
Eduardo Soto-Galera *, Shiomara Herrera-Gutiérrez, Nidia Leticia Mendoza-Castro, Joel Paulo-Maya y Fabian Pérez-Miranda
Instituto Politécnico Nacional, Escuela Nacional de Ciencias Biológicas, Departamento de Zoología, Laboratorio de Bioconservación y Manejo, Prol. de Carpio y Plan de Ayala, Col. Casco de Santo Tomás, 11340 Ciudad de México, México
*Autor para correspondencia: egalera@ipn.mx (E. Soto-Galera)
Recibido: 8 noviembre 2022; aceptado: 17 octubre 2023
Resumen
Los efectos de la introducción de Amatitlania nigrofasciata sobre la ictiofauna nativa han sido poco explorados. El objetivo del presente estudio fue analizar la influencia del espacio y alimento como factores que expliquen la coexistencia de este cíclido con el nativo Herichthys pantostictus en la Reserva de la Biosfera Barranca de Metztitlán. Con la información procedente de recolectas y del monitoreo ambiental de 7 localidades visitadas entre marzo de 2016 y mayo de 2018, se determinaron el uso, selección y traslape de hábitat, mediante índices y métodos multivariados. Adicionalmente, se calculó el traslape de la dieta, con aquellos ejemplares que coincidieron espacial y temporalmente. Los resultados señalan que el uso y la selección diferencial del hábitat, en función de la cobertura de distintos sustratos y profundidades, se manifiesta en un bajo traslape espacial entre las especies, en contraste con el traslape de dieta que es muy alto. Así, el aprovechamiento diferencial del espacio permite explicar la coexistencia de estos cíclidos en la reserva. Esto último no necesariamente es una respuesta a las condiciones ambientales o a la utilización de determinados recursos, sino que puede ser resultado de interacciones de competencia relacionadas con la defensa de territorios de anidación o alimentación.
Palabras clave: Traslape de hábitat; Traslape de dieta; Selección del hábitat; Coexistencia; Convicto; Mojarra de Chairel
© 2024 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Spatial and trophic segregation between the invasive cichlid Amatitlania nigrofasciata and the native Herichthys pantostictus in rivers of the Barranca de Metztitlán Biosphere Reserve, Hidalgo, Mexico
Abstract
The effects of the introduction of Amatitlania nigrofasciata on the native fish fauna have been little assessed. The objective of this paper is to analyze the influence of space and food resources as factors that explain the coexistence of this exotic cichlid with the native Herichthys pantostictus in the Barranca de Metztitlán Biosphere Reserve. Based on information from collections and environmental monitoring of 7 locations visited between March 2016 and May 2018, the use, selection and overlapping of habitat were determined, using multivariate indices and methods. Additionally, diet overlap was calculated, with those specimens that coincided in space and time. The results indicate that the use and differential selection of the habitat, depending on the coverage of different substrates and depths, manifested in low spatial overlap between the species, in contrast, the diet overlap is extremely high. Thus, the differential use of space makes possible to explain the coexistence of these cichlids in the reserve. The latter is not necessarily a response to environmental conditions or the use of certain resources but may also be the result of competition interactions related to the defense of nesting or feeding territories.
Keywords: Habitat overlap; Diet overlap; Habitat selection; Coexistence; Convict; Chairel mojarra
© 2024 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introduction
Dentro del conjunto de especies exóticas, existe un subgrupo conocido como especies invasoras, las cuales son capaces de sobrevivir, establecerse y reproducirse de manera descontrolada fuera de su ambiente natural, desarrollando un comportamiento invasivo, desplazando especies nativas y causando graves daños a la biodiversidad, economía, agricultura o salud pública (Williamson, 1996). Actualmente, se reconoce que una de las principales causas de pérdida de la biodiversidad en el mundo es la presencia de las especies invasoras (McNeely, 2001). En México, la presencia de especies invasoras en ambientes acuáticos es uno de los problemas más severos y menos controlados, en 1904 solo se reconocían 4 especies de peces exóticas o translocadas; 7 en 1969, 55 en 1983, 94 en 1997, 102 en 2008 (Contreras-Balderas, 2008) y 118 en 2009 (Aguirre-Muñoz et al., 2009). Para el 2014, la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad reconoció la presencia de 93 especies de peces invasoras exóticas o translocadas (Mendoza y Koleff, 2014). La escaza regulación en la importación e introducción de nuevas especies al mercado nacional permite el cultivo de casi cualquier organismo acuático con poco control, provocando así una tasa de introducción y establecimiento de nuevas especies para la región, de 1 cada 5 años (Contreras-MacBeath et al., 2014). Una especie que se establece con éxito en un nuevo ecosistema, por lo general, tiene que competir con las especies nativas por los recursos existentes, alimento o espacio (David et al., 2017; Simberloff, 2013); la consecuencia de ello es la modificación de las tramas tróficas locales y/o el desplazamiento de especies nativas (Britton et al., 2018; Catford et al., 2018). Sin embargo, una mínima diferenciación en los recursos utilizados en cualquier nivel (espacio, tiempo o recursos alimentarios) puede ser un factor mediador de la coexistencia entre especies (Bolnick et al., 2011; Chase y Leibold, 2003). Por tanto, estudios sobre el uso de recursos son necesarios para evaluar la competencia, especialmente si en el sistema se presenta una especie no nativa que pueda desplazar a las nativas sintópicas (Gozlan et al., 2010; Roberts y Britton, 2018).
La especie Amatitlania nigrofasciata (Günther, 1867) es un pez pequeño de la familia Cichlidae oriundo de Centroamérica, conocido en acuariofilia como convicto o pez Congo. Su distribución en la vertiente del Pacífico comprende desde el río Sucio en El Salvador hasta el río Suchiate en Guatemala y en la vertiente atlántica desde el río Patuca, Honduras hasta el río Jutiapa, Guatemala (Schmitter-Soto et al., 2007). Es ampliamente utilizado en el comercio de peces ornamentales y es precisamente a partir de liberaciones accidentales durante esta actividad, que ha sido introducido en diversas regiones del mundo: EUA (Courtenay et al., 1984), Israel (Welcomme, 1988), Hawái (Eldredge, 1994), México (Contreras-MacBeath et al., 2014), Reunión (Keith et al., 2006), Perú (Cossios, 2010), Japón (Ishikawa y Tachihara, 2010), Irán (Reza-Esmaeili et al., 2013), Australia (Duffy et al., 2013), Italia (Bianco, 2014), China (Xiong et al., 2015), Hungría (Takács et al., 2017), Colombia (Herrera-R. et al., 2017) y Brasil (Gurgel-Lourenço et al., 2019). Su amplia tolerancia ambiental y la capacidad de colonizar hábitats perturbados, el oportunismo trófico, el cuidado biparental, su agresividad durante la competencia territorial, rápida tasa de crecimiento, maduración temprana y periodos de desove largos con múltiples puestas, muy probablemente contribuyen al éxito del establecimiento de este cíclido en islas y otras regiones (CABI, 2019; Gagliardi-Seeley, 2012; Ishikawa y Tachihara, 2010; McKaye, 1977; Wisenden, 1994; Yamamoto y Tagawa, 2000). En México se le considera como especie exótica invasora con presencia, desde 1984, en la cuenca del río Amacuzac (Conabio, 2016; Mejía-Mojica et al., 2012) y ampliamente distribuida en la cuenca del Pánuco desde el 2000 (Miranda et al., 2012; Montaño-Campos, 2008; Sánchez-Barrera, 2010; Soto-Galera et al., 2018). En la Reserva de la Biosfera Barranca de Metztitlán (RBBM) el convicto fue registrado por primera vez en 2007, se piensa que fue introducido de manera deliberada como consecuencia de su comercio con fines ornamentales. Además de este cíclido, se reporta la presencia de otras especies ajenas a la reserva, 3 exóticas y 3 translocadas. De entre las especies nativas, la única perteneciente a la familia Cichlidae es Herichthys pantostictus (Miranda et al., 2012), endémica de la cuenca Pánuco-Tamesí (Pérez-Miranda et al., 2018); este cíclido comparte con A. nigrofasciata estrategias alimentarias y reproductivas, ya que ambas son omnívoras, tienden a ser territoriales, agresivas y presentan cuidados parentales (De la Maza-Benignos, 2014; De la Maza-Benignos y Lozano-Vilano, 2013; Ishikawa y Tachihara, 2010; Trujillo-Jiménez, 1996). La presencia espacial simultánea de estos 2 cíclidos en la RBBM representa una excelente oportunidad para determinar la influencia de la utilización de los recursos espacio y alimento como factores que explican su coexistencia a más de 10 años de la introducción del convicto a la RBBM, lo que analizamos mediante el uso, selección y traslape de hábitat para el caso de la dimensión espacial, mientras que para el caso de la trófica, lo abordamos con el análisis de sus dietas y el traslape de éstas.
Materiales y métodos
La Reserva de la Biosfera Barranca de Metztitlán (RBBM) se encuentra al centro-este de Hidalgo. Comprende la cuenca de la barranca de Metztitlán, entre los paralelos 98°23’00” y 98°57’08” longitud oeste y 20°14’15” y 20°45’26” latitud norte, presenta gradientes altitudinales que van desde los 1,000 hasta los 2,000 m snm, con una extensión de 96,042.94 ha (Conanp, 2003). Los 2 principales ríos que drenan la RBBM son tributarios altos de la cuenca del río Amajac, uno de los afluentes más importantes de la cuenca del río Pánuco, con un área de 6,884 km². El primero es un río que a lo largo de sus 100 km en la reserva recibe el nombre de acuerdo con la región; identificándose 3 tramos importantes, el primero en su entrada, al sur de la barranca se conoce como río Grande de Tulancingo; el segundo llamado río Venados, en la unión con el río San Sebastián; y el tercer tramo donde inicia el Distrito 08 Metztitlán se denomina río Metztitlán, al norte de la reserva; este río corre de SE a NO, y al N de la reserva desemboca en la laguna de Metztitlán. El segundo es una pequeña porción del río Amajac, antes de la afluencia del río Metztitlán, abajo de la laguna de mismo nombre (este tramo pequeño también se conoce como río Almolón) en la porción noroeste de la reserva (Argota-Quiroz, 2011; Conanp, 2003).
Dentro de la RBBM se ubicaron 7 localidades de estudio, 5 en el río Metztitlán: 1) barranca del Chilar y San Pablo, a 500 m del poblado Los Baños (20°17’50.5”N, 98°28’02.1”O); 2) río Venados, a 7.05 km al SO de Metzquititlán (20°28’18.9”N, 98°40’52.5”O); 3) Geoparque Comarca Minera, a 12 km al NE de Atotonilco el Grande (20°20’22”N, 98°36’37”O); 4) San Pedro Tlatemalco, 12.53 km al SE de Metztitlán (20°32’07”N, 98°43’32”O) y 5) Jihuico, 7.12 km al SE de Metztitlán (20°32’30”N, 98°43’35”O); y 2 en el río Amajac: 6) puente en el libramiento a Cardonal, a 2 km al SE de San Pablo Tetlapayac (20°37’46”N, 98°54’37.7”O) y 7) San Pablo Tetlapayac (20°38’27.2”N, 98°55’15.1”O) (fig. 1).
El trabajo de campo contempló muestreos en las 2 temporadas del año que para la cuenca del río Metztitlán señala Mendoza-Cariño et al. (2018): sequía (de marzo a mayo de 2016 y mayo 2018) y lluvias (agosto-noviembre 2016 y noviembre del 2017). En cada localidad se efectuó la captura de peces y el monitoreo de los factores ambientales, en todas las porciones o áreas (sitios o hábitats) en un tramo de 30 m de río que se identificaron como diferentes en términos de velocidad de corriente, profundidad y porcentaje de cobertura de distintos tipos de sustrato, independientemente de la presencia de peces, de acuerdo con lo señalado por Jowett y Richardson (1995). Los peces fueron recolectados mediante electropesca por vadeo con un equipo de AbP-3TM; posteriormente, se fijaron con una solución de formaldehído al 10%, con la finalidad de detener el proceso de digestión y mantener el contenido estomacal en condiciones aceptables para su análisis posterior, cualitativo y cuantitativo, en el laboratorio (Cailliet et al.,1996). Los ejemplares recolectados fueron debidamente identificados y etiquetados con la fecha, localidad y sitio de recolecta. En cada sitio se cuantificaron las siguientes características del hábitat: profundidad (m) con un estadal graduado; velocidad de corriente a la mitad de la columna de agua (m/s) con un flujómetro (Global Water FP201) y el porcentaje de cobertura de los diferentes tipos de sustratos: roca madre (cantos rodados y materiales pétreos con diámetro > a los 0.5 m), cantos rodados < 0.5 m de diámetro, grava 2-16 mm y arena 0.0625-2 mm; las coberturas de detritus, sedimentos finos y de vegetación acuática, que son comúnmente empleadas en estudios del hábitat ribereño de peces (Reid, 2019), no fueron contempladas ya que fueron muy escasas en los sitios de estudio.
Para la verificación taxonómica de los ejemplares de H. pantostictus y A. nigrofasciata se siguieron los criterios establecidos por Pérez-Miranda et al. (2018) y Schmitter-Soto (2007), respectivamente; para el resto de la ictiofauna se siguió a Miller et al. (2009). Los nombres específicos se presentan de acuerdo con el catálogo de Fricke et al. (2022). Posteriormente, se contabilizó el número de individuos por sitio de muestreo para cada uno de los cíclidos de interés, se determinó la longitud patrón (LP) de cada ejemplar con ayuda de un vernier digital Mitutoyo con precisión de 0.01 mm, se sustituyó el formaldehído al 10% por alcohol etílico al 70% y se depositaron a la Colección Nacional de Peces Dulceacuícolas Mexicanos de la Escuela Nacional de Ciencias Biológicas (ENCB-IPN-P).
Para el análisis de la dimensión espacial, se construyó la matriz de las variables físicas registradas en toda la gama de sitios de muestreo (hábitats), es decir de todas las localidades y temporadas de muestreo, a la que se aplicó un análisis de componentes principales (ACP) a partir de la matriz de correlación de Spearman, utilizando XLSTAT (Addinosoft, 2022), éste último empleado para visualizar en el espacio multivariado la similitud en el uso del hábitat (definido como el número total de ejemplares dentro de cada uno de los hábitats) entre la especie nativa y la invasora. Dado que las variables determinadas se encuentran expresadas en diferentes unidades, éstas fueron estandarizadas de la siguiente forma: velocidad de corriente (m/s) y profundidad (m) con el logaritmo natural (valor del parámetro ambiental + 1), mientras que a las determinaciones de los porcentajes de cobertura de los diferentes sustratos se les aplicó arcoseno (%/100)-2 (Krebs,1999). Componentes principales (CP) con valores propios ˃ 0.9 y variables con fuerte poder explicativo, cargas con valores absolutos ˃ 0.6 fueron retenidos. Los puntajes en los CP de cada uno de los sitios donde se presentaron los cíclidos fueron empleados como variables dependientes, mientras que las especies fueron las variables independientes en un análisis multivariado de varianza (Manova) que probó las diferencias en la utilización del hábitat entre las 2 especies con relación a las variables físicas determinadas (Reid, 2019). El análisis estadístico se realizó con el paquete estadístico PAST versión 1.94 (Hammer et al., 2020), con el empleo de la distribución T² de Hotelling para probar las diferencias y los valores de p ajustados con la corrección secuencial de Bonferoni. Para cada especie se desarrollaron las curvas de selección del hábitat de modo independiente para el porcentaje combinado de roca y cantos rodados menores de 0.5 m, de grava y arena, profundidad (m) y velocidad de corriente (m/s) (Reid, 2019). Las curvas se generaron con el programa “Habitat Selection” 3.10 (Jowett, 2022), los procedimientos numéricos y la terminología de este programa se basan en los trabajos de Jowett y Davey (2007) y Manly et al. (1993). Así, la fórmula para la estimación de la selección del hábitat (selectividad) es la misma que propusieron Manly et al. (1993) para la determinación de tasas de forrajeo: Wi = ui (Ʃui)-1/ai (Ʃui)-1, donde Wi es la tasa de forrajeo para el iecimo de n categorías de hábitat, ui es el número total de organismos en la categoría del hábitat i, Σui es el número total de organismos en todas las categorías de hábitat, ai es el número de muestras de la categoría i y Σai es el número total de muestras. Este índice mide la intensidad de la selección (selectividad) para una categoría de hábitat particular, los valores de W = 1 indican selectividad neutral, es decir, la categoría en cuestión se usa justo en la proporción con la que se encuentra en el ambiente; valores de W < 1 se usan con menos frecuencia de lo esperado por la casualidad; es decir, son hábitats que se evitan y valores con W > 1 son hábitats que se usan con más frecuencia de lo que se espera por la casualidad y, por lo tanto, son los seleccionados o preferidos. Adicionalmente, con los datos del número de ejemplares por sitio, de la especie nativa y la exótica, correspondientes a cada localidad, en cada temporada, se determinó la magnitud del traslape espacial, mediante el índice de Pianka (1973): Ojk = Ʃpijpik / (Ʃpij 2 Ʃpik2)1/2, donde Ojk = traslape de Pianka, Pij = proporción del recurso i del total de los recursos utilizados por la especie j y Pik = proporción del recurso i del total de los recursos utilizados por la especie k. El índice fluctúa de 0 a 1, donde 0 indica que no hay traslape y 1 representa traslape total entre el par de especies analizadas, los valores entre 0.6 y 1.0 indican traslape alto, entre 0.4 y 0.59 intermedio, y menores de 0.4 traslape bajo (Grossman, 1986).
Para el análisis de la dimensión trófica, se determinó el contenido estomacal solo con los ejemplares de ambas especies que fueron recolectados el mismo día y en la misma localidad para evitar el cálculo del traslape entre individuos que no coincidieron en espacio y tiempo (Reinas et al., 2022). El análisis se efectuó para clases de talla de 10 mm: 50 < 70 mm, 70 < 90 mm, 90 < 110 mm y > 110 mm para H. pantostictus, y de 30 < 40 mm, 40 < 50 mm, 50 < 60 mm y > 60 mm para A. nigrofasciata. El procedimiento empleado es una modificación del método volumétrico (Cailliet et al., 1996), que consiste en extender el contenido del primer tercio del tubo digestivo (la parte con el contenido menos digerido) de cada uno de los ejemplares en una caja de Petri marcada con una cuadrícula milimétrica, separar los diferentes componentes y, posteriormente, determinar su identidad taxonómica siguiendo los criterios de Pennak (1978), Needham y Needham (1982) y Pineda-López et al. (2009), para finalmente expresar el área ocupada por cada componente como una proporción del total del área ocupada por todo el contenido estomacal. Puesto que ambas especies son omnívoras y se alimentan de una extensa variedad de componentes alimenticios (Pérez-Miranda et al., 2019; Trujillo-Jiménez, 1998), la determinación de estos se llevó a cabo a una escala de categorías taxonómicas superiores, como lo sugieren Kramer y Bryant (1995) y Lombard et al. (2018). Se analizaron las diferencias de dieta, ubicando las clases de talla y localidades en gradientes de consumo, obtenidos mediante un análisis de componentes principales (ACP) aplicado a la matriz de consumo, previamente transformada con el arco seno de la raíz cuadrada de la proporción del alimento, con el programa XLSTAT (Addinosoft, 2022). Para constatar las diferencias en la composición de la dieta (variable de respuesta) entre las clases de talla de las especies (variable independiente) y variaciones espaciales en su dieta (diferencias entre localidades; variable independiente), se aplicó un análisis de varianza multivariante permutacional (Permanova) (Ferreira et al., 2021; Reinas et al., 2022), los datos fueron previamente transformados (con el arco seno de la raíz cuadrada de la proporción consumida de cada tipo de alimento) con distancias euclidianas y con 9,999 permutaciones para calcular los valores de p, con el paquete estadístico PAST 1.94 (Hammer et al., 2020). Adicionalmente, se calculó el traslape de dieta mediante el índice de Pianka (1973) entre todas las clases de talla de ambas especies que fueron recolectadas simultáneamente, los valores del índice varían de 0 a 1; para evaluar los resultados obtenidos se siguieron los criterios de Grossman (1986). Este índice es utilizado cuando por la naturaleza de los elementos alimenticios que consumen las especies, es poco factible determinar la disponibilidad de estas en el ambiente, además de ser uno de los índices más ampliamente usados cuando se trata de especies omnívoras, como sería el caso de ciclídos y loricáridos entre otros (Ferreira et al., 2021; Gois et al., 2015; Lombard et al., 2018; Maitra et al., 2019; Zengeya y Marshall, 2007).

Resultados
Se efectuaron 82 eventos de muestreo y recolecta de peces (sitios o hábitats) en las 7 localidades, donde un total de 4,715 ejemplares de 16 especie fueron capturados, Amatitlania nigrofasciata fue la segunda más abundante (8.72%) con 417 (209 en lluvias, 208 en secas) en 39 eventos de recolecta procedentes de 3 localidades (río Venados, Jihuico y San Pedro Tlatemalco); H. pantostictus estuvo representada con 322 ejemplares (113 en lluvias, 209 en secas) procedentes de 68 eventos de recolecta en las 7 localidades, siendo la quinta especie más abundante (6.36%). Otras especies muy abundantes fueron Poeciliopsis gracilis (translocada), Oreochromis mossambicus (exótica), Pseudoxiphophorus jonesii (nativa) y Xiphophorus helleri (translocada) con 55.6, 8.72, 6.4 y 6.26%, respectivamente. Mientras que las especies exóticas Abramis brama, Carassius auratus, Cyprinus carpio e Ictalurus punctatus, las translocadas Chirostoma jordani y Goodea atripinnis, así como las nativas Tampichthys ipni, Astyanax argentatus, Ictalurus mexicanus y Poecilia mexicana no sobrepasaron individualmente 1.75% de la abundancia. Herichthys pantostictus y A. nigrofasciata coincidieron en 28 eventos de recolecta en el río Venados, San Pedro Tlatemalco y Jihuico. La media (intervalo, desviación estándar) de la longitud patrón de H. pantostictus fue de 75.93 mm (13.3-134.61, 28.49 mm) y la de A. nigrofasciata fue de 49.31 mm (26.44-77.4, 9.67 mm).
Tabla 1
Valores de las variables del hábitat disponible y utilizado para H. pantostictus y A. nigrofasciata en la RBMB. Media (intervalo de variación) y desviación estándar de los sitios de muestreo.
| Variables del hábitat | Disponible | H. pantostictus | A. nigrofasciata |
| Profundidad (m) | 0.365 (0.08-1.00) 0.20 | 0.374 (0.80-1.00) 0.19 | 0.378 (0.10-1.00) 0.24 |
| Velocidad de corriente (m/s) | 0.435 (0.0-1.717) 0.40 | 0.459 (0.0-1.717) 0.39 | 0.351 (0.0 -1.717) 0.38 |
| Cantos rodados > 0.5m (%) | 10.71 (0-80) 20.70 | 12.17 (0-80) 22.22 | 6.95 (0-70) 17.69 |
| Cantos rodados < 0.5m (%) | 51.49 (0-100) 35.63 | 53.00 (0-100) 34.50 | 41.31(0-100) 37.45 |
| Grava (%) | 14.65 (0-100) 22.65 | 14.64 (0-90) 20.75 | 15.60 (0-100) 27.0 |
| Arena (%) | 19.17 (0-100) 29.52 | 17.41 (0-100) 27.91 | 32.33 (0-100) 35.40 |
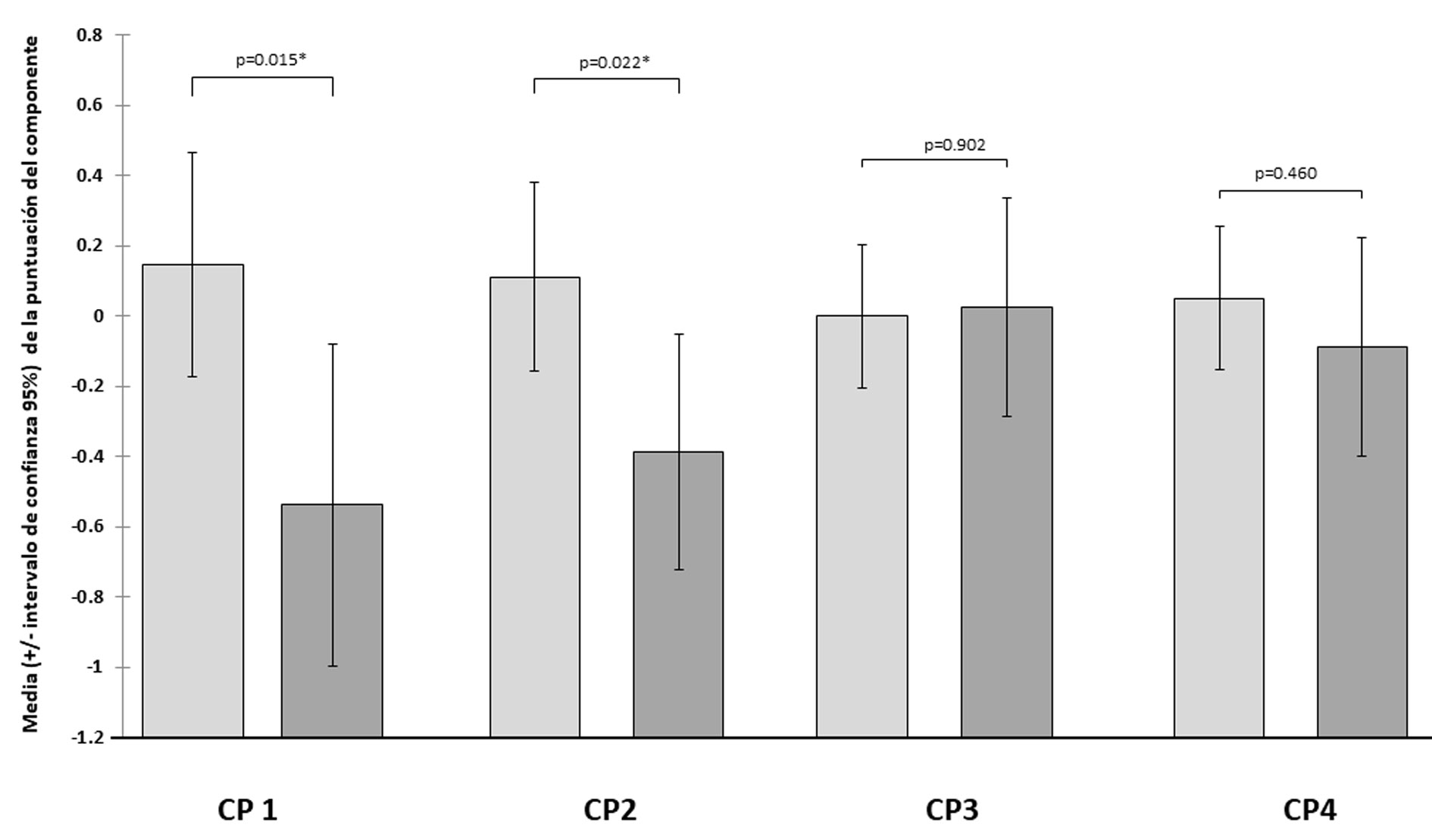
Las condiciones de flujo en las localidades estuvieron caracterizadas por profundidades someras (< 1 m) y velocidades de corriente promedio de 0.435 m/s, los sistemas estudiados presentan desde remansos hasta rápidos con velocidades de 1.71 m/s. El lecho del río estuvo constituido principalmente por sustratos mixtos, típicamente de cantos rodados con diámetros menores a 0.5 m, pero con porcentajes apreciables de grava, arena y roca mayor a 0.5 m (tabla 1). El ACP explicó en sus primeros 4 componentes 86.07% de la varianza del uso y disponibilidad del hábitat. El primer eje (CP1) demuestra diferencias en el uso entre las especies de cobertura de cantos rodados menores a 0.5 m y arena, diferencias en profundidad y cobertura de grava son explicadas en el componente 2, de roca por el componente 3 y de velocidad por el componente 4. Existen diferencias significativas (Manova Wilks Lambda: 0.91, p < 0.0182) de los puntajes de los sitios en los componentes asociados al uso del hábitat entre las especies. Las diferencias en el CP1 indican que H. pantostictus manifestó un mayor uso de hábitats con alta cobertura de cantos rodados menores 0.5 m y baja de arena en comparación con A. nigrofasciata, en el CP2 el cíclido nativo manifiesta un uso más acentuado de hábitats con mayores profundidades y coberturas de grava que el invasor, finalmente los componentes 3 y 4 indican que H. pantostictus utiliza más frecuentemente hábitats con mayor cobertura de roca y velocidades de corriente más elevadas que A. nigrofasciata (tabla 1, fig. 2). Herichthys pantostictus seleccionó hábitats con sustratos mixtos, constituidos por roca-cantos rodados con porcentajes entre 20 y 80%, complementados con grava-arena (20-80%), profundidades de entre 0.85 y 1 m; así como, velocidades de corriente entre 0.60 y 1.15 m/s. La selección del hábitat en función de estas mismas variables para el caso de A. nigrofasciata es totalmente distinta, ya que ésta fue por hábitats con coberturas de sustrato siempre menores a 27% de roca-cantos rodados, mayores de 63% de grava-arena, con profundidades entre 0.4 y 1m y por toda la gama de velocidades registradas, excluyendo las seleccionadas por el cíclido nativo (fig. 3). En cuanto a la temporada de estiaje, el índice de Pianka resulto bajo, con un valor de 0.30 para el traslape espacial entre las especies. Mientras que para la temporada de lluvias este fue de 0.29.
Tabla 2
Composición y abundancia de la dieta de H. pantostictus y A. nigrofasciata para las localidades de río Venados, Jihuico y San Pablo Tlatemalco en la RBBM, de noviembre 2017 a mayo 2018.
| H. pantostictus | A. nigrofasciata | |
| Componentes de la dieta | Contenido en la dieta (%) | |
| Larva Coleoptera (LC) | 5.45 | 6.18 |
| Larva Diptera (LD) | 19.36 | 19.40 |
| Ninfa Ephemeroptera (NE) | 21.67 | 14.45 |
| Larva Megaloptera (LM) | 0.42 | 0.00 |
| Larva Trichoptera (LT) | 3.27 | 8.54 |
| Ninfa Odonata (NO) | 0.00 | 0.15 |
| Restos de insectos adultos no identificables (RIANI) | 8.16 | 1.99 |
| Gastropoda (G) | 0.80 | 1.00 |
| Hydracarina (H) | 0.36 | 2.18 |
| Peces (P) | 2.50 | 6.53 |
| Algas filamentosas (AF) | 31.84 | 33.32 |
| Restos vegetales no identificables (RVNI) | 1.48 | 0.69 |
| Detritus (D) | 4.69 | 5.57 |
Los ejemplares de las 2 especies que coincidieron espacial y temporalmente proceden de las recolectas efectuadas en el río Venados en noviembre de 2017 y mayo de 2018. Así como de Jihuico, y San Pedro Tlatemalco en mayo de 2018. El análisis de la dieta indica la presencia de 13 tipos de alimento (tabla 2), siendo los insectos y las algas filamentosas los elementos más abundantes en ambos cíclidos, con poco más de 50 y 33% del contenido estomacal, respectivamente. El ACP explica en sus 3 primeros componentes 63% de la varianza de los gradientes de consumo, el CP1 está relacionado con el consumo de algas filamentosas y larvas de dípteros, mientras que el gradiente del CP 2 lo está con el consumo de ninfas de efemerópteros y larvas de tricópteros (fig. 4). Sin embargo, como lo señala el Permanova, la composición de la dieta no difiere entre las clases de talla de las especies (pseudo-F = 0.81854, p = 0.3844), pero sí entre localidades (pseudo-F = 2.8663, p = 0.0001), sin interacción entre los 2 factores (pseudo-F = -1.9437, p = 0.5527). La prueba pareada derivada del análisis muestra diferencias en los patrones de consumo de los cíclidos recolectados en Jihuico y las otras 2 localidades, primordialmente por el alto consumo de larvas de tricópteros y bajo de ninfas de efemeróteros, también existen diferentes patrones de consumo entre los peces recolectados en el río Venados en las 2 temporadas, ya que en mayo se da un mayor consumo de ninfas de efemerópteros. Finalmente, no existen diferencias entre las dietas de los peces capturados en el río Venados y San Pedro Tlatemalco (tabla 3, fig. 4). El traslape trófico entre las clases de talla de las 2 especies fue alto; para el caso de los cíclidos capturados en noviembre de 2017 se ubicó entre 0.672 y 0.768, mientras que para los capturados en mayo de 2018 el valor fluctuó entre 0.620 y 0.978 (tabla 4).
Tabla 3
Prueba pareada entre localidades y fecha de colecta resultado del Permanova aplicado a la abundancia de los componentes de la dieta por clase de talla obtenidos para H. pantostictus y A. nigrofasciata en la RBBM. * = Diferencias significativas.
| Río Venados
noviembre 2017 |
Jihuico
mayo 2018 |
Río Venados
mayo 2018 |
San Pedro Tlaltemalco
mayo 2018 |
|
| Río Venados noviembre 2017 | 1 | 0.0001* | 0.0161* | 0.2691 |
| Jihuico mayo 2018 | 0.0001* | 1 | 0.0001* | 0.0001* |
| Río Venados mayo 2018 | 0.0161* | 0.0001* | 1 | 0.2056 |
| San Pedro Tlaltemalco mayo 2018 | 0.2691 | 0.0001* | 0.2056 | 1 |
Tabla 4
Solapamiento de dieta (índice de Pianka) entre clases de talla de H. pantostictus y A. nigrofasciata en la RBBM noviembre 2017 y mayo 2018.
| CLT H. pantostictus (mm) | |||||
| CLT A. nigrofasciata (mm) | 50 < 70 | 70 < 90 | 90 < 110 | > 110 | |
| Río Venados/ noviembre 2017 | n | 5 | |||
| 30 < 40 | 4 | 0.768 | |||
| 40 < 50 | 5 | 0.672 | |||
| Jihuico/ mayo 2018 | n | 10 | 4 | 18 | |
| 50 < 60 | 4 | 0.633 | 0.618 | 0.739 | |
| Río Venados/ mayo 2018 | n | 10 | 4 | 18 | 18 |
| 30 < 40 | 10 | 0.763 | 0.773 | 0.731 | 0.694 |
| 40 < 50 | 33 | 0.822 | 0.642 | 0.647 | 0.620 |
| 50 < 60 | 19 | 0.914 | 0.796 | 0.635 | 0.645 |
| > 60 | 14 | 0.949 | 0.718 | 0.938 | 0.858 |
| San Pedro Tlatemalco / mayo 2018 | n | 7 | 11 | 16 | |
| 40 < 50 | 28 | 0.725 | 0.933 | 0.915 | |
| 50 < 60 | 18 | 0.718 | 0.963 | 0.978 | |
| > 60 | 6 | 0.756 | 0.862 | 0.906 |
Discusión
Si bien el registro de 16 especies ícticas para los ríos de la RBBM es igual al reportado por Miranda et al. (2012), existen pequeñas diferencias. En el presente estudio no fue posible recolectar al mugílido Dajaus monticola en el río Amajac, pero se registró la presencia del ciprínido exótico Carassius auratus en la localidad de Jihuico en ambas temporadas del año, lo que finalmente señala la presencia de 17 especies en la reserva. En cuanto a la abundancia y distribución de los 2 cíclidos de interés, para los años 2007-2008 A. nigrofasciata solo fue detectada en una localidad del río Metztitlán, donde además fue poco frecuente (Miranda et al., 2012), mientras que en 2017-2018 se le registró en 3 localidades del mismo río y fue la segunda especie más abundante del total de los ejemplares recolectados en la RBBM. Estos resultados son una clara manifestación de que la especie ha logrado establecerse, incluso datos preliminares señalan que este cíclido invasor se reproduce exitosamente dentro de la RBBM (Soto-Galera, et al., 2018).
Existen diferencias significativas en el uso y selección del hábitat entre H. pantostictus y A. nigrofasciata relacionadas con la cobertura del tipo de sustrato, profundidad y marginalmente, con la velocidad de corriente. En cuanto al uso del hábitat de H. pantostictus, Miller et al. (2009) mencionan que en la cuenca del río Pánuco, la especie se encuentra asociada a profundidades que alcanzan 2 m, sustratos diversos de lodo, arena, grava y rocas, vegetación asociada con algas, velocidades de la corriente bajas o nulas. En tanto, para la RBBM claramente hace uso más frecuente de sustratos mixtos con amplia cobertura de roca madre y cantos rodados (mayores a 40%), profundidades de 0.1 a 0.60 m y velocidades de corriente de hasta 0.9 m/s. En el río Santa María (cuenca del Pánuco), H. pantostictus selecciona hábitats profundos (> 2 m), con velocidades de corriente entre 0.3 y 0.35 m/s y coberturas a partes iguales de arena y roca (Soto-Galera y Alcántara-Soria, 2016), estos resultados difieren con lo registrado para la RBBM, donde suele seleccionar hábitats más someros, con mayores velocidades de corriente y sustratos mixtos con coberturas de roca madre y cantos rodados de entre 20 y 80%. Las diferencias encontradas en el uso y selección del hábitat son resultado de la amplia distribución de este cíclido en la cuenca del Pánuco, donde manifiesta diferentes morfotipos como resultado del uso de hábitats contrastantes que van desde ambientes lénticos, como lagunas y estuarios, hasta lóticos como ríos y arroyos (De la Maza-Benignos y Lozano-Vilano, 2013). Además, la determinación de la selección del hábitat en entornos donde no existe la gama completa asociada al ámbito de distribución y temporalidad de la especie en el momento de muestreo, solo será aplicable al entorno correspondiente (Jowett y Richardson, 2008).
La especie Amatitlania nigrofasciata habita entre rocas en aguas profundas, con corrientes de moderadas a rápidas, los adultos son frecuentes en la corriente de pequeños ríos o arroyos, hasta en las aguas someras de grandes y caudalosos ríos prefieren hábitats rocosos, donde encuentran refugio en las diversas grietas y hendiduras proporcionadas por este tipo de ambientes (Bussing, 1993; Conkel, 1993). En el río Acahuapa en El Salvador de donde es nativa, su presencia se asocia con hábitats donde la velocidad del agua es de moderada a baja, los sustratos son particularmente de roca, además de presentar ramas y troncos (Álvarez-Calderón, 2014). En la RBBM, si bien el convicto también hace uso de hábitats someros y con velocidades de corriente de moderadas a bajas, difiere en el uso del sustrato, ya que lo hace sobre sustratos mixtos con bajas coberturas de roca madre y cantos rodados. Estas diferencias solo ponen de manifiesto los amplios intervalos de tolerancia de este cíclido a tipos y coberturas de sustratos, velocidad de corriente, altitud y temperatura en las diversas regiones del planeta y en los diferentes ambientes (ríos, canales, embalses y pantanos) donde ha sido introducido (Contreras-MacBeath et al., 1998; Cossíos, 2010; Ishikawa y Tachihara, 2010; Mejía-Mojica et al., 2014; Scoppettone et al., 2011; Yamamoto y Tagawa, 2000). En cuanto a su selectividad por el tipo de hábitat, diversos autores señalan que prefiere desovar en cuevas, sobre sustratos de roca que le proporcionen refugios con las condiciones necesarias para la protección de huevos y crías (Conkel, 1993; Patterson, 1985; Weber y Weber, 1971). En la RBBM los resultados son radicalmente diferentes, ya que la selección es por sustratos mixtos con elevadas coberturas de grava-arena y bajas de roca madre-cantos rodados, que suponen condiciones poco favorables para la anidación, adicionalmente selecciona profundidades menores a 1 m y de la gama de velocidades de corriente disponibles, evita las intermedias (0.65-0.95 m/s), que son precisamente las que selecciona H. pantostictus.
Las diferencias expuestas en el uso y selección del hábitat claramente se manifiestan en bajo traslape espacial entre ambos cíclidos dentro de la reserva, lo que sugiere que a más de una década de la introducción de A. nigrofasciata a la RBBM, esta se encuentra segregada espacialmente con respecto a H. pantostictus, lo que potencialmente ha promovido la coexistencia. Misma situación ha sido presentada por Makocho y Satuffer (2018) para explicar la coexistencia en cíclidos de los hábitats rocosos (4 especies nativas vs. 2 introducidas) en el Parque Nacional del Lago Malawi. La segregación espacial en los cíclidos neotropicales se relaciona con los sitios de anidación, ya que como lo sugieren Buege et al. (2021), la distribución de los mismos se encuentra parcialmente gobernada por controles físicos (sustrato, profundidad, velocidad de corriente), pero también en el caso de especies con comportamiento agresivo, por la defensa territorial de los sitios de anidación, como sería el caso de las especies bajo estudio (De la Maza-Benignos, 2014; Gagliardi-Seeley, 2012; Itzkowitz et al., 1998; Rícan et al., 2016). Situación que también registraron Archundia y Arce (2019) entre Cichlasoma istlanum (endémica de la cuenca del Balsas) y A. nigrofasciata en condiciones de laboratorio, donde la primera tuvo más éxito en la obtención de los recursos en disputa (refugio y alimento). El bajo traslape señalado difiere de lo registrado para el binomio A. nigrofasciata y C. istlanum en la cuenca del Balsas, donde ocupan hábitats y sitios de anidación similares, lo que sugiere la existencia de competencia constante por los recursos y, por ende, el establecimiento de jerarquías de dominancia (Trujillo-Jiménez 1998).

Herichthys pantostictus se encuentra catalogada como una especie omnívora, que ingiere artrópodos, gasterópodos, restos vegetales, algas filamentosas y detritus (De la Maza-Benignos y Lozano-Vilano, 2013; Pérez-Miranda et al., 2019). El convicto ingiere una amplia gama de componentes tanto de origen animal como vegetal e incluso detritus; sin embargo, por la alta ingesta de insectos ha sido considerada como un entomófago especializado en el consumo de larvas de dípteros, ninfas de efemerópteros y zigópteros, aunque también se alimentan de peces, gasterópodos, crustáceos y materia vegetal que incluye semillas, restos de hojas y algas filamentosas (Bussing, 1993, 1998; Burcham, 1988). En ríos de la cuenca del Balsas se comporta como omnívoro con tendencia carnívora, ya que los elementos de origen animal constituyen poco más de 60% del total de su dieta o incluso como un detritívoro-herbívoro (Medina-Nava, 2011; Trujillo-Jiménez, 1998). En la RBBM ambos cíclidos manifiestan la ingesta de diferentes grupos de insectos que comprenden la mitad de su dieta, la materia vegetal alcanza un tercio de ésta, por lo que sí bien ambas especies se comportan como entomófagas especializadas, también pueden ser catalogadas como omnívoras. La similitud y los altos valores del traslape de dieta entre los 2 cíclidos indican la utilización común del recurso alimento, misma que bajo condiciones limitantes se traduciría en el desplazamiento de alguna de las 2 como consecuencia de la competencia, situación contraria, a la observada en la cuenca del Balsas, donde A. nigrofasciata presenta una dieta diferente a C. istlanum, que resultó en bajo traslape y prácticamente se descarta la competencia por este recurso (Trujillo-Jiménez, 1998).
La coexistencia del cíclido nativo H. pantostictus y el exótico A. nigrofasciata dentro de la RBBM es promovida principalmente por la selección y uso diferencial del espacio, mismo que les proporciona alimento, el cual está conformado mayoritariamente por organismos de deriva, estos componentes acuáticos y terrestres son la mayor fuente de alimento para los peces (Flecker, 1992), ya que como mencionan Tamaris-Turizo et al. (2013), los elementos de deriva son de gran importancia para los niveles tróficos en los ríos, debido al subsidio energético que suministran. La coexistencia de una o varias especies como resultado de la segregación espacial en estudios que analizan simultáneamente las dimensiones espacial y trófica, ha sido demostrada en especies de la familia Cyprinidae en América del norte (Gee y Northcote, 1963; Mendelson, 1975), en Salmo truta y Cottus poecilipus en Noruega (Hesthagen et al., 2004), en especies de la familia Loricariidae en Brasil (Reinas et al., 2022). Específicamente en la familia Cichlidae, la coexistencia de especies que utilizan el mismo recurso alimenticio en los grandes lagos africanos se explica también en función de la segregación espacial. Así, en el lago Malawi las diferencias interespecíficas e intraespecíficas en la utilización y defensa de los sitios de alimentación probablemente facilitan la coexistencia de especies del género Petrotilapia (Marsh y Ribbink, 1985). En tanto que la segregación de nichos a través de la microdistribución diferencial de especies del género Haplocromis que se alimentan de “Aufwuchs” (perifiton), se asoció a la profundidad y otros parámetros topográficos en el lago Victoria (Seehausen y Bouton, 1997). También, en 15 especies herbívoras de 4 ecomorfos de alimentación en el lago Tangañica se detectó diferenciación en el uso de profundidad y sustrato entre las especies de un mismo ecomorfo (Hata y Ochi, 2016). Finalmente, en el Parque Nacional del Lago Malawi, la disposición de áreas de alimentación de la fauna que habitan los litorales rocosos sugiere segregación espacial, lo que potencialmente ha promovido la coexistencia entre las especies nativas e introducidas, ésto a más de 30 años de la introducción de estas últimas (Makocho y Stauffer, 2018).

Finalmente, es importante señalar que la selección diferencial del hábitat y por tanto la segregación espacial entre H. pantostictus y A. nigrofasciata en la RBBM, no necesariamente es una respuesta a la utilización de determinados recursos y condiciones, sino que más bien pudiera ser el resultado de las interacciones de competencia relacionadas con la defensa de territorios de anidación o alimentación, ya que como señalan Hata y Ochi (2016), la segregación espacial por profundidad y tipo se sustrato entre los cíclidos que habitan los litorales rocosos del lago Tangañica se presenta en algunas especies como resultado de la defensa de territorios de anidación o alimentación, pero también en especies que no defienden territorios. Es por ello que futuros estudios relacionados con la utilización, selección y segregación del hábitat, deberán encaminarse a dimensionar las interacciones agonísticas entre estas 2 especies presentes en la RBBM, ya sea mediante ensayos de confrontación como los mencionados por Molina et al. (2021) o los de seguimiento de poblaciones en condiciones naturales como los realizados por Makocho y Stauffer (2018), sin dejar de lado el análisis de otras dimensiones del nicho trófico, como pudieran ser la cronología de la alimentación, los sitios específicos de esta actividad o incluso el tamaño de las presas, ello con la finalidad de comprender con mayor certidumbre los mecanismos de competencia que permiten explicar la coexistencia y las consecuencias futuras para estos cíclidos en la RBBM, a más de una década de la introducción de A. nigrofasciata.
Agradecimientos
Los resultados presentados forman parte de los proyectos: Estado actual, identificación de riesgos y propuestas para el manejo de las especies invasoras presentes en las regiones hidrológicas prioritarias. Confluencia de las Huastecas, Media Luna y Cabecera del río de La Laja (LI038), financiado por la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad y Estado actual, identificación de riesgos y propuestas para el manejo de las especies invasoras presentes en los afluentes de la laguna de Metztitlán, Hidalgo (20171914), financiado por la Secretaría de Investigación y Posgrado del Instituto Politécnico Nacional.
Referencias
Addinosoft. (2022). XLSTAT satatistycal an data análisis solution. New York, USA. Recuperado el 16 de agosto, 2022 de: https://www.xlstat.com/es
Aguirre-Muñoz, A., Mendoza-Alfaro, R., Arredondo-Ponce Bernal, H., Arriaga-Cabrera, L., Campos-González, E., Contreras-Balderas, S. et al. (2009). Especies exóticas invasoras: impactos sobre las poblaciones de flora y fauna, los procesos ecológicos y la economía. En R. Dirzo, R. González y I. J. March (Comp.), Capital natural de México, Vol. II. Estado de conservación y tendencias de cambio (pp. 277–318). México D.F.: Conabio.
Álvarez-Calderón, F. S. (2014). Estructura y distribución de la íctiofauna de la subcuenca del río Acahuapa, San Vicente, El Salvador (Tesis). Universidad de El Salvador. San Salvador, San Salvador.
Archundia, M. y Arce, E. (2019). Fighting behaviour in native fish: the Mexican mojarra (Cichlasoma istlanum) wins when confronted with the non‑native convict cichlid fish (Amatitlania nigrofasciata). Journal of Ethology, 37, 67–73. https://doi.org/10.1007/s10164-018-0569-5
Argota-Quiroz, T. (2011). Simulación hidrológica de la cuenca del río Amajac, estado de Hidalgo, aplicando el modelo SWAT (Tesis de maestría). Instituto Politécnico Nacional. México D.F.
Bianco, P. G. (2014). An update on the status of native and exotic freshwater fishes of Italy. Journal of Applied Ichthyology, 30, 62–77. https://doi.org/10.1111/jai.12291
Bolnick, D. I., Amarasekare, P., Araújo, M. S., Bürger, R., Levine, J. M., Novak, M. et al. (2011). Why intraspecific trait variation matters in community ecology. Trends in Ecology and Evolution, 26, 183–192. https://doi.org/10.1016/j.tree.2011.01.009
Britton, J. R., Ruiz-Navarro, A., Verreycken, H. y Amat-Trigo, F. (2018). Trophic consequences of introduced species: Comparative impacts of increased interspecific versus intraspecific competitive interactions. Functional Ecology, 32, 486–495. https://doi.org/10.1111/1365-2435.12978
Buege, E. A., Esselman, P. C. y Praskievicz, S. J. (2021). Hidrogeomorphological control son reach-scale distributions of cichid nest sites a small neotropical river. Ecology of Freshwater Fish, 30, 244–255. https://doi.org/10.1111/eff.12580
Burcham, J. (1988). Fish communities and environmental characteristics of two lowland streams in Costa Rica. Revista de Biología Tropical, 36, 273–285.
Bussing, W. A. (1993). Fish communities and environmental characteristics of a tropical rain forest river in Costa Rica. Revista de Biología Tropical, 41, 791–809.
Bussing, W. A. (1998). Peces de las aguas continentales de Costa Rica. San José: Universidad de Costa Rica.
CABI. (2019). Datasheet Amatitlania nigrofasciata (convict cichlid) invasive species compendium. Recuperado el 22 de septiembre 2022 de: https://www.cabi.org/isc/datasheet/112202#D2D94F15-C38F-49DB-BBAB-CE375C9113FC
Cailliet, G. M., Love, M. S. y Ebeling, A. W. (1996). Fishes: a field and laboratory manual on their structure, identification and natural history. Long Grove, Illinois: Waveland Pr Inc.
Catford, J.A., Bode, M. y Tilman, D. (2018). Introduced species that overcome life history tradeoffs can cause native extinctions. Nature Communications, 9, 1–7. https://doi.org/10.1038/s41467-018-04491-3
Chase, J. M. y Leibold, M. A. (2003). Ecological niches: linking classical and contemporary approaches. Chicago: University of Chicago Press.
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2003). Programa de Manejo Reserva de la Biosfera Barranca de Metztitlán. México D.F.: Instituto Nacional de Ecología.
Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad). (2016). Sistema de información sobre especies invasoras en México. Ciudad de México. Recuperado el 25 abril, 2016 de: http://www.biodiversidad.gob.mx/invasoras
Conkel, D. (1993). Cichlids of North and Central America. New Jersey: T.F.H. Publications Inc.
Contreras-Balderas, S. (2008). Especies de peces introducidas en aguas continentales de México. Catálogo y manuscrito. Bases de datos SNIB-Conabio Proyecto Núm. AE002. México D.F. Recuperado el 28 septiembre, 2022 de: http://www.conabio.gob.mx/institucion/cgi-bin/datos2.cgi?Letras=AE&Numero=2
Contreras-MacBeath, T., Gaspar-Dillanes, M. T., Huidobro-Campos, L. y Mejía-Mojica, H. (2014). Peces invasoras en el centro de México. En R. Mendoza y P. Koleff (Coords.), Especies acuáticas invasoras en México (pp. 413–424). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Contreras-MacBeath, T., Mejía, M. H. y Carrillo, W. R. (1998). Negative impact on the aquatic ecosystems of the state of Morelos from introduced aquarium and other commercial fish. Aquarium Sciences and Conservation, 2, 1–12.
Cossíos, E. D. (2010). Naturalized vertebrates in Peru: history and state of knowledge. Revista Peruana de Biología, 17, 179–189. https://doi.org/10.1023/A:1009676403693
Courtenay, W. R. Jr., Hensley, D. A., Taylor, J. N. y McCann, J. A. (1984). Distribution of exotic fishes in the continental United States. En W. R. Courtenay Jr. y J. R. Stauffer, Jr. (Eds.). Distribution, biology and management of exotic
fishes. Baltimore, Maryland: Johns Hopkins University Press.
David, P., Thebault, E., Anneville, O., Duyck, P. F., Chapuis, E. y Loeuille, N. (2017). Impacts of invasive species on food webs: a review of empirical data. Advances in Ecological Research, 56, 1–60. https://doi.org/10.1016/bs.aecr.2016.10.001
De la Maza-Benignos, M. (2014). Sistemática, zoogeografía y evolución del género Herichthys (Pisces: cichlidae), en el noreste de México (Tesis de doctorado). Universidad Autónoma de Nuevo León. San Nicolás de los Garza, N. L.
De la Maza-Benignos, M. y Lozano-Vilano, M. L. (2013). Description of three new species of the genus Herichthys (Perciformes: Cichlidae) from eastern Mexico, with redescription of H. labridens, H. steindachneri, and H. pantostictus. Zootaxa, 3734, 101–129. https://doi.org/10.11646/zootaxa.3734.2.1
Duffy, R., Snow, M. y Bird, C. (2013). The convict cichlid Amatitlania nigrofasciata (Cichlidae): first record of this non-native species in Western Australia Waterbodies. Records of the Western Australian Museum, 28, 7–12. https://doi.org/10.18195/ISSN.0312-3162.28(1).2013.007-012
Eldredge, L. G. (1994). Freshwater fishes. En L. G. Eldredge (Ed.), Perspectives in aquatic exotic species management in the Pacific Islands. Vol. 1. Introductions of commercially significant aquatic organisms to the Pacific Islands (pp. 73–84). New Caledonia, France: South Pacific Commission.
Ferreira, D. L., Guedes, G. H., da Silva, L. G. y Araújo, F. G. (2021). Resource partitioning among freshwater congeneric fishes (Loricariidae: Hypostomus): trophic, spatial, and temporal dimensions, Studies on Neotropical Fauna and Environment, 58, 622–632. https://doi.org/10.1080/01650521.2021.2010974
Flecker, A. S. (1992). Fish predation and the evolution of invertebrate drift periodicity: evidence from Neotropical Streams. Ecology, 73, 438–448.
Fricke, R., Eschmeyer, W. N. y Van der Laan, R. (2022). Eschmeyer’s catalog of fishes: genera, species, references. Recuperado el 25 de abril, 2022 de: https://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Gagliardi-Seeley, J. (2012). Dominance status reversal based on pair-bond formation, in the convict cichlid (Amatitlania nigrofasciata). Journal of Ethology, 30, 317–323. https://doi.org/10.1007/s10164-012-0329-x
Gee, J. y Northcote, T. (1963). Comparative ecology of two species of dace (Rhynchthys) in the Fraser River System, British Columbia. Journal of the Fisheries Research Board of Canada, 20, 105–118. https://doi.org/10.1139/f63-010
Gois, K. S., Pelicice, F. M., Gomes, L. C. y Agostinho, A. A. (2015). Invasion of an Amazonian cichlid in the Upper Paraná River: facilitation by dams and decline of a phylogenetically related species. Hydrobiologia, 746, 401–413. https://doi.org/10.1007/s10750-014-2061-8
Gozlan, R. E., Britton, J. R., Cowx, I. y Copp, G. H. (2010). Current knowledge on non-native freshwater fish introductions. Journal of Fish Biology, 76, 751–786. https://doi.org/10.1111/j.1095-8649.2010.02566.x
Grossman, G. D. (1986). Food partitioning in a rocky intertidal fish assemblage. Journal of Zoology, 1, 317–355. https://doi.org/10.1111/j.1096-3642.1986.tb00642.x
Gurgel-Lourenço, R. C., Pinto, L. M., Bezerra, L. A. V. y Sánchez-Botero, J. I. (2019). Has a non-native cichlid of the genus Amatitlania (Actinopterygii, Cichlidae) adapted to the headwaters in Brazilian semi-arid? River Research, 36, 1353–1359. https://doi.org/10.1002/rra.3612
Hammer, Ø., Harper, D. A. T. y Ryan, P. D. (2020). PAST. Paleontological statistics sofware pakage for education and data análisis. Paleontología Electrónica, 4, 1.
Hata, H. y Ochi, H. (2016). Depth and substratum differentiations among coexisting herbivorous cichlids in Lake Tanganyika. Royal Society Open Science, 3, 160229. https://doi.org/10.1098/rsos.160229
Herrera, R. G. A., Murcia-Castillo, M. A. y Prada-Pedreros, S. (2017). First record of Amatitlania nigrofasciata (Günther, 1867) (Cichliformes: Cichlidae) as introduced species in natural freshwaters of Colombia. Check List, 12, 1932. https://doi.org/10.15560/12.4.1932
Hesthagen, T., Saksgard, R., Hegge, O., Dervo, B. y Skurdal, J. (2004). Niche overlap between young brown trout (Salmo trutta) and Siberian sculpin (Cottus poecilopus) in subalpine Norwegian river. Hydrobiologia, 52, 117–125. https://doi.org/10.1023/B:HYDR.0000026354.22430.17
Ishikawa, T. y Tachihara, K. (2010). Life history of the nonnative convict cichlid Amatitilania nirgofasciata in the Haebaru Reservoir on Okinawa-jima Island, Japan. Environmental Biology of Fishes, 88, 283–292. https://doi.org/10.1007/s10641-010-9641-x
Itzkowitz, M., Vollmer, G. y Ríos-Carenas, O. (1998). Competition for nesting sites between monogamous pairs of convict cichlids (Cichlasoma nigrofasciatum): Asymmetries in size and prior residence. Behaviour, 135, 261–267. https://doi.org/10.1163/156853998793066294
Jowett, I. G. (2022). Analysis of habitat suitability data. Version 3.1 © Ian Jowett Consulting. Recuperado el 14 septiembre, 2022 de: http://www.jowettconsulting.co.nz/home/habprf
Jowett, I. G. y Davey, A. J. H. (2007). A comparison of composite habitat suitability indices and generalized additive models of invertebrate abundance and fish presence–habitat availability. Transactions of the American Fisheries Society, 136, 428–444. https://doi.org/10.1577/T06-104.1
Jowett, I. G. y Richardson, J. (1995). Habitat preferences of common, riverine New Zealand native fishes and implications for flow management. New Zealand Journal of Marine and Freshwater Research, 29, 13–23. https://doi.org/10.1080/00288330.1995.9516635
Jowett, I. G. y Richardson, J. (2008). Habitat use by New Zealand fish and habitat suitability models. NIWA Science and Technology Series, 5, 1–148.
Keith, P., Marquet, G., Valade, B. P. y Vigneux, E. (2006). Atlas des poissons et des crustacés d’eau douce des Comores, Mascareignes et Seychelles. Patrimoines naturels, 65. París: Muséum National d’Histoire Naturelle.
Kramer, D. L. y Bryant, M. J. (1995). Intestine length in the fishes of a tropical stream: 2. Relationships to diet-the long and short of a convoluted issue. Environmental Biology of Fishes, 4, 129–141. https://doi.org/10.1007/BF00001991
Krebs, C. J. (1999). Ecological methodology. San Mateo, CA.: Addison Wesley Longman.
Lombard, R. J., Chimimba, C. T. y Tsungai, A. Z. (2018). Niche complementarity between an alien predator and native omnivorous fish in the Wilge river, South Africa. Hydrobiologia, 817, 329–340. https://doi.org/10.1007/s10750-017-3352-7
Maitra, S., Harikrishnan, M. y Nidhin, B. (2019). Feeding strategy, dietary overlap, and resource partitioning among four mesopredatory catfishes of a tropical estuary. Journal of Fish Biology, 96, 130–139. https://doi.org/10.1111/jfb.14194
Makocho, P. y Stauffer, J. (2018). Territoriality and the coexistence between the introduced and the native mbuna of the Lake Malawi National Park. Modern Environmental Science and Engineering, 4, 556–569. https://doi.org/10.15341/MESE(2333-2581)/06.04.2018/010
Manly, B. F. J., Mc Donald, L. L., Thomas, D. L., McDonald, T. L. y Erickson, W. P. (1993). Resource selection by animals. London: Chapman and Hall.
Marsh, B. A. y Ribbink, A. J. (1985). Feeding site utilization in three sympatric species of Petrotilapia Pisces (Cichlidae) from Lake Malawi. Biological Journal of the Linnean Society, 25, 331–338. https://doi.org/10.1111/j.1095-8312.1985.tb00399.x
McKaye, K. R. (1977) Competition for breeding sites between the cichlid fishes of Lake Jiloa, Nicaragua. Ecology, 58, 291–302. https://doi.org/10.2307/1935604
McNeely, J. (2001). Invasive species: a costly catastrophe for native biodiversity. Land Use and Water Resources Research, 1, 1–10. https://doi.org/10.22004/ag.econ.47850
Medina-Nava, M., Schmitter-Soto, J. J., Mercado-Silva, N., Rueda-Jasso, R. A., Ponce-Saavedra, J. y Pérez-Munguía, R. M. (2011). Ecological guilds of fishes in streams of an arid subtropical drainage in western Mexico. Journal of Freshwater Ecology, 26, 579–592. http://dx.doi.org/10.1080/02705060.2011.626271
Mejía-Mojica, H., Contreras-MacBeath, T. y Ruiz-Campos, G. (2014). Relationship between environmental and geographic factors and the distribution of exotic fishes in tributaries of the Balsas River basin, Mexico. Environmental Biology of Fishes, 98, 611– 621. https://doi.org/10.1007/s10641-014-0298-8
Mejía-Mojica, H., Rodríguez-Romero, F. J. y Díaz-Pardo, E. (2012). Recurrencia histórica de peces invasores en la Reserva de la Biosfera Sierra de Huautla, México. Revista de Biología Tropical, 60, 669–681. https://doi.org/10.15517/rbt.v60i2.3960
Mendelson, J. (1975). Feeding relationships among species of Notropis (Pisces: Cyprinidae) in a Wisconsin stream. Ecological Monograph, 45, 199–230. https://doi.org/10.2307/1942422
Mendoza, R. y Koleff, P. (coords.). (2014). Especies acuáticas invasoras en México. Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Mendoza-Cariño, M., Bautista-Olivas, A. L., Quevedo-Nolasco, A. y Mendoza-Cariño, D. (2018). Análisis hidrológico de largo plazo en la cuenca del río Metztitlán Hidalgo, México y su relación con el cambio climático. Hidrobiológica, 28, 17–30. https://doi.org/10.24275/uam/izt/dcbs/hidro/2018v28n1/Mendoza
Miller, R. R., Minckley, W. L. y Norris, M. S. (2009). Peces dulceacuícolas de México. México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Sociedad Ictiológica Mexicana A.C./ El Colegio de la Frontera Sur/ Consejo de los Peces del Desierto México-Estados Unidos.
Miranda, R., Galicia, D., Monks, S. y Pulido-Flores, G. (2012). Diversity of freshwater fishes in Reserva de la Biosfera Barranca de Metztitlán, Hidalgo, Mexico, and recommendations for conservation. The Southwestern Naturalist, 57, 285–291. https://doi.org/10.1894/0038-4909-57.3.285
Molina, D., Arce, E. y Mercado-Silva, N. (2021). Mexican mojarra can dominate non-native convict cichlids even when outnumbered. Behavioral Ecology and Sociobiology, 75, 16. https://doi.org/10.1007/s00265-020-02963-5
Montaño-Campos, S. E. (2008). Conocimiento tradicional de los peces aprovechados en dos comunidades ribereñas del municipio de Calnalí, Hidalgo (Tesis). Universidad Autónoma del Estado de Hidalgo. Pachuca, Hidalgo.
Needham, J. G. y Needham, P. R. (1982). Guía para el estudio de los seres de las aguas dulces. Barcelona: Editorial Reverté, S.A.
Patterson, I. J. (1985). Limitation of breeding density through territorial behaviour: experiments with convict cichlids, Cichlasoma nigrofasciatum. En R. M. Sibly y R. H. Smith (Eds.), Behavioural ecology, ecological consequences of adaptive behaviour (pp. 393–405). Oxford: Blackwell Scientific Publications.
Pennak, R. (1978). Freshwater Invertebrates of the United States. New York: Wiley Interscience.
Pérez-Miranda, F., Mejía, O., Soto-Galera, E., Espinosa-Pérez, H., Piálek, L. y Rican, O. (2018). Phylogeny and species diversity of the genus Herichthys (Teleostei: Cichlidae). Journal Zoological Systematics Evolutionary Research, 56, 223–247. https://doi.org/10.1111/jzs.12197
Pérez-Miranda, F., Mejía, O., Zuñiga, G., Soto-Galera, E. y Rican, O. (2019). Feeding ecomorphologies in the fish genus Herichthys (Perciformes: Cichlidae) based on stomach content and lower pharyngeal jaw shape. Revista de Biología Tropical, 67, 643–653. https://doi.org/10.15517/rbt.v67i3.33616
Pianka, E. R. (1973). The Structure of Lizard Communities. Annual Review of Ecology and Systematics, 4, 53–74. https://doi.org/10.1146/ANNUREV.ES.04.110173.000413
Pineda-López, R., Díaz-Pardo, E. y Martínez, M. (2009). Biota acuática de arroyos y ríos: cuencas Lerma-Chapala y Pánuco. Manual de identificación. Querétaro: Universidad Autónoma de Querétaro.
Reid, S. M. (2019). Summer microhabitats use and overlap by the invasive Round Goby (Neogobius melanostomus) and native darters in the Trent River (Ontario, Canada). Knowledge & Management of Aquatic Ecosystems, 420, 23. https://doi.org/10.1051/kmae/2019021
Reinas, G. C. Z., Silva, J. C. y Blaletzki, A. (2022). Coexistence patterns between native and exotic juvenile Loricariidae in a Neotropical floodplain: an approach to resource partitioning. Hydrobiologia, 849, 1713–1726. https://doi.org/10.1007/s10750-022-04816-w
Reza-Esmaeili, H., Gholamifard, A., Sayyadzadeh, G., Parsi, B., Mirghiasi, S. y Ghasemian, S. (2013). New record of the convict cichlid, Amatitlania nigrofasciata (Günther, 1867), from the Middle East (Actinopterygii: Cichlidae). Aqua International Journal of Ichthyology, 19, 4–19.
Rícan, O., Pialek, L., Dragová, K. y Novák, J. (2016). Diversity and evolution of the Middle American cichlid fishes (Teleostei: Cichlidae) with revised classification. Vertebrate Zoology, 66, 1–102. https://doi.org/10.3897/vz.66.e31534
Roberts, C. G. y Britton, J. R. (2018). Trophic interactions in a lowland river fish community invaded by European barbel Barbus (Actinopterygii, Cyprinidae). Hydrobiologia, 819, 259–273. https://doi.org/10.1007/s10750-018-3644-6
Sánchez-Barrera, E. (2010). Comparación de la ictiofauna de dos localidades del río Huzalingo, Hidalgo (Tesis). Universidad Autónoma del Estado de Hidalgo. Pachuca, Hidalgo.
Schmitter-Soto, J. J. (2007). A systematic revision of the genus Archocentrhus (Perciformes: Cichlidae), with the description of two new genera and six new species. Zootaxa, 1603, 1–78. https://doi.org/10.11646/zootaxa.1603.1.1
Scoppettone, G. G., Rissler, P., Johnson, D. y Hereford, M. (2011). Relative abundance and distribution of fishes and crayfish at Ash Meadows National Wildlife Refuge, Nye County, Nevada, 2007-08. Open-File Report. Seattle: Geological Survey. https://doi.org/10.3133/ofr20111017
Seehausen, O. y Bouton, N. (1997). Microdistribution and fluctuations in niche overlap im a rocky cichlid community in Lake Victoria. Ecology of Freswater Fish, 6, 161–173. https://doi.org/10.1111/j.1600-0633.1997.tb00159.x
Simberloff, D. (2013). Invasive species: what everyone needs to know. New York: Oxford University Press.
Soto-Galera, E. y Alcántara-Soria, L. (2016). Preferencia de hábitat de los peces en riesgo del río Santa María, territorio queretano. En R. Jones y C. R. Cárdenas (Eds.), Historia natural de Querétaro (pp. 353–355). Querétaro: Universidad Autónoma de Querétaro.
Soto-Galera, E., Alcántara-Soria, L. y Paulo-Maya, J. (2018). Estado actual, identificación de riesgos y propuestas para manejo de las especies invasoras presentes en las regiones hidrológicas prioritarias, confluencias de las Huastecas, Media Luna y Cabecera del río de La Laja. Instituto Politécnico Nacional. Escuela Nacional de Ciencias Biológicas. Informe final SNIB-Conabio, Proyecto No. LI038. Ciudad de México. Recuperado el 22 septiembre, 2022 de http://www.conabio.gob.mx/institucion/proyectos/resultados/InfLI038.pdf.
Takács, P., Czeglédi, I., Ferincz. Á., Sály, P., Specziár, Á., Vitál, Z. et al. (2017). Non-native fish species in Hungarian waters: historical overview, potential sources, and recent trends in their distribution. Hydrobiologia, 795, 1–22. https://doi.org/10.1007/s10750-017-3147-x
Tamaris-Turizo, C., Rodríguez-Barrios, J. y Ospina-Torres, R. (2013). Deriva de macroinvertebrados acuáticos a lo largo del río Gaira, vertiente noroccidental de la sierra Nevada de Santa Marta, Colombia. Caldasia, 35, 149–163.
Trujillo-Jiménez, P. (1996). Ecología alimentaria de Cichlasoma (Archocentrus) nigrofasciatum (Pisces: Cichlidae) en el río Amacuzac, Morelos. Zoología Informa, 33, 13–32.
Trujillo-Jiménez, P. (1998). Dinámica trófica de la ictiofauna del río Amacuzac, Morelos (Tesis de maestría). Universidad Nacional Autónoma de México. México D.F.
Weber, P. G. y Weber, S. P. (1971). Choice of spawn site in Cichlasoma nigrofasciatum (Gunther) (Pisces: Cichlidae). Zeitschrift fur Tierpsychologie, 28, 475–478. https://doi.org/10.1111/j.1439-0310.1971.tb00830.x
Welcomme, R. L. (1988). International introductions of inland aquatic species. FAO Fisheries Technical Paper, 294. Rome, Italy: FAO.
Williamson, M. (1996). Biological invasions. New York: Chapman and Hall, International Thomson Publishing.
Wisenden, B. D. (1994). Factors affecting reproductive success in free-ranging convict cichlids (Cichlasoma nigrofasciatum). Canadian Journal of Zoology, 72, 2177–2185. https://doi.org/10.1139/z94-29
Xiong, W., Sui, X., Liang, S. H y Chen, Y. (2015). Non-native freshwater fish species in China. Reviews in Fish Biology and Fisheries, 25, 651–687. https://doi.org/10.1007/s11160-015-9396-8
Yamamoto, M. N. y Tagawa, A. W. (2000). Hawai’i’s native and exotic freshwater animals. Honolulu: Mutual Publishing.
Zengeya, T. A. y Marshall, B. E. (2007). Trophic interrelationships amongst cichlid fishes in a tropical African reservoir (Lake Chivero, Zimbawe). Hydobiologia, 592, 175–182. https://doi.org/10.1007/s10750-007-0790-7


