
Marco Violante-Huerta a, *, Laura Sanvicente-Añorve b, Aurora Marrón-Becerra a, Elia Lemus-Santana c, César Flores-Coto b
a Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Circuito Exterior s/n, Ciudad Universitaria, 04510 Ciudad de México, Mexico
b Departamento de Ecología y Biodiversidad Acuática, Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Circuito Exterior s/n, Ciudad Universitaria, 04510 Ciudad de México, Mexico
c Departamento de Biología Comparada, Facultad de Ciencias, Universidad Nacional Autónoma de México, Circuito Exterior s/n, Ciudad Universitaria, 04510 Ciudad de México, Mexico
*Corresponding author: marco_violante@hotmail.com (M. Violante-Huerta)
Received: 5 March 2019; accepted: 5 May 2020
Abstract
Within the planktonic Amphipoda, the non-hyperiid species have been scarcely studied in the southern Gulf of Mexico. Zooplankton samples were obtained over a grid of 25 oceanographic stations in the southwestern area of the gulf using a stratified opening-closing plankton net system in epi- and mesopelagic zones (0-1,000 m depth). Sixty-nine individuals of non-hyperiid planktonic amphipods were obtained. Their taxonomic identification indicate the presence of 3 species of Amphilochidea, Cyphocaris anonyx Boeck, 1871, C. challengeri Stebbing, 1888, Eusiropsis riisei Stebbing, 1897, and 1 species of Senticaudata, Stenopleura atlantica Stebbing, 1888, representing the first records of these species in the region. A taxonomic account and information on their horizontal and vertical distribution in the studied area are provided. The presence of eyes in E. riisei is for the first time reported. An identification key for 7 species of Amphilochidea and Senticaudata recorded in the Gulf of Mexico is provided.
Keywords: Mesopelagic zone; Pelagic amphipods; Stenopleura atlantica; Stratified sampling; Vertical distribution; Western Atlantic Ocean
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Primeros registros de anfípodos planctónicos no hiperídeos (Amphilochidea y Senticaudata) de aguas profundas del suroeste del golfo de México, con una clave de identificación
Resumen
Entre los Amphipoda planctónicos, las especies no hiperídeas del sur del golfo de México han sido escasamente estudiadas. Se obtuvieron muestras de zooplancton sobre una red de 25 estaciones oceanográficas ubicadas en el suroeste del golfo utilizando un sistema de redes planctónicas estratificadas de apertura y cierre en zonas epi- y mesopelágicas (0-1,000 m de profundidad). Se obtuvieron 69 individuos de anfípodos plantónicos no hiperídeos. Los resultados de su identificación taxonómica indican la presencia de 3 especies de Amphilochidea, Cyphocaris anonyx Boeck, 1871, C. challengeri Stebbing, 1888, Eusiropsis riisei Stebbing, 1897, y 1 especie de Senticaudata, Stenopleura atlantica Stebbing, 1888, lo cual representa sus primeros registros en la región. Se presenta un reporte taxonómico de las especies y se proporciona información sobre su distribución horizontal y vertical en el área de estudio. Se reporta por primera vez la presencia de ojos en E. riisei. Se proporciona una clave de identificación de 7 especies de Amphilochidea y Senticaudata registradas en el golfo de México.
Palabras clave: Zona mesopelágica; Anfípodos pelágicos; Stenopleura atlantica; Muestreo estratificado; Distribución vertical; Océano Atlántico occidental
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introduction
Among the peracarid crustaceans, the order Amphipoda Latreille, 1816 is the most abundant and has the greatest species richness (LeCroy et al., 2009); this order includes 6 suborders (Lowry & Myers, 2017). The amphipods inhabit mainly benthic systems and have been mostly studied in epicontinental aquatic ponds and in coral reefs, coastal lagoons, mangroves, interacting with other benthic or pelagic organisms (Bowman, 1955; LeCroy et al., 2009; Lowry & Stoddart, 1997; Marrón-Becerra et al., 2018; Thomas & Barnard, 1992; Violante-Huerta, 2018). In planktonic systems, amphipods can be free-living or associated with gelatinous organisms. Some species have been recognized as active predators of other zooplankters (Vinogradov et al., 1996).
Within the planktonic Amphipoda, the non-hyperiid species have been scarcely studied because most of them inhabit deep waters. Generally, they show a broad distribution in the water column. Some species inhabit the epi- and mesopelagic zones, and others are exclusively found in the bathypelagic or hadal zones (Hughes & Lowry, 2015; Lowry & Stoddart, 1997; Thurston, 1976; Vinogradov, 1999). Knowledge of the vertical distribution of non-hyperiid planktonic amphipods is scarce. In the Atlantic Ocean, the last study was published by Thurston (1976) upon stratified samples analyzing the diel vertical migration and distribution of 8 non-hyperiid species down to 950 m depth.
In the Gulf of Mexico, studies concerning planktonic amphipods are mainly referred to surface waters (Barnard, 1972; Barnard & Thomas, 1989; Gasca, 2003, 2004; Gasca et al., 2009), with few studies in deep waters (Lowry & Stoddart, 1997; Shoemaker, 1956). In this gulf, 2 suborders of planktonic Amphipoda have been recorded: Amphilochidea Boeck, 1871 with 3 families and 3 species, the Cyphocarididae with Cyphocaris tunicola Lowry & Stoddart, 1997, the Eurytheneidae with Eurythenes obesus (Chevreux, 1905), and the Synopiidae with Synopia ultramarina Dana, 1853, and Hyperiidea H. Milne Edwards, 1830 with about 100 species (Barnard, 1972; Barnard & Thomas, 1989; LeCroy et al., 2009; Lowry & Stoddart, 1997; Shoemaker, 1956). However, there are no studies on non-hyperiid planktonic amphipods addressing the mesopelagic zone in Mexico. In this work we report the first records of 4 species of non-hyperiid planktonic amphipods of the suborders Amphilochidea and Senticaudata Lowry & Myers, 2013 from the southwestern Gulf of Mexico and provide information on their horizontal and vertical distribution using stratified samples collected up to 1,000 m depth. An identification key for 7 species of Amphilochidea and Senticaudata recorded in the Gulf of Mexico is provided.
Materials and methods
Zooplankton samples were collected in the southwestern Gulf of Mexico from January 23 to February 3, 2013 during the oceanographic campaign “Zooplancton de la capa mesopelágica del sur del Golfo de México” (ZOOMEP-1) with the Research Vessel “Justo Sierra”. Samples were obtained over a grid of 25 oceanographic stations (Fig. 1, Table 1) covering oceanic waters, using a stratified opening-closing plankton net system (nets with 75 cm mouth and 505 µm mesh) with a flowmeter at the mouth of each net to estimate the volume of filtered water. At each station, 2 sets of stratified sampling nets were made: the first set with 3 nets to sample the waters between 0-600 m depth, and the second set with 2 nets to sample the waters between 600-1,000 m depth. This effort was carried out at most at 5 levels of the water column, depending on bottom depth (Table 1): level 1: 0-200 m, level 2: 200-400 m, level 3: 400-600 m, level 4: 600-800 m,
and level 5: 800-1,000 m. Thus, a total of 104 samples were obtained. The samples were fixed in 4% formalin and preserved in 70% ethanol. In the laboratory, amphipods were sorted, dissected, and body parts mounted on semi-permanent slides. The taxonomic identification was made following the publications of Hughes and Lowry (2015), LeCroy (2000), Lowry and Stoddart (1997), Stebbing (1888, 1897), and Vinogradov (1999). One individual of each species was deposited in the Colección Nacional de Crustáceos (CNCR), Instituto de Biología, Universidad Nacional Autónoma de México, and the rest of examined individuals are deposited in the zooplankton collection of the Laboratorio de Ecología de Sistemas Pelágicos, Instituto de Ciencias del Mar y Limnología (ICML-LESP), Universidad Nacional Autónoma de Mexico.
For the species account we follow the taxonomic classification of Horton et al. (2019). The diagnoses of the species are mainly based on publications of several authors, which are indicated in the correspondent species account. In addition, mean density (ind/1,000 m3) value of each species was calculated by the following formula:
Mean density = ((∑N ((ni × 1000)/V(i)))/N)
ni = number of individuals at sampling station i; V(i) = volume of filtered water at sampling station i; N = number of sampling stations
Descriptions
Order Amphipoda Latreille, 1816
Suborder Amphilochidea Boeck, 1871
Infraorder Lysianassida Dana, 1849
Parvorder Lysianassidira Dana, 1849
Superfamily Lysianassoidea Dana, 1849
Family Cyphocarididae Lowry & Stoddart, 1997
Cyphocaris Boeck, 1871
Cyphocaris anonyx Boeck, 1871
(Fig. 2)
Diagnosis. Slender and smooth body, total length 3-15 mm, antenna 1 shorter than antenna 2, extending beyond pereonite 6. Head narrow, deeper than longer; rostrum absent. Eyes present, rounded. Pereionite 1 conspicuously produced anterodorsally. Coxae of pereopods 1-3 smaller than the following coxae. Gnathopods 1-2 weakly subchelate. Pereopods 3-4 with an expanded distal end of the propodus. Pereopod 3 coxa not anteriorly produced. Pereopod 4 coxa anterior margin subacute. Pereopod 5 coxa smaller than coxa 4, basis posterior margin spur with margins serrate, almost reaching the length of merus, not reaching beyond pleonite 1; basis posterior margins of pereopods 5-7 dentate. Telson relatively short, not reaching the end of uropod 3. Sources: Hughes and Lowry (2015), Lowry and Stoddart (1997), and Vinogradov (1999).
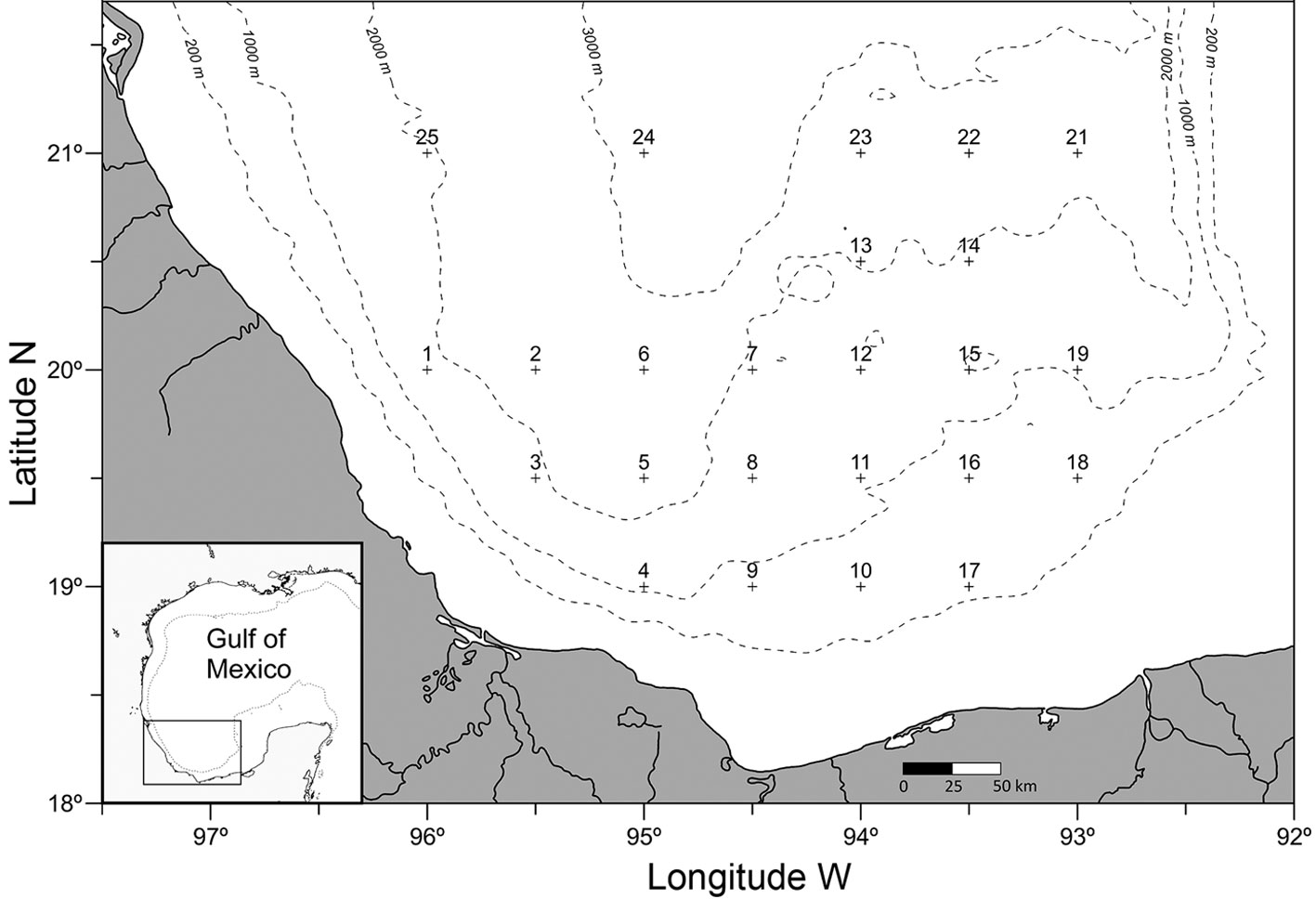
Taxonomic summary
Type locality. Greenland, North Atlantic Ocean (Hughes & Lowry, 2015).
Depth range. 0 – 2,523 m (Hughes & Lowry, 2015).
Geographical distribution. North Atlantic: Greenland, central north Atlantic, New York-USA, Bermuda, Caribbean Sea, Iceland, Norway, Ireland, west Bay of Biscay; Gulf of Mexico; South Atlantic: oceanic waters between America and Africa, Brazil, Gulf of Guinea, south Africa; North Pacific: Bering Sea, Gulf of Alaska, Punta Rompiente-Mexico; South Pacific: Solomon Sea, New Caledonia, oceanic waters off Peru and Chile; Western Indian Ocean: next to Madagascar; Australia: north-east and south oceanic waters; Sub-Antarctic Ocean: south-east of Campbell Island (Bellan-Santini & Costello, 2001; GBIF.org, 2019; Hughes & Lowry, 2015; Hurley, 1963; Lalana et al., 2005; Miloslavich et al., 2010; OBIS, 2019; Thurston, 1976; Vinogradov, 1999, 2004; this study).
Table 1
Geographical coordinates, depth, and sampled levels of sampling stations in the southwestern Gulf of Mexico.
|
Station |
Latitude |
Longitude W |
Depth |
Sampled levels |
|
1 |
20 |
96 |
0 – 1,000 |
1-5 |
|
2 |
20 |
95.5 |
0 – 1,000 |
1-5 |
|
3 |
19.5 |
95.5 |
0 – 1,000 |
1-5 |
|
4 |
19 |
95 |
0 – 1,000 |
1-5 |
|
5 |
19.5 |
95 |
0 – 1,000 |
1-5 |
|
6 |
20 |
95 |
0 – 1,000 |
1-5 |
|
7 |
20 |
94.5 |
0 – 1,000 |
1-5 |
|
8 |
19.5 |
94.5 |
0 – 1,000 |
1-5 |
|
9 |
19 |
94.5 |
0 – 600 |
1-3 |
|
10 |
19 |
94 |
0 – 400 |
1-2 |
|
11 |
19.5 |
94 |
0 – 800 |
1-4 |
|
12 |
20 |
94 |
0 – 1,000 |
1-5 |
|
13 |
20.5 |
94 |
0 – 1,000 |
1-5 |
|
14 |
20.5 |
93.5 |
0 – 1,000 |
1-5 |
|
15 |
20 |
93.5 |
0 – 600 |
1-3 |
|
16 |
19.5 |
93.5 |
0 – 400 |
1-2 |
|
17 |
19 |
93.5 |
0 – 400 |
1-2 |
|
18 |
19.5 |
93 |
0 – 600 |
1-3 |
|
19 |
20 |
93 |
0 – 1,000 |
1-5 |
|
21 |
21 |
93 |
0 – 1,000 |
1-5 |
|
22 |
21 |
93.5 |
0 – 1,000 |
1-5 |
|
23 |
21 |
94 |
0 – 1,000 |
1-5 |
|
24 |
21 |
95 |
0 – 1,000 |
1-5 |
|
25 |
21 |
96 |
0 – 1,000 |
1-5 |
Material examined. Seven adults ♀♀ and 3 juveniles collected from: station 1, level 2, 1 ♀ ICML-LESP 0001; station 5, level 4, 1 juvenile ♀ CNCR 35502 and 1 juvenile ICML-LESP 0002; station 7, level 4, 1 ♀ ICML-LESP 0003; station 11, level 2, 1 ♀ ICML-LESP 0004; station 19, level 3, 1 ♀ ICML-LESP 0005; station 22, level 3, 1 juvenile ICML-LESP 0006; station 24, level 3, 2 ♀♀ ICML-LESP 0007; station 25, level 3, 1 ♀ ICML-LESP 0008.
Remarks
Cyphocaris anonyx and C. ananke Hughes & Lowry, 2015 are morphologically very similar. C. anonyx has the shape of the pereopod 4 coxa anterior margin subacute and the length of the telson shorter than the uropod 3 length, whilst in C. ananke the pereopod 4 coxa anterior margin is broadly rounded and the telson is as long as the uropod 3 (Hughes & Lowry, 2015). All the C. anonyx studied individuals have translucent rounded eyes as reported by Hughes and Lowry (2015), who annotated that this character was previously ignored possibly due to the bleaching effect of alcohol. The color of the examined individuals preserved in alcohol is yellowish. The mean density of this species was 0.15 ind/1,000 m3.
Cyphocaris challengeri Stebbing, 1888
(Fig. 3)
Diagnosis. Slender and smooth body with 2.7 to 5.7 mm of total length, antennae long; antenna 1 shorter than antenna 2. Head narrow, deeper than longer; rostrum absent. Eyes present, reniform. Pereoionite 1 produced anteriorly, overlapping head. Coxae of pereopods 1-3 smaller than the following coxae. Gnathopods 1-2 weakly subchelate. Pereopod 4 coxa anterior margin rounded, with a strongly convex anterior margin covering coxae of pereopods 1-3. Pereopod 5 basis proximal posterior margin serrate and the distal posterior margin spur smooth, exceeding length of merus and not reaching beyond pleonite 1; basis posterior margins of pereopods 5-7 dentate. Telson relatively short, not reaching the end of uropod 3. Sources: Hughes and Lowry (2015), Lowry and Stoddart (1997), Stebbing (1888), and Vinogradov (1999).
Taxonomic summary
Type locality. Hawaiian Islands, North Pacific Ocean (Hughes & Lowry, 2015).
Depth range. 0 – 5,987 m depth (Hughes & Lowry, 2015).
Geographical distribution. North Atlantic: oceanic waters of New York-USA, Bermuda, oceanic waters in front of Ireland, Canary Islands; Gulf of Mexico; South Atlantic: oceanic waters between America and Africa, Brazil, south Africa; North Pacific: Japan, Hawaiian Islands, Bering Sea, Gulf of Alaska, Canada, USA; South Pacific: Solomon Sea; Indian Ocean: Arabian Sea, southernmost Indian Ocean next to Madagascar; Southern Australia Basin (GBIF.org, 2019; Hughes & Lowry, 2015; OBIS, 2019; Thurston, 1976; Vinogradov, 1999, 2004; Yamada & Ikeda, 2000; this study).
Material examined. Station 3, level 2, 1 adult ♀, CNCR 35503; station 22, level 5, 1 juvenile, ICML-LESP 0009.
Remarks
Cyphocaris challengeri is similar to C. bouvieri Chevreux, 1916 in the shape of the pereopod 5 basis, but the main difference is the spur that exceeds the length of merus in C. challengeri but not in C. bouvieri (Hughes & Lowry, 2015). C. challengeri differs from C. tunicola (the only cyphocarid species reported in the Gulf before the present study) by having the dorsal margin of the spur of pereopod 5 basis smooth, and a shorter length of spur, whilst in C. tunicola the posterior margin of the spur is entirely serrated and the length of spur much longer (Hughes & Lowry, 2015; Lowry & Stoddart, 1997). The color of the examined individuals preserved in alcohol is yellowish. The mean density of this species was 0.04 ind/1,000 m3.
Infraorder Amphilochida Boeck, 1871
Parvorder Eusiridira Stebbing, 1888
Superfamily Eusiroidea Stebbing, 1888
Family Eusiridae Stebbing, 1888
Eusiropsis Stebbing, 1897
Eusiropsis riisei Stebbing, 1897
(Fig. 4)
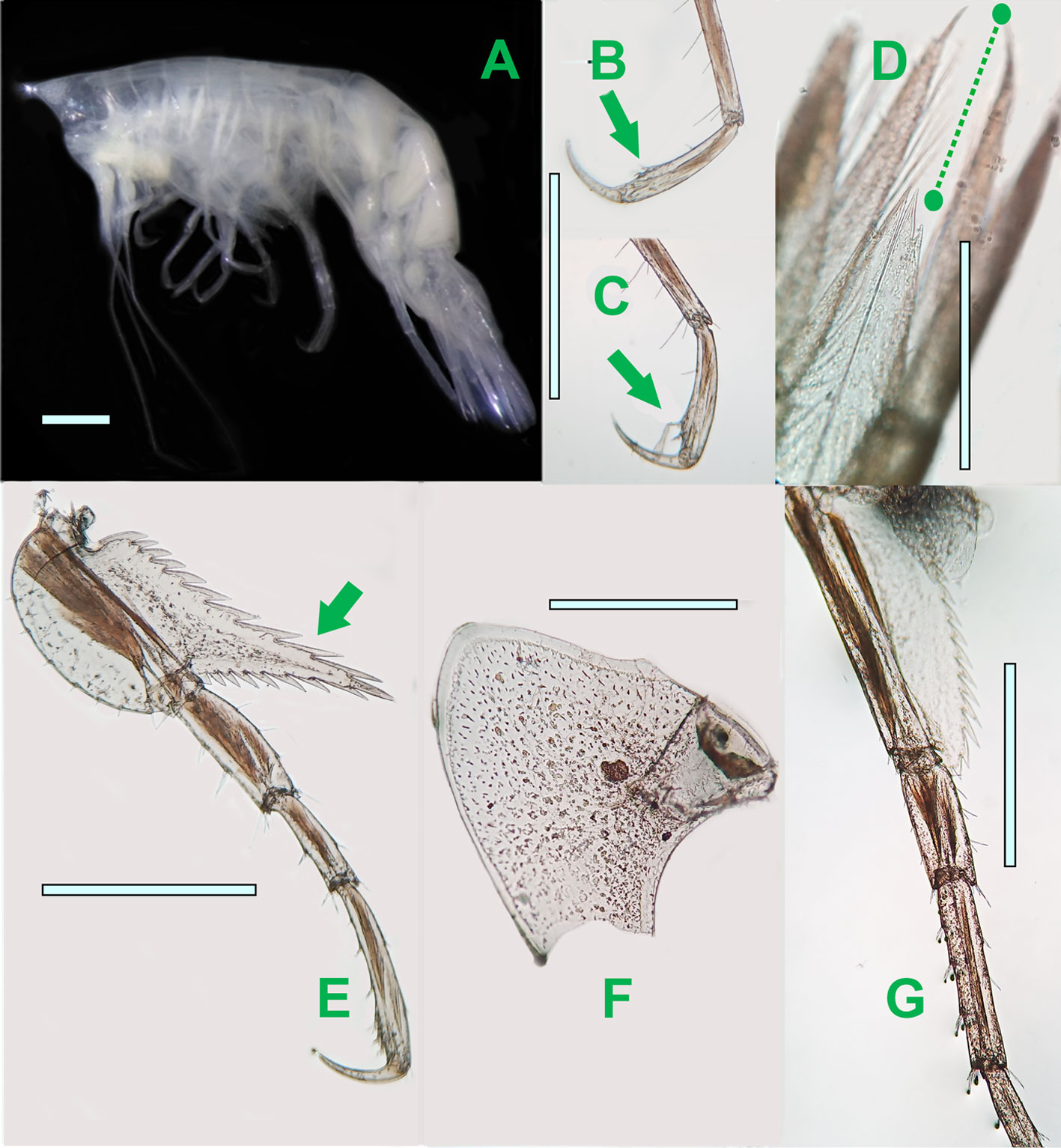
Diagnosis. Body slender, smooth, total length 7 mm, without mucronations on dorsal surface of pleonites. Eyes present, translucent, reniform. Head with triangular rostrum longer than broad. Antenna 1 and antenna 2 subequal in length, lower margin with some calceoli in antenna 1, calceoli in upper margin in antenna 2. Gnathopods 1-2 subchelate, subequal with eusirid form. Gnathopods 1-2 merus with posterior lobe, carpus lacking posterior lobe, propodus with a spinigerous process on posteriodistal edge. Pereopods 3-7 elongate and simple, the inferior margin setose, with dactylus simple. Basis of pereopods 5-7 with serrations in the posterior margin. Dorsal surface of all segments of the pleon smooth. Telson elongate, narrow, apically incised with the end margin with 4 very long plumose setae. Sources: Stebbing (1897), Vinogradov (1999), and this study.
Taxonomic summary
Type locality. Tropical Atlantic (Stebbing, 1897).
Depth range. 0 – 500 m (Vinogradov, 1999).
Geographical distribution. North Atlantic: Canary Islands; Gulf of Mexico; South Atlantic: Brazil; South Pacific: central part, Indonesia; North Indian Ocean: Arabian Sea (GBIF.org, 2019; OBIS, 2019; Thurston, 1976; Vinogradov, 1999; this study).
Material examined. Station 10, level 2, 1 adult ♀, CNCR 35504.
Remarks
There are only 2 species in the genus Eusiropsis and E. riisei is the type species (Horton et al., 2019). E. riisei differs from the other species E. spinidorsalis Gamo, 1981 by the lack of the conspicuous transverse rows of spinules on the dorsum surface of the pleon segments (Gamo, 1981). This characteristic is the most important and easily distinguishable morphologic feature among the 2 species, since E. riisei has the dorsal surface smooth in all segments (Stebbing, 1897). Previous studies indicated the absence of eyes in E. riisei (Stebbing, 1897; Vinogradov, 1999), but in the studied individual we observed translucents non-pigmented reniform ommatidia in each side of the head (Fig. 4B). According to Gamo (1981), E. spinidorsalis has a pair of distinct reddish eyes. The distribution range of E. spinidorsalis is restricted to the northwestern Pacific Ocean, only recorded in Japan (Gamo, 1981; GBIF.org, 2019). The color of the examined specimen preserved in alcohol is yellowish. The mean density of this species was 0.02 ind/1,000 m3.
Suborder Senticaudata Lowry & Myers, 2013
Infraorder Hadziida S. Karaman, 1943
Parvorder Hadziidira S. Karaman, 1932
Superfamily Calliopioidea G.O. Sars, 1895
Family Calliopiidae G.O. Sars, 1893
Stenopleura Stebbing, 1888
Stenopleura atlantica Stebbing, 1888
(Fig. 5)
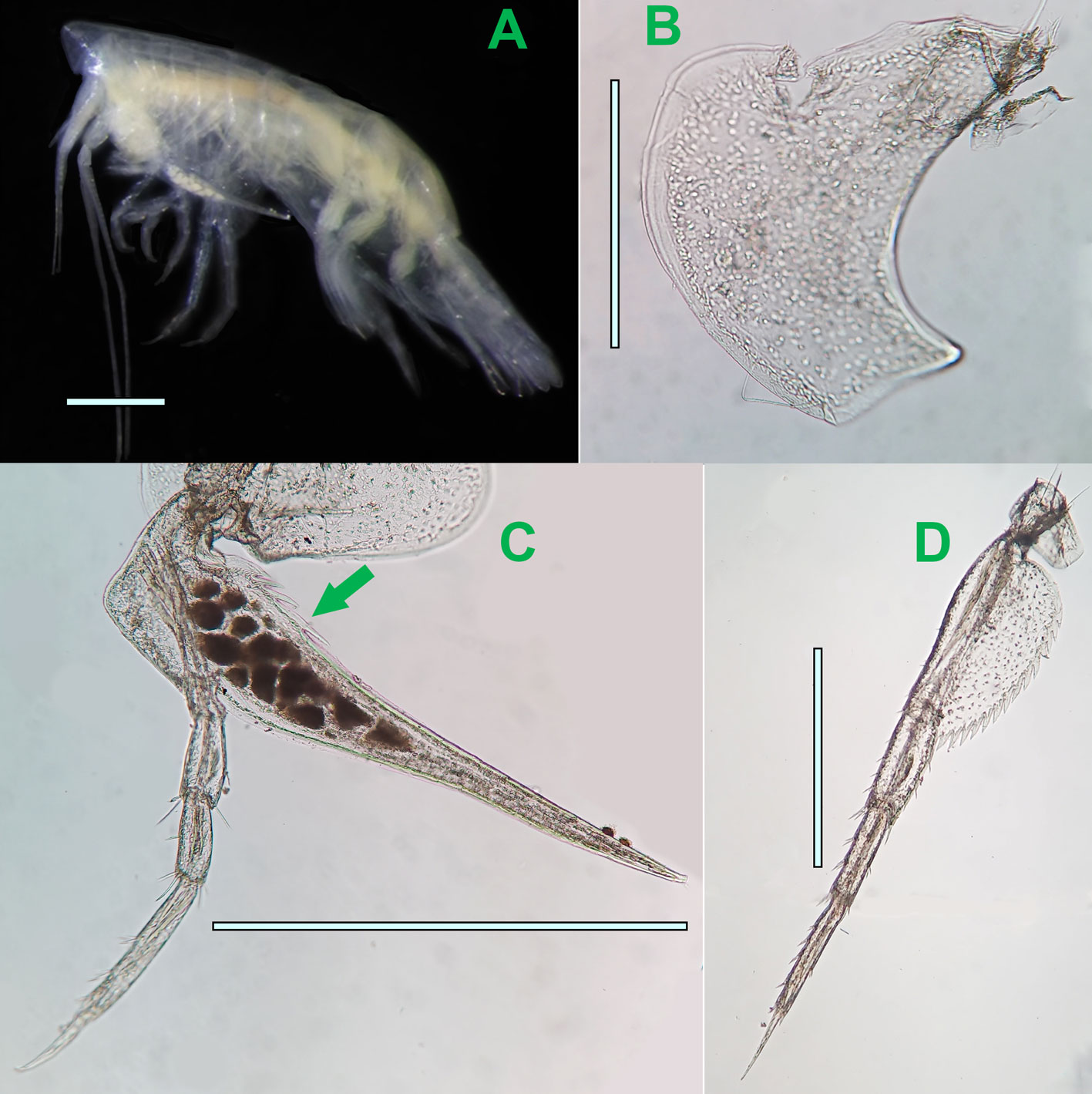
Diagnosis. Body slender, smooth, total length 2.5-4.7 mm. Antenna 1 peduncle short, second article much shorter than article 1; antenna 1 slightly longer than antenna 2. Eyes clearly defined, reniform, pigmented in red. Short coxae. Gnathopods 1 and 2 with large posterior lobe in merus, smaller than pereopods; gnathopod 1 prehensile and gnathopod 2 elongate and slender subchelate. Pereopods 3-7 elongate and simple; pereopod 7 with broad basis as long as merus. Uropods armed with lateral spines. Telson entire relatively short, as broad as long, apically incised. Sources: Stebbing (1888), Vinogradov (1999), this study.
Taxonomic summary
Type locality. Mid Atlantic Ocean 1°47’ N, 24°26’ W (Stebbing, 1888).
Depth range. 0 – 800 m, mainly at surface waters (Thurston, 1976).
Geographical distribution. North Atlantic: Cuba, Ireland, Canary Islands; Gulf of Mexico; South Atlantic: oceanic waters between America and Africa, Brazil, Gulf of Guinea, South Africa; South Pacific: New Zealand; Eastern Tropical Pacific: coast of Panama; North Indian Ocean: Arabian Sea; Antarctic: Davis Sea (Bellan-Santini & Costello, 2001; De Broyer et al., 2007; GBIF.org, 2019; Lalana et al., 2005; Miloslavich et al., 2010; OBIS, 2019; Vinogradov, 1999; this study).

Material examined. Forty-seven adult ♀♀, 9 ovigerous ♀♀ collected from: station 1, level 1, 1 ♀ ,CNCR 35505, 9 ♀♀ and 1 ovigerous ♀ ICML-LESP 0010; station 1, level 2, 1 ♀ ICML-LESP 0011; station 1, level 5, 3 ♀♀, ICML-LESP 0012; station 2, level 1, 1 ♀ and 1 ovigerous ♀, ICML-LESP 0013; station 2, level 2, 1 ♀, ICML-LESP 0014; station 2, level 5, 1 ♀, ICML-LESP 0015; station 3, level 1, 1 ♀, ICML-LESP 0016; station 3, level 2, 1 ♀, ICML-LESP 0017; station 4, level 2, 1 ♀ and 1 ovigerous ♀, ICML-LESP 0018; station 4, level 3, 2 ♀♀, ICML-LESP 0019; station 4, level 4, 1 ♀, ICML-LESP 0020; station 4, level 5, 1 ♀, ICML-LESP 0021; station 5, level 1, 5 ♀♀, ICML-LESP 0022; station 5, level 2, 1 ♀, ICML-LESP 0023; station 5, level 3, 1 ♀, ICML-LESP 0024; station 5, level 4, 1 ♀, ICML-LESP 0025; station 7, level 1, 2 ♀♀, ICML-LESP 0026; station 7, level 4, 1 ♀, ICML-LESP 0027; station 8, level 1, 1 ♀, ICML-LESP 0028; station 8, level 3, 2 ♀♀, ICML-LESP 0029; station 11, level 1, 2 ovigerous ♀♀, ICML-LESP 0030; station 12, level 3, 1 ♀, ICML-LESP 0031; station 12, level 4, 1 ♀, ICML-LESP 0032; station 12, level 5, 2 ovigerous ♀♀, ICML-LESP 0033; station 13, level 1, 1 ♀, ICML-LESP 0034; station 13, level 3, 1 ♀, ICML-LESP 0035; station 14, level 4, 1 ♀, ICML-LESP 0036; station 16, level 1, 1 ovigerous ♀, ICML-LESP 0037; station 19, level 3, 1 ♀, ICML-LESP 0038; station 21, level 3, 1 ovigerous ♀, ICML-LESP 0039; station 22, level 2, 1 ♀, ICML-LESP 0040; station 23, level 3, 1 ♀, ICML-LESP 0041; station 25, level 2, 1 ♀, ICML-LESP 0042.
Remarks
Stebbing (1888) noted that the genus Stenopleura is morphologically intermediate between the Atylidae G. O. Sars, 1882 and the Eusiridae Stebbing, 1888 due to its shape of mouth parts, and gnathopods 1-2 with an apparent eusirid form, but with important differences in the rest of pereopods, uropods and telson in comparison to the genus Eusirus Krøyer, 1845, so, finally the author established the new monotypic genus Stenopleura in the family Atylidae. Later, this species was reassigned by Sars (1895) in the family Calliopiidae. Stenopleura, currently considered as a monotypic genus differs from Calliopius Lilljeborg, 1865 by the shape of the distal margin of telson, in Calliopius is rounded, and in Stenopleura, the distal margin of telson is incised (Barnard, 1964). In this study, all individuals collected were adult females. The number of ovigerous females (Fig. 5B) were highest with 5 specimens at level 1, in contrast with 1 specimen in level 2 and level 3 each, and 2 in level 5. In level 4 ovigerous females were absent. The color of the examined specimens preserved in alcohol is yellowish. The mean density of this species was 1.13 ind/1,000 m3.
Discussion
This is the first attempt to study the deep-water (by stratified sampling) non-hyperiid planktonic amphipods (up to 1,000 m depth) in the southwestern Gulf of Mexico (Fig. 1). Four species of non-hyperiid planktonic amphipods are recorded for the first time in this area: Cyphocaris anonyx, C. challengeri, Eusiropsis riisei, and Stenopleura atlantica. All of them were observed as free-living forms, not associated with gelatinous organisms. With these new records, the species richness of non-hyperiid planktonic amphipods in the Gulf of Mexico rises to 7 (Table 2). In the western Atlantic Ocean, the 4 species here encountered had been mainly recorded at temperate latitudes (GBIF.org, 2019; OBIS, 2019; Vinogradov, 1999). Our records in the Gulf of Mexico extend their geographical range in the Atlantic (Table 2), indicating their affinity to tropical waters in the western Atlantic. Only C. anonyx and S. atlantica have previous records in the Caribbean Sea (Lalana et al., 2005; Miloslavich et al., 2010). In the northeastern Gulf of Mexico, near Florida, the species C. tunicola was recorded in association with pelagic tunicates between 200 to 500 m depth (Lowry & Stoddart, 1997). The vertical distribution of S. atlantica is extended up to 1,000 m depth; previously, Thurston (1976) recorded this species up to 800 m depth, referring as the greatest depth where the species inhabits in the north Atlantic Ocean.
Regarding the abundance, we consider that C. challengeri and E. riisei are rare species in the study area, since only 1 or 2 individuals are represented in our samplings. In contrast, S. atlantica was found in 17 (from 25) oceanographic stations at all sampling levels (Table 2), with its highest density at surface waters (0 – 200 m). Thurston (1976) found this species in the third abundance position of the non-hyperiid planktonic amphipods at surface waters (40 – 85 m depth) in the North Atlantic. The occurrence of ovigerous females of S. atlantica more frequently at level 1 (0 – 200 m depth) in our study suggests that the species mostly reproduces in the epipelagic zone, in concordance with the findings of Thurston (1976), where S. atlantica juveniles were present only in surface waters, however, further studies are needed to determine the environmental factors affecting the reproduction of the species.
The presence of eyes is an important morphological characteristic in the taxonomy of the non-hyperiid species. In a key to Lysianassoid taxa from western Atlantic Ocean published by Lowry and Stoddart (1997), the species C. anonyx was distinguished by the absence of eyes. However, Hughes and Lowry (2015) re-described this species with eyes and discussed that this character was previously ignored possibly due to the bleaching effect of alcohol. All the examined specimens of the 4 species of non-hyperiid amphipods reported in this work have eyes. The C. anonyx specimens have translucent rounded eyes as previously reported by Hughes and Lowry (2015), and C. challenger specimens have reniform eyes as reported by Vinogradov (1999). The studied specimen of Eusiropsis riisei has translucent reniform eyes. In the original description of this species, Stebbing (1897) reported the absence of eyes, and even Vinogradov (1999) extended this absence as a character for the diagnosis of the genus Eusiropsis. Therefore, the presence of eyes in E. riisei is for the first time here reported (Fig. 4B). The specimens of S. atlantica have reniform eyes as reported by Stebbing (1988) and Vinogradov (1999), the eyes are red in the present material.
Finally, in the Gulf of Mexico and the Caribbean Sea, new records of planktonic amphipods (hyperiids and non-hyperiids) are still being found in the region (Gasca, 2003, 2009; Lowry & Stoddart, 1997; Miloslavich et al., 2010). Previous studies and our results indicate the need for more taxonomic studies addressing the diversity of planktonic amphipods in Mexican waters, specially mesopelagic ones.
Table 2
Summary of records of non-hyperiid amphipods in the Gulf of Mexico, and their general geographical distribution.
|
Taxon |
Depth (m) |
Geographical distribution |
References |
|
|
Previous records |
Current observation |
|||
|
Suborder Amphilochidea Family Cyphocarididae Cyphocaris anonyx |
0 – 2,500 |
200 – 800 |
North and south Atlantic; Gulf of Mexico, Caribbean Sea; north and south Pacific; Indian Ocean; Australia; Antarctic. |
Bellan-Santini & Costello (2001); GBIF.org (2019); Hughes & Lowry (2015); Hurley (1963); Lalana et al. (2005); Miloslavich et al. (2010); OBIS (2019); Thurston (1976); Vinogradov (1999, 2004); this study. |
|
Cyphocaris challengeri |
0 – 5,987 |
200 – 1,000 |
North and south Atlantic; Gulf of Mexico, north and south Pacific; north Indian Ocean; Australia. |
GBIF.org (2019); Hughes & Lowry (2015); OBIS (2019); Thurston (1976); Vinogradov (1999, 2004); Yamada & Ikeda (2000); this study. |
|
Cyphocaris tunicola |
200-500 |
– |
Gulf of Mexico (endemic). |
Lowry & Stoddart (1997); LeCroy et al. (2009); OBIS (2019). |
|
Family Eurytheneidae Eurythenes obesus |
1,518-1,700 |
– |
North and south Atlantic; Gulf of Mexico; south Pacific; Indian Ocean; Australia; Antarctic. |
Shoemaker (1956); LeCroy et al. (2009); OBIS (2019). |
|
Family Synopiidae Synopia ultramarina |
0-219 |
– |
North and south Atlantic; Gulf of Mexico; north and south Pacific; Indian Ocean; Australia. |
Barnard (1972); Barnard & Thomas (1989); LeCroy et al. (2009); OBIS (2019). |
|
Family Eusiridae Eusiropsis riisei |
0 – 500 |
200 – 400 |
North and south Atlantic; Gulf of Mexico, south Pacific; Indian Ocean. |
GBIF.org (2019); OBIS (2019); Thurston (1976); Vinogradov (1999); this study. |
|
Suborder Senticaudata Family Calliopiidae Stenopleura atlantica |
0 – 800 |
0 – 1,000 |
North and south Atlantic; Gulf of Mexico, Caribbean Sea; east tropical and south western Pacific; Indian Ocean; Antarctic. |
Bellan-Santini & Costello (2001); De Broyer et al. (2007); GBIF.org (2019); Lalana et al. (2005); Miloslavich et al. (2010); OBIS (2019); Vinogradov (1999); this study. |
Key to species of non-hyperiid planktonic amphipods from the Gulf of Mexico.
Adapted from Barnard and Thomas (1989), Hughes (2009), Hughes and Lowry (2015), Lowry and Stoddart (1997), Stebbing (1888, 1897), and Stoddart and Lowry (2004).
1. Gnathopod 1 simple, weakly subchelate or parachelate. Gnathopod 2 simple or minute subchelate. Merus and carpus without posterior lobe 2
– Gnathopod 1-2 subchelate with the propodus much wider than basis or eusirid like very similar shape and size. Merus or carpus with posterior lobe 6
2. Coxae 1-2 very reduced or vestigial, coxa 3 smaller than coxa 4. Gnathopod 1 simple, weakly subchelate or parachelate, Gnathopod 2 minute subchelate 3
– Coxae 1-2 smaller than coxa 3, coxa 3 very large enfolding coxa 4. Gnathopod 1 weakly subchelate. Gnathopod 2 simple. (Synopiidae) Synopia ultramarina Dana, 1853
3. Gnathopod 1 simple or weakly subchelate. Coxae 1-3 small. Pereopod 5 basis posterior margin with spur. Pereopod 5-7 merus not expanded (Cyphocarididae: Cyphocaris) 4
– Gnathopod 1 parachelate, dactylus over-reaching palm. Pereopod 5 basis posterior margin without spur. Pereopods 5-7 merus expanded posteriorly. (Eurytheneidae) Eurythenes obesus (Chevreux, 1905)
4. Pereopod 5 posterior margin spur not reaching beyond pleonite 1. Eye present and small. Pereonite 1 without a large, narrow process 5
– Pereopod 5 posterior margin spur extremely elongate, reaching beyond pleonite 1. Eye conspicuous greatly enlarged, length at least 3 times as long as wide. Pereonite 1 forming long, narrow, slightly down-turned processC. tunicola Lowry & Stoddart, 1997
5. Pereopod 5 basis posterior spur margins serrate, not extending more than merus. Pereopod 4 coxa anterior margin is subacute. Telson short not reaching the end of uropod 3 C. anonyx Boeck, 1871 (Fig. 2)
– Pereopod 5 basis posterior margin serrate but spur distal margins smooth, exceeding merus length. Pereopod 4 coxa anterior margin rounded. Telson short not reaching the end of uropod 3C. challengeri Stebbing, 1888 (Fig. 3)
6. Antennae 1-2 with calceoli. Gnathopods 1-2 subchelate, subequal with eusirid form, propodus with a spinigerous pocket on posterior edge. Telson very large, apically incised, with 4 very long plumose setae. (Eusiridae) Eusiropsis riisei Stebbing, 1897 (Fig. 4)
– Antenna 1-2 without calceoli. Gnathopod 1-2 subchelate, subequal without eusirid form, propodus without a spinigerous pocket on posterior edge. Telson entire relative short, as broad as long, apically incised, without long setae (Calliopiidae) Stenopleura atlantica Stebbing, 1888 (Fig. 5)
Acknowledgments
To the Universidad Nacional Autónoma de México (UNAM) for the financial support of the oceanographic cruise ZOOMEP-1, coordinated by CFC. Authors appreciate the assistance of Faustino Zavala García during the sampling processes, José L. Villalobos Hiriart for the facilities to deposit the specimens in the Colección Nacional de Crustáceos, Instituto de Biología, UNAM, and Tomohiko Kikuchi from Yokohama Nacional University of Japan, for providing specialized literature. Also, the authors thank two anonymous reviewers who kindly reviewed the manuscript. This work is part of the Master of Science thesis of the first author, who is grateful for the scholarship granted by Consejo Nacional de Ciencia y Tecnología/Programa Nacional de Posgrados de Calidad (Conacyt/PNPC, 862851) of Mexico.
References
Barnard, J. L. (1964). Revision of some families, genera and species of Gammaridean Amphipoda. Crustaceana, 7, 49–74. https://doi.org/10.1163/156854064X00263
Barnard, J. L. (1972). A review of the family Synopiidae (= Tironidae), mainly distributed in the deep sea (Crustacea: Amphipoda). Smithsonian Contributions to Zoology, 124, 1–94. https://doi.org/10.5479/si.00810282.124
Barnard, J. L., & Thomas, J. D. (1989). Four species of Synopiidae from the Caribbean region (Crustacea: Amphipoda). Proceedings of the Biological Society of Washington, 102, 362–374.
Bellan-Santini, D., & Costello, M. J. (2001). Amphipoda. In M. Costello (Ed.), European register of marine species: a check-list of the marine species in Europe and a bibliography of guides to their identification (pp. 295–308). Paris: Collection Patrimoines Naturels.
Bowman, T. E. (1955). A new genus and species of whale-louse (Amphipoda: Cyamidae) from the false killer whale. Bulletin of Marine Science of the Gulf and Caribbean, 5, 315–320.
De Broyer, C., Lowry, J. K., Jazdzewski, K., & Robert, H. (2007). Catalogue of the Gammaridean and Corophiidean Amphipoda (Crustacea) of the Southern Ocean, with distribution and ecological data. In De Broyer C. (Ed.), Census of Antarctic marine life: synopsis of the Amphipoda of the Southern Ocean. Vol. I. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Biologie, Suppl. 1, 77, 1–325.
Gamo, S. (1981). A new bathypelagic Gammaridean amphipod, Eusiropsis spinidorsalis sp. nov. (Crustacea) taken off the Ogasawara Islands. Science Reports of the Yokohama National University, Sec. II, 28, 1–10.
Gasca, R. (2003). Hyperiid amphipods (Crustacea: Peracarida) in relation to a cold-core ring in the Gulf of Mexico. Hydrobiologia, 510, 115–124. https://doi.org/10.1023/B:HYDR.0000008637.16933.6f
Gasca, R. (2004). Distribution and abundance of hyperiid amphipods in relation to summer mesoscale features in the southern Gulf of Mexico. Journal of Plankton Research, 26, 993–1003. https://doi.org/10.1093/plankt/fbh091
Gasca, R. (2009). Diversity of hyperiid amphipods (Crustacea: Peracarida) in the western Caribbean Sea: news from the deep. Zoological Studies, 48, 63–70.
Gasca, R., Manzanilla, H., & Suárez-Morales, E. (2009). Distribution of hyperiid amphipods (Crustacea) of the southern Gulf of Mexico, summer and winter, 1991. Journal of Plankton Research, 31, 1493–1504. https://doi.org/10.1093/plankt/fbp096
GBIF.org. (2019). GBIF Occurrence Download (https://doi.org/10.15468/39omei). Retrieved on 24 September 2019 from: https://www.gbif.org
Horton, T., Lowry, J., De Broyer, C., Bellan-Santini, D., Coleman, C. O., Corbari, L. et al. (2019). World Amphipoda database. Amphipoda. Retrieved on 09 January 2019 from: http://www.marinespecies.org/aphia.php?p=taxdetails&id=1135
Hughes, L. E. (2009). Synopiidae. In J. K. Lowry, & A. A. Myers (Eds), Benthic Amphipoda (Crustacea: Peracarida) of the Great Barrier Reef, Australia. Zootaxa, 2260, 1–930. https://doi.org/10.11646/zootaxa.2260.1.50
Hughes, L. E., & Lowry, J. K. (2015). A review of the world Cyphocarididae with description of three new species (Crustacea, Amphipoda, Lysianassoidea). Zootaxa, 4058, 1–40. https://doi.org/10.11646/zootaxa.4058.1.1
Hurley, D. E. (1963). Amphipoda of the family Lysianassidae from the west coast of North and Central America. Los Angeles: University of Southern California Press.
Lalana, R., Ortiz, M., & Varela, C. (2005). Primera adición a la lista de los crustáceos no decápodos de Cuba. Revista de Biología, 19, 50–56.
LeCroy, S. (2000). An illustrate identification guide to the nearshore marine and estuarine Gammaridean Amphipoda of Florida. Volume 1. Families Gammaridae, Hadziidae, Isaeidae, Melitidae and Oedicerotidae. Tallahassee: Florida Department of Environmental Protection.
LeCroy, S., Gasca, R., Winfield, I., Ortiz, M., & Escobar-Briones, E. (2009). Amphipoda (Crustacea) of the Gulf of Mexico. In D. Felder, & D. Camp (Eds), Gulf of Mexico: origins, waters and biota, Volume 1, Biodiversity (pp. 941–972). College Station: Texas A&M University Press.
Lowry, J. K., & Myers, A. A. (2017). A Phylogeny and classification of the Amphipoda with the establishment of the new order Ingolfiellida (Crustacea: Peracarida). Zootaxa, 4265, 1–89. https://doi.org/10.11646/zootaxa.4265.1.1
Lowry, J. K., & Stoddart, H. E. (1997). Amphipoda Crustacea IV. Families Aristiidae, Cyphocarididae, Endevoursidae, Lysianassidae, Scopelocheiridae, Uristidae. Memoirs of the Hourglass Cruises, 10, 1–148.
Marrón-Becerra, A., Hermoso-Salazar, M., & Solís-Weiss, V. (2018). Hyalella maya, a new Hyalellidae species (Crustacea: Amphipoda) from a cenote in the Yucatan peninsula, Mexico. Journal of Cave and Karst Studies, 80, 1–11. https://doi.org/10.4311/2017LSC0115
Miloslavich, P., Díaz, J. M., Klein, E, Alvarado, J. J., Díaz, C., Gobin, J. et al. (2010). Marine biodiversity in the Caribbean: regional estimates and distribution patterns. Plos One, 5, e11916. https://doi.org/10.1371/journal.pone.0011916
OBIS (Ocean Biogeographic Information System). (2019). Retrieved on 21 February 2019 from: https://obis.org/
Sars, G. O. (1895). An account of the Crustacea of Norway, with short descriptions and figures of all the species. Amphipoda. Vol. I. Amphipoda. Christiania and Copenhagen: Alb. Cammermeyers Forlag.
Shoemaker, C. R. (1956). Notes on the amphipods Eurythenes gryllus (Lichtenstein) and Katius obesus Chevreux. Proceedings of the Biological Society of Washington, 69, 177–178.
Stebbing, T. R. R. (1888). Report on the Amphipoda collected by H.M.S. Challenger during the years 1873-1876. Report on the scientific results of the Voyage of H.M.S. Challenger during the years 1873-76. Zoology, 29, 1–1737.
Stebbing, T. R. R. (1897). Amphipoda from the Copenhagen Museum and other sources. The Transactions of the Linnean Society of London: Zoology, 2, 25–45. https://doi.org/10.1111/j.1096-3642.1897.tb00400.x
Stoddart, H. E., & Lowry, J. K. (2004). The deep-sea lysianassoid genus Eurythenes (Crustacea, Amphipoda, Eurytheneidae n. fam.). Zoosystema, 26, 425–468.
Thomas, J. D., & Barnard, J. L. (1992). Podocerus cheloniphilus, a testudinous amphipod newly recorded from the western Atlantic Ocean. Bulletin of Marine Science, 50, 108–116.
Thurston, M. L. H. (1976). The vertical distribution and diurnal migration of the Crustacea Amphipoda collected during the Sond Cruise, 1965: I. The Gammaridea. Journal of the Marine Biological Association of the United Kingdom, 56, 359–382. https://doi.org/10.1017/S002531540001897X
Vinogradov, G. (1999). Amphipoda. In D. Boltovskoy (Ed.), South Atlantic zooplankton (pp. 1141–1240). Leiden: Backhuys Publishers.
Vinogradov, G. (2004). Near-bottom and pelagic gammaridean amphipods in the western Indian Ocean. Annals of the South African Museum, 112, 39–88.
Vinogradov, M. E., Volkov, A. F., & Semenova, T. N. (1996). Hyperiid amphipods (Amphipoda, Hyperiidea) of the world oceans. Washington D.C.: Smithsonian Institution Libraries.
Violante-Huerta, M. (2018). La epibiosis en los grandes vertebrados marinos de México: una revisión y su relevancia ecosistémica. Revista Peruana de Biología, 25, 335–342. https://doi.org/10.15381/rpb.v25i3.14786
Yamada, Y., & Ikeda, T. (2000). Development, maturation, brood size and generation length of the mesopelagic amphipod Cyphocaris challengeri (Gammaridea: Lysianassidae) off southwest Hokkaido, Japan. Marine Biology, 137, 933–942. https://doi.org/10.1007/s002270000397


