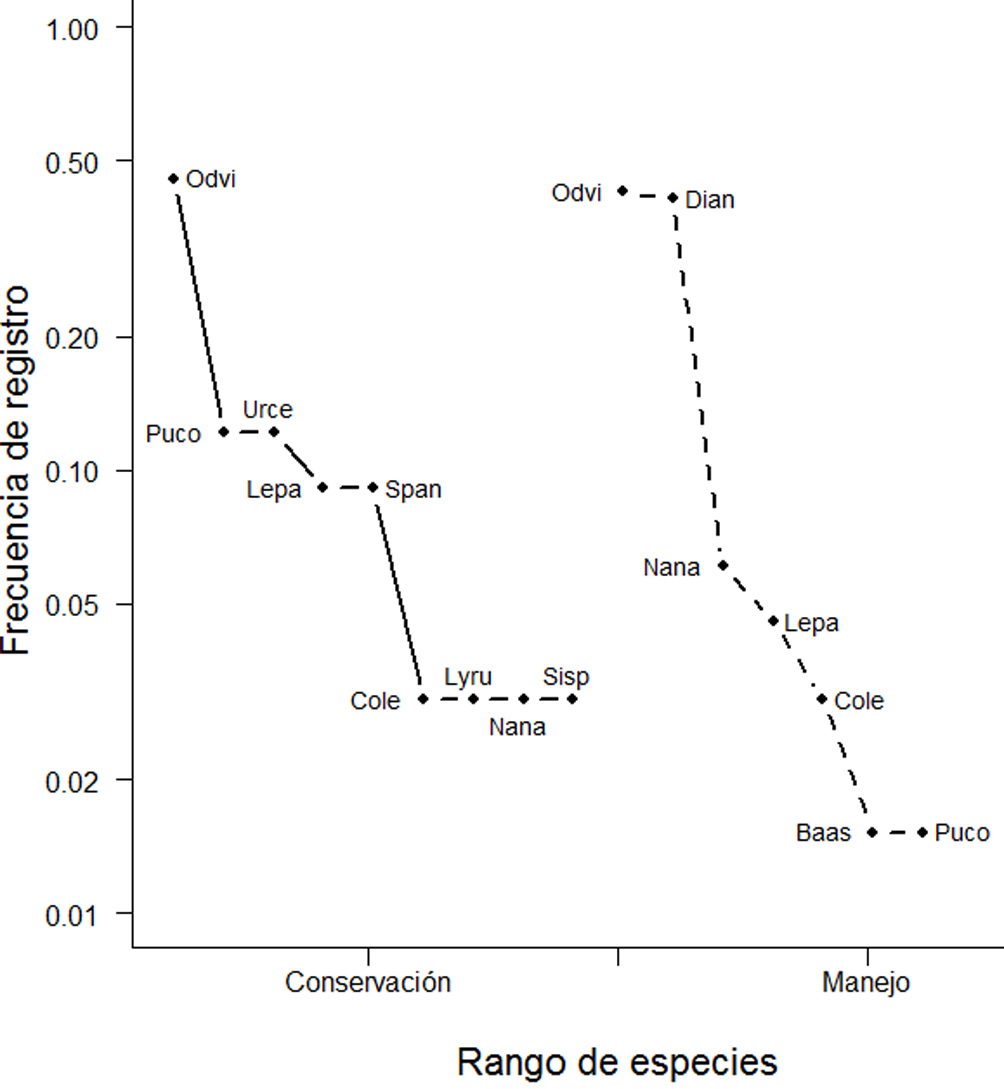
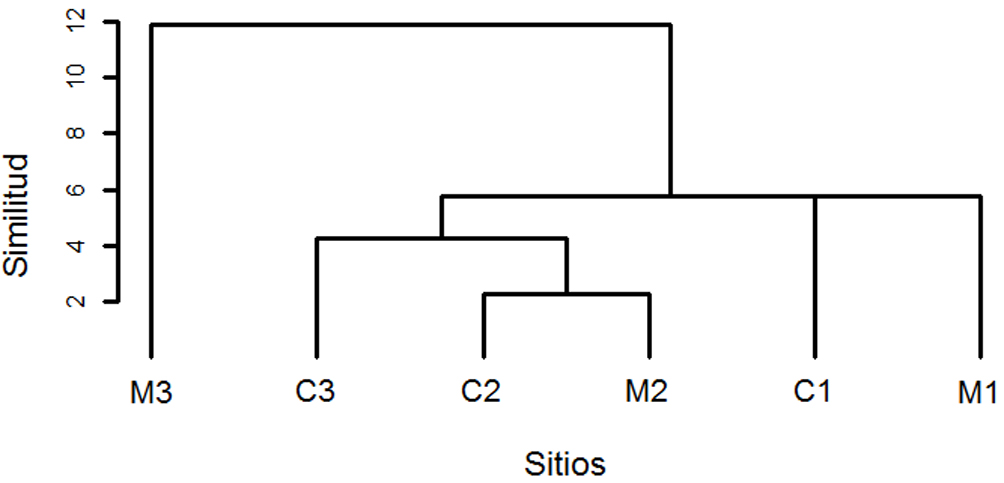
Las gráficas rango-abundancia muestran un patrón similar en la frecuencia de registro de las especies de mamíferos en ambas condiciones. La especie O. virginianus, tiene una mayor frecuencia que la mayoría de las especies, aunque similar en ambas condiciones (fig. 4). El análisis de conglomerados revela que no hay una segregación de los sitios conservados y manejados en función de la composición del ensamble de especies presentes (fig. 5). El análisis parcial de Mantel mostró que no existe correlación entre la composición de especies de mamíferos medianos y grandes, la distancia geográfica de las localidades de estudio ni su condición como sitio de conservación o manejo (r = 0.0075, p = 0.44).
Discusión
Los resultados de este estudio indican que la diversidad del ensamble de mamíferos medianos y grandes en esta zona no es afectada de manera evidente por el manejo forestal de tala de impacto reducido. Esto concuerda con lo reportado por otros estudios en los que los mamíferos medianos y grandes pueden ser resilientes al manejo (Azevedo-Ramos et al., 2006; Cruz-Espinoza et al., 2012). Así mismo, estos resultados coinciden con lo reportado en el meta-análisis hecho por Putz et al. (2012) en el que de 85 a 100% de la riqueza de aves, mamíferos (en su mayoría pequeños) e invertebrados es mantenida después de la aplicación de este tipo de manejo en bosques tropicales.
Con respecto a la diversidad β, los valores explicados en su mayor parte por el recambio (βrepl) y en menor proporción por diferencias en riqueza (βrich), reafirman que el número de especies en ambas condiciones es similar. Esta similitud en la diversidad β, entre y dentro de condiciones, sugiere que el recambio de especies no está influenciado por el manejo forestal y que más bien, factores como la heterogeneidad topográfica y ambiental del paisaje pueden favorecer la actividad diferencial de la fauna (Briones-Salas, 2000; García-Mendoza et al., 2004; Gittleman et al., 2001). La disimilitud del sitio M3 con los demás se debió a los registros de varios individuos de D. angulatus, especie gregaria que durante el periodo de estudio sólo se presentó en esta localidad (fig. 3). En los registros restantes se encuentran especies herbívoras (e.g., O. virginianus) y carnívoras (e.g., P. concolor) cuya presencia en ambas condiciones sugiere que procesos como la herbívora y la depredación de especies animales se pueden estar manteniendo (Briones-Salas, 2000; Parry et al., 2013; Rumiz, 2010).
Existen una serie de características del sistema de manejo analizado que probablemente se están combinando para generar los resultados obtenidos en términos de los patrones en la diversidad α, β y la similitud de ensambles de mamíferos. Por una parte, está el hecho que el tamaño de las áreas bajo tala de impacto reducido es pequeño y que el manejo no involucra un cambio notable del bosque (Chaudhary et al., 2016). A la par, las áreas bajo manejo se encuentran embebidas en un paisaje en el que se combinan grandes bloques de bosques destinados a la conservación junto con pequeñas áreas donde se realizan otras actividades agrícolas y se implementan diferentes sistemas de tala. Esto puede por una parte amortiguar el impacto de la actividad humana y por otro lado, favorecer la existencia de una heterogeneidad ambiental lo que a su vez puede favorecer la coexistencia de distintas especies animales (Socolar et al., 2016). De esta manera es importante considerar que es el manejo de la región, en un sentido más amplio, el que puede estar favoreciendo que a una escala espacial más reducida la extracción forestal tenga un bajo impacto sobre la fauna (Pazos-Almada y Bray, 2018) manteniendo la diversidad y distribución de mamíferos medianos y grandes que tienen amplios hábitos hogareños (Grebner et al., 2013; Laidlaw, 2000; Naughton-Treves et al., 2003; Pattanavibool y Dearden, 2002; Putz et al., 2012). Por otra parte, al haber establecido un vínculo de trabajo con las comunidades locales se abre la posibilidad de realizar estudios en el futuro que permitan analizar el impacto que distintas configuraciones (e.g., distintas relaciones entre el área bajo manejo y el área de bosque circundante) del paisaje pueden tener sobre la fauna de mamíferos silvestres.
Gracias al compromiso de conservar su patrimonio biocultural, las comunidades de la sierra Juárez han establecido áreas voluntarias de conservación (AVC) que contemplan la designación de espacios para proyectos productivos, ecoturísticos, de servicios ambientales locales, y para la preservación de los recursos naturales, así como espacios destinados exclusivamente al manejo forestal (Bray et al., 2007; Briones-Salas et al., 2016; Fuente-Carrasco et al., 2012). Estos últimos no solo contemplan extracción de madera, sino también involucran acciones como la aplicación de estrategias de prevención y control de plagas, mitigación de incendios forestales, así como acciones de reforestación que contribuyan a mantener la productividad y calidad de los bosques (Bray et al., 2007; Chapela, 2007; Klooster y Masera, 2000;Ortega, 2004). Es importante señalar que la tala de impacto reducido no supone ingresos económicos como los que se podrían obtener aplicando otros regímenes (Chaudhary et al., 2016), por lo que es muy importante el desarrollo de otras estrategias de manejo (e.g., tala rasa, sistemas de retención) y la implementación de estrategias de conservación apoyadas por instituciones nacionales e internacionales (e.g., pagos por servicios ambientales por Conafor, reconocimientos a actividades de conservación por la WWF) para generar ingresos económicos adicionales que contribuyen al beneficio de la región (Briones-Salas et al., 2016; UZACHI, 2003).
Finalmente, aunque la técnica por tala de impacto reducido parece ser un método compatible con la conservación de mamíferos medianos y grandes, es importante evaluar si procesos como las interacciones ecológicas también son mantenidos. Así mismo, se requiere evaluar si los efectos reportados para mamíferos medianos y grandes en este estudio son los mismos para otros taxones. La implementación de programas de evaluación y el monitoreo a largo plazo de la fauna y otros componentes de la biodiversidad podrá brindar un mejor panorama de la efectividad de esta técnica silvicultural para la conservación de la naturaleza.
Agradecimientos
Este estudio se llevó a cabo gracias a las becas otorgadas al primer autor por parte del Consejo Nacional de Ciencia y Tecnología (Núm. 595991) y al programa The Sigma Xi Grant-in-Aid of Research (ID G201603152071095), así como por el apoyo económico de Red PRODEP: “Conservación de la biodiversidad en ambientes antropizados” otorgado a E. Mendoza. Agradecemos a Jesús López Santiago y a Jorge Arturo Núñez Joaquín por su apoyo en el trabajo de campo, así como a las comunidades de las sierra Juárez por habernos permitido desarrollar este trabajo en sus áreas naturales. Este artículo forma parte de los requisitos para que EHR obtuviera el grado en el programa de Maestría en Ciencias en Ecología Integrativa de la Universidad Michoacana de San Nicolás de Hidalgo.
Referencias
Álvarez, S. y Rubio, A. (2013). Línea base de carbono en bosque mixto de pino-encino de la sierra Juárez (Oaxaca, México): Aplicación del modelo CO2FIX v. 3.2. Serie Ciencias Forestales y del Ambiente, 19, 125–137. https://doi.org/10.5154/r.rchscfa.2012.01.005
Azevedo-Ramos, C., Carvalho, De, O. y Benedito, D. A. (2006). Short-term effects of reduced-impact logging on eastern Amazon fauna. Forest Ecology and Management, 232, 26–35. https://doi.org/10.1016/j.foreco.2006.05.025
Benchimol, M. (2016). Medium and large – sized mammals. En T. H. Larsen (Ed.), Core standard methods for rapid biological field assessment (pp. 37–48). Arlington: Conservation International.
Bray, D. B., Merino, P. L. y Barry, D. (2007). El manejo comunitario en sentido estricto: las empresas forestales comunitarias de México. En D. Bray, L. Merino y D. Barry (Eds.), Los bosques comunitarios de México: manejo sustentable de paisajes forestales (pp. 21–49). México D.F.: Secretaría de Medio Ambiente y Recursos Naturales/ Instituto Nacional de Ecología/ Instituto de Geografía-Universidad Nacional Autónoma de México/ Consejo Civil Mexicano para la Silvicultura Sostenible A.C./ Florida International University.
Briones-Salas, M. (2000). Los mamíferos de la región Sierra Norte de Oaxaca, México. Oaxaca, México. Recuperado el 20 de mayo de 2018 de: http://www.conabio.gob.mx/institucion/proyectos/resultados/InfR104.pdf
Briones-Salas, M., Cortés-Marcial, M. y Lavariega, M. C. (2015). Diversidad y distribución geográfica de los mamíferos terrestres del estado de Oaxaca, México. Revista Mexicana de Biodiversidad, 86, 685–710. https://doi.org/10.1016/j.rmb.2015.07.008
Briones-Salas, M., Lavariega-Nolasco, M. C., Cortés-Marcial, M., Monroy-Gamboa, A. G. y Masés-García, C. A. (2016). Iniciativas de conservación para los mamíferos de Oaxaca, México. En M. Briones-Salas, Y. Hortelano-Moncada, G. Magaña-Cota, G. Sánchez-Rojas, y J. E. Sosa-Escalante (Eds.), Riqueza y conservación de los mamíferos en México a nivel estatal, Vol. I (pp. 329–366). Ciudad de México: Instituto de Biología, Universidad Nacional Autónoma de México/ Asociación Mexicana de Mastozoología A.C./ Universidad de Guanajuato.
Cardoso, P., Rigal, F. y Carvalho, J. C. (2017). BAT: Biodiversity assessment tools. Recuperado el 24 de octubre de 2017 de: https://cran.r-project.org/package=BAT
Carvalho, J. C., Cardoso, P. y Gomes, P. (2012). Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Global Ecology and Biogeography, 21, 760–771. https://doi.org/10.1111/j.1466-8238.2011.00694.x
Ceballos, G. y Oliva, G. (2005). Los mamíferos silvestres de México. Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Universidad Nacional Autónoma de México/ Fondo de Cultura Económica.
Chao, A., Gotelli, N. J., Hsieh, T. C., Sander, E. L., Ma, K. H., Colwell, R. K. y Ellison, A. M. (2014). Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecological Monographs, 84, 45–67. https://doi.org/10.1890/13-0133.1
Chao, A. y Jost, L. (2012). Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size. Ecology, 93, 2533–2547. https://doi.org/10.1890/11-1952.1
Chapela, F. (2007). El manejo forestal comunitario indígena en la sierra de Juárez, Oaxaca. En D. B. Bray, L. Merino-Pérez y D. Barry (Eds.), Los bosques comunitarios de México: manejo sustentable de paisajes forestales (pp. 123–145). México D.F.: Secretaría de Medio Ambiente y Recursos Naturales/ Instituto Nacional de Ecología/ Consejo Civil Mexicano para la Silvicultura Sostenible/ Instituto de Geografía-UNAM/ Florida International Institute.
Chaudhary, A., Burivalova, Z., Koh, L. P. y Hellweg, S. (2016). Impact of forest management on species richness: Global meta-analysis and economic Trade-Offs. Scientific Reports, 1–10. https://doi.org/10.1038/srep23954
Cruz-Espinoza, A., González-Pérez, G. E. y Ronel-Vázquez, P. (2012). Nota de la variación en la riqueza específica de mamíferos entre áreas de conservación y de aprovechamiento forestal en la sierra Madre de Oaxaca. Therya, 3, 327–332. https://doi.org/10.12933/therya-12-88
Dirzo, R. y Miranda, A. (1990). Contemporary neotropical defaunation and sorest structure, function, and diversity-A sequel to John Terborgh. Conservation Biology, 4, 444–447. https://doi.org/10.1111/j.1523-1739.1990.tb00320.x
Dirzo, R., Young, H. S., Galetti, M., Ceballos, G., Isaac, N. J. B. y Collen, B. (2014). Defaunation in the anthropocene. Science, 401, 401–406. https://doi.org/10.1126/science.1251817
FAO (Organización de las Naciones Unidas para la Alimentación y la Agricultura). (2015). Global forest resources assessment 2015: how are the world’s forests changing? Rome: Food and Agriculture Organization of the United Nations.
Fuente-Carrasco, M. E., Ruiz-Aquino, F. y Aquino-Vásquez, C. (2012). Conocimiento indígena contemporáneo y patrimonio biocultural en la sierra Juárez de Oaxaca: aportaciones empíricas y analíticas hacia la sustentabilidad. Oaxaca, México: Universidad de la Sierra Juárez.
Galetti, M. y Dirzo, R. (2013). Ecological and evolutionary consequences of living in a defaunated world. Biological Conservation, 163, 1–6. https://doi.org/10.1016/j.biocon.2013.04.020
García-Mendoza, A. J., Ordoñez, M. J. y Briones-Salas, M. (2004). Biodiversidad de Oaxaca. México D.F.: Instituto de Biología-Universidad Nacional Autónoma de México/ Fondo Oaxaqueño para la Conservación de la Naturaleza/ World Wildlife Fund.
Gibson, L., Lee, T. M., Koh, L. P., Brook, B. W., Gardner, T. A., Barlow, J. et al. (2011). Primary forests are irreplaceable for sustaining tropical biodiversity. Nature, 478, 378–381. https://doi.org/10.1038/nature10425
Gittleman, J. L., Funk, S., Macdonald, D. y Wayne, R. (2001). Carnivore conservation. United Kigdom: Cambridge University Press.
Grebner, D. L., Bettinger, P. y Siry, J. P. (2013). Introduction to forestry and natural resources. San Diego, California: Academic Press. https://doi.org/10.1016/C2010-0-64966-2
Grillo, O. y Gianfranco, V. (2011). Biodiversity loss in a changing planet. Rijeka, Croatia: InTech. Recuperado el 25 de mayo de 2018 de: http://www.intechopen.com
Günter, S., Weber, M., Stimm, B. y Mosandl, R. (2011). Silviculture in the Tropics. Freising, Germany: Springer.
Hernández-Huerta, A. (1992). Los carnívoros y sus perspectivas de conservación en las áreas protegidas de México. Acta Zoológica Mexicana, 54, 1–23.
Hsieh, T. C., Ma, K. H. y Chao, A. (2016). iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods in Ecology and Evolution, 7, 1451–1456. https://doi.org/10.1111/2041-210X.12613
Inskip, C. y Zimmermann, A. (2009). Human-felid conflict: a review of patterns and priorities worldwide. Oryx, 43, 18–34. https://doi.org/10.1017/S003060530899030X
ITTO. (2018). Reduced impact logging. Recuperado el 1 de mayo de 2018 de: http://www.itto.int/feature15/
Klooster, D. y Masera, O. (2000). Community forest management in Mexico: carbon mitigation and biodiversity conservation through rural development. Global Environmental Change, 10, 259–272. https://doi.org/10.1016/S0959-3780(00)00033-9
Laidlaw, R. K. (2000). Effects of habitat disturbance and protected areas on mammals of Peninsular Malaysia. Conservation Biology, 14, 1639–1648. https://doi.org/10.1111/j.1523-1739.2000.99073.x
Legendre, P. y Legendre, L. (2012). Numerical Ecology (3rd Ed.). Oxford, GB: Elsevier. https://doi.org/10.1016/c2010-0-66470-4
Magurran, A. E. (2004). Measuring biological diversity. Oxford, GB: Blackwell Science.
Miller, B., Dugelby, B., Foreman, D., Martinez, C., Noss, R., Phillips, M. et al. (2001). The importance of large carnivores to healthy ecosystems. Endangered Species Update, 18, 202–210.
Miller, S. D., Goulden, M. L., Hutyra, L. R., Keller, M., Saleska, S. R., Wofsy, S. C. et al. (2011). Reduced impact logging minimally alters tropical rainforest carbon and energy exchange. Proceedings of the National Academy of Sciences, 108, 19431–19435. https://doi.org/10.1073/pnas.1105068108
Morrison, J. C., Sechrest, W., Dinerstein, E., Wilcove, D. S. y Lamoreux, J. F. (2007). Persistence of large mammal faunas as indicators of global human impacts. Journal of Mammalogy, 88, 1363–1380. https://doi.org/10.1644/06-MAMM-A-124R2.1
Naughton-Treves, L., Mena, J. L., Treves, A., Alvarez, N. y Radeloff, V. C. (2003). Wildlife survival beyond park boundaries: the impact of slash-and-burn agriculture and hunting on mammals in Tambopata, Peru. Wildlife Survival in Tropical Agroforest Ecosystems, 17, 1106–1117. https://doi.org/10.1046/j.1523-1739.2003.02045.x
Ojasti, O. (2000). Manejo de fauna silvestre neotropical (Vol. 144). Washington: SI/MAB Series # 5. Smithsonian Institution/MAB Biodiversity Program. https://doi.org/10.4067/S0370-41062004000400012
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D. et al. (2018). Vegan: community ecology package. Recuperado el 24 de octubre de 2018 de: https://cran.r-project.org/package=vegan
Ortega, L. P. (2004). Las comunidades indígenas forestales de la sierra de Juárez Oaxaca, México. Estudio de caso sobre innovación participativa. México D.F.: Programa de Desarrollo Forestal Comunitario PROCYMAF.
Ortiz-Pérez, M. A., Hernández-Santana, J. R. y Figueroa-Mah-Eng, J. M. (2004). Reconocimiento fisiográfico y geomorfológico. En A. J. García-Mendoza, M. J. Ordoñez y M. Briones-Salas (Eds.), Biodiversidad de Oaxaca (pp. 43–54). México D.F.: Instituto de Biología-Universidad Nacional Autónoma de México/ Fondo Oaxaqueño para la Conservación de la Naturaleza/ World Wildlife Fund.
Parry, L., Barlow, J., Peres, C. A., Parry, L., Barlow, J. y Peres, C. A. (2013). Brazilian Amazon large-vertebrate assemblages of primary and secondary forests in the Brazilian Amazon. Journal of Tropical Ecology, 23, 653–662. https://doi.org/10.1017/S0266467407004506
Pattanavibool, A. y Dearden, P. (2002). Fragmentation and wildlife in montane evergreen forests, northern Thailand. Biological Conservation, 107, 155–164. https://doi.org/10.1016/s0006-3207(02)00056-3
Pazos-Almada, B. y Bray, D. B. (2018). Community-based land sparing: territorial land-use zoning and forest management in the Sierra Norte of Oaxaca, Mexico. Land Use Policy, 78, 219–226. https://doi.org/10.1016/j.landusepol.2018.06.056
Powell, R. A. (2012). Movements, home ranges, activity, and dispersal. En L. Boitani y R. A. Powell (Eds.), Carnivore ecology and conservation: a handbook of techniques (pp. 188–217). New York: Oxford University Press. https://doi.org/10.1093/acprof:oso/9780199558520.001.0001
Putz, F. E., Redford, K. H., Robinson, J. G., Fimbel, R. y Blate, G. M. (2000). Biodiversity conservation in the context of tropical forest management. Washington D.C.: The World Bank.
Putz, F. E., Sist, P., Fredericksen, T. y Dykstra, D. (2008). Reduced-impact logging: challenges and opportunities. Forest Ecology and Management, 256, 1427–1433. https://doi.org/10.1016/j.foreco.2008.03.036
Putz, F. E., Zuidema, P. A., Synnott, T., Peña-Claros, M., Pinard, M. A., Sheil, D. et al. (2012). Sustaining conservation values in selectively logged tropical forests: the attained and the attainable. Conservation Letters, 5, 296–303. https://doi.org/10.1111/j.1755-263X.2012.00242.x
R Core Team. (2018). R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Recuperado el 22 de febrero de 2018 de: https://www.r-project.org/
Ramírez-Pulido, J., Noé, G. R., Gardner, A. L. y Arroyo-Cabrales, J. (2014). List of recent land mammals of Mexico. Texas, United States: Special Publications of the Museum of Texas Tech University. https://doi.org/10.5962/bhl.title.142891
Reid, A. F. (2009). A field guide to the mammals of Central America and Southeast Mexico (2nd. Ed.). New York: Oxford University Press.
Ripple, W. J., Estes, J. A., Beschta, R. L., Wilmers, C. C., Ritchie, E. G., Hebblewhite, M. et al. (2014). Status and ecological effects of the world’s largest carnivores. Science, 343, 1241484. https://doi.org/10.1126/science.1241484
Rowcliffe, J. M., Field, J., Turvey, S. T. y Carbone, C. (2008). Estimating animal density using camera traps without the need for individual recognition. Journal of Applied Ecology, 45, 1228–1236. https://doi.org/10.1111/j.1365-2664.2008.0
Rumiz, D. I. (2010). Roles ecológicos de los mamíferos medianos y grandes. En R. B. Wallace, H. Gómez, Z. R. Porcel y D. I. Rumiz (Eds.), Distribución, ecología y conservación de los mamíferos medianos y grandes de Bolivia (pp. 53–73). Santa Cruz de la Sierra, Bolivia: Centro de Ecología Difusión Simón I. Patiño.
Rumiz, D. I., Eulert, C. F. y Arispe, R. (1998). Evaluación de la diversidad de mamíferos medianos y grandes en el Parque Nacional Carrasco (Cochabamba – Bolivia). Revista Boliviana de Ecología y Conservación Ambiental, 4, 77–90.
Sistema Estatal de Información Forestal. (2015). Estudio de cuenca de abasto para el desarrollo industrial forestal maderable de la región sierra Juárez, Oaxaca. Oaxaca, México: Comisión Nacional Forestal y Colegio de Profesionales Forestales de Oaxaca, A.C.
Socolar, J. B., Gilroy, J. J., Kunin, W. E. y Edwards, D. P. (2016). How should beta-diversity inform biodiversity conservation? Trends in Ecology y Evolution, 31, 67–80. https://doi.org/http://dx.doi.org/10.1016/j.tree.2015.11.005
Torres-Colín, R. (2004). Tipos de vegetación. En A. J. García-Mendoza, M. J. Ordoñez, y M. Briones-Salas (Eds.), Biodiversidad de Oaxaca (pp. 105–117). México D.F.: Instituto de Biología-Universidad Nacional Autónoma de México/ Fondo Oaxaqueño para la Conservación de la Naturaleza/ World Wildlife Fund.
UZACHI (Unión de Comunidades Productoras Forestales Zapotecos-Chinantecos de la Sierra Juárez). (2003). Programa de manejo forestal persistente para el aprovechamiento maderable de la comunidad de Capulálpam de Méndez, Ixtlán, Oaxaca. Oaxaca: U. de comunidades productoras forestales Z. y Chinantecas.


