Ontogenia de los tricomas foliares de Tilia caroliniana subsp. floridana (Malvaceae)
Marysol Ramírez-Díaz a, Teresa Terrazas a, *, Silvia Aguilar-Rodríguez b, Alicia Rojas-Leal a y Dorismilda Martínez-Cabrera c
a Instituto de Biología, Universidad Nacional Autónoma de México, Apartado postal 70-633, Ciudad Universitaria, 04510 Ciudad de México, México
b Laboratorio de Botánica, Unidad de Morfología y Función, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autónoma de México, Av. de los Barrios 1, Los Reyes Iztacala, 54090 Tlalnepantla de Baz, Estado de México, México
c Instituto Tecnológico de Huejutla, Km 5.5 carretera Huejutla-Chalahuiyapa, Apartado postal 94, 43000 Huejutla de Reyes, Hidalgo, México
*Autor para correspondencia: tterrazas@ib.unam.mx (T. Terrazas)
Resumen
En este trabajo se describe la ontogenia de los tricomas presentes en las láminas foliares de individuos de Tilia caroliniana subsp. floridana, utilizando microscopía de luz y electrónica de barrido. El objetivo fue reconocer, con base en la ontogenia, los diferentes tipos de tricomas y sus posibles transiciones. Yemas y hojas, en varios estados de desarrollo, se recolectaron en campo y procesaron para las diferentes microtecnias. Se describe el desarrollo de tricomas aciculares, fasciculados, estrellados y glandulares. La ontogenia reveló que todos los tricomas inician su desarrollo a partir de una célula protodérmica. Con excepción del tricoma acicular, que es unicelular, los otros 3 tipos multicelulares se distinguen por el tipo y el número de divisiones de la célula protodérmica que les da origen y determina su forma. Las divisiones anticlinales predominan en el desarrollo de los tricomas fasciculados y estrellados y las periclinales en los glandulares. Aquí encontramos 2 tipos más de tricomas glandulares que los registrados en otros estudios para Tilia. Los tricomas fasciculados se diferencian de los estrellados porque sus brazos se observan erectos, mientras que los estrellados los tienen postrados. No se observaron estadios transicionales entre los distintos tipos de tricomas, lo que permitirá una codificación rigurosa de los mismos en futuros estudios filogenéticos.
Palabras clave:
Anatomía foliar; Microscopio electrónico de barrido; Series de transición; Tricomas estrellados; Tricomas glandulares
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Foliar trichome ontogeny of Tilia caroliniana subsp. floridana (Malvaceae)
Abstract
The trichome ontogeny of Tilia caroliniana subsp. floridana leaf blades is described, using light and scanning electronic microscopy. Our aim was to recognize the different trichome types based on ontogeny as well as its possible transitions. Buds and leaves, in different development stages, were collected in the field and processed by different microtechnics. The development of acicular, fasciculate, stellate and glandular trichomes is described. Ontogeny revealed that all trichomes start their development from a protodermal cell. With the exception of the acicular trichome, which is unicellular, the other 3 multicellular trichome types differ in the type and number of divisions of the protodermal cell that determine their shape. The anticlinal divisions predominate in the development of the fasciculate and stellate trichomes and the periclinal cell divisions in the glandular ones. We found 2 additional glandular trichome types than those recorded in previous studies for Tilia. The fasciculate trichomes differ from the stellate because their arms are erect, while the stellate trichomes have prostrate arms. No transitional stages were observed between the different trichome types, which will allow a rigorous coding of them in future phylogenetic studies.
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Keywords:
Foliar anatomy; Scanning electron microscopy; Transition series; Stellate trichomes; Glandular trichomes
Introducción
Los tricomas son apéndices epidérmicos especializados, cuyas funciones se relacionan con la defensa contra herbívoros y diversos patógenos, así como con la fotoprotección (Agati et al., 2013; Kim et al., 2012; Levin, 1973). Por ello, el estudio de las características de los tricomas permite conocer el papel que desempeñan en varios aspectos de la adaptación fisiológica y ecológica de las plantas, así como en sus procesos evolutivos (Bickford, 2016; Nogueira et al., 2013; Payne, 1978). Los tricomas son uno de los atributos que frecuentemente se emplean en la taxonomía de numerosos grupos de plantas; más de 300 tipos se han descrito para caracterizar su variación morfológica (Kolb y Müller, 2004). Debido a dicha diversidad, algunos autores han propuesto clasificarlos en 2 grupos: glandulares y no glandulares (Inamdar et al., 1983; Theobald et al., 1980). Sin embargo, caracterizarlos cuando han alcanzado un desarrollo completo, puede resultar complicado, por lo que los estudios ontogenéticos de los tricomas permiten validar la presencia de algunos tipos en ciertos taxones (Ma et al., 2016). Ejemplo de ello es el trabajo de Inamdar (1967) quien confirmó la presencia de tricomas peltados en la familia Oleaceae y reconoció 3 tipos más de tricomas según el plano de división (anticlinal, periclinal, oblicua o irregular) de la célula inicial del escudo. Por otra parte, Gama et al. (2013) reconocen tricomas peltados en Bignonia aequinoctialis L. (Bignoniaceae), los cuales se desarrollan a partir de una célula protodérmica con divisiones periclinales que originan la célula del pedúnculo y la célula inicial de la cabeza. Varios autores han recalcado la importancia de la ontogenia de los tricomas con un enfoque evolutivo (Carlquist, 1961; Cowan, 1950). Ramayya (1972) propone una clasificación filogenética de los tricomas y reconoce 5 sistemas de desarrollo de los mismos. Con base en esos sistemas de desarrollo, Rao (1990) aclara la posición taxonómica del género Muntingia L. (Muntingiaceae). Varios estudios sobre las características morfológicas de los tricomas, han revelado un papel relevante en la sistemática de plantas a nivel de género (Hardin, 1979; Rajput et al., 1985; Redonda-Martínez et al., 2012) y especie (Carvalho-Sobrinho et al., 2009; Celep et al., 2014; Martínez-Gordillo y Espinosa-Matías, 2005; Shaheen et al., 2009).
Para las Malvaceae hay estudios que describen con detalle sus tricomas y evidencian su valor para distinguir especies (Garcia et al., 2014; Rendón-Carmona et al., 2006). Un caso particular es el género Tilia L. (Malvaceae), género de la subfamilia Tilioideae que reúne cerca de 45 especies (que se distribuyen exclusivamente en las zonas templadas del hemisferio norte; Judd et al., 2007). Más de la mitad de las especies se reconocen de Europa y Asia (Pigott, 2012) y un número aún indeterminado de Norteamérica (McCarthy y Mason-Gamer, 2016). Jones (1968) hace una revisión para el género en Norteamérica y menciona que se pueden separar las especies con base en los tricomas estrellados de 4 y 8 brazos. En México, Tilia se encuentra en el bosque mesófilo de montaña, entre 770 y 2,600 m snm, desde Chihuahua hasta Oaxaca (Hardin, 1990). Está representado por árboles caducifolios con hojas alternas, largamente pecioladas, láminas ovadas a elípticas, margen aserrado o dentado-mucronado e inflorescencias terminales o axilares, dispuestas sobre una bráctea foliácea, con flores hermafroditas. Para México, Hardin (1990) reconoce tan solo a Tilia americana var. mexicana Schltdl. con base exclusivamente en la morfología y combinación de 4 tipos de tricomas (acicular, glandular, fasciculado y estrellado). Por su parte, Pigott (2012) en su monografía de Tilia, considera que en México solo está presente Tilia caroliniana Mill. con 2 subespecies (T. caroliniana subsp. floridana (Small) A.E. Murray y T. caroliniana subsp. occidentalis (Rose) Pigott), las cuales pueden separarse por la presencia y abundancia de los tricomas estrellados que se distribuyen en la cara abaxial de la lámina de las ramas reproductivas, pero no hace referencia a los otros tipos de tricomas. Este autor también enfatizó que las características más importantes de los tricomas estrellados son el número de brazos y la longitud de éstos. A pesar de los trabajos mencionados, la taxonomía de Tilia sigue sin resolverse. No obstante que los tricomas en Tilia son fundamentales para la delimitación de sus taxones, se desconoce cómo es el desarrollo de los mismos y si es posible diferenciar los 4 tipos (glandular bulboso uniseriado, acicular, estrellado y fasciculado) reconocidos por Hardin (1990) desde las primeras divisiones de las células epidérmicas y si hay estados intermedios en ellos. El objetivo de esta investigación fue describir la ontogenia de los apéndices epidérmicos foliares en los individuos de T. caroliniana subsp. floridana de distribución en Sierra Madre Oriental, México para corroborar los 4 tipos definidos por Hardin (1990) y evaluar la presencia de formas intermedias o un mayor número de tipos de tricomas.
Materiales y métodos
Se seleccionaron 2 poblaciones de Tilia caroliniana subsp. floridana para seguir el desarrollo de las hojas desde el estadio de yema a partir de octubre del 2016 hasta la expansión de las hojas en marzo del 2017. La primer población se encuentra en Tlanchinol, Hidalgo (20°59’16” N, 98°39’36” O; D. Martínez-Cabrera 341, 342, 343) y la segunda en Cuetzalan, Puebla (19°46’23” N, 97°38’36” O; M. Ramírez-Díaz 12, 13). En ambas poblaciones se recolectaron yemas vegetativas en diferentes estadios de desarrollo y hojas maduras, se fijaron en CRAF III por 48 h (Ruzin, 1999); posteriormente, se lavaron y conservaron con etanol al 50%. Los ejemplares de respaldo están depositados en el Herbario Nacional de México (MEXU).
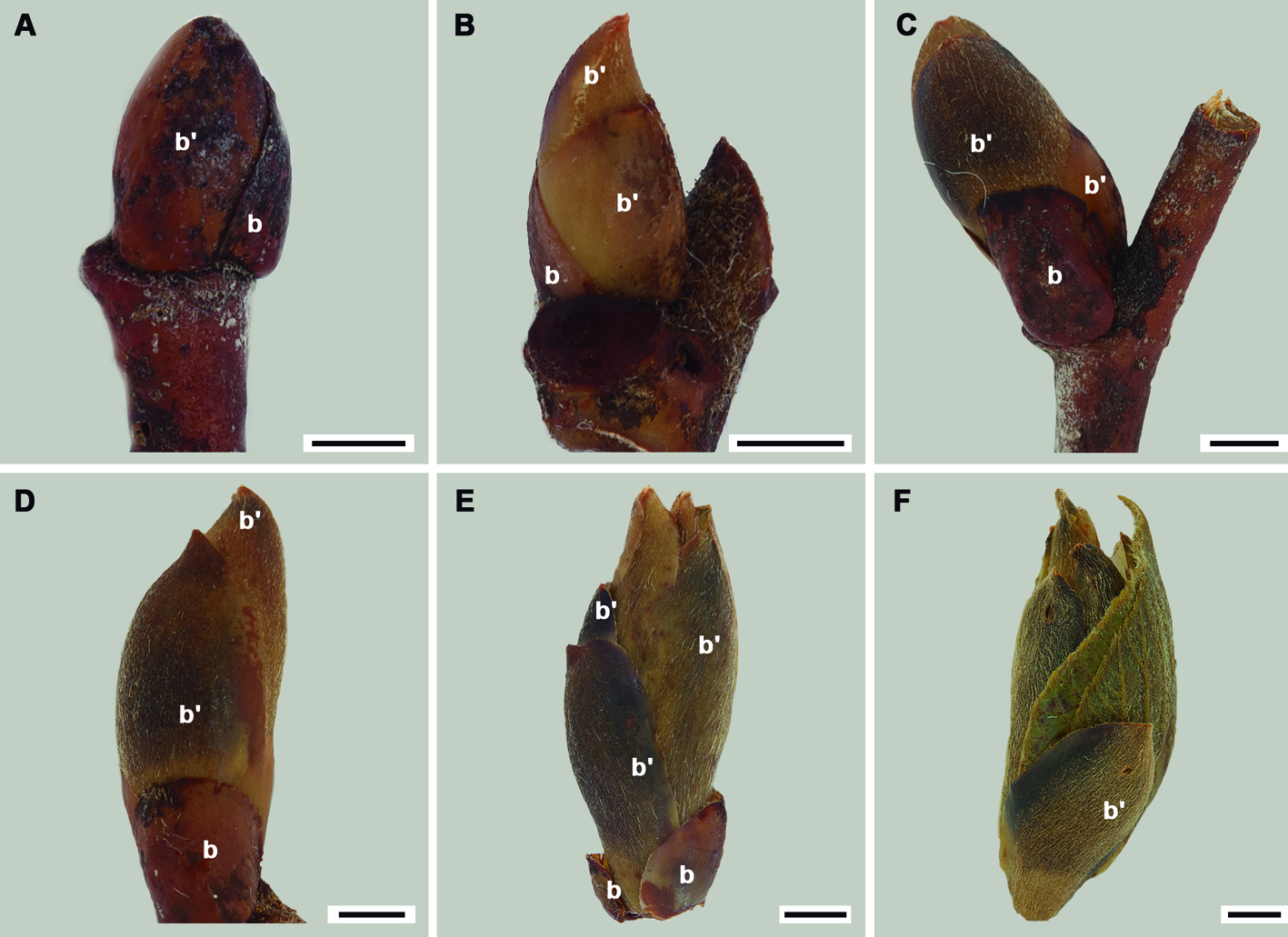
Las yemas recolectadas (más de 50) se clasificaron en 6 estadios de desarrollo según el tamaño (fig. 1) y sus hojas se numeraron comenzando por la más externa. Para microscopía de luz se emplearon yemas completas de los estadios 1 y 2; así como 3 hojas de las yemas de los estadios 1 al 4, las cuales se incluyeron en glicol metacrilato y se cortaron a 4 µm de grosor siguiendo la técnica de Zarlavzky (2014). Las hojas de los estadios 5 y 6 presentaron todos los tipos de tricomas en su fase de maduración por lo que no se ilustran en este trabajo. Además, se realizaron cortes a mano alzada de hojas maduras y se tiñeron con safranina-verde rápido (Ruzin, 1999). Las preparaciones se revisaron con un microscopio óptico Olympus BX51 y se fotografiaron con el analizador de imágenes Image Pro Plus versión 6.01. Para las observaciones en el microscopio electrónico de barrido, las hojas disectadas de las yemas de los estadios 2 a 4 y las maduras, se deshidrataron en una serie gradual de alcohol etílico (50-100%), llevadas al punto crítico y cubiertas con una película de oro en una ionizadora (Hitachi-S-2460N). Las fotografías de sus superficies adaxial y abaxial se tomaron en un microscopio Hitachi-S-2460N (15 Kv) en el Instituto de Biología, Universidad Nacional Autónoma de México.
Resultados
Estructura de las yemas y desarrollo de las hojas. La yema del estadio 1 alcanza un máximo de 5 mm de largo, está cubierta por brácteas coriáceas de color café obscuro y contiene 4 láminas foliares, cada una de ellas con una bráctea de textura membranosa; la lámina foliar más pequeña mide 0.3 mm. En este estadio (1) la yema pasa todo el invierno (octubre-febrero, fig. 1A). Una vez que se inicia el crecimiento de las yemas éstas se hinchan y aumentan su tamaño, separándose las brácteas coriáceas y exponiendo las brácteas membranosas de color café claro (fig. 1B-E) hasta que las láminas quedan expuestas (fig. 1F). En las yemas, las brácteas se distinguen además de su color y textura por sus abundantes tricomas aciculares (fig. 2A, B); en la lámina foliar, las células protodérmicas abaxiales se encuentran en diferentes etapas de desarrollo de los tricomas, excepto en los dientes en formación. Como se mencionó, la mayoría de los tricomas están en desarrollo, pero algunos de los fasciculares y los glandulares bulbosos ya completaron su desarrollo (fig. 2C). En contraste, la protodermis adaxial presenta escasos tricomas diferenciándose únicamente sobre las venas (fig. 2D). A nivel de meristemo, los primordios foliares tienen protodermis con divisiones anticlinales abundantes, algunas células ya diferenciándose en tricomas aciculares (fig. 2E), los cuales son escasos hacia ambas superficies foliares. El mesofilo ya es evidente, con 2 estratos celulares e inicia la diferenciación del tejido vascular en la vena media (fig. 2F). En hojas de 0.7 mm de longitud, la vena media ya está diferenciada y la lámina continúa su expansión; se inicia la diferenciación de las venas secundarias, el mesofilo presenta 3 estratos y en la epidermis abaxial hay mayor abundancia de tricomas, los cuales se encuentran aún en diferentes estados de desarrollo (fig. 2G). En hojas más desarrolladas (> 0.7 mm de longitud) se aprecian en la superficie abaxial los tricomas, muy abundantes, en especial los glandulares y los aciculares. Ambos tipos se encuentran principalmente sobre las venas (fig. 2H) aunque también se observan en las regiones intercostales; en este momento se inicia el desarrollo de los fasciculares y estrellados. Cuando la lámina, alcanza 1.9 mm de longitud, conserva los 3 estratos celulares en el mesofilo; además la vena media y las secundarias han completado su desarrollo y sus elementos traqueales también. Se diferencian las venas de órdenes mayores y asociados a éstas, los tricomas en la superficie abaxial de la epidermis (fig. 2I). Se siguen conservando pocos tricomas en la superficie adaxial de la lámina. Una vez que la hoja se libera de las escamas de la yema, ésta se expande rápidamente alcanzado su tamaño final (fig. 2J) y es cuando los tricomas estrellados son más abundantes que en etapas previas. Estos tricomas estrellados, junto con los otros tipos, forman una intrincada red que no permite ver las células epidérmicas abaxiales (fig. 2K).
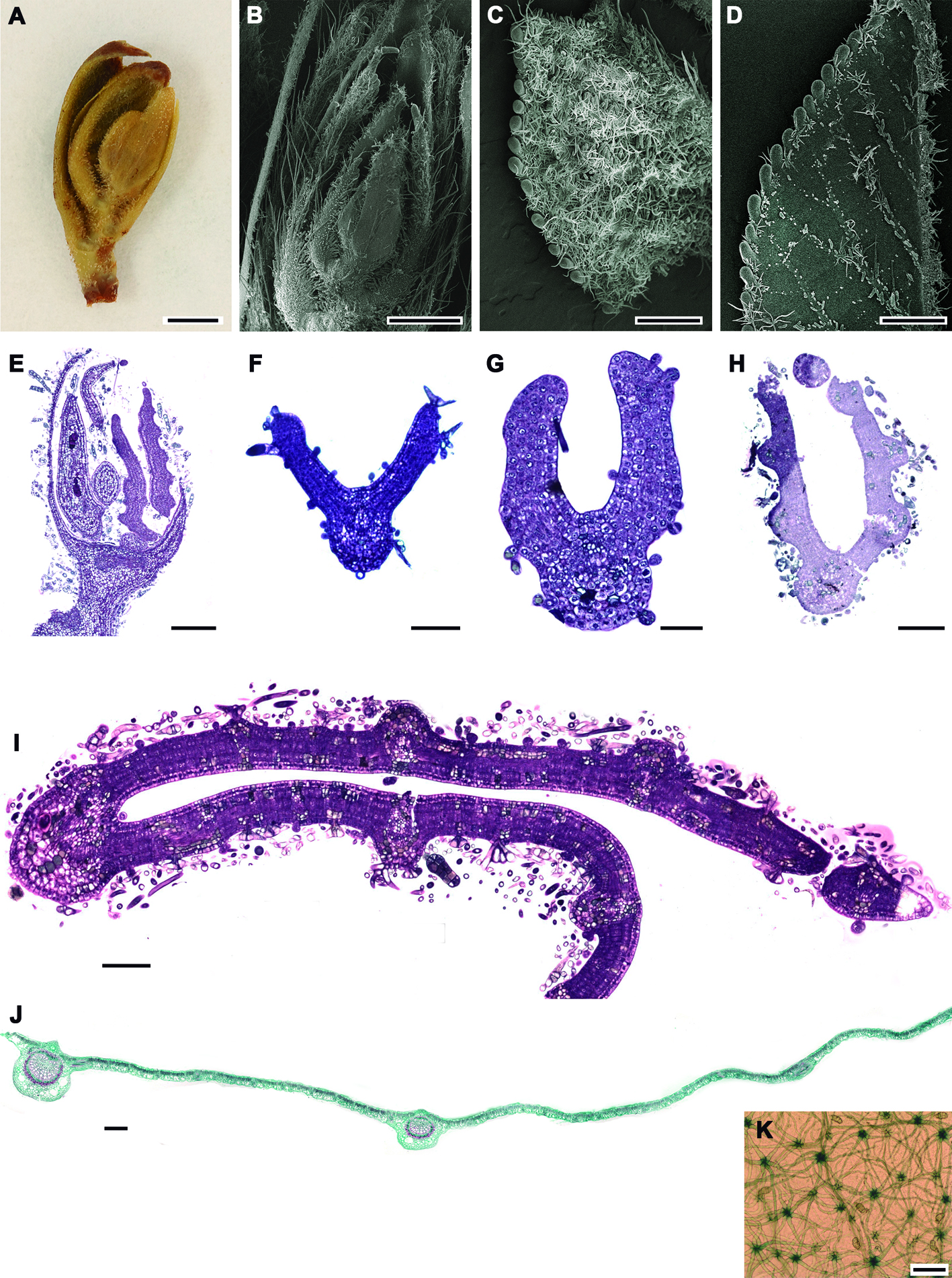
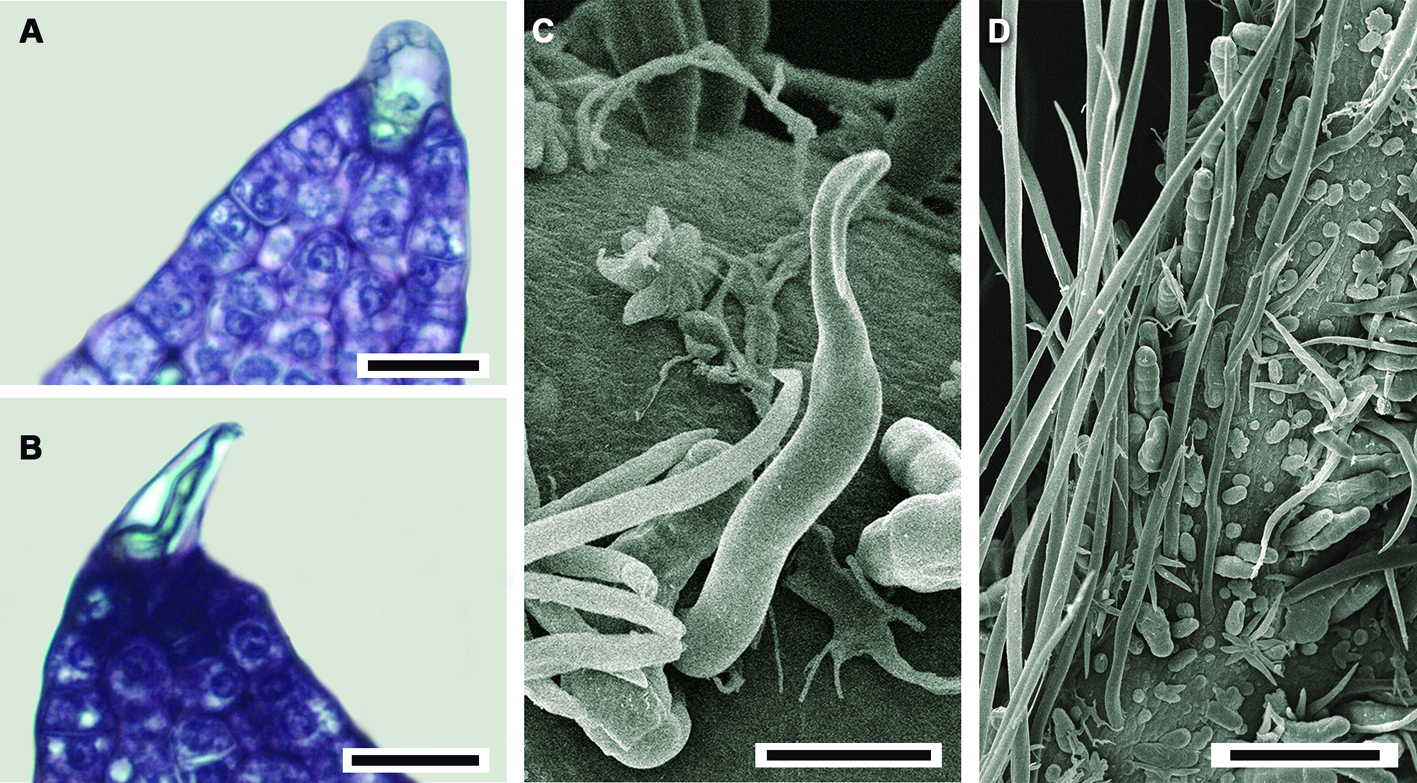
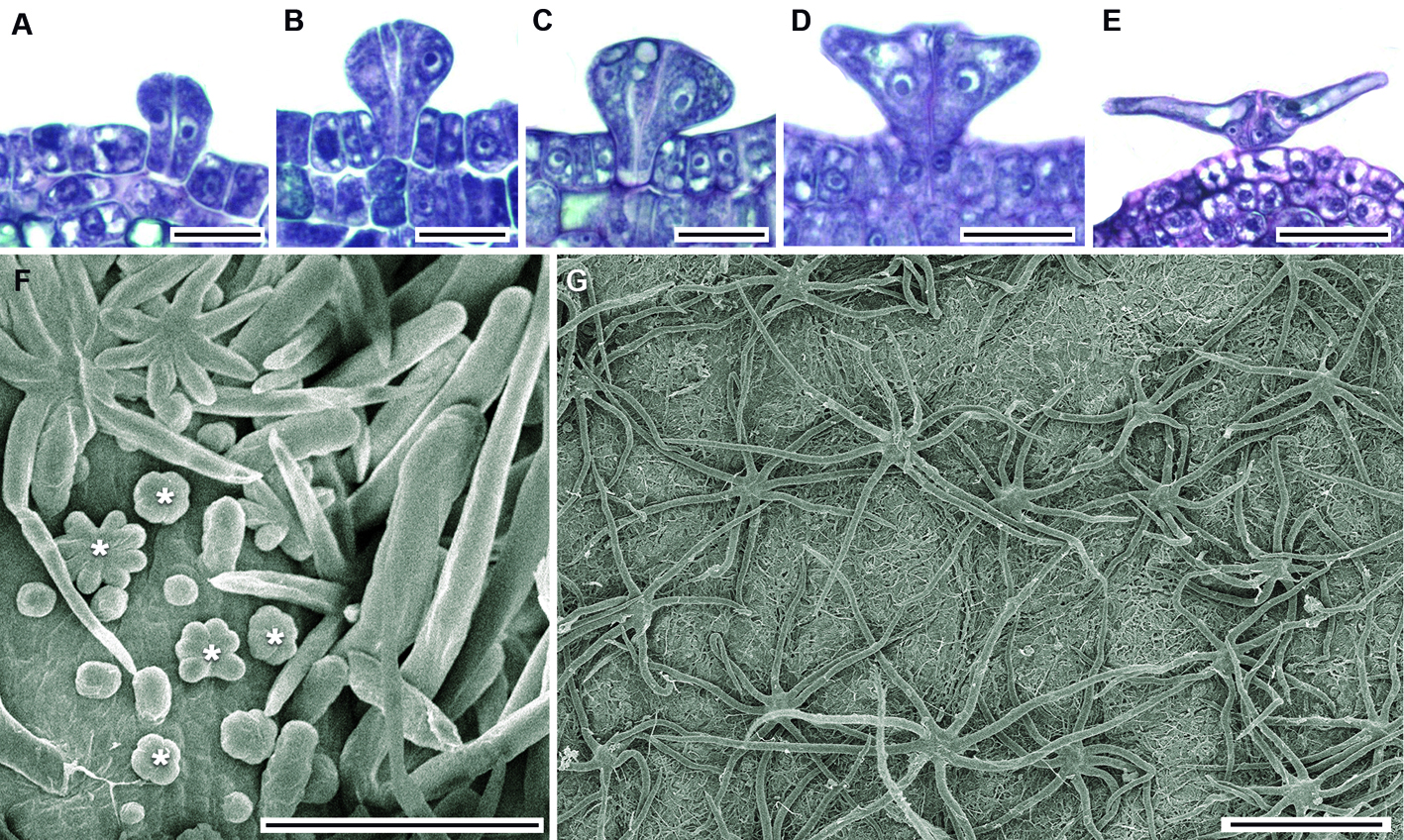
Ontogenia de los tricomas. Tricomas aciculares: inician su desarrollo con el alargamiento de una célula protodérmica que multiplica su tamaño hasta 5 veces y posteriormente agudiza su extremo distal (fig. 3A, B). No se aprecia ningún contenido en su lumen celular, son muy abundantes sobre las venas y alcanzan su tamaño máximo mucho antes que los estrellados (fig. 3C, D) y con la madurez de la hoja muchos de estos tricomas se caen. Tricomas fasciculados: se desarrollan a partir de una división anticlinal de una célula protodérmica, le siguen 2 o más divisiones anticlinales sucesivas, llegando a tener un máximo de 6 brazos. Cada una de las células resultantes se alargan y agudizan diferencialmente (fig. 4A-F), manteniéndose erguidas; las células epidérmicas que se encuentran alrededor de la base del tricoma son más pequeñas (fig. 4G) y se levantan formando una especie de anillo visto en superficie (fig. 4H). Éstos también llegan a caerse con la madurez de la hoja. Tricomas estrellados: su formación inicia con una división anticlinal de una célula protodérmica, posteriormente se suceden varias divisiones anticlinales; el número de divisiones celulares determina el número de brazos (4-8) que conformarán al tricoma maduro (fig. 5A-C), que para ambos sitios fue de 8. En vista superficial se aprecian como lobulaciones (fig. 5F).
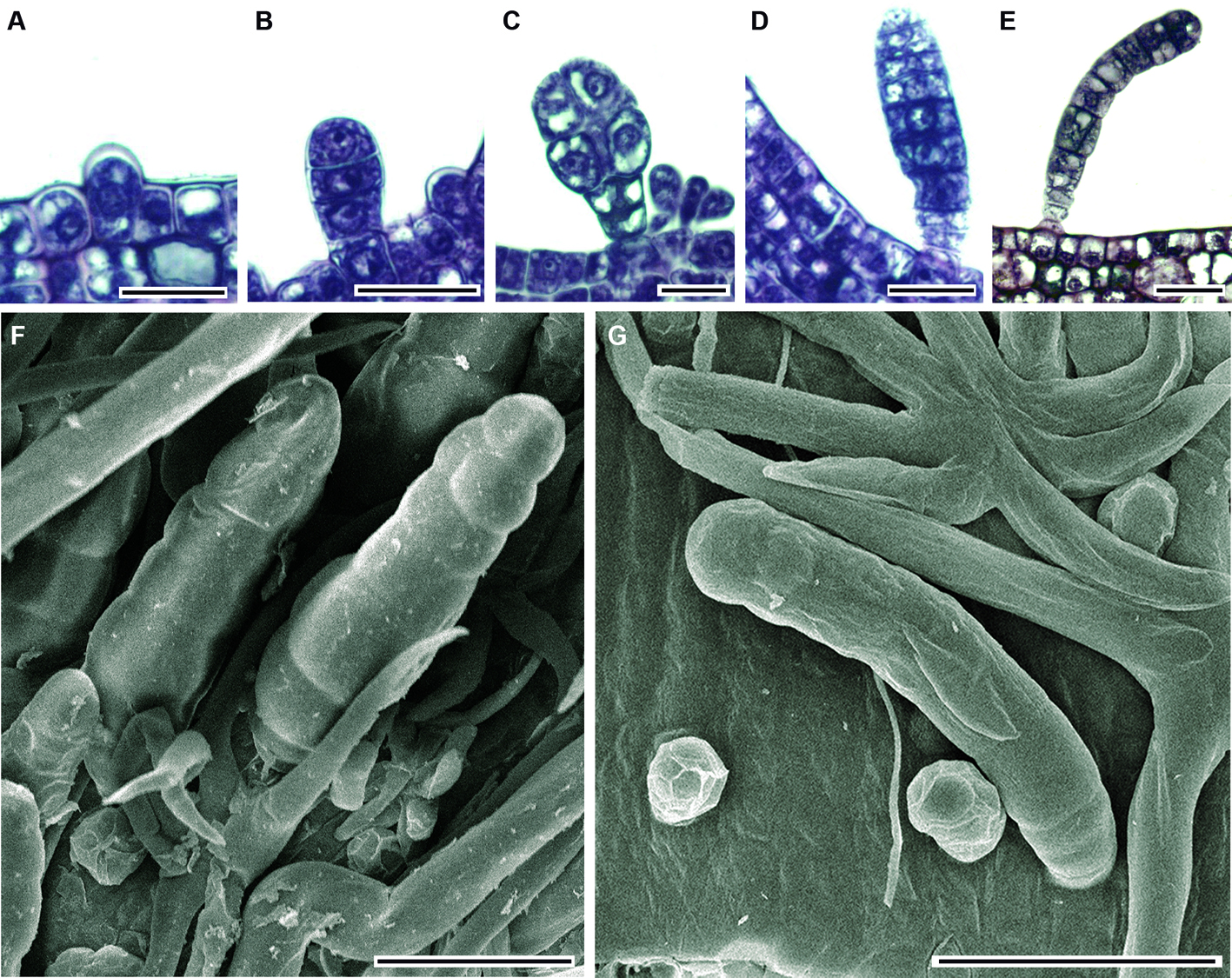
Más tarde cada una de estas células se alarga diferencialmente en forma paralela a la superficie epidérmica y agudiza su ápice. Finalmente, los brazos yacen postrados sobre la lámina (fig. 5D-F); al alcanzar el tricoma la madurez, la longitud de los brazos varía entre 202 y 439 µm con una media de 320 µm; cuando la lámina se expande estos tricomas cubren la región intercostal (fig. 5G). Tricomas glandulares: se encontraron 3 tipos de tricomas glandulares (bulboso, de botella y largo), en todos los casos su desarrollo inicia a partir de una célula protodérmica que se alarga y sobresale de las células protodérmicas adyacentes que la rodean; enseguida se divide periclinalmente (fig. 6A, B).
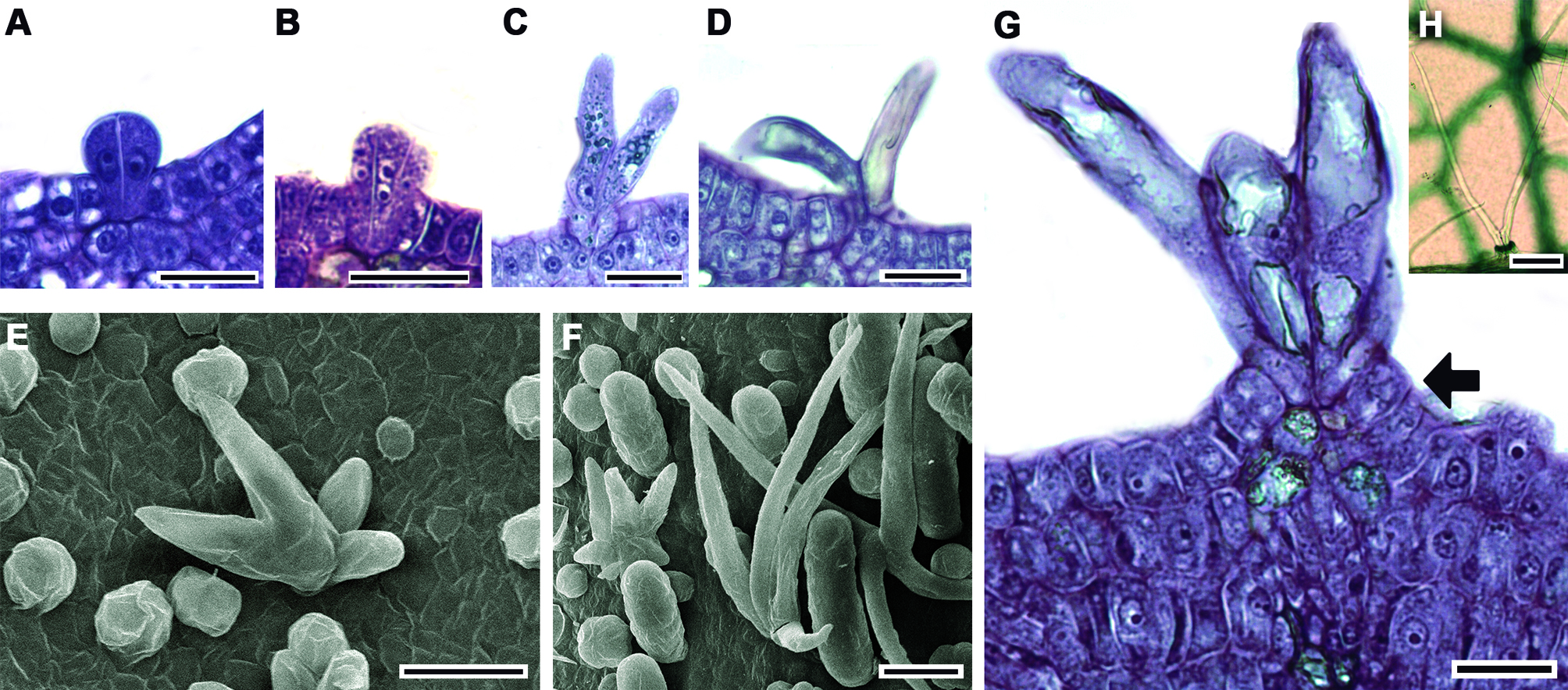
El número y orientación de las divisiones celulares determina la forma del tricoma glandular. El tricoma glandular bulboso inicia con 3 células, las 2 células superiores se dividen de forma anticlinal originando 4 células que posteriormente se ensanchan, mientras tanto la célula basal se divide periclinalmente para formar un pie 3-celular (fig. 6C). En el tricoma glandular con forma de botella, la célula epidérmica inicial se divide periclinalmente una vez, la célula apical resultante experimenta sucesivas divisiones periclinales hasta formar 8 células, al mismo tiempo, la célula basal se alarga; enseguida las 2 células centrales (de la serie de 8 células) se dividen anticlinalmente originando 4 células que se enchanchan; a su vez, la célula apical se divide anticlinalmente y la superficie externa de las células resultantes se redondean (fig. 6D, F). En el tricoma glandular largo, la célula inicial se divide periclinalmente para formar una columna de hasta 11 células; en este estado, la célula apical se redondea (fig. 6E, G). Los tricomas glandulares largos son los primeros en aparecer en la lámina; posteriormente sobre las venas en desarrollo y las regiones intercostales se encuentran los otros 2 tipos de tricomas glandulares que pronto y en las hojas maduras se llegan a colapsar.
Discusión
El tipo de tricoma se define con la primera división de la célula protodérmica o por la ausencia de divisiones celulares, como sucede en los tricomas aciculares, cuya célula epidérmica únicamente se alarga y agudiza su ápice. El número de brazos de los tricomas estrellados y fasciculados también se define por las primeras divisiones anticlinales de la célula protodérmica y la elongación de los brazos se lleva a cabo de manera distintiva para cada uno de ellos. Los tricomas glandulares son más diversos en orientación de las divisiones, así como en el número de células que los conforman. En este trabajo se encontraron 6 tipos de tricomas en las diferentes etapas del desarrollo de las láminas foliares, cuando éstas aún se encuentra en la yema. Además, se evidencia a través de la ontogenia que no hay formas intermedias en los tricomas de Tilia. Todos los tricomas en Tilia tienen origen exclusivo en las células protodérmicas, incluyendo a los tricomas glandulares como en otras familias de angiospermas (Akers et al., 1978; Ascensão y Pais, 1998; Celep et al., 2014; Kolb y Müller, 2004; Nielsen et al., 1991).
Identificación de los tipos de tricomas. La falta de conocimiento sobre todos los tipos de tricomas dentro de las angiospermas vuelve problemática su delimitación (Ramayya, 1962); lo que causa confusión en la terminología y descripción entre los tricomas estrellados y fasciculados. Por ejemplo, Inamdar y Chohan (1969) definen el tricoma estrellado como un apéndice multicelular que tiene 2-8 brazos rodeados por 8-10 células epidérmicas, mientras que Ramayya y Rao (1976) distinguen los tricomas estrellados de los fasciculados porque estos últimos presentan brazos erectos sobre la lámina. Nuestras observaciones son congruentes con Ramayya y Rao (1976), ya que los tricomas fasciculados también tienen los brazos erectos. Además, presentan una serie de células epidérmicas más pequeñas (fig. 4G) alrededor de su base que en superficie parece un anillo (fig. 4H) y es similar a lo descrito por Rendón-Carmona et al. (2006) como un estípite para el género Mortoniodendron Standl. et Steyerm. En contraste, los tricomas estrellados, desde sus primeras etapas de diferenciación son lobulados (fig. 5F) y a medida que los brazos se extienden lo hacen en forma paralela a la lámina conforme ésta se expande (fig. 5) similar a lo que se ha registrado en Mortoniodendron (Rendón-Carmona et al., 2006), por lo que sus brazos son postrados. En Theobroma cacao L., Garcia et al. (2014) observaron tricomas estrellados con un pie y sésiles, en Tilia únicamente encontramos tricomas estrellados sésiles. Distintivamente la longitud (320 µm) y número de brazos (8) de los tricomas estrellados que aquí registramos corresponden a las dimensiones que señala Pigott (2012) para la subespecie en estudio.
Hardin (1990) describe tricomas glandulares bulbosos uniseriados para las especies de Tilia presentes en América. Sin embargo, los únicos tricomas glandulares uniseriados que se observaron en este trabajo son los denominados glandulares largos. Además, también se registraron tricomas glandulares multiseriados en forma de botella, como los descritos por Ragonese (1960) para otra Malvaceae, Hibiscus rosa-sinensis L. Se sabe que los tricomas glandulares tienen un desarrollo excesivamente rápido en los primordios foliares (Duke y Paul, 1993). En el caso de las tilias, los tricomas del tipo glandular largo son los primeros que se desarrollan y se presentaron con mayor abundancia en las yemas colectadas en otoño, mientras el árbol está en latencia. Dicha abundancia puede estar relacionada a la lubricación y protección contra la desecación de la yema, así como contra patógenos y herbívoros (Fahn, 1979; 1990; Kronestedt-Robards y Robards, 1991; Thomas, 1991; Uphof, 1962). Este resultado es contrario a lo encontrado en el género Vitis L., donde los tricomas glandulares aparecen tardíamente en el desarrollo de la lámina una vez que los aciculares se desprenden (Ma et al., 2016).
Los tricomas glandulares largos y los no glandulares aciculares predominan sobre las venas en Tilia. Una vez que la yema se hincha comienza el desarrollo de los otros tricomas aquí descritos, principalmente sobre las venas y los márgenes. Los estrellados son los últimos en completar su desarrollo y cubren las regiones intercostales como también se ha descrito para otras Malvaceae (Gual, 1998; Rendón-Carmona et al., 2006). A partir de nuestras observaciones los tricomas de Tilia podrán codificarse por su número de células como unicelular (0), multicelular (1); por su secreción como glandular (0) y no glandular (1). Los multicelular glandulares como globoso (0), botella (1), largo (2) y los multicelulares no glandular como fascicular (0), estrellado (1). Estas hipótesis de homología podrán incluirse en futuros análisis filogenéticos para Tilia. Con base en los resultados de este trabajo, concluimos que no hay estados intermedios entre los diferentes tipos de tricomas encontrados en el género Tilia, que con excepción de los tricomas estrellados, los otros pueden o no encontrarse en las láminas maduras. Caracterizar el tamaño de los brazos en los tricomas estrellados, como ya señaló Pigott (2012), podría contribuir a la delimitación de las especies con base en los atributos de los tricomas estrellados y no en la ausencia-presencia de los otros tipos.
Agradecimientos
Se agradecen las facilidades del Instituto de Biología para desarrollar esta investigación, así como a Jorge Plancarte Gómez por colaborar en la recolecta en Cuetzalan, a Mayte Jiménez-Noriega por supervisar la microtecnia de inclusión en glicol metilcrilato, a Berenit Mendoza-Garfías por ayudar en el microscopio electrónico de barrido y a Julio César Montero Rojas por elaborar las figuras digitales. Se agradecen los comentarios de un revisor anónimo y de Mahinda Martínez.
Referencias
Agati, G., Brunetti, C., Di Ferdinando, M., Ferrini, F., Pollastri, S. y Tattini, M. (2013). Functional roles of flavonoids in photoprotection: new evidence, lessons from the past. Plant Physiology and Biochemistry, 72, 35–45.
Akers, C. P., Weybrew, J. A. y Long, R. C. (1978). Ultrastructure of glandular trichomes of leaves of Nicotiana tabacum L., cv Xanthi. American Journal of Botany, 65, 282–292.
Ascensão, L. y Pais, M. S. (1998). The leaf capitate trichomes of Leonotis leonurus: histochemistry, ultrastructure and secretion. Annals of Botany, 81, 263–271.
Bickford, C. P. (2016). Ecophysiology of leaf trichomes. Functional Plant Biology, 43, 807–814.
Carlquist, S. (1961). Comparative plant anatomy. New York: Holt, Rinehart, Winston.
Carvalho-Sobrinho, J. G. D., Santos, F. D. A. R. D. y Queiroz, L. P. D. (2009). Morfologia dos tricomas das pétalas de espécies de Pseudobombax Dugand (Malvaceae, Bombacoideae) e seu significado taxonômico. Acta Botanica Brasilica, 23, 929–934.
Celep, F., Kahraman, A., Atalay, Z. y Doğan, M. (2014). Morphology, anatomy, palynology, mericarp and trichome micromorphology of the rediscovered Turkish endemic Salvia quezelii (Lamiaceae) and their taxonomic implications. Plant Systematics and Evolution, 300, 1945–1958.
Cowan, J. M. (1950). The Rhododendron leaf; a study of epidermal appendages. London: Oxford and Boyd.
Duke, S. O. y Paul, R. N. (1993). Development and fine structure of the glandular trichomes of Artemisia annua L. International Journal of Plant Sciences, 154, 107–118.
Fahn, A. (1979). Secretory tissues in plants. London: Academic Press.
Fahn, A. (1990). Plant anatomy. Oxford: Pergamon Press.
Gama, T. S. S., Demarco, D. y Aguiar-Dias, A. C. A. (2013). Ontogeny, histochemistry, and structure of the glandular trichomes in Bignonia aequinoctialis (Bignoniaceae). Brazilian Journal of Botany, 36, 291–297.
Garcia, T. B., Potiguara, R. C. D. V., Kikuchi, T. Y. S., Demarco, D. y Aguiar-Dias, A. C. A. D. (2014). Leaf anatomical features of three Theobroma species (Malvaceae s.l.) native to the Brazilian Amazon. Acta Amazonica, 44, 291–300.
Gual, M. (1998). La familia Tiliaceae Juss. en el estado de Guerrero (Tesis de maestría). Facultad de Ciencias, Universidad Nacional Autónoma de México, México D.F.
Hardin, J. (1979). Patterns of variation in foliar trichomes of eastern North American Quercus. American Journal of Botany, 6, 576–585.
Hardin, J. (1990). Variation patterns and recognition of varieties of Tilia americana s.l. Systematic Botany, 15, 33–48.
Inamdar, J. A. (1967). Studies on the trichomes of some Oleaceae, structure and ontogeny. Proceedings of the Indian Academy of Sciences-Section B, 66, 164–177.
Inamdar, J. A. y Chohan, A. J. (1969). Epidermal structure and stomatal development in some Malvaceae and Bombacaceae. Annals of Botany, 33, 865–878.
Inamdar, J. A., Bhat, R. B. y Rao, T. R. (1983). Structure, ontogeny, classification, and taxonomic significance of trichomes in Malvales. Korean Journal of Botany, 26, 151–160.
Jones, G. N. (1968). Taxonomy of American species of Lindens (Tilia). Illinois Biological Monographs 39. Champaign-Urbana, Illinois: University of Illinois Press.
Judd, W. S., Campbell, C. S., Kellog, E. A., Stevens, P. F. y Donoghue, M. J. (2007). Plant systematics: a phylogenetic approach, 3a Ed. Sunderland, Massachusetts: Sinauer.
Kim, H. J., Seo, E., Kim, J. H., Cheong, H., Kang B. C. y Choi, D. (2012). Morphological classification of trichomes associated with possible biotic stress resistance in the genus Capsicum. Plant Pathologist Journal, 28, 107–113.
Kolb, D. y Müller, M. (2004). Light, conventional and environmental scanning electron microscopy of the trichomes of Cucurbita pepo subsp. pepo var. styriaca and histochemistry of glandular secretory products. Annals of Botany, 94, 515–526.
Kronestedt-Robards, E. y Robards, A. W. (1991). Exocytosis in gland cells. En C. R. Hawes, J. O. D. Coleman y D. E. Evans (Eds.), Endocytosis, exocytosis and vesicle traffic in plants (pp. 199–232). Cambridge: Cambridge University Press.
Levin, D. A. (1973). The role of trichomes in plant defense. Quarterly Review of Biology, 48, 3–15.
Ma, Z. Y., Wen, J., Ickert-Bond, S. M., Chen, L. Q. y Liu, X. Q. (2016). Morphology, structure, and ontogeny of trichomes of the grape genus (Vitis, Vitaceae). Frontiers in Plant Science, 7, 704.
Martínez-Gordillo, M. y Espinosa-Matías, S. (2005). Tricomas foliares de Croton sección Barhamia (Euphorbiaceae). Acta Botanica Mexicana, 72, 39–51.
McCarthy, D. M. y Mason-Gamer, R. J. (2016). Chloroplast DNA-based phylogeography of Tilia americana (Malvaceae). Systematic Botany, 41, 865–880.
Nielsen, M. T., Akers, C. P., Järlfors, U. E., Wagner, G. J. y Berger, S. (1991). Comparative ultrastructural features of secreting and nonsecreting glandular trichomes of two genotypes of Nicotiana tabacum L. Botanical Gazette, 152, 13–22.
Nogueira, A., Ottra, J. H. L., Guimarães, E., Rodrigues-Machado, S. y Lohmann, L. G. (2013). Trichome structure and evolution in Neotropical lianas. Annals of Botany, 112, 1331–1350.
Payne, W. (1978). A glossary of plant hair terminology. Brittonia, 30, 239–255.
Pigott, D. (2012). Lime-trees and basswoods: a biological monograph of the genus Tilia. Cambridge: Cambridge University Press.
Ragonese, A. M. (1960). Ontogenia de los distintos tipos de tricomas de Hibiscus rosa-sinensis L. (Malvaceae). Darwiniana, 12, 58–66.
Rajput, M. T. M., Carolin, R. C. y Tahir, S. S. (1985). The indumentum of genus Dampiera R. Br. (Goodeniaceae). Pakistan Journal of Botany, 17, 181–194.
Ramayya, N. (1962). Studies on the trichomes of some Compositae. II. Phylogeny and classification. Nelumbo, 4, 189–192.
Ramayya, N. (1972). Classification and phylogeny of the trichomes of angiosperms. En A. K. M. Ghouse y M. Yunus (Eds.), Research trends in plant anatomy (pp. 91–102). Bombay: McGraw-Hill Pub.
Ramayya, N. y Rao, S. R. (1976). Morphology phylesis and biology of the peltate scale, stellate and tufted hairs in some Malvaceae. Journal of the Indian Botanical Society, 55, 75–79.
Rao, S. R. (1990). Trichome ontogenesis in some Tiliaceae. Beiträge zur Biologie der Pflanzen, 65, 363–375.
Redonda-Martínez, R., Villaseñor, J. L. y Terrazas, T. (2012). Trichome diversity in the Vernonieae (Asteraceae) of Mexico I: Vernonanthura and Vernonia (Vernoniinae). The Journal of the Torrey Botanical Society, 139, 235–247.
Rendón-Carmona, N., Ishiki-Ishihara, M., Terrazas, T. y Nieto-López, M. G. (2006). Indumento y tricomas en la caracterización de un grupo de nueve especies del género Mortoniodendron (Tiliaceae). Revista Mexicana de Biodiversidad, 77, 169–176.
Ruzin, S. E. (1999). Plant microtechnique and microscopy. New York: Oxford University Press.
Shaheen, N. I. G. H. A. T., Ajab, M., Yasmin, G. y Hayat, M. Q. (2009). Diversity of foliar trichomes and their systematic relevance in the genus Hibiscus (Malvaceae). International Journal of Agriculture and Biology, 11, 279–284.
Theobald, W. L., Krahnlik, J. L. y Rollins, R. C. (1980). Trichome description and classification. En C. R. Metcalfe y L. Chalk (Eds.), Anatomy of dicotyledons (pp. 40–53). Oxford: Clarendon Press.
Thomas, V. (1991). Structural, functional and phylogenetic aspects of the colleter. Annals of Botany, 68, 287–305.
Uphof, J. C. T. (1962). Plant hairs. En W. Zimmermann y P. G. Ozenda (Eds.), Handbuch der pflanzenanatomie (pp.1–292). Berlin: Gebrüder Borntraeger.
Zarlavsky, G. E. (2014). Histología vegetal: técnicas simples y complejas. Buenos Aires: Sociedad Argentina de Botánica.


