
Claudio Delgadillo-Moya, Ana Paola Peña-Retes
Universidad Nacional Autónoma de México, Instituto de Biología, Tercer Circuito, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
*Corresponding author: moya@unam.mx (C. Delgadillo-Moya)
Received: 10 October 2022; accepted: 03 February 2023
Resumen
Pilotrichella, a genus of epiphytic mosses comprising 6 species and 1 form, is represented in Mexico by P. flexilis (Hedw.) Aongstr., P. flexilis fo. nudiramulosa (Müll. Hal.) B. H. Allen & Magill, P. mauiensis (Sull.) A. Jaeger, and P. reesei B. H. Allen & Magill. The last species is known from a single specimen. The Peruvian P. vermiformis B. H. Allen & Magill is reported here as new to Mexico from Hidalgo, Veracruz, and Oaxaca. In total, 260 specimens deposited at MEXU served to illustrate, map, and discuss the basic morphology of the local taxa.
Keywords: Disjunct; Tropical mosses
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Pilotrichella (Bryophyta: Lembophyllaceae) en México
Abstract
Pilotrichella es un género de musgos epifitos que comprende 6 especies y 1 forma; en México está representado por P. flexilis (Hedw.) Aongstr., P. flexilis fo. nudiramulosa (Müll. Hal.) B. H. Allen & Magill, P. mauiensis (Sull.) A. Jaeger y P. reesei B. H. Allen & Magill. La última especie se conoce en México por un solo ejemplar. En este estudio se cita a P. vermiformis B. H. Allen & Magill como nueva para la flora mexicana a partir de material proveniente de Hidalgo, Veracruz y Oaxaca. En total, se estudiaron 260 ejemplares depositados en MEXU para ilustrar, situar y discutir la morfología de los taxones locales.
Palabras clave: Disyunto; Musgos tropicales
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
The genus Pilotrichella (Müll. Hal.) Besch. is a small essentially tropical American-African genus originally placed in the Meteoriaceae. It was transferred to the Lembophyllaceae by Buck (1994), a view supported by Buck and Goffinet (2000) and Allen and Magill (2003) citing morphological arguments. Prior to the last-mentioned authors, only P. flexilis (Hedw.) Aongstr. and P. rigida (Müll. Hal.) Besch. were recognized as Mexican taxa (Sharp et al., 1994). However, further systematic treatment of the genus Pilotrichella excluded various costate or ecostate species with spirally ranked leaves and placed them in Orthostichella (Allen & Magill, 2007). The generic revision by Allen and Magill (2003) described and illustrated 6 species; a nomenclatural history was given there and in Allen (2010). The former recognized Pilotrichella flexilis, P. flexilis fo. nudiramulosa (Müll. Hal.) B. H. Allen & Magill, P. mauiensis (Sull.) A. Jaeger, and P. reesei B. H. Allen & Magill as present in Mexico. Recent work extended the distribution of the Peruvian P. vermiformis B. H. Allen & Magill to Mexico (see below). The Mexican distribution of Pilotrichella, while supported by herbarium specimens, has not been mapped to include areas known from local collections. This study aims to revise specimens of Pilotrichella at MEXU following current concepts to update the known range of the genus in Mexico.
Materials and methods
The Bryophyte Collection at MEXU contains about 260 specimens of Pilotrichella from Mexican states, which were examined to validate previous identification or to update their name according to Allen and Magill’s (2003) taxonomic concepts. These specimens represent areas visited by bryologists and other collectors since 1904 including recent fieldwork collections by the authors; although specimens cited in the literature and reported in other herbaria were included in this study to further clarify the generic range of Pilotrichella in Mexico, they were not used in mapping local distribution of species and no effort was made to examine them directly in order to confirm their identity. The geographical information for the maps in the present contribution was obtained from specimens that conform with current taxonomic concepts. Arc.Map 10.1 software was used to produce the maps.
Results
Although the original labels indicated that all Mexican specimens at MEXU belonged in Pilotrichella flexilis, upon re-identification the set was found to comprise P. flexilis (129 specimens), P. flexilis fo. nudiramulosa (13 specimens), P. mauiensis (112 specimens), and P. vermiformis (7 specimens) according to current concepts. No collections of Pilotrichella reesei were
observed.
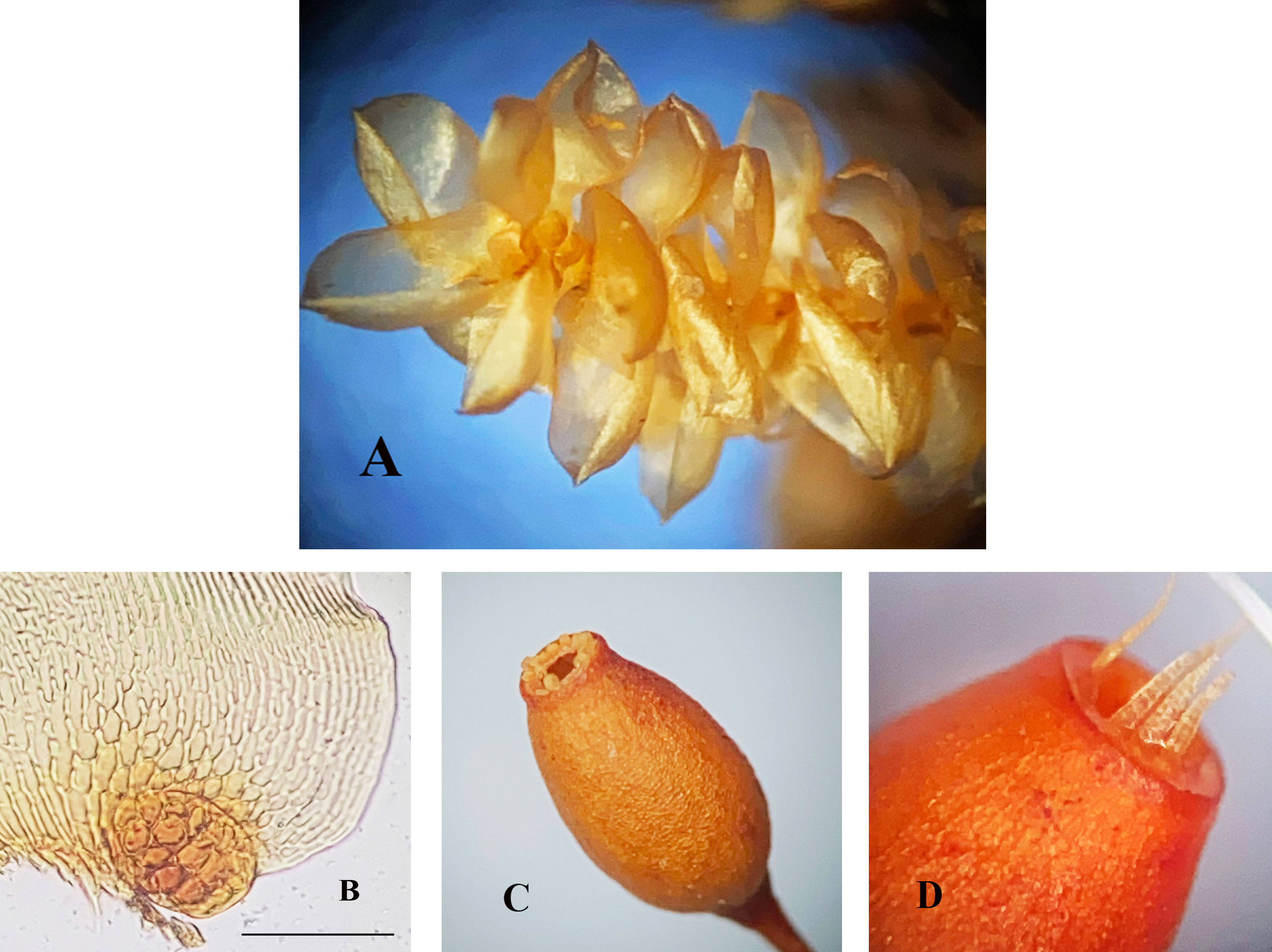
Distinguishing morphological features are given in Table 1. Others are illustrated as indicated, e.g., the primary stems in P. flexilis that produce long, pendent secondary stems with short branches (Fig. 1A, B); the leaves are comparatively large with reflexed apices, involute laminae, and conspicuous alar cells in a convex group (Fig. 1C-E). Stems of fo. nudiramulosa (Fig. 2) are usually smaller than those of P. flexilis and have deciduous leaves and nude branches. In P. mauiensis the stems are less robust than those of P. flexilis, the branch tips may be attenuate (Fig. 3A), but the lateral branches may be rather robust (Fig. 3B). Although the leaves may be similar in both taxa, in P. mauiensis the cuspidate apices are straight (Fig. 3E), not recurved, and microphyllous branches (Fig. 3C) or nude branches may be present (Fig. 3D). In P. vermiformis, on the other hand, the stems are less robust than in other species of the genus, the spoon-shaped leaves have margins nearly erect when wet (Fig. 4A), the apices may be broadly rounded, mucronate, straight or reflexed, and the alar cells form a small but distinct convex group (Fig. 4B).
Nude branches are a common feature of P. flexilis fo. nudiramulosa (Fig. 2); its leaves are rounded, shortly cuspidate, but no microphyllous branches are evident as in P. mauiensis. However, microphyllous branches are sometimes present in the robust stems of P. flexilis, e.g., in Sharp 663a (MEXU).
The distinction between P. flexilis and P. mauiensis is sometimes tenuous. Specimens from the same area, collected in the same habitat, bearing consecutive collecting numbers, are similar morphologically but key characters place them in different species. For example, Delgadillo 1046 (MEXU) shows comparatively thin, attenuate stems, and cuspidate leaves with straight apices as in P. mauiensis. By comparison, in Delgadillo 1047 (MEXU) the generally robust stems with obtuse-rounded, apiculate, distinctly reflexed leaf apices identify it as P. flexilis.
Table 1
Distinguishing features of Pilotrichella species in Mexico.
| Character states | flex | nudi | maui | verm | rees* |
| Robust stems | × | × | |||
| Rounded stem and branch tips | × | × | |||
| Leaf apex reflexed, rounded | × | × | |||
| Leaf apex cuspidate, straight, not reflexed | × | ||||
| Leaf margin involute when dry | × | × | × | × | |
| Leaf margin erect | × | ||||
| Attenuate branches | × | × | |||
| Nude branches, deciduous leaves | × | × | |||
| Microphyllous branches frequent | × | ||||
| Alar cell group small, distinct | × | ||||
| Alar cell group large, well-developed | × |
* = After Allen and Magill (2003).


Shared character states should receive careful evaluation. Deciduous-leaved branches are characteristic of P. flexilis fo. nudiramulosa, but are also present in specimens of P. mauiensis (and P. vermiformis); round-obtuse leaf apices may also occur in parts of stems P. mauiensis although they are the norm in P. flexilis. Peña 511 (MEXU) is a small form of P. mauiensis with well-developed erect cuspidate leaf apices where some leaves resemble those of P. vermiformis.
Pilotrichella vermiformis B. H. Allen & Magill was described as a new species from Peru (Allen & Magill 2003). A specimen from Veracruz (Juárez 958A) identified by Allen as a member of this taxon shows the peculiar spoon-shaped leaves with erect, hardly involute laminae and strongly differentiated alar cells. A single sporophyte was found in this specimen, but its morphology was not known at the time of the original description of the species; the seta is ca. 10 mm long, the ovoid capsule is smooth-walled and the single peristome is erect when wet thus opening the capsule mouth (Fig. 4C, D). Upon drying the teeth are inflexed and apparently leave the mouth fully open. Similar peristomial movement was reported by Patterson (1953) in Forsstroemia ohioensis [= F. producta (Hornsch.) Paris].
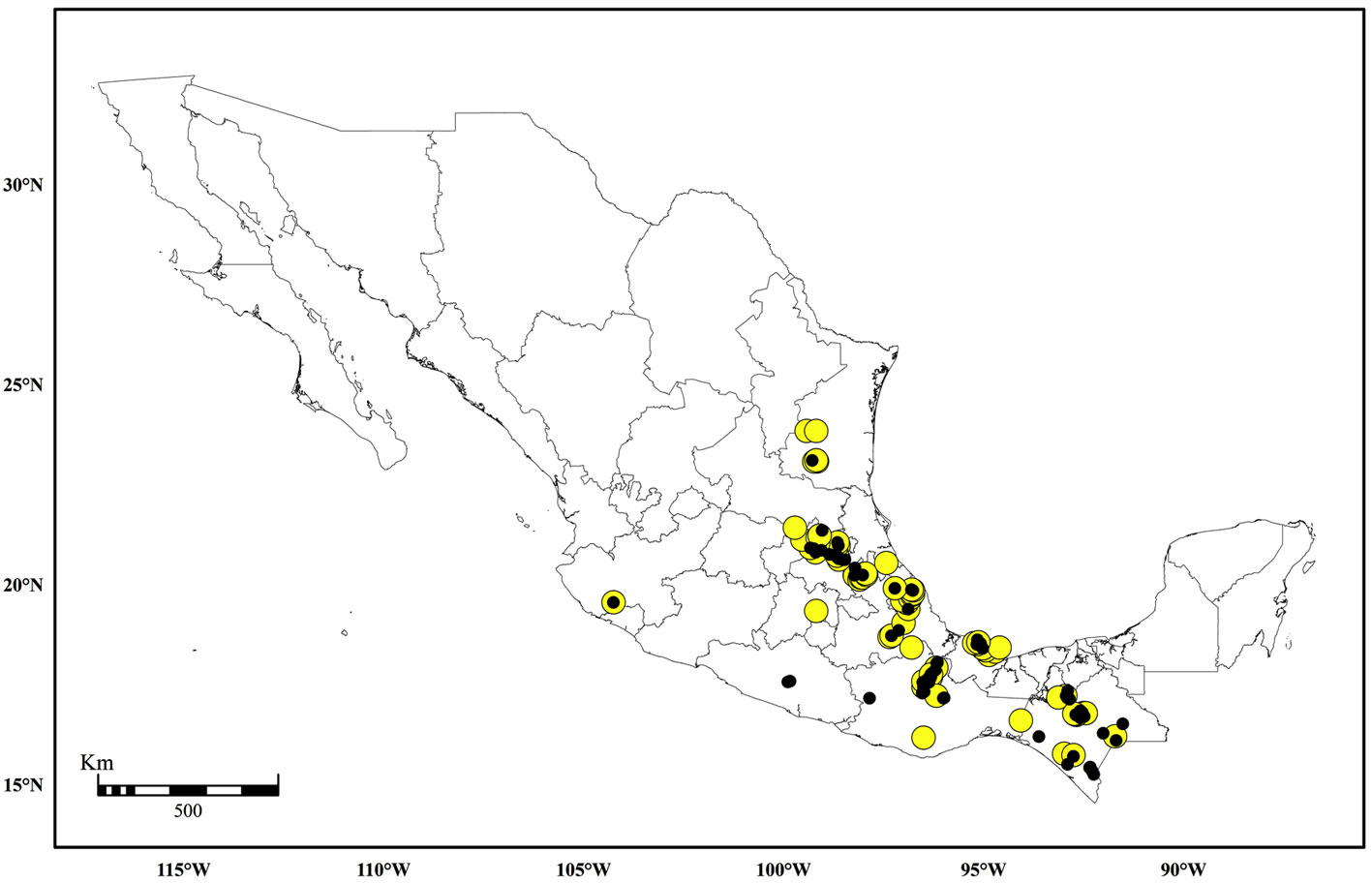
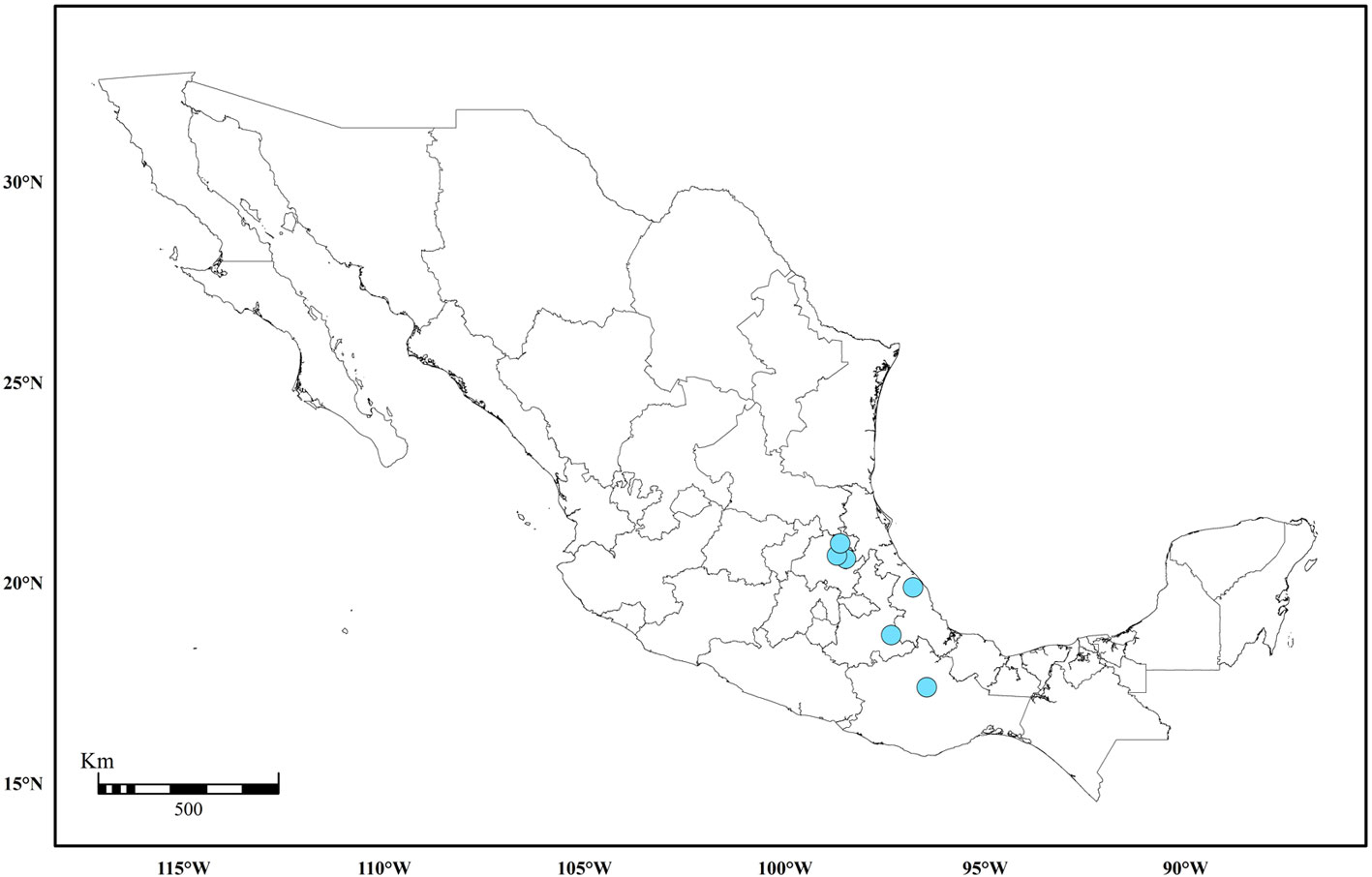
The map in Figure 5 shows the general distribution of Pilotrichella in Mexico. The collecting sites are localities in eastern and southern Mexico with deciduous, evergreen or pine-oak forests where P. flexilis and P. mauiensis are most frequent. A westward locality occurs in the state of Jalisco. The collecting localities for other Pilotrichella taxa also seem part of these forests except for the habitat of P. reesei which is known from a single record in Chiapas (Allen & Magill 2003). A collection record reported in the literature indicates that P. flexilis occurs in Baja California (Allen & Magill, 2003; Crum, 1951). However, the specimen (Brandegee 11, NY) bears no locality information so that the distribution of the genus cannot be validated in Lower California. In any event this locality is well outside the generic geographical range in Mexico. Brandegee visited localities in southern Mexico (Moran, 1952), but there is no indication that he would have obtained Pilotrichella samples there. Figure 6 shows the known distribution of P. vermiformis in areas of the states of Hidalgo, Veracruz, and Oaxaca that are also part of the general distribution of the genus.
Discussion
Because of its characteristic morphology and comparatively large size, Pilotrichella has been frequently collected and its generic range may be established with ease. However, as there are few characters restricted to a single taxon or they have intermediate values, their taxonomic separation is sometimes obscured, as for example, to mark a clear-cut distinction between P. flexilis and P. mauiensis. For practical reasons identification in this group should use a combination of character states and determination of habitat preferences during field work.

Understanding phenotypic variation in terms of environmental response would help in making good taxonomic evaluations in Pilotrichella and its significance in defining their geographical ranges. Thus, for instance, do mucronate or cuspidate leaf apices or the presence of nude or attenuate branches represent stable features of the species or an ecological response to habitat differences? Allen (2010) refers to variation exhibited by P. mauiensis throughout its range, especially with regard to cuspid length, but he offers no explanation as to its cause. Nude branches in P. flexilis fo. nudiramulosa are also present in P. mauiensis and might be related to microclimatic variables or drier habitats along their range. The geographic and taxonomic implications of the morphological variation need further analysis, especially in species that share other features of leaf apices and branch morphology or in such species as P. reesei whose “distinctive alar cell development” (Allen & Magill, 2003) is perhaps its major distinguishing character. A single specimen of this taxon is known from Mexico.
The known distribution of Pilotrichella seems mostly associated with moist forests in tropical and subtropical areas of Mexico. Comparison against the distribution of the mesophyllous forests indicates that the genus is a frequent epiphyte growing on deciduous trees; its range extends from Tamaulipas in the north southwards to Chiapas, Guerrero, and Oaxaca (Fig. 5). Lack of collection data for Baja California does not support its past or present existence in Mexico; the presumed occurrence of mesophyllous forests in extreme northern localities in nearby Sinaloa and Durango (Gual-Díaz & Rendón-Correa, 2017) are also devoid of known records. Southwards, the range comprises stations in Central and South America, i.e., Pilotrichella is an entirely tropical taxon that is part of a flora that intermixes with temperate vascular plant species (cf. Miranda & Sharp, 1950) in the mesophyllous forests of eastern Mexico since Tertiary times (cf. Graham, 1993; Martin & Harrell, 1957; Rzedowski & Palacios-Chávez, 1977). Its range in Africa speaks for an ancient distribution and perhaps a Gondwanan relationship of the group.
Pilotrichella is far from being well understood. Morphologically, it offers numerous expressions that may be special adaptations as an epiphyte, but which require multiple explanations. The narrow pitted thick-walled cells that make up most of the leaf lamina ending at base in an excavated thick-walled alar group are perhaps associated with water relationships in long stems with no conducting cells. The peculiar behavior of the peristome observed in the exserted sporophytes is also a matter worth investigating. The capsule mouth seems widely open both in moist and dry conditions (Fig. 4C, D), and dispersal may occur at any time after operculum dehiscence, but the spores being comparatively large in the genus (18-28 or 34-64 µm; Allen & Magill, 2003) may not disperse easily under cloudy or rainy weather despite the exserted capsule and the pendulous growth form.
Acknowledgements
We are indebted to Bruce Allen, Missouri Botanical Garden, for the original identification of a specimen of Pilotrichella vemiformis and for review of a draft of the manuscript. Specimens collected by Enrique Hernández-Rodríguez in Oaxaca included a specimen of P. vermiformis. Two anonymous reviewers contributed helpful comments to the manuscript.
References
Allen, B. (2010). Pilotrichella (Müll. Hal.) Besch. Moss Flora of Central America. Part 3. Anomodontaceae – Symphyodontaceae. Monographs in Systematic Botany from the Missouri Botanical Garden, 117, 173–181.
Allen, B., & Magill, R. E. (2003). A revision of Pilotrichella (Lembophyllaceae: Musci). Acta Academiae Paedagogicae Agriensis, Sectio Biologiae, 24, 43–83.
Allen, B., & Magill, R. E. (2007). A revision of Orthostichella (Neckeraceae). The Bryologist, 110, 1–45. https://doi.org/10.1639/0007-2745(2007)110[1:aroon]2.0.co;2
Buck, W. R. (1994). A new attempt at understanding the Meteoriaceae. Journal of the Hattori Botanical Laboratory, 75, 51–72. https://doi.org/10.18968/jhbl.75.0_51
Buck, W. R., & Goffinet, B. (2000). Morphology and classification of mosses. In A. J. Shaw, & B. Goffinet (Eds.). Bryophyte Biology (pp. 71–123). Cambridge: Cambridge University Press. https://doi.org/10.1017/cbo9781139171304.004
Crum, H. A. (1951). The Appalachian-Ozarkian element in the moss flora of Mexico with a check-list of all known Mexican mosses (Ph. D. Thesis). University of Michigan. Ann Arbor.
Graham, A. (1993). Historical factors and biological diversity in Mexico. In T. P. Ramamoorty, R. Bye, A. Lot, & J. Fa (Eds.), Biological diversity of Mexico. Origins and distribution (pp.109–127). New York: Oxford University Press.
Gual-Díaz, M., & Rendón-Correa, A. (2017). Los bosques mesó-
filos de montaña de México. Agro-Productividad, 10, 3–9.
Martin, P. S., & Harrell, B. E. (1957). The Pleistocene history of temperate biotas in Mexico and eastern United States. Ecology, 38, 468–480. https://doi.org/10.2307/1929892
Miranda, F., & Sharp, A. J. (1950). Characteristics of the vegetation in certain temperate regions of eastern Mexico. Ecology, 31, 313–333. https://doi.org/10.2307/1931489
Moran, R. (1952). The Mexican itineraries of T. S. Brandegee. Madroño, 11, 221–252.
Rzedowski J., & Palacios-Chávez, R. (1977). El bosque de Engelhardtia (Oreomunnea) mexicana en la región de la Chinantla (Oaxaca, México): una reliquia del Cenozoico. Boletín de la Sociedad Botánica de México, 36, 93–123. https://doi.org/10.17129/botsci.1161
Patterson, P. M. (1953). The aberrant behavior of the peristome teeth of certain mosses. The Bryologist, 56, 157–159. https://doi.org/10.2307/3240111
Sharp, A. J., Crum, H., & Eckel, P. M. (1994). The moss flora of Mexico. Memoirs of the New York Botanical Garden. No. 69. New York, USA.


