Diversidad y conservación de los mamíferos terrestres de Chiapas, México
Consuelo Lorenzo⁎ ✉ , Jorge Bolaños-Citalán, Eugenia Sántiz, Darío Navarrete
Resumen
Evaluamos el estado actual de la diversidad, distribución y conservación de mamíferos terrestres en Chiapas a través de registros históricos y actuales. Chiapas es el segundo estado mexicano más diverso en este grupo, con 210 especies (42.3% del total nacional), de las cuales 9 son endémicas del estado. Los quirópteros representan 108 especies, los roedores 51 y los carnívoros 19. Las regiones más diversas en mamíferos en Chiapas presentan una gran variedad de ambientes, principalmente bosques templados, bosques mesófilos y selvas, y una gran heterogeneidad topográfica en las subprovincias de los Altos de Chiapas, Sierras del Sur y Norte de Chiapas, así como en selvas en la Sierra Lacandona. Los patrones de distribución se relacionan con la historia evolutiva de las especies, sus requerimientos de hábitat y los efectos de la actividad humana (pérdida de hábitat, cacería sin control, tráfico ilegal e introducción de especies exóticas). Para fundamentar las prácticas de conservación requeridas es esencial generar información más completa sobre la distribución, la abundancia de las poblaciones de mamíferos endémicos, amenazados y en peligro de extinción, así como su respuesta a la pérdida y fragmentación del hábitat, la sobreexplotación y otras actividades humanas.
Palabras clave
Diversity and conservation of terrestrial mammals from Chiapas, Mexico
Abstract
We evaluated the current state of diversity, distribution and conservation of terrestrial mammals in Chiapas, through historical and current records. Chiapas is the second Mexican state more diverse in this group, with 210 species (42.3% of the national total), of which 9 are endemic to the state. The bats represent 108 species, rodents 51 and carnivores 19. The most diverse regions in mammals in Chiapas maintain a variety of environments mainly temperate forest, cloud forest and tropical forest and large topographic heterogeneity in the subprovinces of the Chiapas highlands, mountains of the southern and northern Chiapas, as well as the Lacandon Sierra forests. The patterns of distribution of mammals of Chiapas are related with the evolutionary history of the species, their habitat requirements, and the effects of human activity (loss of habitat, hunting without control, illegal trafficking, and introduction of exotic species). To substantiate the required conservation practices, it is essential to generate more comprehensive information on the distribution, abundance of populations of endemic mammals, threatened and in danger of extinction, as well as its response to the loss and fragmentation of habitat, overexploitation, and other human activities.
© 2017 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Keywords:
Biodiversity; Conservation; Mammals; Mexico; Species richness
Introducción
El estado de Chiapas es ampliamente reconocido como una de las regiones con mayor riqueza de mamíferos silvestres en México ( Ceballos y Oliva, 2005; Naranjo, Lorenzo y Horváth, 2005 ) debido a su historia biogeográfica, la diversidad de hábitats, los cambios climáticos glaciales e interglaciales y la dinámica tectónica. Esto ha permitido un ciclo de especiación de gran importancia para este grupo taxonómico ( Gutiérrez-García y Vázquez-Domínguez, 2013 ). El conocimiento sobre los mamíferos en Chiapas se remonta varios milenios atrás, cuando los grupos humanos que colonizaron la región utilizaron intensamente a la fauna silvestre como fuente de alimento y muchos otros productos derivados, como pieles y huesos, necesarios para su supervivencia. Posteriormente, los mayas y sus descendientes dieron cuenta del gran significado de los mamíferos en su cultura al emplear materiales y valores derivados de este grupo, que aún hoy día podemos observar en el arte, lenguaje, nomenclatura, tradiciones culinarias y rituales de los grupos étnicos, que constituyen una proporción muy significativa de la población estatal ( Naranjo et al., 2005; Naranjo, Lorenzo, Bolaños-Citalán y Horváth 2016 ).
Durante los últimos 50 años el conocimiento sobre los mamíferos en el estado se incrementó notablemente gracias a publicaciones pioneras, como las de Álvarez-del Toro (1966, 1991) y Álvarez y Álvarez (1991) , seguidas por aportaciones de los cada vez más numerosos mastozoólogos que han trabajado en el estado, provenientes tanto de instituciones estatales como federales (Universidad de Ciencias y Artes de Chiapas, Universidad Autónoma de Chiapas, El Colegio de la Frontera Sur, Universidad Nacional Autónoma de México, Instituto Politécnico Nacional, entre otras). Gracias al trabajo de estos mastozoólogos, Chiapas es en la actualidad uno de los estados del sur del país con mejores niveles de conocimiento en cuanto a su mastofauna. Sin embargo, indudablemente aún falta mucho por hacer en materia de investigación mastozoológica con miras a la conservación y uso sustentable de las especies de este grupo.
El presente estudio tiene por objeto presentar una evaluación del estado que guarda la diversidad de mamíferos silvestres terrestres (incluyendo voladores) en Chiapas. Se realiza un análisis histórico del conocimiento de la mastofauna, de los aspectos de la riqueza (con la actualización de la lista taxonómica de especies) y de la distribución geográfica de especies (por regiones fisiográficas, altitud y coberturas de vegetación, y uso de suelo), de la problemática en las comunidades mastofaunísticas y necesidades de conservación e investigación.
Materiales y métodos
Chiapas se ubica entre los 14°32′ y los 17°59′ N y entre los 90°22′ y los 94°14′ O. Su fisonomía la determinan 2 grandes cadenas montañosas que la recorren con orientación noroeste-sureste: 1) la Sierra Madre de Chiapas, que corre casi paralela a la costa del océano Pacífico e incrementa su altitud desde aproximadamente los 1,000 m en los límites con Oaxaca hasta más de 4,000 m en la frontera con Guatemala, y 2) la Cordillera Central o Centroamericana, que conforma la Altiplanicie Central o los Altos de Chiapas y se desplaza por la parte central del estado de manera paralela a la Sierra Madre de Chiapas formando la Depresión Central y extendiéndose hacia el norte del estado para formar las Montañas del Norte; esta cordillera proviene de Guatemala (sierra de los Cuchumatanes) y desde ahí penetra en territorio mexicano, alcanzando sus máximas altitudes en el cerro Tzontehuitz, a 2,880 m, cerca de San Cristóbal de Las Casas ( Lorenzo, Kraker-Castañeda y Bolaños-Citalán, 2015 ). Chiapas se divide en 122 municipios que se agrupan en 10 subprovincias fisiográficas: 1) Depresión Central de Chiapas (DC); 2) Llanura Costera de Chiapas y Guatemala (LLC); 3) Llanura del Istmo (LLI); 4) Altos de Chiapas (AC); 5) Llanura y Pantanos Tabasqueños (LLPT); 6) Sierra Lacandona (SL); 7) Sierras Bajas del Petén (SBP); 8) Sierras del Norte de Chiapas (SN); 9) Sierras del Sur de Chiapas (SS), y 10) Volcanes de Centroamérica (VCA; Inegi, 1981).
El clima del estado es consecuencia de esta topografía y presenta una gran variedad. En algunas partes bajas de la costa y en la depresión que se encuentra entre los Altos y la Sierra Madre (el valle formado por el río Grijalva) el clima es cálido y seco, con una temporada lluviosa corta (6 meses). La vegetación es una selva baja espinosa o selva caducifolia en las vegas de los ríos. No obstante, buena parte de esta vegetación original ha sido sustituida por praderas para la cría de ganado bovino. Por su parte, las partes bajas a sotavento de los vientos alisios, en el norte y noreste de la entidad, reciben grandes cantidades de precipitación durante 8 o 10 meses del año, lo que, aunado a las altas temperaturas, favorece la presencia de selvas medianas y altas perennifolias (selva Lacandona). Estas áreas forestales han sufrido fuertes impactos negativos debido a la introducción de ganado bovino o a prácticas no controladas de extracción de madera y cultivos ( Carazo, 2007).
En contraste, y debido a su altitud, las zonas elevadas de la sierra y de los Altos atrapan el remanente de humedad que no se depositó en sus vertientes, por lo que presentan una estacionalidad marcada y característica. Sus inviernos son secos y muy fríos (con temperaturas por debajo de los 0 o C), mientras que los veranos tienden a ser templados y muy húmedos (7 u 8 meses de lluvia). La vegetación de estas regiones se ve dominada por bosques de pino-encino. Ambas cadenas cuentan con bosque mesófilo de montaña, aunque prácticamente ha desaparecido de los Altos, y en la Sierra Madre se encuentra la Reserva de la Biosfera El Triunfo, con más de 100,000 ha de este tipo de bosque. La zona costera, por su parte, está compuesta de manglares y su clima es también tropical ( Inegi, 2015).
Para el análisis histórico del conocimiento de la mastofauna de Chiapas y el estado que guarda, se utilizó la base de datos Global Biodiversity Information Facility ( GBIF, 2013; www.gbif.org ), que contiene información de ejemplares depositados en 57 colecciones nacionales e internacionales, así como una búsqueda bibliográfica. Primeramente, la base de datos fue depurada, eliminando los registros que no presentaron información completa, como localidad, género y especie, número de catálogo de la colección y coordenadas geográficas; posteriormente, se actualizó la taxonomía de algunas especies. Adicionalmente, se incluyeron en la base de datos general los registros de ejemplares recolectados en los últimos 15 años por el equipo de trabajo de los autores, los cuales se encuentran depositados en la Colección Mastozoológica de El Colegio de la Frontera Sur en San Cristóbal de Las Casas, Chiapas, con registro por parte de la Secretaría de Medio Ambiente y Recursos Naturales (CHI-MA-0013-0497).
Para el análisis de datos se realizaron curvas de acumulación de registros y especies en el tiempo, en intervalos de 10 años a partir de los primeros registros formales de colecta (1860). Se elaboró una lista de especies que fue actualizada en su nomenclatura y arreglo sistemático de acuerdo con Ramírez-Pulido, González-Ruiz, Gardner y Arroyo-Cabrales (2014) , y se consideraron los cambios taxonómicos recientes y los nuevos registros de zorrillos ( Dragoo, Bradley, Honeycutt y Templeton, 1993 ), venados (Geist, 1998), roedores ( Carroll y Bradley, 2005; Carroll, Peppers y Bradley, 2005; Voss, 1998 ), musarañas ( Guevara, Sánchez-Cordero, León-Paniagua y Woodman, 2014; Woodman y Timm, 1999 ), murciélagos ( Baird, Marchán-Rivadeneira, Pérez y Baker, 2012; Baker, Solari y Hoffmann, 2002; Escobedo-Morales, León-Paniagua, Arroyo-Cabrales y Greenaway, 2006; Lee, Hoofer y van den Bussche, 2002; Simmons y Handley, 1998 ) y didélfidos (Voss y Jansa, 2003) en Chiapas.
Se identificaron las especies endémicas del estado, así como la situación de conservación y protección de cada especie con base en la lista roja de la Unión Internacional para la Conservación de la Naturaleza ( IUCN, 2015 ), los apéndices de la Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres ( CITES, 2014 ) y la Norma Oficial Mexicana 059 (NOM-059-SEMARNAT-2010; Semarnat, 2010).
Las coordenadas geográficas de todos los registros considerados en la base de datos se proyectaron en los mapas de las subprovincias fisiográficas ( Inegi, 1981 ), de altitud y de uso de suelo, y vegetación ( Inegi, 2015 ) del estado. Esto permitió: 1) identificar áreas de Chiapas poco exploradas o con pocas recolectas para mamíferos; 2) determinar la riqueza de especies, géneros, familias y órdenes a nivel estatal, altitudinal y por subprovincia fisiográfica ( Ortiz-Pérez, Hernández-Santana y Figueroa-Mah Eng, 2004 ), y 3) evaluar los cambios históricos en los diferentes tipos de vegetación y uso de suelo en los que fueron colectados los ejemplares y compararlo con los actuales. Posteriormente se calculó el coeficiente de Jaccard con el método de pares asociados al azar sin peso aritmético (UPGMA, por sus siglas en inglés) para obtener la similitud de especies entre las subprovincias fisiográficas con el programa MVSP 3.22 y se estimó el índice de Whittaker (B W; Magurran, 1988; Whittaker, 1972 ) para obtener la diversidad beta con el programa Species Diversity and Richness 3.02.
Resultados
A partir de las exploraciones biológicas en México, cuyo fin era conocer los recursos naturales con los que contaba la nación, se crea en 1831 el Museo Nacional Mexicano. En 1846 se funda la Smithsonian Institution y se realiza una recolecta intensiva en México, destacando la participación de Spencer Fullerton Baird, Elliot Coues y Edgar A. Mearns. En 1868 se funda la Sociedad Mexicana de Historia Natural y en 1877 se forma la Comisión Geográfica Exploradora, y se exploran diversas regiones del país con apoyo de la Secretaría de Agricultura ( Ramírez-Pulido y González-Ruiz, 2006; Retana-Guiascón, 2006 ). Posteriormente, en los Anales del Museo Nacional de Historia Natural se publican datos para la zoología de Chiapas, en los que se incluye a los mamíferos ( Herrera, 1897 ). Para la década de 1890 se tienen 689 registros de 62 diferentes especies, producto de las recolectas científicas de mamíferos en el estado de Chiapas ( fig. 1).
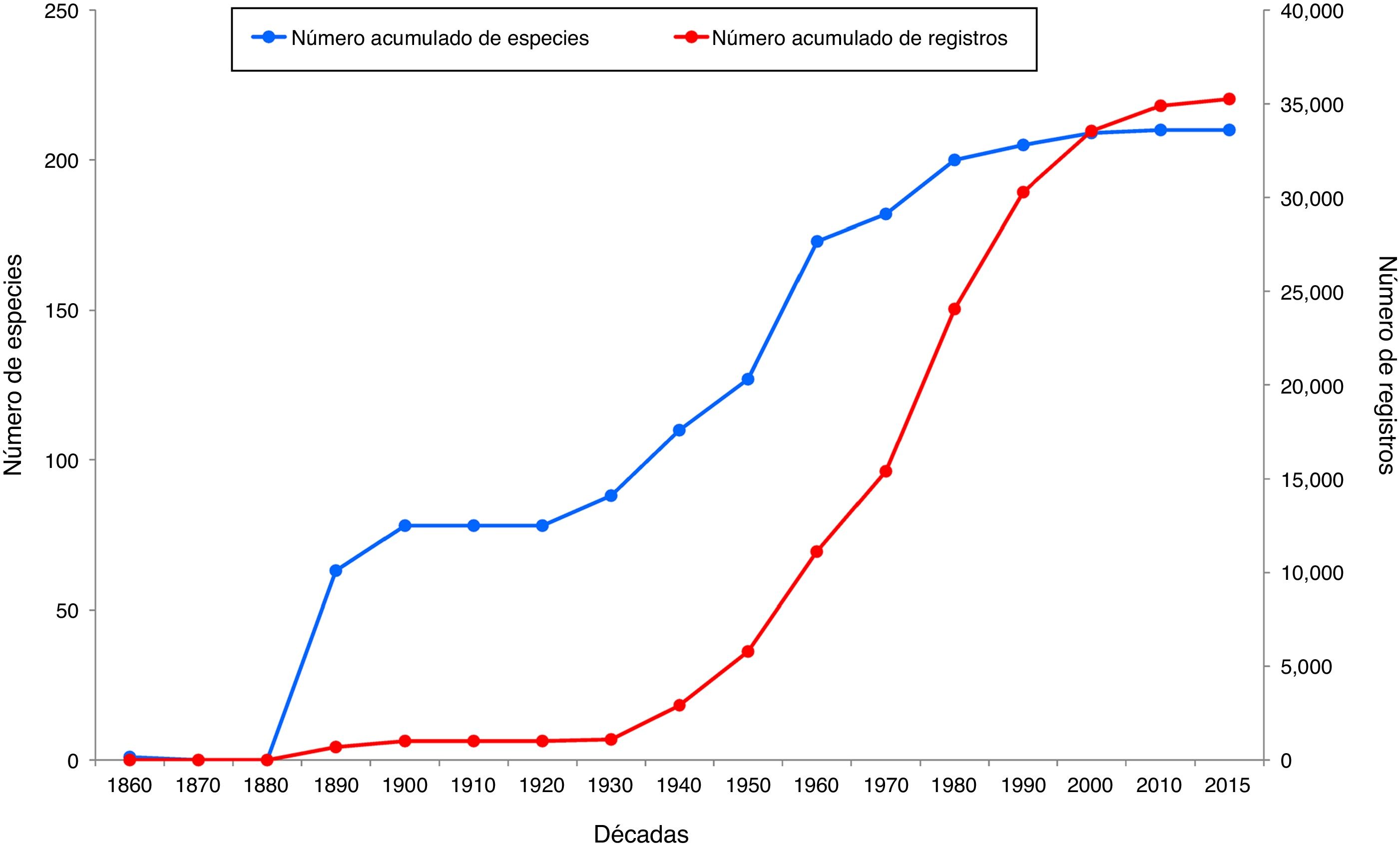
Figura 1
Número acumulado de especies y número acumulado de registros de mamíferos terrestres en el estado de Chiapas, a partir de la primera década de recolecta.
De 1895 a 1915 funcionó el United States Bureau of Biological Survey, dependencia de cuyo trabajo derivaron expediciones con recolectas de mamíferos en Chiapas, supervisadas por C.H. Merriam (jefe de la División de Ornitología y Mamíferos del Departamento de Agricultura de los Estados Unidos), y sus principales recolectores fueron E.W. Nelson y E.A. Goldman ( Ramírez-Pulido y González-Ruiz, 2006 ). Los registros de ejemplares de Chiapas datan de 1895 a 1904 por Nelson y Goldman, quienes, junto con Merriam, describieron 93 especies y 271 subespecies de mamíferos para México y Chiapas ( Ramírez-Pulido y González-Ruiz, 2006 ).
En la década de 1930 comienza el trabajo de autores mexicanos en el campo taxonómico, con L. Martínez y B. Villa, quienes describen varias especies de mamíferos ( Ramírez-Pulido y González-Ruiz, 2006 ); también existen registros de esa década de N. Hartweg de la Universidad de Michigan, reflejándose un incremento en el esfuerzo de muestreo en la recolecta de mamíferos en Chiapas, y se adicionan 10 especies y 93 registros ( fig. 1).
A partir de 1940 se intensificaron las exploraciones mastozoológicas en Chiapas. En la curva acumulada de registros para esta fecha se muestran 2,924 registros, con recolectas de M. Álvarez-del Toro, B. Villa y H.O. Wagner; en la década de 1950, con A. Barrera, A. Johnson, A.A. Alcorn, D.C. Carter, R.W. Dickerman, E.T. Hooper y T. Álvarez, asciende a 5,796 registros, y se alcanzan 24,075 en 1980. Posteriormente, el incremento fue paulatino hasta 2015. En cuanto al número acumulado de especies, en la década de 1900 se conocían 78 especies, cifra que aumenta para 1930 en 10 especies más, incrementándose a 200 en 1980, década a partir de la cual la adición de nuevas especies es menor. A partir del año 2000, el número acumulado de registros rebasa el número acumulado de especies, lo que se ha mantenido hasta 2015, ya que no se han descrito más especies para Chiapas ( fig. 1).
La base de datos de mamíferos terrestres para el estado de Chiapas estuvo conformada por 35,972 registros de ejemplares albergados en colecciones científicas, de los cuales 22,726 registros (63.2%) pertenecen a 10 colecciones nacionales y 13,246 registros (36.8%) a 47 colecciones extranjeras. Las colecciones mastozoológicas que cuentan con mayor número de registros de especies de mamíferos colectados en Chiapas se encuentran en México y son: la Colección Nacional de Mamíferos, Universidad Nacional Autónoma de México (CNMA: 7,475), Colección Mastozoológica de El Colegio de la Frontera Sur (ECO-SC-M 4,822), Colección Mastozoológica de la Escuela Nacional de Ciencias Biológicas del Instituto Politécnico Nacional (IPN: 4,728) y la Colección Zoológica Regional de la Secretaría de Medio Ambiente, Vivienda e Historia Natural (CZRMA: 3,936; tabla 1 ). Las colecciones extranjeras que cuentan con el mayor número de registros se encuentran en las siguientes instituciones: University of Kansas Biodiversity Institute (KU: 2,898), University of Michigan, Museum of Zoology (UMMZ: 2,381), Los Angeles County Museum of Natural History (LACM: 1,406) y National Museum of Natural History, Smithsonian Institution (NMNH: 1,201; tabla 1 ). Del total de registros, 33,961 (94.4%) cuentan con nombre de colector (916 colectores en total). C. Lorenzo es el colector con mayor número de registros (2,006), seguido por A. Ocaña (1,619), A. Riechers (1,358), A. Gardner (1,352) y R. Medellín (1,304).
Tabla 1
Número de registros de mamíferos terrestres de Chiapas depositados en colecciones científicas nacionales y extranjeras. El nombre de la colección (incluyendo los acrónimos) se tomaron de acuerdo a Lorenzo et al. (2012).
| Acrónimo de la colección | Colecciones | Número de registros |
| CNMA | Universidad Nacional Autónoma de México, Colección Nacional de Mamíferos | 7,475 |
| ECO-SC-M | El Colegio de la Frontera Sur, Colección Mastozoológica | 4,822 |
| ENCB | Instituto Politécnico Nacional, Escuela Nacional de Ciencias Biológicas | 4,728 |
| CZRMA | Secretaría de Medio Ambiente, Vivienda e Historia Natural | 3,936 |
| KU | University of Kansas, Biodiversity Institute | 2,898 |
| UMMZ | University of Michigan, Museum of Zoology | 2,381 |
| LACM | Los Angeles County Museum of Natural History | 1,406 |
| NMNH | National Museum of Natural History, Smithsonian Institution | 1,201 |
| MZ-ICACH | Universidad de Ciencias y Artes de Chiapas, Museo de Zoología | 1,177 |
| TTU | Texas Tech University, The Museum | 855 |
| MVZ | University of California, Berkeley, Museum of Vertebrate Zoology | 736 |
| ROM | Royal Ontario Museum | 695 |
| MHP | Fort Hays State University, Museum of the High Plains | 476 |
| MSU | Michigan State University | 465 |
| MZFC-M | Universidad Nacional Autónoma de México, Museo de Zoología «Alfonso L. Herrera» | 453 |
| ASVRC | Angelo State University | 293 |
| ASU | Arizona State University | 206 |
| CAS | California Academy of Sciences | 195 |
| MCZ | Harvard University Provider | 182 |
| FMNH | Field Museum | 176 |
| UW-WSM | University of Washington, Burke Museum | 170 |
| INATURALIST | I Naturalist Research Grade Observations | 144 |
| LSUMZ | Louisiana State University Museum of Natural Science | 120 |
| UAM-I | Universidad Autónoma Metropolitana, Unidad Iztapalapa | 114 |
| HMHM | Hungarian Natural History Museum | 105 |
| TCWC | Texas A & M University, Texas Cooperative Wildlife Collection | 97 |
| CU | Cornell University | 85 |
| CM | Carnegie Museum of Natural History | 82 |
| MMNH | University of Minnesota, J. F. Bell Museum of Natural History | 73 |
| AMNH | American Museum of Natural History | 48 |
| CU | Cornell University | 37 |
| UF | Florida Museum Natural History | 22 |
| MSB | University of New Mexico, Museum of Southwestern Biology | 20 |
| UND | University of North Dakota | 16 |
| OC | Occidental College, More Laboratory of Zoology | 11 |
| UANL | Universidad Autónoma de Nuevo León | 10 |
| CRD | Instituto Politécnico Nacional, Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Unidad Durango | 9 |
| OSU | Oklahoma State University Museum | 8 |
| ISU | Illinois State University | 6 |
| CUMV | Cornell University Museum of Vertebrates | 5 |
| JDS | James Dale Smith (colección personal) | 4 |
| Naturgucker | Naturgucker.de | 4 |
| PSM | University of Puget Sound, Slater Museum of Natural History | 4 |
| QM | Queensland Museum | 4 |
| CAEC | Charlotte Area Educational Consortium | 3 |
| MHNG | Muséum d’Histoire Naturelle de la Ville de Genève | 2 |
| MMMM | Manitoba Museum of Man and Nature | 2 |
| OSUFW | Oregon State University, Fish and Wildlife Department | 2 |
| UCLA | University of California at Los Angeles | 2 |
| VMHSUC | Vertebrate Museum Humboldt State University | 2 |
| ZOOMAT | Zoologíco Miguel Alvarez del Toro | 2 |
| BJH | Private Collection of Bruce J. Hayward, Silver | 1 |
| BYU | Brigham Young University, Life Science Museum | 1 |
| YPM | Yale University Peabody Museum | 1 |
La composición actual de la mastofauna de Chiapas está conformada por 210 especies, 119 géneros, 30 familias y 11 órdenes ( apéndice 1 ), lo que representa el 42.3% de la riqueza nacional de especies de mamíferos terrestres ( Ramírez-Pulido et al., 2014 ). Esto implica que Chiapas ocupa el segundo lugar nacional en diversidad de mamíferos terrestres, después del estado de Oaxaca ( Briones-Salas, Cortés-Marcial y Lavariega, 2015; Retana-Guiascón y Lorenzo, 2002 ).
Los órdenes de mamíferos terrestres con mayor número de registros en Chiapas son: Chiroptera, Rodentia y Carnivora. Chiroptera aporta el mayor número de especies (108) y géneros (60), los cuales representan el 51.4 y el 50% de la diversidad de mamíferos terrestres, respectivamente. Rodentia aporta 51 especies y 21 géneros, que constituyen, respectivamente, el 24.3 y el 17.5% de la riqueza estatal. Por último, Carnivora aporta 19 especies y 17 géneros, que equivalen al 9.05 y al 14.2% de la mastofauna de la entidad, respectivamente. Los restantes 8 órdenes de mamíferos presentes en Chiapas que están representados por 10 especies o menos son: Soricomorpha (musarañas, 10); Didelphimorphia (tlacuaches, 8); Artiodactyla (venados y pecaríes, 4); Primates (monos, 3); Cingulata (armadillos, 2); Pilosa (hormigueros, 2); Lagomorpha (conejos, 2), y Perissodactyla (tapires, 1; tabla 2).
Tabla 2
Número de registros por categoría taxonómica de mamíferos terrestres de Chiapas.
| Orden | Familia | Número de registros | Porcentaje del número de registros | Número de especies |
| Rodentia | Cricetidae | 11,238 | 31.24 | 35 |
| Heteromyinae | 1,746 | 4.85 | 5 | |
| Sciuridae | 521 | 1.44 | 5 | |
| Cuniculidae | 289 | 0.8 | 1 | |
| Geomydae | 184 | 0.5 | 2 | |
| Dasyproctinae | 37 | 0.1 | 2 | |
| Erethizontidae | 23 | 0.06 | 1 | |
| Total Rodentia | 14,038 | 38.99 | 51 | |
| Chiroptera | Phyllostomidae | 15,207 | 42.34 | 52 |
| Molossidae | 1,065 | 2.96 | 15 | |
| Vespertilionidae | 965 | 2.68 | 23 | |
| Moormoopidae | 921 | 2.56 | 5 | |
| Emballonuridae | 763 | 2.19 | 9 | |
| Noctilionidae | 298 | 0.82 | 2 | |
| Natalidae | 114 | 0.31 | 1 | |
| Thyropteridae | 19 | 0.05 | 1 | |
| Total Chiroptera | 19,352 | 53.91 | 108 | |
| Carnivora | Procyonidae | 244 | 0.67 | 4 |
| Felidae | 161 | 0.44 | 5 | |
| Canidae | 110 | 0.3 | 2 | |
| Mustelidae | 95 | 0.26 | 4 | |
| Mephitidae | 73 | 0.2 | 4 | |
| Total Carnivora | 683 | 1.87 | 19 | |
| Artiodactyla | Tayassuidae | 347 | 0.96 | 2 |
| Cervidae | 265 | 0.73 | 2 | |
| Total Artiodactyla | 612 | 1.69 | 4 | |
| Didelphimorphia | 517 | 1.43 | 8 | |
| Total Didelphimorphia | 517 | 1.43 | 8 | |
| Primates | 265 | 0.73 | 3 | |
| Total Primates | 265 | 0.73 | 3 | |
| Soricomorpha | 188 | 0.52 | 10 | |
| Total Soricomorpha | 188 | 0.52 | 10 | |
| Lagomorpha | 154 | 0.42 | 2 | |
| Total Lagomorpha | 154 | 0.42 | 2 | |
| Cingulata | 92 | 0.25 | 2 | |
| Total Cingulata | 92 | 0.25 | 2 | |
| Pilosa | 36 | 0.1 | 2 | |
| Total Pilosa | 36 | 0.1 | 2 | |
| Perissodactyla | 35 | 0.09 | 1 | |
| Total Perissodactyla | 35 | 0.09 | 1 | |
De las 210 especies de mamíferos terrestres presentes en Chiapas, 9 son endémicas del estado: 3 musarañas ( Cryptotis lacandonensis, Sorex stizodon y S. sclateri), 2 murciélagos (Rhogeessa bickhami y R. genowaysi) y 4 roedores (Peromyscus zarhynchus, Tylomys bullaris, T. tumbalensis y Heteromys nelsoni ). Estas especies se distribuyen en las regiones de los Altos, las cañadas de la selva Lacandona, Soconusco y la porción este de la Sierra Madre de Chiapas; algunas de ellas ( R. bickhami, R. genowaysi, T. tumbalensis, C. lacandonensis, S. stizodon y S. sclateri ) conocidas solamente de la localidad tipo ( Ceballos y Rodríguez, 1993; Guevara et al., 2014 ; apéndice 1).
Chiapas cuenta con 66 especies de mamíferos, incluidas en alguna categoría de protección dentro de la Norma Oficial Mexicana NOM-059-SEMARNAT-2010 ( Semarnat, 2010 ): 31 amenazadas (A), 21 sujetas a protección especial (Pr) y 14 en peligro (P). La mayoría de las especies de mamíferos terrestres de Chiapas se encuentran en la lista roja de la Unión Internacional para la Conservación de la Naturaleza ( IUCN, 2015 ) bajo la categoría de menor preocupación, y 32 especies se incluyen en las siguientes categorías de riesgo: datos deficientes (3), vulnerable (7), críticamente en peligro (5), casi amenazada (11) y en peligro (6). En la lista de la Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres se encuentran 21 especies: 8 en el apéndice I, 3 en el apéndice II y 10 en el apéndice III (CITES, 2014; apéndice 1).
Las subprovincias con la mayor cantidad de registros en orden decreciente son: Altos de Chiapas (AC), con 11,977 (33.3%), de los que 638 corresponden a especies endémicas; las Sierras del Sur de Chiapas (SS), con 6,964 (19.3%), de los que 73 corresponden a especies endémicas, y la Sierra Lacandona (SL), con 5,377 (14.9%), de los que 6 son endémicos. En comparación, las menos representadas (cada una con menos de 500 registros) son: Llanura del Istmo (LLI), con 352 (0.9%), y Sierras Bajas del Petén (SBP), con solo 19 (0.05%), ambas sin registros de especies endémicas ( tabla 3; fig. 2).
Tabla 3
Representatividad taxonómica de mamíferos terrestres en las subprovincias fisiográficas de Chiapas.
| Subprovincia fisiográfica | Registros | Órdenes | Familias | Géneros | Número de especies no endémicas (número de registros) | Número de especies endémicas (número de registros) |
| Altos de Chiapas (AC) | 11,977 | 11 | 27 | 97 | 157 (11,339) | 3 (638) |
| Sierras del Sur de Chiapas (SS) | 6,964 | 11 | 29 | 90 | 155 (6,891) | 3 (73) |
| Sierra Lacandona (SL) | 5,377 | 11 | 30 | 105 | 150 (5,371) | 2 (6) |
| Sierras del Norte de Chiapas (SN) | 5,344 | 10 | 25 | 88 | 134 (4,997) | 3 (347) |
| Llanura Costera de Chiapas y Guatemala (LLC) | 2,838 | 11 | 28 | 67 | 113 (2,836) | 1 (2) |
| Depresión Central de Chiapas (DC) | 1,256 | 7 | 17 | 38 | 51 (1,256) | 0 |
| Volcanes de Centroamérica (VCA) | 1,037 | 8 | 20 | 54 | 91 (1,035) | 1 (2) |
| Llanura y Pantanos Tabasqueños (LLPT) | 808 | 8 | 21 | 57 | 75 (808) | 0 |
| Discontinuidad Llanura del Istmo (LLI) | 352 | 6 | 16 | 33 | 44 (352) | 0 |
| Sierras Bajas del Petén (SBP) | 19 | 2 | 6 | 12 | 15 (19) | 0 |
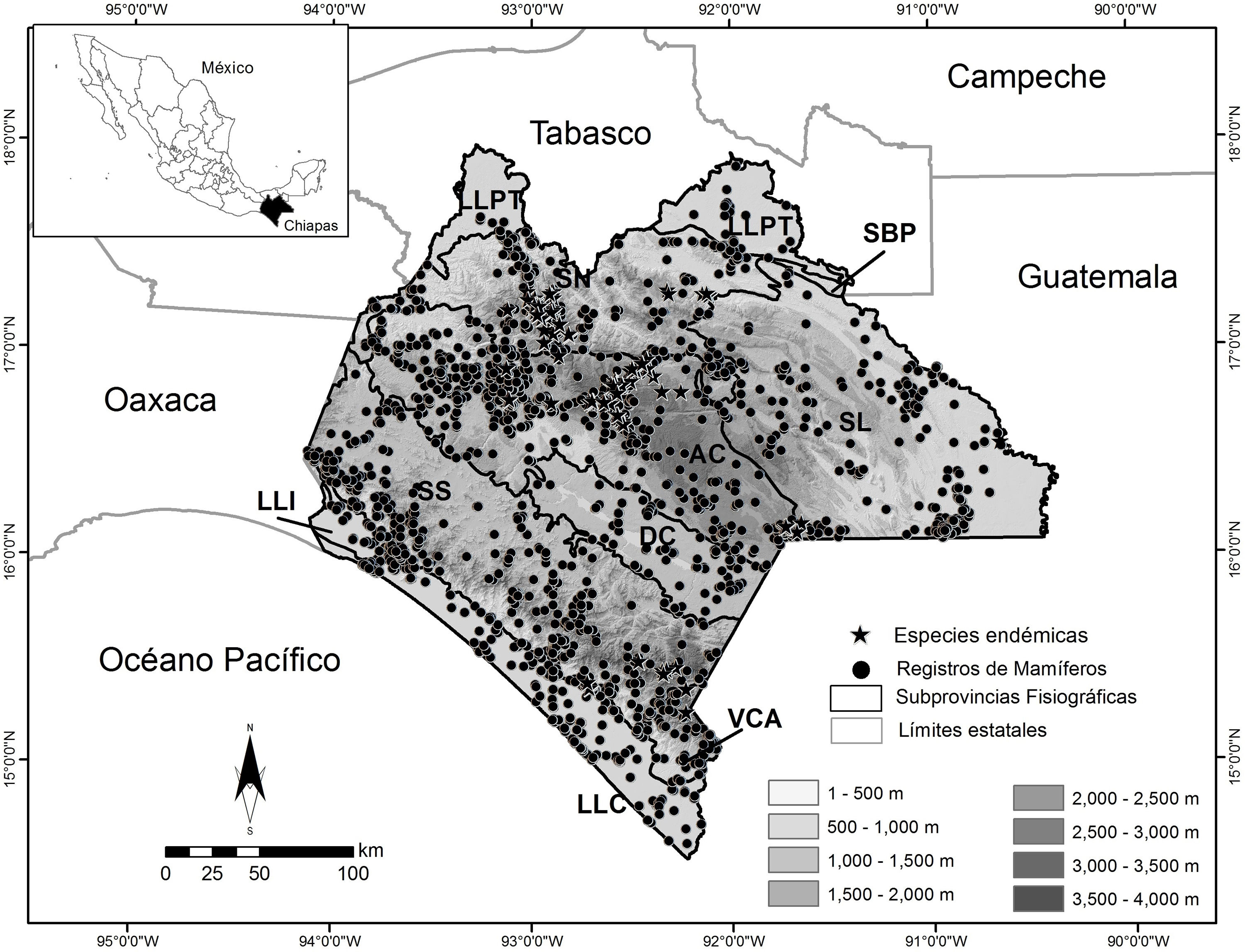
Figura 2
Localización de los mamíferos terrestres registrados en el estado de Chiapas, por subprovincia fisiográfica y altitud.AC: Altos de Chiapas; DC: Depresión Central de Chiapas; LLC: Llanura Costera de Chiapas y Guatemala; LLI: Llanura del Istmo; LLPT: Llanura y Pantanos Tabasqueños; SBP: Sierras Bajas del Petén; SL: Sierra Lacandona; SN: Sierras del Norte de Chiapas; SS: Sierras del Sur de Chiapas; VCA: Volcanes de Centroamérica.
Con respecto a la presencia de las especies en los diferentes tipos de vegetación y uso de suelo se tienen: 184 en vegetación secundaria (VS), 169 en pastizales (PZ), 157 en asentamientos humanos (AH), 149 en áreas agrícolas (AG), 126 en selvas (altas, medianas y bajas, perennifolias, subperennifolias, caducifolias; SE), 108 en bosque mesófilo de montaña (BM), 92 en bosques templados (pino, encino, cedro, oyamel y sus asociaciones; BQ), 52 en hábitats asociados a cuerpos de agua (CA), 49 en manglar (MG), 38 en sitios sin vegetación aparente (SV), 27 en sabana (SA), 17 en praderas de montaña (PM), 14 en tulares (TU) y 4 en vegetación de galería (VG). De las especies endémicas de Chiapas, C. lacandonensis se distribuye en SE; S. sclateri en AG; S. stizodon en AH; R. bickhami en BM; R. genowaysi en AG, BM, PZ; H. nelsoni en AG, BM, VS; P. zarhynchus en AG, BM, BQ, SV, PZ, VS y AH; T. bullaris en AH, y T. tumbalensis en AG, BM, VS (apéndice 1; fig. 3).
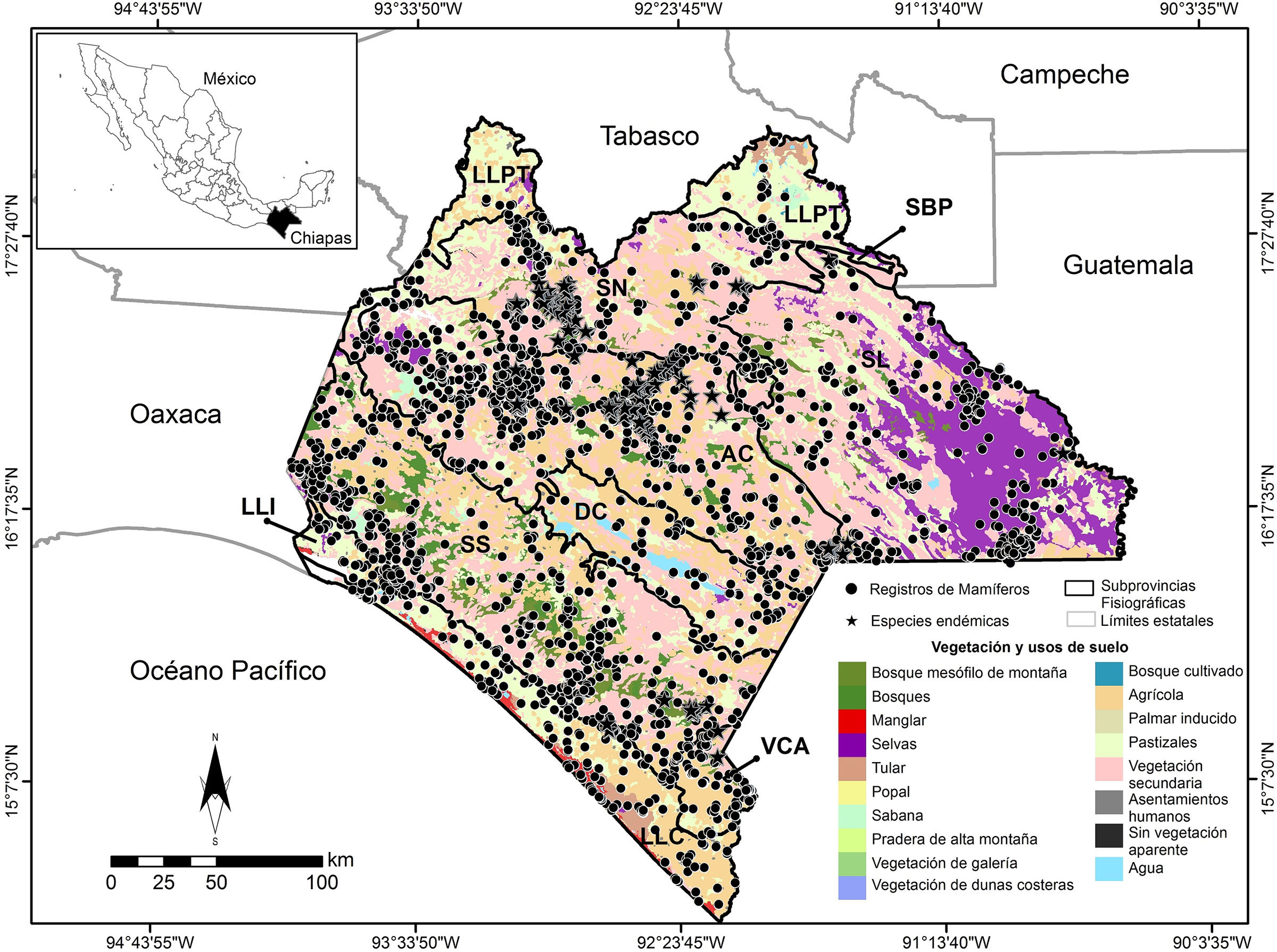
Figura 3
Localización de los mamíferos terrestres registrados en el estado de Chiapas, por subprovincia fisiográfica (ver nombres y acrónimos en la fig. 2 ) y uso de suelo y vegetación.AG: agrícola; AH: asentamientos humanos; BM: bosque mesófilo de montaña; BQ: bosques (pino, encino, cedro, oyamel y sus asociaciones); CA: cuerpos de agua; MG: manglar; PM: praderas de montaña; PZ: pastizales; SA: sabana; SE: selvas (altas, medianas y bajas, perennifolias, subperennifolias, caducifolias); SV: sin vegetación aparente; TU: tulares; VG: vegetación de galería; VS: vegetación secundaria.
El número de especies de mamíferos varió notoriamente en cada una de las subprovincias fisiográficas, desde 160 en AC hasta 15 en SBP ( tabla 3 ). El número de especies endémicas y el número de especies en alguna categoría de protección mostraron grandes diferencias entre subprovincias. En general, AC, SS y SN son las subprovincias que cuentan con la mayor cantidad de especies endémicas y de especies en alguna categoría de protección ( tabla 3).
El valor de diversidad beta de 1, calculado con el índice de Whittaker (B W ), indica una baja similitud de especies entre subprovincias fisiográficas. En el dendrograma de similitud se observa un grupo conformado por todas las subprovincias fisiográficas, excepto las Sierras Bajas del Petén. Las subprovincias más distanciadas fueron LLI, DC y LLPT, y las más similares entre sí son SS y AC, seguidas de SN y SL ( fig. 4).
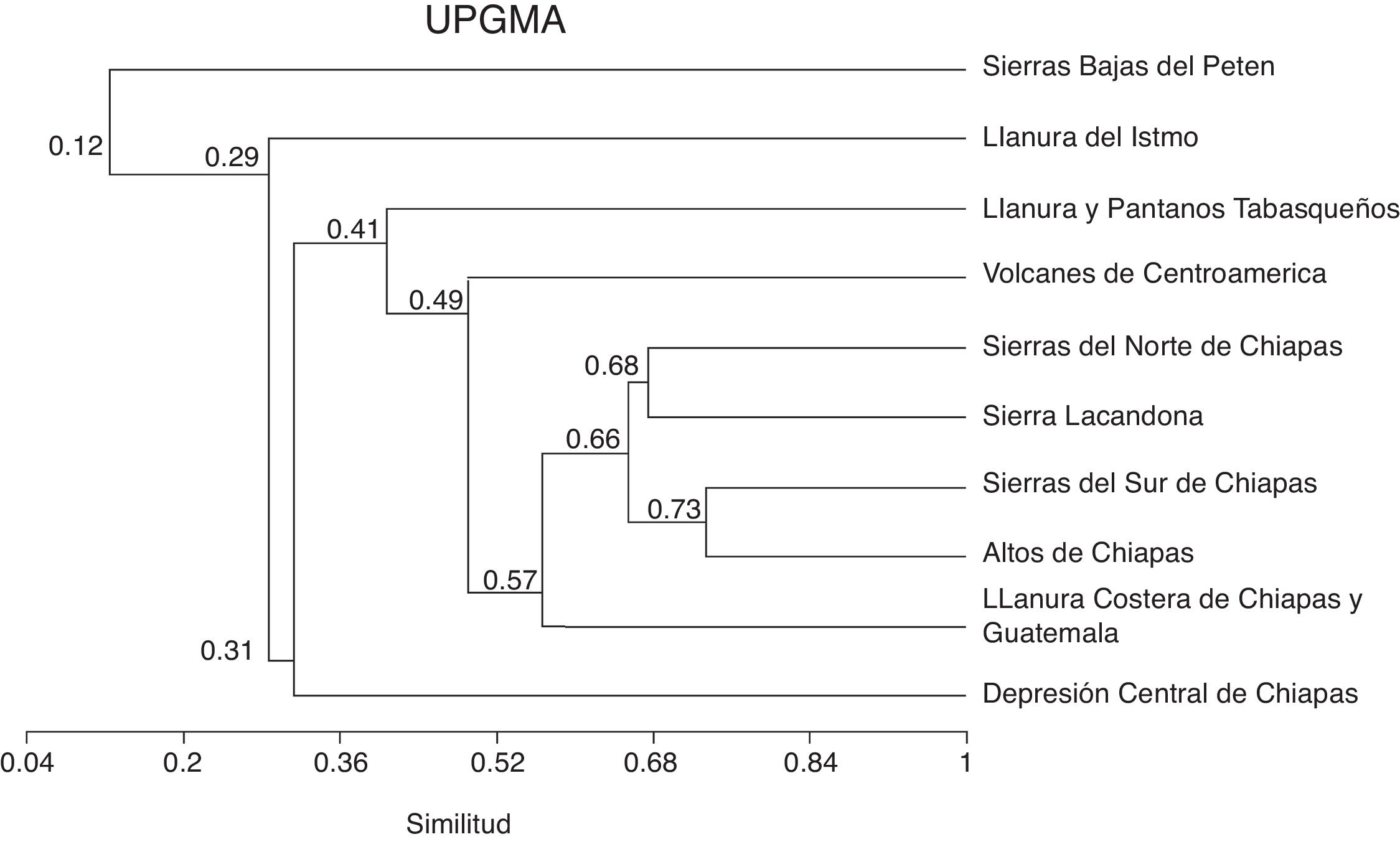
Figura 4
Dendrograma de similitud entre subprovincias fisiográficas de Chiapas, elaborado con un análisis de agrupamiento por el método UPGMA, mediante el coeficiente de Jaccard.
Discusión
Debido a la rica historia cultural en el aprovechamiento y manejo de recursos faunísticos (y florísticos) en Chiapas, así como por la heterogeneidad en climas, ambientes y fisiografía, dicho estado es ampliamente reconocido por su diversidad y riqueza mastofaunística. Las exploraciones biológicas nacionales y extranjeras en México (y Chiapas) desde el siglo XIX dieron cuenta de ello, y desde entonces despertó el interés primeramente de extranjeros por conocer este recurso, y posteriormente de los investigadores mexicanos ( Lorenzo et al., 2012 ), de tal forma que los ejemplares de mamíferos recolectados en el siglo xx (muchos de ellos tipos) se depositaron en colecciones extranjeras de diversas instituciones ( López-Wilchis, 2006; Ramírez-Pulido y González-Ruiz, 2006 ). En Chiapas, en particular, existe un aumento en el número de registros de mamíferos en la década de 1930, al igual que la descripción de nuevas especies de estos, la última en el 2014 ( Guevara et al., 2014 ), lo que lo ha llevado a ser el segundo estado mexicano con mayor riqueza de especies (210) de mamíferos silvestres.
El número de especies, géneros u otras categorías supraespecíficas de mamíferos del estado también se ha incrementado, en parte por los cambios taxonómicos en diversos taxones registrados en Chiapas, los cuales se mencionan a continuación. Dentro del orden Didelphimorphia se describe un género nuevo a partir de Marmosa canescens, nombrado Tlacuatzin (Voss y Jansa, 2003 ). En el orden Rodentia se analiza y recomienda como válido el cambio de la familia Cuniculidae y la del género Cuniculus como las categorías taxonómicas válidas para reemplazar Agoutidae y Agouti, respectivamente ( Comisión Internacional en la Nomenclatura Zoológica, 1998 ). Se agrupa a Sigmodon hispidus , una especie sin distribución en el estado y de la cual se derivan S. toltecus y S. zanjonensis ( Bradley, Henson y Durish, 2008; Carroll et al., 2005; Henson y Bradley, 2009 ). Se agrupa al género Liomys dentro del complejo Heteromys ( Anderson, Weksler y Rogers, 2006; Hafner et al., 2007 ). Se reconoce a Heteromys desmarestianus goldmani como especie, asignándole el nombre de H. goldmani ( Espinoza, Lorenzo y Rios, 2011; Hafner et al., 2007; Rogers y González, 2010 ). En el orden Carnivora se separa Spilogale angustifrons de S. putorius , siendo la primera la que se presenta en Chiapas ( Wozencraft, 2005 ). En el orden Artiodactyla cambia la sinonimia de Pecari tajacu por Dicotyles crassus y se restringe Mazama americana a Sudamérica, reconociendo a Mazama temama como la especie presente en Chiapas ( Ashley, Norman y Stross, 1996; Groves y Grubb, 2011; Grubb, 2005 ). Se propone el cambio del género Tapirus a Tapirella (Groves y Grubb, 2011). En el orden Primates, Alouatta pigra es un sinónimo de A. villosa , este último considerado como válido ( Brandon-Jones, 2006 ). En el orden Soricomorpha, Cryptotis goldmani cambia a C. griseoventris; la subespecie C. parvus tropicalis se asigna a nivel de especie como C. tropicalis (Carraway, 2007). Se agrupa a Sorex saussurei dentro de S. salvini (Woodman et al., 2012 ). Se describe una nueva especie de musaraña para Chiapas, C. lacandonensis (Guevara et al., 2014).
Para el orden Chiroptera, se asigna como especie a Eumops ferox con distribución en México y Eumops glaucinus se restringe a Sudamérica (McDonough et al., 2008). Eumops nanus, considerada como subespecie de Eumops bonariensis, se considera como especie (Eger, 2008). Se asigna a Natalus stramineus como Natalus mexicanus ( López-Wilchis et al., 2012; Tejedor, 2005 ). Se asigna el cambio de nombre de Leptonycteris curasoae a L. yerbabuenae ( Cole y Wilson, 2006a, b; Simmons, 2005; Simmons y Wetterer, 2002 ). Se registra a Trinycteris nicefori en el estado, la cual no estaba reportada para México ( Escobedo-Morales et al., 2006). Se trata a Mimon cozumelae como una especie diferente de M. bennetti , la cual incluso ya no existe para México ( Williams y Genoways, 2008). Se agrupa a Vampyrodes major como especie diferente de V. caraccioli (Velazco y Simmons, 2011). Se cambia la sinonimia de Sturnira ludovici por S. hondurensis (Gardner, 2008). Se usa el nombre de Sturnira parvidens , el cual había sido reconocido anteriormente como subespecie de Sturnira lilium (Velazco y Patterson, 2013). Se modifica el nombre de Uroderma bilobatum a Uroderma convexum (Mantilla-Meluk, 2014 ). Se describe una nueva especie, Rhogeessa bickhami, la cual es simpátrica con Rhogeessa genowaysi (Baird et al., 2012). Rhogeessa gracilis estaba considerada dentro de las especies con distribución estatal ( Naranjo et al., 2005, 2013, 2016; Retana-Guiascón y Lorenzo, 2002 ); sin embargo, durante la revisión de la base de registros y recolectas obtenidas no se encontró ninguna evidencia de esta especie para el estado, por lo que se procedió a eliminarla de la lista de especies presentes en Chiapas.
La diversidad beta muestra que existe una baja similitud (pocas especies compartidas) de especies entre las subprovincias fisiográficas de Chiapas; por lo tanto, existe una alta diversidad beta, lo cual refleja la heterogeneidad ecológica entre las diferentes subprovincias fisiográficas del estado ( Koleff et al., 2008 ). El mayor número de registros y la mayor riqueza de mamíferos se presentan en 2 grandes subprovincias, AC y SS; ambas representan el 52.6% del total de registros y el 33.3 y el 19.3%, respectivamente, del total de especies en el estado. AC y SS coinciden con las cadenas montañosas de la Cordillera Central (en la parte central del estado) y la Sierra Madre de Chiapas (paralela a la llanura costera del Pacífico), respectivamente, y presentan altitudes que van de los 1,000 a los 4,000 m. El hecho de que ambas subprovincias fisiográficas sean las más similares en número de especies (ver dendrograma de similitud) se debe a que presentan condiciones ambientales únicas que propician ecosistemas compuestos por bosques templados (bosques de encinos, bosques de pinos) y bosques mesófilos de montaña, así como una amplia variedad de comunidades vegetales como selvas (en SS) que albergan esa cantidad de especies, incluyendo endémicas ( P. zarhynchus, H. nelsoni, S. stizodon, T. bullaris y R. genowaysi ); sin embargo, estos bosques y selvas están muy perturbados debido a diversas actividades humanas.
Las subprovincias SN en la región de las Montañas del Norte del estado y SL, al este del estado, presentan también una buena proporción de registros con respecto al total (29.8%), y 14.8 y 15.0% del total de especies, respectivamente, y ambas se agrupan por la similitud en número de especies. SN es un terreno montañoso donde el clima es cálido húmedo todo el año y se presentan lluvias intensas que superan los 5,000 mm durante los nortes; en esta se localizan selvas y en las partes más altas, bosques de pino. En ella se encuentran especies endémicas, como P. zarhynchus, T. tumbalensis y S. sclateri . SL está formada por serranías de diferentes altitudes y, por su orientación, los vientos cargados de humedad (provenientes del golfo de México) penetran en su territorio y permiten la existencia de selvas, en las que se encuentran gran cantidad de especies de mamíferos tropicales y especies endémicas ( C. lacandonensis y P. zarhynchus ); en esta, es necesario llevar a cabo mayor exploración en ciertas áreas para contar con registros de mamíferos en zonas poco representadas.
Por el contrario, el menor número de registros y la menor riqueza de mamíferos se presentan en DC, y 3 más pequeñas, LLPT, LLI y SBP. Estas subprovincias representan el 3.49, el 2.25, el 0.98 y el 0.05% del total de registros, respectivamente. No tienen especies endémicas y presentan alturas que van de los de 500 a los 2,000 m; son diferentes del resto en número de especies, por lo que su posición en el dendrograma de similitud es externo, debido a que representan mayormente ecosistemas que no son adecuados para albergar gran cantidad de especies, como lo son sitios de vegetación secundaria, pastizales y áreas agrícolas.
Al analizar el mapa actual de las coberturas de vegetación, en el que se presenta cada especie en Chiapas, podemos observar el grado de perturbación que ha sufrido la vegetación original, lo que ha ocasionado una alteración en la distribución de las especies. Un ejemplo de lo anterior son los registros actuales del tapir Tapirella bairdii y el murciélago falso vampiro Vampyrum spectrum (ambas con altos requerimientos en la calidad del hábitat para sobrevivir), que actualmente están representadas en lugares con asentamientos humanos (AH), vegetación secundaria (VS) y pastizales (PZ), cuando originalmente se registraron en selva en 1945 la primera y en 1994 la segunda. Adicionalmente, existen registros de especies en tipos de vegetación que no concuerdan con su distribución actual, como es el caso del pecarí de labios blancos, Tayassu pecari , que tiene registros (en 1953) en bosques mesófilos en la Sierra Madre de Chiapas, cuando su distribución actual está restringida a la selva Lacandona. El mono araña, Ateles geoffroyii , es una especie sensible a la perturbación humana y cuenta con registros en 1944 en selva mediana cercanos a Tonalá, en la región de la Llanura Costera de Chiapas y Guatemala; dicha selva ha sido transformada en la actualidad por la presencia de asentamientos humanos. Caso similar ocurre con el jaguar, Panthera onca , con registros en la década de 1950 en Comitán y la Trinitaria, correspondientes a la subprovincia AC, hoy ocupada por grandes asentamientos humanos. Lo anterior son ejemplos de la contracción en el área de distribución de muchas especies ante el impacto de las actividades humanas en el estado.
La distribución de mamíferos en Chiapas se concentra en ecosistemas frágiles (bosques húmedos) en la porción norte de la entidad. La diversidad de mamíferos de las selvas bajas caducifolias prácticamente se encuentra sin protección en el estado, especialmente a lo largo de la franja fronteriza con Guatemala. Esta situación también se presenta en las Montañas del Norte, donde no existen áreas naturales protegidas, por lo que la aplicación de medidas que permitan proteger los bosques remanentes es prioritaria ( Naranjo et al., 2016).
Entre los mamíferos de Chiapas más vulnerables a la extinción se encuentran las especies endémicas, la mayoría incluidas en alguna categoría de riesgo dentro de la NOM-059-SEMARNAT-2010. En particular se requieren medidas inmediatas de conservación para estas, ya que también están incluidas en la Norma Oficial Mexicana y la lista roja de la IUCN, lo que implica que enfrentan actualmente un riesgo extremadamente alto de extinción en estado silvestre. Todas estas tienen distribuciones restringidas (< 100 km2), excepto P. zarhynchus , el cual presenta una distribución fragmentada, compuesta por 7 poblaciones conocidas dentro de la región de los Altos de Chiapas y Sierras del Norte de Chiapas ( tabla 3 ) en un área no mayor a los 20,000 km2 , de aquí que sea la especie con mayor número de registros de recolecta en 20 colecciones mastozoológicas (7 nacionales y 13 extranjeras). Es importante mencionar que desde 1898 (año en que se describe a P. zarhynchus ) se recolectaron 36 ejemplares, y tuvieron que pasar 55 años (1950) para tener nuevos registros de esta especie, con solo 4 ejemplares; a partir de entonces continúan sus recolectas, pero se desconoce si estas afectan negativamente la abundancia de sus poblaciones, en particular en localidades cuyos pobladores hacen uso de esta especie para su consumo, por ejemplo en Oxchuc ( Barragán, Retana-Guiascón y Naranjo, 2007 ).
De todas las especies endémicas, S. stizodon se encuentra en la Reserva Ecológica Huitepec, al igual que una población de P. zarhynchus ( Lorenzo, Álvarez-Castañeda, Pérez-Consuegra y Patton, 2016 ). Adicionalmente, C. lacandonensis se encuentra en el Monumento Natural Yaxchilán, por lo que su hábitat está protegido. El hábitat del resto de las especies endémicas se ha modificado por actividades humanas (agricultura, ganadería, tala, incendios forestales), en adición a los cambios climáticos que han afectado a aquellas que viven en las cimas de montañas (al grado de contar con un proceso de extinción a corto plazo), como H. nelsoni ( Rios, Lorenzo y Álvarez-Castañeda, 2016 ), por lo que su supervivencia se ha visto fuertemente afectada. Las diferencias entre R. bickhami y R. genowaysi se basan por cariotipos, por lo que es importante realizar estudios taxonómicos entre ellas ( Lorenzo, Briones-Salas y Álvarez-Castañeda, 2016 ).
Por otra parte, es importante hacer notar el caso de las musarañas: S. sclateri , descrita en 1897 por Merriam, que es conocida por 5 ejemplares (depositados en la Colección Mastozoológica de la Escuela Nacional de Ciencias Biológicas, IPN y en el NMNH) de 2 localidades, y S. stizodon , descrita en 1895 por Merriam, conocida solo por un ejemplar (NMNH) de la localidad tipo, Reserva Ecológica Huitepec, en San Cristóbal de Las Casas ( Carraway, 2007). Lo mismo ocurre con T. tumbalensis , descrita en 1901 por Merriam, la cual es conocida por menos de 10 ejemplares y solo de la localidad tipo, Tumbalá, y que no ha sido recolectada en los últimos 40 años ( Álvarez-Castañeda y Castro-Arellano, 2008 ).
A lo largo de casi 20 años los autores hemos realizado recolectas no intensivas para la búsqueda de estas especies, sin éxito de registrarlas; solamente al llevar a cabo búsquedas extensivas en campo podremos evaluar su estado de conservación, redefinir su categoría de riesgo y determinar si están extintas en vida silvestre; por el momento, continuamos considerando que estas especies existen dentro del estado. Las especies de musarañas están en una situación de conservación grave, debido a la transformación de los bosques de encinos maduros y su cambio a bosques homogéneos de pinos.
En una situación menos apremiante pero vulnerable se encuentran el armadillo de cola desnuda (Cabassous centralis), el leoncillo (Herpailurus yagouaroundi), el ocelote (Leopardus pardalis), el jaguar (Panthera onca), la nutria (Lontra longicaudis), el cacomixtle (Bassariscus sumichrasti) y el tapir (Tapirella bairdii) , las cuales se incluyen en la NOM-059-SEMARNAT-2010, la lista roja de la IUCN y el CITES. Estos y otros mamíferos regionales enfrentan una compleja variedad de amenazas para su supervivencia, por lo que las soluciones para evitar su extinción no son fáciles de identificar y aplicar. La conservación y buen manejo del hábitat remanente (protegido o no) y la creación y mantenimiento de corredores biológicos, a través de procesos de participación comunitaria, son estrategias deseables para favorecer a la mayoría de las especies ( Naranjo et al., 2013, 2016).
La acelerada transformación de los ecosistemas en Chiapas (con tasas anuales de deforestación superiores al 3%; Soto-Pinto, Castillo-Santiago y Jiménez-Ferrer, 2012 ) está ocasionando graves consecuencias en la estructura y composición de las comunidades de mamíferos, así como en la distribución y la abundancia de muchas especies, en particular en aquellas con mayor vulnerabilidad debido a su distribución restringida y a sus bajas densidades. La pérdida y fragmentación de los ecosistemas naturales en el estado facilita el desplazamiento de especies nativas por mamíferos oportunistas (i.e., roedores domésticos, gatos y perros) que, además de competir por los mismos recursos de espacio y alimento, son depredadores de la fauna nativa y portadores potenciales de enfermedades tanto para esta como para los humanos ( Naranjo et al., 2016).
La investigación sobre los mamíferos de Chiapas debe enfocarse, en la medida de lo posible, a evaluar ciertas áreas poco exploradas (principalmente por su difícil acceso) para ampliar el conocimiento de la distribución de los mamíferos, en particular en las subprovincias fisiográficas Sierra Lacandona, Sierras del Sur de Chiapas, Sierras del Norte de Chiapas, Discontinuidad Depresión Central de Chiapas y Llanura y Pantanos Tabasqueños. Se deben hacer revisiones detalladas de ejemplares en colecciones científicas, hacer trabajo de campo y contar con nuevos datos no solo de las especies en sí, sino también del hábitat en el que se encuentran. Los diversos factores que motivan el proceso de cambio en el estado de Chiapas requieren conocer con mayor precisión lo que ocurre con las especies tanto histórica como actualmente. Hay especies que están perdiendo el hábitat y podrían desaparecer en los siguientes años. También se deben realizar monitoreos intensivos para buscar las especies endémicas (la mayoría se encuentran únicamente en colecciones extranjeras) y generar el conocimiento biológico de las mismas. Es de igual importancia evaluar los cambios en los procesos ecológicos y evolutivos de las especies silvestres, resultantes de actividades humanas tales como la transformación del hábitat, la extracción de especies ( Naranjo et al., 2009 ) y los efectos del cambio climático. En particular, es urgente aplicar medidas de protección del hábitat (bosques tropicales y de montaña, humedales y manglares), así como impulsar alternativas realistas de uso sustentable de la flora y fauna silvestres a través de la conformación de grupos organizados capaces de crear unidades de manejo para la conservación de la vida silvestre (UMA) bien administradas, asesoradas y financiadas que permitan mejorar la economía de las comunidades rurales para hacerlas partícipes de este proceso. Si no se llevan a cabo estas acciones, difícilmente podremos avanzar en generar conocimiento científico biológico actual de los mamíferos de Chiapas y, en particular, del estado de conservación que guardan las poblaciones. De no ponerse en práctica una estrategia viable de conservación, la pérdida de la diversidad de mamíferos en Chiapas tendría costos ecológicos y económicos muy elevados.
Agradecimientos
A ECOSUR por el apoyo brindado a lo largo de dos décadas de estudio en Chiapas. A dos revisores anónimos por aportar sugerencias y comentarios valiosos.
Apéndice 1
Lista taxonómica de los mamíferos terrestres de Chiapas.
Subprovincia fisiográfica: AC: Altos de Chiapas; DC: Depresión Central de Chiapas; LLC: Llanura Costera de Chiapas y Guatemala; LLI: Llanura del Istmo; LLPT: Llanura y Pantanos Tabasqueños; SBP: Sierras Bajas del Peten; SL: Sierra Lacandona; SN: Sierras del Norte de Chiapas; SS: Sierras del Sur de Chiapas; VCA: Volcanes de Centroamérica.
Vegetación: AG: agrícola; AH: asentamientos humanos; BM: bosque mesófilo de montaña; BQ: bosques (pino, encino, cedro, oyamel y sus asociaciones); CA: cuerpos de agua; MG: manglar; PM: praderas de montaña; PZ: pastizales; SA: sabana; SE: selvas (altas, medianas y bajas, perennifolias, subperennifolias, caducifolias); SV: sin vegetación aparente; TU: tulares; VG: vegetación de galería; VS: vegetación secundaria.
Altitud (metros): 1: 0-300; 2: 301-500; 3: 501-1,000; 4: 1,001-1,500; 5: 1,501-2,000; 6: 2,001-2,500; 7: 2,501-3,000; 8: 3,001-3,500; 9: 3,501-4,000.
Estado de conservación: CITES: I, II, III. IUCN: CR: en peligro crítico; DD: datos insuficientes; EN: en peligro; LC: menor preocupación; NT: casi amenazada; VU: vulnerable.
NOM (NOM-059-SEMARNAT-2010): A: amenazada; P: en peligro de extinción; Pr: sujeta a protección especial.
Endemismo: CHIS: endémico de Chiapas; MX: endémico de México.
| Categoría taxonómica | Subprovincia fisiográfica | Vegetación | Altitud | NOM 059 | IUCN | CITES |
| Orden Didelphimorphia | ||||||
| Familia Didelphidae | ||||||
| Subfamilia Caluromyinae | ||||||
| Caluromys derbianus (Waterhouse, 1841) | LLC, AC, SL, SN, SS | AG, PZ, VS, AH | 1, 3 | A | LC | |
| Subfamilia Didelphinae | ||||||
| Chironectes minimus (Zimmermann, 1780) | LLPT, SL, SN | AG, PZ, SE, VS | 1, 4, 6 | P | LC | |
| Didelphis marsupialis Linnaeus, 1758 | DC, LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, PZ, SE, SV, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Didelphis virginiana Kerr, 1792 | DC, LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, SV, PZ, SE, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Philander opossum (Linnaeus, 1758) | LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, MG, PZ, SE, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Marmosa mexicana Merriam, 1897 | LLC, AC, SL, SN, SS, VCA | AG, AH, BM, BQ, PZ, PM, VS | 1, 3, 4, 5, 6, 8, 9 | LC | ||
| Tlacuatzin canescens (J. A. Allen, 1893) MX | AC, SS | BM, VS, AH | 3, 5 | LC | ||
| Metachirus nudicaudatus (É. Geoffroy St.-Hilaire, 1803) | SL | AG | 1 | A | LC | |
| Orden Cingulata | ||||||
| Familia Dasypodidae | ||||||
| Subfamilia Dasypodinae | ||||||
| Dasypus novemcinctus Linnaeus, 1758 | LLC, AC, SL, SN, SS | AG, BM, CA, MG, PZ, SE, SV, VS, AH | 1, 2, 3, 4, 5 | LC | ||
| Subfamilia Tolypeutinae | ||||||
| Cabassous centralis (Miller, 1899) | SL, SN | PZ, SE, VS | 1, 2, 3, 4 | P | DD | III |
| Orden Pilosa | ||||||
| Familia Myrmecophagidae | ||||||
| Tamandua mexicana (de Saussure, 1860) | DC, LLC, AC, SL, SS, VCA | AG, AH, BM, SV, PZ, VS | 1, 2, 3, 4, 5 | P | LC | III |
| Familia Cyclopedidae | ||||||
| Cyclopes didactylus (Linnaeus, 1758) | LLPT, SL, SN, SS | PZ, VS, AH | 1, 2, 3 | P | LC | |
| Orden Soricomorpha | ||||||
| Familia Soricidae | ||||||
| Subfamilia Soricinae | ||||||
| Cryptotis goodwini Jackson, 1933 | SS, VCA | AG, VS | 4, 5, 6 | LC | ||
| Cryptotis griseoventris Jackson, 1933 MX | AC | AG, BQ, VS, AH | 3, 4, 5, 6, 7 | VU | ||
| Cryptotis lacandonensis Guevara et al., 2014 CHIS | SL | SE | 1 | |||
| Cryptotis merriami Choate, 1970 | AC, SL | AG, VS | 4, 5 | LC | ||
| Cryptotis mexicanus (Coues, 1877) MX | AC, SN | PZ, VS | 3, 4, 5, 6 | LC | ||
| Cryptotis parvus (Say, 1822) | DC, LLC, AC, SN, SS, VCA | AG, AH, BQ, SV, PZ, PM, VS | 1, 3, 4, 5, 6, 9 | LC | ||
| Cryptotis tropicalis (Merriam, 1895) | LLPT, SS, VCA | AG, AH | 1, 4 | Pr | DD | |
| Sorex salvini Merriam, 1892 MX | AC, SN, SS, VCA | AG, BM, PZ, VS, AH | 4, 5, 6, 7 | LC | ||
| Sorex sclateri Merriam, 1897 CHIS | SN | AG | 4 | A | CR | |
| Sorex stizodon Merriam, 1895 CHIS | AC | AH | 6 | A | CR | |
| Sorex veraepacis Alston, 1877 | AC, SN, SS, VCA | BM, VS, AH | 4, 5, 6, 7 | A | LC | |
| Orden Chiroptera | ||||||
| Familia Emballonuridae | ||||||
| Subfamilia Emballonurinae | ||||||
| Balantiopteryx io Thomas, 1904 | AC, LLPT, SL, SN | CA, PZ, SE, VS | 1, 2, 3, 4 | VU | ||
| Balantiopteryx plicata Peters, 1867 | DC, LLC, LLI, AC, LLPT, SN, SS | AG, AH, BM, BQ, MG, PZ, VS | 1, 2, 3, 4, 5 | LC | ||
| Centronycteris centralis Thomas, 1912 | LLPT, SS | BQ, AH | 1, 4 | Pr | LC | |
| Diclidurus albus Wied-Neuwied, 1820 | SL | PZ | 1 | LC | ||
| Peropteryx kappleri Peters, 1867 | LLPT, SL, SN | AH, PZ, SE, VS | 1, 2 | Pr | LC | |
| Peropteryx macrotis (J. A.Wagner, 1843) | AC, LLPT, SL, SN, SS | AG, CA, PZ, SE, VS, AH | 1, 2, 3, 5 | LC | ||
| Rhynchonycteris naso (Wied-Neuwied, 1820) | LLC, SL, SBP, SN, SS, VCA | AG, AH, CA, MG, PZ, SE, TU, VS | 1, 2, 3 | Pr | LC | |
| Saccopteryx bilineata (Temminck, 1838) | LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, CA, MG, PZ, SE, VS | 1, 2, 3, 4, 5 | LC | ||
| Saccopteryx leptura (Schreber, 1774) | SS | VS | 3 | LC | ||
| Familia Molossidae | ||||||
| Subfamilia Molossinae | ||||||
| Cynomops mexicanus (J. K. Jones y Genoways, 1967) | SL | SE | 1 | Pr | LC | |
| Eumops auripendulus (Shaw, 1800) | SS | PZ, VS | 1 | LC | ||
| Eumops ferox (Gundlach, 1861) | LLC, LLPT, SS | PZ, VS, AH | 1, 3 | |||
| Eumops hansae Sanborn, 1932 | SL, SS | PZ | 1, 2 | LC | ||
| Eumops nanus (Miller,1900) | SS | VS | 2, 3 | LC | ||
| Eumops underwoodi Goodwin, 1940 | LLC, LLI, AC, SS | AG, PZ, VS, AH | 1, 2, 3 | LC | ||
| Molossus aztecus de Saussure, 1860 | AC, SS | BQ, PZ, VS | 2, 3, 6 | LC | ||
| Molossus coibensis J. A. Allen, 1904 | LLC | PZ | 1 | LC | ||
| Molossus molossus (Pallas, 1766) | DC, LLC, AC, LLPT, SL, SBP, SS, VCA | AG, AH, BQ, MG, PZ, SE, VS | 1, 3, 5 | LC | ||
| Molossus rufus É. Geoffroy St.-Hilaire, 1805 | DC, LLC, LLI, AC, LLPT, SL, SBP, SN, SS, VCA | AG, AH, MG, PZ, SE, VS | 1, 2, 3, 5 | LC | ||
| Molossus sinaloae J. A. Allen, 1906 | LLC, SS | AH, VS | 1, 2 | LC | ||
| Nyctinomops laticaudatus (É. Geoffroy Saint – Hilaire, 1805) | LLC, SN, SS | PZ, VS, AH | 1, 3, 4 | LC | ||
| Nyctinomops macrotis (Gray, 1839) | SN | PZ, VS | 3, 4 | LC | ||
| Promops centralis Thomas, 1915 | AC | BQ | 6 | LC | ||
| Tadarida brasiliensis (I. Geoffroy Saint -Hilaire, 1824) | DC, AC, SN, SS | AG, BM, BQ, PZ, VS, AH | 3, 4, 5, 6 | LC | ||
| Familia Natalidae | ||||||
| Natalus mexicanus Miller, 1902 | AC, SL, SS | AG, BM, BQ, PZ, SE, VS, AH | 1, 2, 3, 4, 5 | LC | ||
| Familia Thyropteridae | ||||||
| Thyroptera tricolor Spix, 1823 | LLC, LLPT, SL, SN | AG, AH, PZ, SE, VS | 1 | Pr | LC | |
| Familia Mormoopidae | ||||||
| Mormoops megalophylla (Peters, 1864) | LLC, AC, LLPT, SL, SBP, SN, SS | AG, AH, BM, BQ, CA, SV, PZ, SE, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Pteronotus davyi Gray, 1838 | LLC, AC, LLPT, SL, SBP, SN, SS | AG, BM, BQ, CA, PZ, SE, VS, AH | 1, 2, 3, 4, 5 | LC | ||
| Pteronotus gymnonotus (J. A. Wagner, 1843) | SL | CA | 1 | A | LC | |
| Pteronotus parnellii (Gray, 1843) | DC, LLC, LLI, AC, LLPT, SL, SBP, SN, SS, VCA | AG, BM, BQ, CA, SV, PZ, SA, SE, VS AH | 1, 2, 3, 4, 5, 6, 7 | LC | ||
| Pteronotus personatus (J. A. Wagner, 1843) | LLC, LLI, AC, LLPT, SL, SS | AG, MG, PZ, SE, SV, VS, AH | 1, 2, 3, 4 | LC | ||
| Familia Noctilionidae | ||||||
| Noctilio albiventris Desmarest, 1818 | LLC | PZ | 1 | Pr | LC | |
| Noctilio leporinus (Linnaeus, 1758) | LLC, LLI, SL, SN, SS | AG, AH, BQ, CA, MG, PZ, SE, TU, VS | 1, 2, 3, 4, 5 | LC | ||
| Familia Phyllostomidae | ||||||
| Subfamilia Carollinae | ||||||
| Carollia perspicillata (Linnaeus, 1758) | LLC, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, MG, PZ, SE, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Carollia sowelli R. J. Baker et al., 2002 | LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, SV, MG, PZ, SA, SE, VS | 1, 2, 3, 4, 5 | LC | ||
| Carollia subrufa (Hahn, 1905) | DC, LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, SV, MG, PZ, PM, SA, SE, TU, VS | 1, 2, 3, 4, 5, 6, 8 | LC | ||
| Subfamilia Desmodontinae | ||||||
| Desmodus rotundus (É. Geoffroy Saint.-Hilaire, 1810) | DC, LLC, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, SV, MG, PZ, PM, SA, SE, VS | 1, 2, 3, 4, 5, 6, 8 | LC | ||
| Diaemus youngi (Jentink, 1893) | SL, SN | PZ, VS, AH | 1, 3 | Pr | LC | |
| Diphylla ecaudata Spix, 1823 | LLC, AC, SL, SN, SS, VCA | AG, AH, BQ, PZ, SE, VS | 1, 2, 3, 4, 5 | LC | ||
| Subfamilia Glossophaginae | ||||||
| Anoura geoffroyi Gray, 1838 | DC, AC, SL, SN, SS, VCA | AG, AH, BM, BQ, SV, PZ, PM, SE, VS | 1, 2, 3, 4, 5 6, 7, 8 | LC | ||
| Choeroniscus godmani (Thomas, 1903) | LLC, AC, LLPT, SL, SN, SS, VCA | AG, MG, PZ, SE, VS, AH | 1, 3, 4, 5, 6 | LC | ||
| Choeronycteris mexicana Tschudi, 1844 | AC | PZ, SV, VS | 2, 3, 4 | A | NT | |
| Hylonycteris underwoodi Thomas, 1903 | AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, PZ, PM, SE, VS | 1, 2, 3, 4, 5, 8 | LC | ||
| Lichonycteris obscura Thomas, 1895 | SL, SN | PZ, SE | 1 | LC | ||
| Glossophaga commissarisi Gardner, 1962 | DC, LLC, AC, SL, SN, SS, VCA | AG, AH, BM, BQ, MG, PZ, SA, SE, TU, VS | 1, 2, 3, 4, 5 6 | LC | ||
| Glossophaga leachii (Gray, 1844) | DC, LLC, LLI, AC, LLPT, SL, SBP, SN, SS, VCA | AG, AH, BM, MG, PZ, SE, VS | 1, 2, 3, 4, 5 | LC | ||
| Glossophaga morenoi Martínez y Villa, 1938 MX | DC, LLC, AC, SL, SN, SS, VCA | AG, BM, BQ, MG, PZ, SE, SV, VS, AH | 1, 2, 3, 4, 5, 6 | LC | ||
| Glossophaga soricina (Pallas, 1766) | DC, LLC, LLI, AC, LLPT, SL, SBP, SN, SS, VCA | AG, AH, BM, BQ, CA, SV, MG, PZ, SA, SE, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Leptonycteris yerbabuenae Martí nez y Villa, 1940 | LLC, LLI, AC, SN, SS | AG, AH, BM, BQ, PZ, VS | 1, 3, 4 | A | VU | |
| Leptonycterys nivalis (de Saussure, 1860) | LLC, LLI | PZ, AH | 1 | A | EN | |
| Subfamilia Glyphonycterinae | ||||||
| Glyphonycteris sylvestris Thomas, 1896 | LLC, AC, SL | MG, SE, VS | 1, 3 | LC | ||
| Subfamilia Lonchorhininae | ||||||
| Lonchorhina aurita Tomes, 1863 | LLC, LLI, AC, SL, SN, SS | AG, CA, PZ, SE, VS | 1, 2, 3, 4 | A | LC | |
| Subfamilia Macrotinae | ||||||
| Macrotus waterhousii Gray, 1843 | LLPT | AH | 1 | LC | ||
| Subfamilia Micronycterinae | ||||||
| Lampronycteris brachyotis (Dobson, 1879) | AC, SL | PZ, SE, VS | 1, 3 | A | LC | |
| Micronycteris microtis Miller, 1898 | DC, LLC, AC, LLPT, SL, SN, SS | AG, AH, BM, SV, MG, PZ, SE, VS | 1, 2, 3, 4, 5 6 | LC | ||
| Micronycteris schmidtorum Sanborn, 1935 | LLC, AC, SL | AG, CA, MG, VS | 1, 3, 4 | A | LC | |
| Trinycteris nicefori (Sanborn, 1949) | SL | SE | 1 | LC | ||
| Subfamilia Phyllostominae | ||||||
| Macrophyllum macrophyllum (Schinz, 1821) | SL | PZ, SE | 1, 3 | A | LC | |
| Trachops cirrhosus (Spix, 1823) | LLC, AC, SL, SN, SS, VCA | AG, BQ, MG, PZ, SE, VS | 1, 2, 3, 4, 5 | A | LC | |
| Lophostoma brasiliense Peters, 1867 | SL | SE | 1, 3 | A | LC | |
| Lophostoma evotis (W. B. Davis y Carter, 1978) | LLC, SL, SS | MG, PZ, SE, VS | 1 | A | LC | |
| Mimon cozumelae Goldman, 1914 | AC, LLPT, SL, SN | CA, PZ, SE, VS, AH | 1, 2, 3 | A | LC | |
| Mimon crenulatum (É. Geoffroy Saint- Hilaire, 1803) | AC, SL, SN | AG, AH, PZ, SE, VS | 1, 3, 4 | A | LC | |
| Phyllloderma stenops Peters, 1865 | SL | PZ, SE | 1 | A | LC | |
| Phylllostomus discolor (J. A. Wagner, 1843) | LLC, AC, LLPT, SL, SN, SS, VCA | AG, BM, BQ, MG, PZ, SA, SE, VS, AH | 1, 2, 3, 4 | LC | ||
| Tonatia saurophila Koopman y Williams, 1951 | SL | AH, CA, PZ, SE, VS | 1, 2 | A | LC | |
| Chrotopterus auritus (Peters, 1856) | AC, SL, SN, SS | AG, BM, PZ, SE, VS | 1, 2, 3, 4, 5 6 | A | LC | |
| Vampyrum spectrum (Linnaeus, 1758) | SL | PZ, SE, VS | 1 | P | NT | |
| Subfamilia Stenodermatinae | ||||||
| Artibeus jamaicensis Leach, 1821 | DC, LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, SV, MG, PZ, SA, SE, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Artibeus lituratus (Olfers, 1818) | DC, LLC, LLI, AC, LLPT, SL, SBP, SN, SS, VCA | AG, AH, BM, BQ, CA, SV, MG, PZ, SE, TU, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Dermanura azteca (Andersen, 1906) | AC, SL, SN, SS, VCA | AG, AH, BM, PZ, VS | 3, 4, 5, 6, 7 | LC | ||
| Dermanura phaeotis Miller, 1902 | LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, BM, PZ, SE, VS, AH | 1, 2, 3, 4, 5 | LC | ||
| Dermanura tolteca (de Saussure, 1860) | DC, LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, SV, PZ, PM, SA, SE, VS | 1, 2, 3, 4, 5, 6, 8 | LC | ||
| Dermanura watsoni (Thomas, 1901) | AC, LLPT, SL, SN, SS | AH, PZ, SE, VS | 1, 2, 3, 4, 6 | Pr | LC | |
| Enchisthenes hartii (Thomas, 1892) | LLC, LLI, AC, SL, SN, SS, VCA | AG, BM, PZ, SA, SE, TU, VS, AH | 1, 2, 3, 4, 5, 6, 8 | Pr | LC | |
| Centurio senex Gray, 1842 | DC, LLC, AC, LLPT, SL, SN, SS, VCA | AG, BM, BQ, SV, MG, PZ, SA, SE, VS AH | 1, 2, 3, 4, 5, 6, 7 | LC | ||
| Chiroderma salvini Dobson, 1878 | LLC, AC, SL, SN, SS, VCA | AG, BM, MG, PZ, SE, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Chiroderma villosum Peters, 1860 | DC, LLC, AC, SL, SN, SS, VCA | AG, BM, BQ, MG, PZ, SA, SE, VS, AH | 1, 2, 3, 4, 5 | LC | ||
| Platyrrhinus helleri (Peters, 1866) | DC, LLC, LLI, AC, LLPT, SL, SBP, SN SS, VCA | AG, AH, BM, BQ, CA, PZ, SA, SE, VS | 1, 2, 3, 4, 5, 6, 7 | LC | ||
| Uroderma convexum Peters, 1866 | LLC, LLI, AC, LLPT, SL, SBP, SN, SS, VCA | AG, AH, BM, BQ, CA, SV, MG, PZ, SE, TU, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Uroderma magnirostrum W. B. Davis, 1968 | LLC, SS, VCA | AG, PZ, VS | 1, 2 | LC | ||
| Vampyressa thyone Thomas, 1909 | LLC, AC, LLPT, SL, SBP, SN, SS | AG, CA, MG, PZ, SE, VS, AH | 1, 2, 3, 4, 5 | LC | ||
| Vampyrodes major G. M. Allen, 1908 | LLC, AC, LLPT, SL, SN, SS | AG, BQ, PZ, SE, VS, AH | 1, 2, 3, 4, 5 | LC | ||
| Sturnira hondurensis Goodwin, 1940 | LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, PZ, PM, SA, SE, SV, VS | 1, 2, 3, 4, 5, 6, 7, 8 | LC | ||
| Sturnira parvidens Goldman, 1917 | DC, LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, MG, PZ, SA, SE, SV, TU, VS | 1, 2, 3, 4, 5, 6, 7 | LC | ||
| Familia Vespertilionidae | ||||||
| Sufamilia Antrozoinae | ||||||
| Bauerus dubiaquercus (Van Gelder, 1959) | LLC, AC, SL | PZ, SE, VS | 1, 3 | NT | ||
| Subfamilia Myotiinae | ||||||
| Myotis albescens (É. Geoffroy Saint.-Hilaire, 1806) | AC, SL, SN, SS, VCA | AG, AH, BM, BQ, PZ, SE, VS | 1, 2, 3, 4, 5, 6 | Pr | LC | |
| Myotis californicus (Audubon y Bachman, 1842) | AC, VCA | AG, VS | 4, 5, 6 | LC | ||
| Myotis elegans Hall, 1962 | LLC, AC, SL, SS, VCA | AG, BM, PZ, VS, AH | 1, 2, 3, 4, 5, 6 | LC | ||
| Myotis fortidens Miller y G. M. Allen, 1928 | DC, LLC, AC, LLPT, SL, SN, SS | AG, AH, BM, PZ, TU | 1, 3, 5 | LC | ||
| Myotis keaysi J. A. Allen, 1914 | AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, PZ, PM, SA, SE, VS | 1, 2, 3, 4, 5, 6, 8 | LC | ||
| Myotis nigricans (Schinz, 1821) | LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, BM, BQ, SV, MG, PZ, TU, VS, AH | 1, 2, 3, 4, 5, 6 | Pr | LC | |
| Myotis thysanodes Miller, 1897 | AC | AG, BQ | 6 | LC | ||
| Myotis velifer (J. A. Allen, 1890) | AC, SS | AG, AH, BQ, VS | 3, 5, 6 | LC | ||
| Subfamilia Vespertilioninae | ||||||
| Perimyotis subflavus (F. Cuvier, 1832) | LLPT, SL, SN | AH, PZ, SE, VS | 1, 2 | LC | ||
| Eptesicus brasiliensis (Desmarest, 1819) | LLC, AC, LLPT, SL, SN, SS | BM, PZ, SE, VS, AH | 1, 2, 3, 4, 5 6 | LC | ||
| Eptesicus furinalis (d’Orbigny y Gervais, 1847) | LLC, LLI, AC, LLPT, SL, SBP, SN, SS, VCA | AG, BM, PZ, SE, VS, AH | 1, 2, 3, 5, 6 | LC | ||
| Eptesicus fuscus (Palisot de Beauvois, 1796) | AC, SL; SN, SS | AG, AH, BM, BQ, SE, VS | 1, 3, 4, 5, 6 7 | LC | ||
| Lasiurus blossevillii (Lesson, 1826) | LLC, AC, SL, SS | AG, AH, BM, BQ, PZ, SE | 1, 4, 5, 6 | LC | ||
| Lasiurus cinereus (Palisot de Beauvois, 1796) | LLC, AC, SL, SS | AG, BM, PZ, SE, VS, AH | 1, 3, 4, 6 | LC | ||
| Lasiurus ega (Gervais, 1856) | LLC, LLI, AC, SL, SN, SS | AG, AH, BM, PZ, SE, VS | 1, 2, 3, 4, 6 | LC | ||
| Lasiurus intermedius H. Allen, 1862 | LLC, AC, SL, SN, SS | AH, BQ, PZ, VS | 1, 3, 4, 6 | LC | ||
| Rhogeessa bickhami Baird, Marchán-Rivadeneira, Pérez, y R. J. Baker, 2012 CHIS | SS | BM | 5 | |||
| Rhogeessa genowaysi R. J. Baker, 1984 CHIS | LLC, SS | AG, BM, PZ | A | EN | ||
| Rhogeessa parvula H. Allen, 1866 MX | LLC, AC, SS, VCA | PZ, VS, AH | 1, 3 | LC | ||
| Rhogeessa tumida H. Allen, 1866 | DC, LLC, AC, SL, SS, VCA | AG, AH, BM, BQ, PZ, SA, SE, TU, VS | 1, 2, 3, 4, 5 | LC | ||
| Corynorhinus mexicanus (G. M. Allen, 1916) | AC | VS | 3 | NT | ||
| Orden Primates | ||||||
| Familia Atelidae | ||||||
| Subfamilia Atelinae | ||||||
| Ateles geoffroyi Kuhl, 1820 | LLC, AC, LLPT, SL, SN, SS | AG, AH, BQ, CA, PZ, SE, SV, TU, VS | 1, 2, 3, 4, 5 | P | EN | II |
| Subfamilia Alouattinae | ||||||
| Alouatta palliata (Gray, 1849) | AC, SL, SN, SS | AG, PZ, VS, AH | 1, 3, 4 | P | LC | I |
| Allouatta villosa (Gray, 1845) | AC, LLPT, SL, SN, SS | CA, PZ, SE, VS, AH | 1, 2, 3, 4, 5 | P | EN | I |
| Orden Lagomorpha | ||||||
| Familia Leporidae | ||||||
| Subfamilia Leporinae | ||||||
| Sylvilagus brasiliensis (Linnaeus, 1758) | LLC, AC, SL, SN, VCA | AG, AH, CA, PZ, SE, VS | 1, 2, 3 | LC | ||
| Sylvilagus floridanus (J. A. Allen, 1890) | DC, LLC, LLI, AC, SL, SN, SS, VCA | AG, AH, BQ, PZ, SE, VS | 1, 2, 3, 4, 5, 6, 7 | LC | ||
| Orden Rodentia | ||||||
| Familia Sciuridae | ||||||
| Subfamilia Pteromyinae | ||||||
| Glaucomys volans (Linnaeus, 1758) | AC, SL, SN | AG, AH BQ, PZ, VS | 3, 4, 5, 6, 7 | A | LC | |
| Sciurus aureogaster F. Cuvier, 1829 | DC, LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH BM, BQ CA, PZ, PM, SA, SE, SV, VS | 1, 3, 4, 5, 6 7, 8 | LC | ||
| Sciurus deppei Peters, 1864 | LLC, AC, LLPT, SL, SN SS, VCA | AG, AH BM, CA MG, PZ, PM, SE, VS | 1, 3, 4, 5, 6, 7, 8 | LC | ||
| Sciurus variegatoides Ogilby, 1839 | LLC, SN, SS, VCA | AG, AH MG, PZ, TU, VS | 1, 2, 3, 5 | Pr | LC | |
| Sciurus yucatanensis J. A. Allen, 1877 | AC, SL | AH, PZ, SA, VS | 1, 3, 4 | LC | ||
| Familia Geomyidae | ||||||
| Subfamilia Geomyinae | ||||||
| Orthogeomys grandis (Thomas, 1893) | LLC, LLI, AC, LLPT, SS VCA | AG, AH BM, BQ, PZ, SV, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Orthogeomys hispidus (J. L. Le Conte, 1852) | DC, AC, LLPT, SL, SN, SS | AG, AH BM, BQ CA, PZ, SE, VS | 1, 2, 3, 4, 5, 6, 7 | LC | ||
| Familia Heteromyidae | ||||||
| Subfamilia Heteromyinae | ||||||
| Heteromys desmarestianus Gray, 1868 | LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH BM, BQ CA, PZ, PM, SE, VS | 1, 2, 3, 4, 5, 6, 8 | LC | ||
| Heteromys goldmani Merriam, 1902 | LLC, SN, SS, VCA | AG, AH, BM, BQ, PZ, SE, VS | 1, 2, 3, 4, 5, 6, 7 | LC | ||
| Heteromys nelsoni Merriam, 1902 CHIS | SS, VCA | AG, BM, VS | 5, 7, 8 | Pr | EN | |
| Heteromys pictus Thomas, 1893 | DC, LLC, LLI, AC, SL, SN SS | AG, AH BM, BQ, CA, SV, PZ, SA, SE, VG, VS | 1, 2, 3, 4, 5 | LC | ||
| Heteromys salvini Thomas, 1893 | LLC, LLI, SS, VCA | AG, AH BM, MG, PZ, SE, VS | 1, 2, 3, 4, 5 | LC | ||
| Familia Erenthizontidae | ||||||
| Subfamilia Erethizontinae | ||||||
| Coendou mexicanus (Kerr, 1792) | LLC, AC, SL, SS | AG, AH BQ, CA, MG, PZ, SA, SE, VS | 1, 2, 3, 5 | A | LC | III |
| Familia Agoutidae | ||||||
| Subfamilia Dasyproctinae | ||||||
| Dasyprocta mexicana de Saussure, 1860 MX | AC, SS | SE, VS, AH | 3, 5 | CR | ||
| Dasyprocta punctata Gray, 1842 | LLC, AC, LLPT, SL, SS, VCA | AG, AH, CA, MG, PZ, SE, VS | 1, 2, 3 | LC | III | |
| Familia Cuniculidae | ||||||
| Cuniculus paca (Linnaeus, 1766) | LLC, AC, LLPT, SL, SN, SS | AG, BM, BQ, CA MG, PZ, SE, SV, VS, AH | 1, 2, 3, 4, 5, 6 | LC | III | |
| Familia Cricetidae | ||||||
| Subfamilia Arvicolinae | ||||||
| Microtus guatemalensis Merriam, 1898 | AC, SN | AG, BQ, VS | 3, 5, 6, 7 | A | NT | |
| Subfamilia Neotominae | ||||||
| Baiomys musculus (Merriam, 1892) | DC, LLC, LLI, AC, LLPT, SL, SN, SS | AG, AH, BM, BQ, PZ, SA, SE, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Scotinomys teguina (Alston, 1877) | AC | AG, SE, VS | 2, 3, 4 | Pr | LC | |
| Neotoma mexicana Baird, 1855 | DC, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, PZ VG, VS | 1, 2, 3, 4, 5 6 7, 8 | LC | ||
| Habromys lophurus (Osgood, 1904) | AC, SS, VCA | AG, AH, BM, BQ, VS | 2, 3, 5, 6, 7 | NT | ||
| Peromyscus aztecus (de Saussure, 1860) | DC, AC, SL, SN, SS, VCA | AG, AH, BM, BQ SV, PZ, SE, VS | 1, 2, 3, 4, 5, 6, 7 | LC | ||
| Peromyscus guatemalensis Merriam, 1898 | LLC, AC, SL, SN, SS, VCA | AG, BM, BQ, PZ, PM, SE, VS | 1, 2, 3, 4, 5,6, 7, 8 | LC | ||
| Peromyscus gymnotis Thomas, 1894 | DC, LLC, AC, SL, SN, SS, VCA | AG, AH, BM, PZ, SE, VS | 1, 2, 3, 4, 5 | LC | ||
| Peromyscus leucopus (Rafinesque, 1818) | DC, AC, SS | AG, PZ, SE, VS | 3, 4, 5 | LC | ||
| Peromyscus levipes Merriam, 1898 | DC, AC, SL, SN, SS, VCA | AG, AH, BM, BQ, PZ, PM, SA, SE, VS | 1, 2, 3, 4, 5, 6, 7, 8 | LC | ||
| Peromyscus melanophrys (Coues, 1874) MX | DC, LLI, AC, SL, SN, SS, VCA | AG, AH, BM, BQ, PZ SE, VG, VS | 1, 2, 3, 4, 5 6 | LC | ||
| Peromyscus mexicanus (de Saussure, 1860) | DC, LLC, LLI, AC, LLPT, SL, SBP, SN, SS, VCA | AG, AH, BM, BQ, CA, SV, PZ, SA, SE, VG, VS | 1, 2, 3, 4, 5, 6, 7 | LC | ||
| Peromyscus zarhynchus Merriam, 1898 CHIS | AC, SL, SN | AG, BM, BQ, SV, PZ, VS, AH | 2, 3, 4, 5, 6, 7 | Pr | VU | |
| Reithrodontomys fulvescens J. A. Allen, 1894 | DC, AC, SL, SN, SS, VCA | AG, AH, BM, BQ, PZ, VS | 3, 4, 5, 6 | LC | ||
| Reithrodontomys gracilis J. A. Allen y Chapman, 1897 | LLC, AC, SN, SS, VCA | AG, AH, MG, PZ, VS | 1, 2, 3, 4, 5 6 | LC | ||
| Reithrodontomys megalotis (Baird, 1857) | AC, SL, SN, SS | AG, BM, BQ, SE, VS | 1, 3, 4, 5, 6, 7 | LC | ||
| Reithrodontomys mexicanus (de Saussure, 1860) | DC, AC, SL, SN, SS | AG, AH, BM, BQ, PZ, VS | 3, 4, 5, 6, 7 | LC | ||
| Reithrodontomys microdon Merriam, 1901 | AC, SN, SS | AG, BM, BQ, PZ, VS, AH | 4, 5, 6, 7, 8 | A | LC | |
| Reithrodontomys sumichrasti (de Saussure, 1860) | AC, SL, SN, SS, VCA | AG, AH, BM, BQ, PZ, PM, VS | 1, 2, 3, 4, 5, 6, 7, 8 | LC | ||
| Reithrodontomys tenuirostris Merriam, 1901 | AC, SS | AG, BM, BQ, VS | 5, 6, 7, 8 | VU | ||
| Subfamilia Sigmodontinae | ||||||
| Oligoryzomys fulvescens (de Saussure, 1860) | DC, LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, PZ, SA, VS | 1, 2, 3, 4, 5 6 | LC | ||
| Oryzomys alfaroi (J. A. Allen, 1891) | DC, LLC, AC, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, PZ, SE, VS | 1, 2, 3, 4, 5 6 7, 8 | LC | ||
| Oryzomys couesi (Alston, 1877) | DC, LLC, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ CA, MG, PZ, SA, SE, TU, VS | 1, 2, 3, 4, 5 6, 7 | A | LC | |
| Oryzomys rhabdops Merriam, 1901 | AC, SN, SS, VCA | AG, BM, VS, AH | 5, 6, 7, | VU | ||
| Oryzomys rostratus Merriam, 1901 | LLC, AC, LLPT, SL, SN, SS, VCA | AG, BM BQ, MG, PZ, SE, VS, AH | 1, 2, 3, 4, 5, 7 | LC | ||
| Oryzomys saturatior Merriam, 1901 | AC, SL, SN, SS, VCA | AG, AH BM, CA, PZ, SE, VS | 1, 2, 3, 4, 5, 8 | NT | ||
| Rheomys thomasi Dickey, 1928 | SS | AG, VS | 4 | Pr | NT | |
| Sigmodon mascotensis J.A. Allen, 1897 MX | SN, SS | BQ, PZ, VS, AH | 2, 3 | LC | ||
| Sigmodon toltecus (de Saussure, 1860) | DC, LLC, AC, LLPT, SL, SN, SS | AG, AH, BM, BQ, CA, PZ, SA, SE, VS | 1, 2, 3, 4, 5, 6, 7 | LC | ||
| Sigmodon zanjonensis Goodwin, 1932 | LLC, LLI, SS, VCA | AG, AH, BM BQ, PZ, TU, VS | 1, 2, 3, 4, 5 | LC | ||
| Subfamilia Tylomyinae | LC | |||||
| Nyctomys sumichrasti (de Saussure, 1860) | LLC, AC, SL, SN, SS, VCA | AG, BM, PZ SE, VS, AH | 1, 3, 4, 5, 6, 7, 8 | |||
| Ototylomys phyllotis Merriam, 1901 | AC, LLPT, SL, SBP, SN | AG, BM, CA, PZ, SE, VS, AH | 1, 2, 3, 4, 5 | LC | ||
| Tylomys bullaris Merriam, 1901 CHIS | AC | AH | 3 | A | CR | |
| Tylomys nudicaudus (Peters, 1866) | LLC, AC, LLPT, SL, SN, SS, VCA | AG, BM, BQ, PZ, SE, VS, AH | 1, 2, 3, 4, 5 | LC | ||
| Tylomys tumbalensis Merriam, 1901 CHIS | SN | AG, BM, VS | 1, 2, 4, 6 | Pr | CR | |
| Orden Carnivora | ||||||
| Familia Felidae | ||||||
| Subfamilia Felinae | ||||||
| Herpailurus yagouaroundi (É. Geoffroy Saint.- Hilaire, 1803) | DC, LLC, AC, SL, SS | AG, CA, PZ, VS, AH | 1, 2, 3, 5 | A | LC | I |
| Leopardus pardalis (Linnaeus, 1758) | LLC, AC, LLPT, SL, SS | AG, BM, CA, PZ, SE, VS, AH | 1, 2, 3, 4, 5, 6 | P | LC | I |
| Leopardus wiedii (Schinz, 1821) | AC, SL, SN, SS | BM, BQ, PZ, SE, VS, AH | 1, 2, 3, 4 | P | NT | I |
| Puma concolor (Linnaeus, 1771) | AC, SL, SS | AH, CA, SE, VS | 1, 2, 3, 5 | LC | I | |
| Subfamilia Pantherinae | ||||||
| Panthera onca (Linnaeus, 1758) | LLC, AC, LLPT, SL, SN, SS | AG, CA, MG, PZ, SE, VS, AH | 1, 2, 3, 4, 5 | P | NT | |
| Familia Canidae | ||||||
| Subfamilia Caninae | ||||||
| Canis latrans Say, 1822 | LLC, AC, SL, SN | MG, PZ, VS, AH | 1, 3 | LC | ||
| Urocyon cinereoargenteus (Schreber, 1775) | LLC, LLI, AC, SL, SN, SS, VCA | AG, AH, BM, BQ, MG, PZ, SE, SV, VS | 1, 2, 3, 4, 5 6 | LC | ||
| Familia Mephitidae | ||||||
| Conepatus leuconotus (Lichtenstein, 1832) | DC, LLC, AC, SN, SS, VCA | AG, BM, BQ, SV, PZ, PM, VS, AH | 1, 2, 3, 4, 5 6, 8 | LC | ||
| Conepatus semistriatus (Boddaert, 17854) | SL, SN | SE, VS | 1, 2, 3 | Pr | LC | |
| Mephitis macroura Lichtenstein, 1832 | DC, LLC, AC, SL, SN, SS | AG, AH, BQ, PZ, VS | 1, 3, 5, 6 | LC | ||
| Spilogale angustifrons A. H. Howell, 1902 | LLC, AC, SN, SS, VCA | AH, BM, PZ, VS | 1, 3, 4, 5, 7 | LC | ||
| Familia Mustelidae | ||||||
| Subfamilia Lutrinae | ||||||
| Lontra longicaudis (Olfers, 1818) | DC, LLC, AC, SL, SN, SS | AG, BQ, PZ, SE, VS, AH | 1, 2, 3, 4, 5 | A | NT | I |
| Subfamilia Mustelinae | ||||||
| Eira barbara (Linnaeus, 1758) | DC, AC, LLPT, SL, SS VCA | AG, AH, BQ, PZ, SE, VS | 1, 2, 3, 4, 5 | P | LC | III |
| Galictis vittata (Schreber, 1776) | AC, SL | PZ, SE, VS, AH | 1, 2, 3, 4, 5 | A | LC | III |
| Mustela frenata Lichtenstein, 1831 | AC, SL, SN, SS, VCA | AG, BM, BQ, PZ, VS, AH | 1, 3, 4, 5, 6, 7 | LC | ||
| Familia Procyonidae | ||||||
| Subfamilia Bassariscinae | ||||||
| Bassariscus sumichrasti (de Saussure, 1860) | AC, SN, SS, VCA | AH, BM, PZ, PM, VS | 1, 2, 3, 4, 5, 8 | Pr | LC | III |
| Potos flavus (Schreber, 1774) | DC, LLC, AC, LLPT, SL, SN, SS, VCA | AG, BM, CA, PZ, SE, VS, AH | 1, 2, 3, 4, 5, 6 | Pr | LC | III |
| Subfamilia Procyoninae | ||||||
| Nasua narica (Linnaeus, 1766) | LLC, LLI, AC, LLPT, SL, SN, SS | AG, AH, BM, CA, SV, MG, PZ, SE, VS | 1, 2, 3, 4, 5 | LC | III | |
| Procyon lotor (Linnaeus, 1758) | DC, LLC, AC, SL, SN, SS, VCA | AG, AH, MG, PZ, SV, VS | 1, 2, 3, 4, 5 | LC | ||
| Orden Artiodactyla | ||||||
| Familia Tayassuidae | ||||||
| Subfamilia Tayassuinae | ||||||
| Dicotyles crassus (Merriam, 1901) | LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, BQ, CA, SV, MG, PZ, SE, VS | 1, 2, 3, 4, 5 6 | LC | II | |
| Tayassu pecari (Link, 1795) | AC, SL, SN, SS | BM, CA, SV, PZ, SE, VS, AH | 1, 2, 3, 4 | P | VU | II |
| Familia Cervidae | ||||||
| Subfamilia Capreolinae | ||||||
| Mazama temama (Kerr, 1792) | AC, LLPT, SL, SN, SS | AG, AH, BM, CA, PZ, SE, VS | 1, 2, 3, 4, 5 | DD | ||
| Odocoileus virginianus (Zimmermann, 1780) | LLC, LLI, AC, LLPT, SL, SN, SS, VCA | AG, AH, BM, CA, PZ, SE, VS | 1, 2, 3, 4, 5, 6 | LC | ||
| Orden Perissodactyla | ||||||
| Familia Tapiridae | ||||||
| Tapirella bairdii (Gill, 1865) | LLC, AC, SL, SS | AG, AH, MG, PZ, SE, VS | 1, 2, 3, 4, 5 | P | EN | I |
Referencias
Álvarez, S. y T. Álvarez. 1991. Los murciélagos de Chiapas. Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional. Ciudad de México, México.
Álvarez-Castañeda, S.T. e I. Castro-Arellano. 2008. Tylomys tumbalensis. The IUCN Red List of Threatened Species 2008: e.T22575A9377455. Recuperado el 24 abril 2016 de http://dx.doi.org/10.2305/IUCN.UK.2008.RLTS.T22575A9377455
Álvarez del Toro, M. 1966. A note on the breeding of Baird’s tapir at Tuxtla Gutiérrez Zoo. International Zoo Yearbook 6:196-197.
Álvarez del Toro, M. 1991. Los mamíferos de Chiapas. Segunda ed. Gobierno del Estado, Tuxtla Gutiérrez, Chiapas, México.
Anderson, R. P., M. Weksler y D. S. Rogers. 2006. Phylo¬genetic analyses of pocket mice (Heteromyidae: Het¬eromyinae) based on allozymic and morphological data. Journal of Mammalogy 87:1218–1233.
Ashley, M., J. Norman y L. Stross. 1996. Phylogenetic analysis of the Perissodactyla family Tapiridae using mitochondrial cytochrome c oxidase (COII) sequences. Journal of Mammalian Evolution 3:315–326.
Baird, A. B., M. R. Marchán–Rivadeneira, S. G. Pérez y R. J. Baker. 2012. Morphological analysis and description of two new species of Rhogeessa (Chiroptera: Vespertilionidae) from the neotropics. Ocassional Papers, Museum of Texas Tech University 307:1–25.
Baker, R. J., S. Solari y F. G. Hoffmann. 2002. A new Central American species from the Carollia brevicauda complex. Occasional Papers, Museum of Texas Tech University 217:1-12.
Bradley, R. D., D. D. Henson y N. D. Durish. 2008. Re–evalu¬ation of the geographic distribution and phylogeography of the Sigmodon hispidus complex based on mitochondrial DNA sequences. The Southwestern Naturalist 53:301–310.
Brandon–Jones, D. 2006. Apparent confirmation that Alouatta villosa (Gray, 1845) is a senior synonym of A. pigra Lawrence, 1933 as the species–group name for the black howler monkey of Belize, Guatemala and Mexico. Pri¬mate Conservation 21:41–43.
Briones-Salas, M., M. Cortès-Marcial y M. C. Lavariega. 2015. Diversidad y distribución geográfica de los mamíferos terrestres del estado de Oaxaca, México. Revista Mexicana de Biodiversidad 86:685-710.
Carazo, E. 2007. Atlas socioeconómico de la selva. Chiapas, México: PRODESIS. EPYPSA.
Carraway, L. N. 2007. Shrews (Eulypotyphla:Soricidae) of Mexico. Monographs of the Western North American Naturalist 3:1-91.
Carroll, D. S. y R. D. Bradley. 2005. Systematics of the genus Sigmodon: DNA sequences from beta–fibrinogen and cytochrome–b. The Southwestern Naturalist 50:342–349.
Carroll, D. S., L. Peppers y R. D. Bradley. 2005. Molecular systematics and phylogeography of the Sigmodon hispidus species group. In Contribuciones Mastozoológicas en Homenaje a Bernardo Villa, V. Sánchez-Cordero y R. A. Medellín (eds.). Instituto de Biología e Instituto de Ecología, Universidad Nacional Autónoma de México-Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, D. F. p. 87-99.
Ceballos, G. y P. Rodríguez. 1993. Diversidad y conservación de los mamíferos de México: II. Patrones de Endemicidad. In Avances en el estudio de los mamíferos de México Publicaciones Especiales I, R. Medellín y G. Ceballos (eds.). Asociación Mexicana de Mastozoología, A. C., México, D. F. p. 87-108.
Ceballos, G. y G. Oliva. 2005. Los Mamíferos Silvestres de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad-Fondo de Cultura Económica, México, D.F.
CITES (Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres). 2014. Appendices I, II, and III. Convention on international trade in endangered species of wild fauna and flora. Ginebra: United Nations Environment Programme.
Cole, F. R. y D. E. Wilson. 2006a. Leptonycteris curasoae. Mammalian Species 796:1–3.
Cole, F. R. y D. E. Wilson. 2006b. Leptonycteris yerbabuenae. Mammalian Species 797:1–7.
Comisión Internacional en la Nomenclatura Zoológica (ICZN). 1998. Opinion 1894. Regnum Animale…, Ed. 2 (M. J. Brisson, 1762): rejected for nomenclatural purposes, with the conservation of the mammalian generic names for Philander (Marsupialia), Pteropus (Chiroptera), Glis, Cuniculus and Hydrochoerus (Rodentia), Meles, Lutra and Hyaena (Carnivora), Tapirus (Perissodactyla), Tragulus and Giraffa (Artiodactyla). Bulletin of Zoological Nomenclature 55:64–71.
Daza, J., T. Castoe y C. Parkinson. 2010. Using regional comparative phylogeographic data from snake lineages to infer historical processes in Middle America. Ecography 33:343–354.
Dragoo, J. W., R. D. Bradley, R. L. Honeycutt y J. W. Templeton. 1993. Phylogenetic relationships among the skunks: a molecular perspective. Journal of Mammalian Evolution 1:255-267.
Eger, J. L. 2008. Family Molossidae P. Gervais, 1856. In Mammals of South America. Vol. 1: Marsu¬pials, xenarthrans, shrews, and bats, A. L. Gardner (ed.). University of Chicago Press. p. 399–439.
Escobedo-Morales, L. A., L. León-Paniagua, J. Arroyo-Cabrales y F. Greenaway. 2006. Distributional records for mammals from Chiapas, México. The Southwestern Naturalist 51:269-272.
Espinoza, J., C. Lorenzo y E. Rios. 2011. Variación mor¬fológica y morfométrica de Heteromys desmarestianus en Chiapas, México. Therya 2:139–154.
GBIF ¬(Global Biodiversity Information Facility). 2013. En Global Biodiversity Information Facility Database. Recuperado el 11 diciembre 2013 de http://www.gbif.org/species/2459582
Gardner, A. L. 2008. Tribe Sturnirini [Miller, 1907]. In Mammals of South America. Vol. 1: Marsu¬pials, xenarthrans, shrews, and bats, A. L. Gardner (ed.). University of Chicago Press. p. 363–376.
Geist, V. 1998. Deer of the World. The Evolution, behavior and Ecology. Stackpole Books. Mechanicsburg, Pensylvania.
Groves, C. y P. Grubb. 2011. Ungulate taxonomy. The Johns Hopkins University Press, Baltimore.
Grubb, P. 2005. Order Artiodactyla. In Mammal species of the world. A taxonomic and geographic refer¬ence, D. E. Wilson y Reeder D. A. M. (eds.). The Johns Hopkins University Press, Baltimore. p. 637–722.
Guevara, L., V. Sánchez-Cordero, L. León-Paniagua y N. Woodman. 2014. A new species of small-eared shrew (Mammalia, Eulipotyphla, Cryptotis) from the Lacandona rainforest, México. Journal of Mammalogy 95:739-753.
Hafner, M. S., J. E. Light, D. J. Hafner, M. S. Hafner, E. Reddington, D. S. Rogers y B. R. Riddle. 2007. Basal clades and molecular systematic of heteromyid rodents. Journal of Mammalogy 88:1129–1145.
Henson, D. D. y R. D. Bradley. 2009. Molecular systematics of the genus Sigmodon: results from mitochondrial and nuclear gene sequences. Canadian Journal of Zoology 87:211–220.
Herrera, A. L. 1897. Datos para la zoología de Chiapas. Anales del Museo Nacional de México, Época 1ª 4:136-139 mamíferos y 139-143 aves.
IUCN (International Union for Conservation of Nature). 2015. IUCN Red list of threatened species. International Union for Conservancy of Nature and Natural Resources. Recuperado el 15 enero 2010 de http://www.iucnredlist.org/
INEGI (Instituto Nacional de Estadística, Geografía e Informática). 1981. Subprovincias fisiográficas, escala 1:1000000, SEOP (Secretaría de Obras Públicas, del Gobierno del Estado de Chiapas).
INEGI (Instituto Nacional de Estadística, Geografía e Informática). 2015. Guía para la interpretación de cartografía: uso del suelo y vegetación: escala 1:250, 000: serie V / Instituto Nacional de Estadística y Geografía.– México: INEGI, c2014. 195 p.
Koleff, P., J. Soberón, H. T.Arita, P. Dávila, O. Flores-Villela, J. Golubov, G. Halffter, A. Lira-Noriega, C. E. Moreno, E. Moreno, M. Munguía, M. Murguía, A. G. Navarro-Sigüenza, O. Téllez, L. Ochoa-Ochoa, A. T. Peterson y P. Rodríguez. 2008. Patrones de diversidad espacial en grupos selectos de especies. In Capital natural de México, vol. I: Conocimiento actual de la biodiversidad, J. Soberón, G. Halffter y J. Llorente Bousquets (comps.). Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, D. F. p. 323-364.
Lee, T. E., Jr., S. R. Hoofer y R. A.Van den Bussche. 2002. Molecular phylogenetics and taxonomy revision of the genus Tonatia (Chiroptera: Phyllostomidae). Journal of Mammalogy 83:49-57.
López-Wilchis, R. 2006. Especies y ejemplares de mamíferos mexicanos depositados en colecciones extranjeras. In Colecciones Mastozoológicas de México, C. Lorenzo, E. Espinoza, M. Briones y F. A. Cervantes (eds.). Instituto de Biología, UNAM-Asociación Mexicana de Mastozoología, A. C., México, D. F. p. 111-120.
López–Wilchis, R., L. M. Guevara–Chumacero, N. P. Ángeles, J. Juste, C. Ibáñez y I. D. L. A. Barriga–Sosa. 2012. Taxonomic status assessment of the Mexican populations of funnel–eared bats, genus Natalus (Chiroptera: Natali¬dae). Acta Chiropterologica 14:305–316.
Lorenzo, C., S. T. Álvarez-Castañeda, E. Arellano, J. Arroyo-Cabrales, J. Bolaños et al. 2012. Los mamíferos de México en las colecciones científicas de Norteamérica. Therya 3:239-262.
Lorenzo, C., C. Kraker-Castañeda y J. Bolaños-Citalán. 2015. Los murciélagos filostómidos de Chiapas, México y Guatemala. El Colegio de la Frontera Sur, Museo de Historia Natural de la Universidad de San Carlos de Guatemala, Programa para la Conservación de Murciélagos de Guatemala, Estrategia de Conservación Murciélagos de Centroamérica.
Magurran, A. E. 1988. Ecological diversity and its measurement. Princeton University Press.
McDonough, M. M., L. K. Ammerman, R. M. Timm, H. H. Ge¬noways, P. A. Larsen y R. J. Baker. 2008. Speciation within bonneted bats (genus Eumops): the complexity of morphological, mitochondrial, and nuclear data sets in systematics. Journal of Mammalogy 89:1306–1315.
Naranjo, E. J., C. Lorenzo y A. Horváth. 2005. La diversidad de mamíferos en Chiapas. In Diversidad biológica en Chiapas, M. González-Espinosa, N. Ramírez-Marcial y L. Ruiz-Montoya (eds.). Plaza y Valdés, México, D. F. p. 221-263.
Naranjo, E. J., R. Dirzo, J. C. López-Acosta, J. Rendón-von Osten, A. Reuter y O. Sosa-Nishizaki. 2009. Impacto de los factores antropogénicos de afectación directa a las poblaciones silvestres de flora y fauna. In Capital natural de México, Vol. II: Estado de conservación y tendencias de cambio, J. Soberón, G. Halffter y J. Llorente-Bousquets (eds.). Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, D. F. p. 247-276.
Naranjo, E. J., C. Lorenzo, A. Horváth, A. Riechers, E. Espinoza-Medinilla, J. E. Bolaños-Citalán, R. Vidal-López y E. Cruz-Aldán. 2013. Diversidad y conservación de los mamíferos. In La biodiversidad en Chiapas: Estudio de Estado, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad-Gobierno del Estado de Chiapas, México, D. F. p. 351-361.
Naranjo, E. J., C. Lorenzo, J. Bolaños-Citalán y A. Horváth. En prensa. Diversidad y conservación de los mamíferos terrestres de Chiapas, México. In Riqueza y Conservación de los Mamíferos en México a Nivel Estatal, M. Briones-Salas, Y. Hortelano-Moncada, G. Magaña-Cota, G. Sánchez-Rojas y J. E. Sosa-Escalante (eds.). Instituto de Biología, Universidad Nacional Autónoma de México-Asociación Mexicana de Mastozoología A. C.-Universidad de Guanajuato, México, D. F.
Ortiz-Pérez, M. A., J. R. Hernández-Santana y J. M. Figueroa-Mah-Eng. 2004. Reconocimiento fisiográfico y geomorfológico. In Biodiversidad de Oaxaca, A. J. García-Mendoza, M. J. Ordóñez y M. Briones-Salas (eds.). Instituto de Biología, Universidad Nacional Autónoma de México-Fondo Oaxaqueño para la Conservación de la Naturaleza-World Wildlife Found, México, D. F. p. 43–53.
Ramírez-Pulido, J. N. y González-Ruiz. 2006. Las colecciones de mamíferos de México: origen y destino. In Colecciones Mastozoológicas de México, C. Lorenzo, E. Espinoza, M. Briones y F. A. Cervantes (eds.). Instituto de Biología, UNAM-Asociación Mexicana de Mastozoología, A. C., México, D. F. p. 73-110.
Ramírez-Pulido, J., N. González-Ruiz, A. L. Gardner y J. Arroyo-Cabrales. 2014. List of recent land mammals of Mexico, 2014. Special Publications, Museum of Texas Tech University 63:1-69.
Retana, O., y C. Lorenzo. 2002. Lista de mamíferos terrestres de Chiapas: endemismo y estado de conservación. Acta Zoológica Mexicana 85:25-49.
Retana-Guiascón, O. G. 2006. Raíces históricas de las colecciones zoológicas en México y del Museo Nacional de Historia Natural. In Colecciones Mastozoológicas de México, C. Lorenzo, E. Espinoza, M. Briones y F. A. Cervantes (eds.). Instituto de Biología, UNAM-Asociación Mexicana de Mastozoología, A. C., México, D. F. p. 57-72.
Rogers, D. S. y M. W. González. 2010. Phylogenetic rela¬tionships among spiny pocket mice (Heteromys) inferred from mitochondrial and nuclear sequence data. Journal of Mammalogy 91:914–930.
SEMARNAT (Secretaría de Medio Ambiente y Recursos Naturales). 2010. Norma Oficial Mexicana NOM-059-SEMARNAT-2010. Protección ambiental, especies nativas de flora y fauna silvestres de México, categorías de riesgo y especificaciones para su inclusión, exclusión o cambio, y lista de especies en riesgo. Diario Oficial de la Federación, 30 de diciembre de 2010, Segunda Sección. México, 2454, 1-77.
Simmons, N. B. y C.O. Handley, Jr. 1998. A revision of Centronycteris Gray (Chiroptera: Emballonuridae) with notes on natural history. American Museum Novitates 3239:1-28.
Simmons, N. B. y A. L. Wetterer. 2002. Phylogeny and con¬vergence in cactophilic bats. In Columnar Cacti and their Mutualists: Evolution, Ecology, and Con¬servation, T. H. Fleming y A. Valiente–Banuet (eds.). The University of Arizona Press. p. 87–121.
Simmons, N. B. 2005. Order Chiroptera. In Mammal species of the world. A taxonomic and geographic refer¬ence, D. E. Wilson y D. A. M. Reeder (eds.). Baltimore: The Johns Hopkins University Press. p. 312–529.
Soto-Pinto, L., M. A. Castillo-Santiago y G. Jiménez-Ferrer. 2012. Agroforestry systems and local institutional development for preventing deforestation in Chiapas, Mexico. In Deforestation Around the World, P. Moutinho (ed.). InTech, Rijeka, Croacia. p. 333-350. Recuperado el 24 marzo 2016 de http://www.intechopen.com/books/deforestation-around-the-world.
Tejedor, A. 2005. A new species of funnel–eared bat (Na¬talidae: Natalus) from Mexico. Journal of Mammalogy 86:1109–1120.
Velazco, P. M. y B. D. Patterson. 2013. Diversification of the yellow-shouldered bats, genus Sturnira (Chiroptera, Phyllostomidae), in the New World tropics. Molecular Phylogenetics and Evolution 68:683–698.
Velazco, P. M. y N. B. Simmons. 2011. Systematics and taxonomy of great striped–faced bats of the genus Vampyrodes Thomas, 1900 (Chiroptera: Phyllostomidae). American Museum Novitates 3710:1–35.
Voss, R. S. 1998. Systematics and ecology of Ichthyomyne rodents (Muroidea): patterns of morphological evolution in a small adaptative radiation. Bulletin of the American Museum of Natural History 210:1-113.
Voss, R. S. y S. A. Jansa. 2003. Phylogenetic studies on didelphid marsupials II. Nonmolecular data and new IRBP sequences: separate and combined analyses of didelphine relationships with denser taxon sampling. Bulletin of the American Museum of Natural History 276:1-82.
Whittaker, R. H. 1972. Evolution and Measurement of Species Diversity. International Association for Plant Taxonomy 21:213-251.
Williams, S. L. y H. H. Genoways. 2008. Subfamily Phyllos¬tominae Gray, 1825. In Mammals of South America. Vol. 1: Marsu¬pials, xenarthrans, shrews, and bats, A. L. Gardner (ed.). University of Chicago Press. p. 255–300.
Woodman, N. y R. M. Timm. 1999. Geographic variation and evolutionary relationships among broad-clawed shrews of the Cryptotis goldmani-group (Mammalia: Insectivora: Soricidae). Fieldiana Zoology 91:1-35.
Woodman, N., J. O. Matson, J. J. McCarthy, R. P. Eckerlin, W. Bulmer y N. Ordóñez-Garza. 2012. Distributional records of shrews (Mammalia, Soricomorpha, Soricidae) from northern Central America with the first records of Sorex from Honduras. Annals of Carnegie Museum 80:207-237.
Wozencraft, W. C. 2005. Order Carnivora. In Mammal species of the world. A taxonomic and geographic refer¬ence, D. E. Wilson y D. A. M. Reeder (eds.). Baltimore: The Johns Hopkins University Press. p. 512–628.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.*


