Potencial reproductivo de Stenocerus queretaroensis (Cactaceae) de San José de Cosalima, Zacatecas
Oliva Solís-Márquez a, F. Ofelia Plascencia-Escalante a, *, Angélica Romero-Manzanares b, Juan Antonio Cruz-Rodríguez c, Gregorio Ángeles-Pérez a, Juan Carlos López-Acosta d y Petra Yáñez- Jiménez b
a Posgrado en Ciencias Forestales, Colegio de Postgraduados, Km. 36.5 carretera México-Texcoco, 56230 Montecillo, Texcoco, Estado de México, México
b Posgrado en Botánica, Colegio de Postgraduados, Km. 36.5 carretera México-Texcoco, 56230 Montecillo, Texcoco, Estado de México, México
c Departamento de Agroecología, Universidad Autónoma Chapingo, Km 38.5 carretera México-Texcoco, 56230 Montecillo, Texcoco, Estado de México, México
d Centro de Investigaciones Tropicales, Universidad Veracruzana, Casco de la Ex-hacienda Lucas Martín, Privada de Araucarias s/n, Col. Periodistas, 91019 Xalapa, Veracruz, México
*Autor para correspondencia: fplascen@colpos.mx (F.O. Plascencia-Escalante)
Resumen
Se estudió el potencial reproductivo de una población de Stenocereus queretaroensis, localizada en San José de Cosalima, municipio Tabasco, Zacatecas. Los objetivos fueron: estudiar la variación temporal en la producción de flores, frutos y semillas durante 2 años en 3 parajes; determinar la viabilidad y germinación de la semilla; analizar el efecto de un tratamiento pregerminativo en la eficiencia y velocidad de la germinación, y cuantificar el porcentaje de emergencia y supervivencia de plántulas en campo. De febrero de 2014 a julio de 2015 se siguió la fenología reproductiva del pitayo. Se encontraron diferencias en la producción de flores, frutos y semillas entre parajes y años de evaluación. Se observaron diferencias estadísticas significativas en la viabilidad y germinación de las semillas, así como en la emergencia de plántulas entre parajes y microhábitats diferenciados por tipo de cobertura de dosel. Los resultados de este estudio mostraron que el potencial reproductivo de S. queretaroensis se incrementó en el año de mayor precipitación (2015), con el uso de un tratamiento pregerminativo y bajo el dosel vegetal de la propia especie. Estos resultados pueden ser aplicados en programas de aprovechamiento y conservación del pitayo in situ.
Palabras clave:
Conservación; Emergencia de establecimiento; Fenofases reproductivas; Germinación; Pitayo; Viabilidad
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Richness, geographic distribution and conservation of Lycianthes series Meizonodontae (Capsiceae, Solanaceae)
Abstract
The reproductive potential of a population of Stenocereus queretaroensis was studied in San José de Cosalima, Municipality Tabasco, Zacatecas. The objectives were: to study the temporal variation in flower, fruit and seed production during 2 years in 3 sites; to determine the viability and germination of the seeds; to analyze the effect of a pregerminative treatment on the efficiency and speed of seed germination, and to quantify the emergence and survival percentages of seedlings in the field. From February 2014 to July 2015, the reproductive phenology of pitayo was evaluated. Differences in flower, fruit and seed production were found between sites and the evaluation times. Production of reproductive structures was higher in 2015 than 2014. Significant statistical differences were observed in viability and seed germination, likewise in the emergence of seedlings between sites and microsites differentiated by canopy cover type, being favorable the canopy of Celtis pallida and of the species itself. The results of this study showed that the reproductive potential of S. queretaroensis increased in the year with the greatest precipitation (2015), as well as with the use of a pregerminative treatment, and under the canopy of the species itself. These results can be applied in programs of use and conservation of the pitayo in situ.
Keywords:
Conservation; Emergence recruitment; Reproductive phenophases; Germination; Pitayo; Viability
Introducción
El potencial reproductivo indica la capacidad que tiene una población para producir descendencia viable y contribuir eventualmente al crecimiento poblacional (Marshall et al., 1998; Trippel, 1999) y se estima mediante la producción de flores, frutos y semillas, aunque la reproducción es un proceso que involucra a la floración, el número de flores polinizadas, el número de óvulos fertilizados, la producción de frutos y semillas, la viabilidad y germinación de las semillas (Bowers, 1998; Gaol y Fox, 2002). El potencial reproductivo se expresa al máximo al cumplirse ciertas condiciones físicas y biológicas, como que el clima sea adecuado, que el suelo sea rico en nutrimentos, que la planta parental tenga suficiente tamaño/edad (Tremblay, 2006) y haya madurado para ejercer la reproducción, que la duración del período de crecimiento sea extenso al igual que el ciclo de vida, y que haya presencia y efecto de los agentes polinizadores.
Los estudios relacionados con el potencial reproductivo contribuyen significativamente al conocimiento de la biología reproductiva y de la dinámica poblacional de las especies, además, resultan ser importantes indicadores para la conservación y uso sustentable de los recursos (Rivas y Barilani, 2004), principalmente en aquellas especies altamente susceptibles a los disturbios naturales y perturbaciones antrópicas. Ejemplo de ello son las cactáceas, que presentan un alto grado de endemismo, tienen un crecimiento lento y la incorporación de nuevos individuos en las poblaciones es esporádica, características que las hacen vulnerables (Godínez-Álvarez et al., 2003; Jiménez, 2011; Martínez-Ramos et al., 2016).
El potencial reproductivo en cactáceas se ha estudiado desde la perspectiva del análisis de la dinámica poblacional (Nassar y Emaldi, 2008) y se ha demostrado que las condiciones ambientales del hábitat son determinantes para la persistencia de esta familia (Figueroa-Castro y Valverde, 2011; Godínez-Álvarez et al., 2003; León-de la Luz y Domínguez, 1991). Ciertamente, la propagación vegetativa (ramets y genets) es un aspecto biológico que repercute favorablemente en la dinámica poblacional de éstas (Ferrer et al., 2011).
En este sentido, conocer el potencial reproductivo de las cactáceas, particularmente de aquellas especies que han sido diezmadas de sus comunidades, se vuelve una poderosa herramienta para la toma de decisiones para su aprovechamiento y conservación, como es el caso de Stenocereus queretaroensis (F.A.C.Weber) F. Buxb., una cactácea arborescente, endémica de México (Granados et al., 1999; Jiménez, 2011; Rojas-Aréchiga y Vázquez-Yanes, 2000 ), con amplia distribución en el centro-occidente del país (Ibarra-Cerdeña et al., 2005; Pimienta, 1999a; Salcedo, 1991 ), pero citada con afectaciones a sus poblaciones silvestres, principalmente por la actividad humana (Arriaga et al., 2000; Cabrera y Gómez, 2005; Pimienta, 1999b).
El hábitat de S. queretaroensis es la selva baja caducifolia y el matorral xerófilo (Arreola-Nava y Terrazas, 2003; Balleza y Villaseñor, 2002; Casas, 2002). El pitayo, conocido así popularmente, es una especie de alta importancia económica, social (Arriaga-Ruiz et al., 2015; Ibarra-Cerdeña et al., 2005) y ecológica (Bustamante y Búrquez, 2005; Hernández, 2002), ya que sus frutos son comestibles y altamente comercializados en las regiones donde se producen, además de que son fuente de alimento para la fauna silvestre.
En el sur del estado de Zacatecas, las poblaciones silvestres de S. queretaroensis han disminuido con el probable riesgo para su persistencia, principalmente, a causa de los efectos de la cría de ganado bovino y del cambio de uso de suelo para la agricultura (Pimienta, 1999a). En este sentido, el estudio del potencial reproductivo de S. queretaroensis en el área, permitirá tener las bases científicas para contribuir a la preservación de este recurso fitogenético, mediante la aplicación del conocimiento biológico y ecológico.
Destaca entre la literatura, la falta de información que conjunte evaluaciones acerca del potencial reproductivo del pitayo y el proceso de establecimiento en campo, por lo que el objetivo general del presente trabajo fue: cuantificar y comparar el potencial reproductivo de una población de S. queretaroensis en 3 parajes de estudio en la comunidad de San José de Cosalima, Tabasco, Zacatecas, y entre 2 años de observación (2014 y 2015). Específicamente, se planteó: 1) estudiar la variación temporal en la producción de flores, frutos y semillas durante 2 años de observación y en 3 parajes de estudio, 2) determinar la viabilidad y germinación de las semillas, 3) analizar el efecto del hipoclorito de sodio como tratamiento pregerminativo en la proporción y velocidad de la germinación y 4) cuantificar el porcentaje de emergencia y supervivencia de plántulas en campo y de las condiciones que facilitan este
proceso.
Materiales y métodos
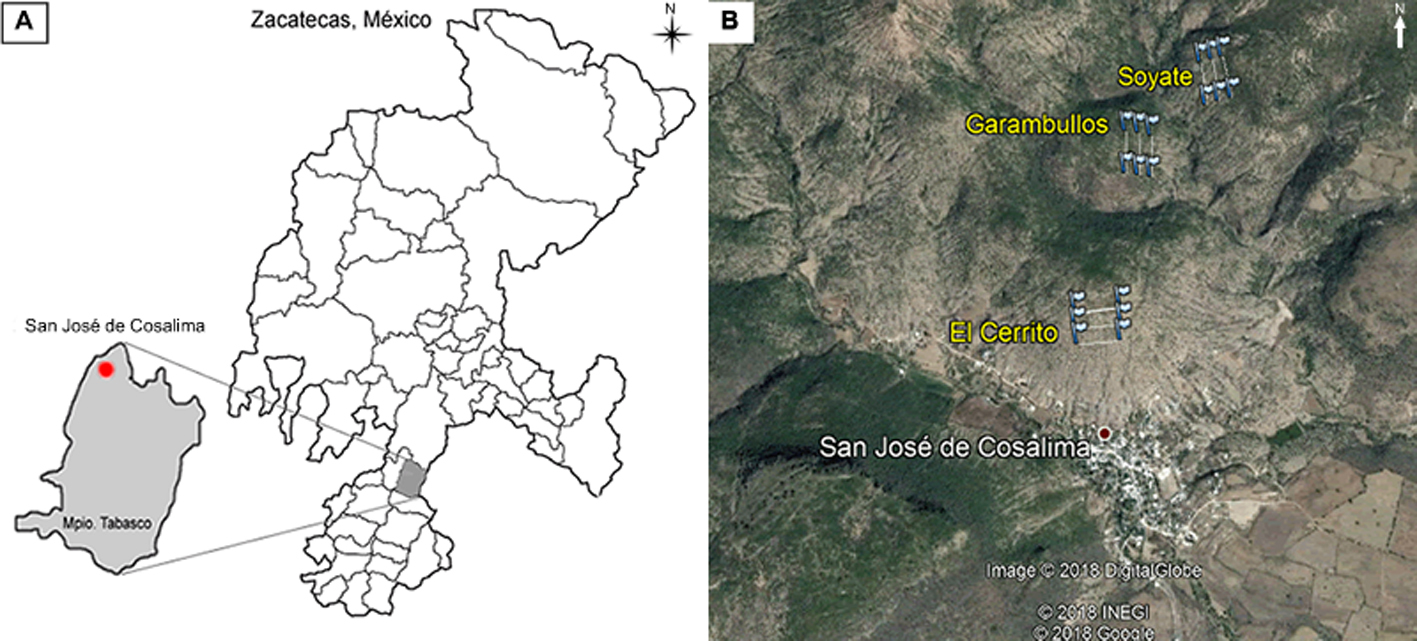
El estudio se realizó en la población silvestre de S. queretaroensis localizada en San José de Cosalima, municipio de Tabasco, Zacatecas (22°00’36” N, 102°58’13” O; 1,844 m snm) (fig. 1A). El clima de la región de acuerdo con la clasificación de Köppen, modificada por García (2004), es BSw, que se describe como un semiseco con lluvias en verano. La precipitación y temperatura media anual en el año 2014, con base en los datos de la estación meteorológica más cercana (El Chique-00032013) fueron de 593.80 mm y 20.99 °C, respectivamente, mientras que para el año 2015, la precipitación fue de 942 mm y una temperatura media anual de 21.54 °C (Conagua-SMN, 2016).
El suelo predominante es de tipo Feozem, presente en las partes bajas donde se realiza la agricultura de temporal, caracterizado por tener una capa superficial de suelo de color oscuro, rico en materia orgánica. En las partes altas de cerros y laderas escarpadas hasta de 45° de pendiente, donde se establecen los pitayos, predomina el Leptosol, un suelo con profundidad menor a los 15 cm, arenoso con poca o nula cantidad de suelo fino, originado de material parental ígneo extrusivo (Inegi, 2004, 2010). La vegetación está conformada por una ecotonía florística tropical seca, compuesta por especies compartidas tanto del bosque tropical caducifolio como del matorral xerófilo (Rzedowski, 1978). Destacan por su abundancia, especies como Celtis pallida Torr., y cactáceas como S. queretaroensis, Myrtillocactus geometrizans (Mart. ex Pfeiff.) Console y diferentes especies de Opuntia spp. (González, 2012).
Se eligieron 3 parajes dentro de la población silvestre de S. queretaroensis (fig. 1B): El Cerrito, Garambullos y Soyate. Cada paraje tuvo diferente orientación del relieve y distancia del centro de la comunidad de San José de Cosalima. El paraje El Cerrito tiene una orientación sur y se encuentra a 0.6 km de distancia de la comunidad; Garambullos tiene una orientación este y se encuentra a una distancia de 1.7 km; Soyate tiene una orientación oeste y se encuentra situado a una distancia de 2.7 km. Se asume que el tipo de suelo y la vegetación presente en los parajes de estudio es uniforme, ya que se observó una misma topografía y una composición similar de especies vegetales.
En cada paraje se establecieron 3 transectos, cada uno con una dimensión de 20 m de ancho y 200 m de longitud (4,000 m2). Los transectos se establecieron de forma paralela, con una separación de 60 m (fig. 1B). Dentro de los transectos, se marcaron todos los individuos de S. queretaroensis que presentaron estructuras reproductivas.
Durante las fenofases de floración y fructificación de S. queretaroensis (febrero a julio) de los años 2014 y 2015, se realizaron recuentos del número total de yemas florales, flores en antesis y frutos. Los recuentos se realizaron cada 7 días.
Con base en los recuentos por fenofase, se estimó por diferencia (tiempo 2 – tiempo 1) el número de flores y frutos producidos por mes en cada paraje. Posteriormente, se comparó la distribución temporal de estos números dentro del mismo año de observación, mediante una tabla de contingencia, trabajada con el programa R (R-Development Core Team, 2015). El mismo análisis se realizó para comparar la distribución de estructuras de cada fenofase por paraje entre los años 2014 y 2015. De lo anterior, se identificaron los períodos de mayor y menor cantidad de estructuras reproductivas para cada fenofase por paraje y año de observación.
En el año 2014, se tomó una muestra de 44 frutos maduros, 35 correspondientes al paraje Garambullos y 9 correspondientes a Soyate. Los frutos recolectados provenían de 15 árboles seleccionados al azar. En el año 2015, se tomó una muestra de 10 frutos maduros por paraje; los frutos también correspondieron a 15 árboles seleccionados al azar. Los frutos recolectados fueron disectados para obtener las semillas, las cuales fueron aisladas del mucílago por medio de lavado con agua y con ayuda de un cernidor metálico. Posteriormente, las semillas se secaron a temperatura ambiente durante un periodo de 30 días, se contaron y almacenaron en bolsas de papel.
El número de semillas por fruto en el año 2014, se comparó con la prueba de t de Student mediante el programa Statistical Analysis Software (SAS, 2002). El número de semillas por fruto colectado en el año 2015, se comparó con un Anova y una prueba de comparación de medias de Tukey. El número de semillas por fruto por paraje en los 2 años de observación se comparó con una prueba de t de Student (Zar, 1988).
Se realizó la prueba de viabilidad de las semillas colectadas en los años 2014 y 2015. Para cada año de observación, se utilizaron 3 lotes de 50 semillas por paraje. Cada lote de semillas se mantuvo sobre papel filtro hidratado dentro de una caja de Petri durante 18 horas. Posteriormente, se retiró la testa y los embriones se depositaron en una solución de tetrazolio a una concentración de 1%, durante un periodo de 18 horas. Con un microscopio estereoscópico se realizó la observación y clasificación de los embriones. Se consideró semilla viable aquella donde el embrión presentó tinción fuerte en la mayor parte de sus tejidos, principalmente en el hipocótilo y en la radícula. La proporción de la viabilidad de la semilla por paraje y año de observación se comparó con un modelo logístico mediante el programa SAS (MacDonald, 2014).
La prueba de germinación se realizó con semillas colectadas en el año 2014. Se utilizaron 6 lotes (100 semillas c/u) por paraje (Garambullos y Soyate); 3 de estos lotes correspondientes a cada paraje se trataron con hipoclorito de sodio al 10% durante 5 minutos, con la finalidad de propiciar la oxidación de la materia orgánica (lípidos) y dar pauta a la presencia de oxígeno y agua dentro de la semilla (Barros y Killian, 2013). Los 3 lotes restantes de cada paraje permanecieron sin tratamiento. Cada lote de semillas se colocó sobre papel filtro en cajas de Petri previamente esterilizadas y se aplicó agua destilada hasta saturar el papel. Las cajas de Petri se colocaron en una germinadora sin control de fotoperiodo a una temperatura de 21.5 ºC (±1). La humedad se reguló de forma manual, aplicando 1.5 ml de agua destilada cada tercer día. Se consideró semilla germinada la que mostró emergencia de la radícula. El periodo de observación fue de 31 días, debido a que al llegar a ese tiempo los porcentajes de germinación se estabilizaron.
Para determinar el efecto de la procedencia de la semilla y la influencia del tratamiento pregerminativo, se construyó un modelo de regresión logística (Zar, 1988). Mediante las técnicas del análisis de supervivencia (Allison, 1995) se construyeron y compararon curvas de la dinámica de germinación, para lo cual se utilizó el procedimiento LIFETEST (SAS, 2015).
Para evaluar la emergencia y supervivencia de plántulas de S. queretaroensis en campo se realizó un experimento factorial que consideró, además de los 3 parajes de estudio, 3 microhábitats (bajo dosel de S. queretaroensis, bajo dosel de C. pallida, y espacio sin dosel vegetal), y un tratamiento pregerminativo de las semillas y un testigo (sin tratamiento). Cada una de las 18 combinaciones tuvo 3 repeticiones. Cada unidad experimental fue un cuadro de 0.5 m2 en el que se colocó un lote de 100 semillas.
Las plántulas emergidas en cada unidad experimental fueron marcadas y se les dio seguimiento en el proceso de supervivencia. Las observaciones se realizaron a intervalos de 7 días durante los primeros 3 meses del experimento, posteriormente se evaluaron cada 15 días hasta finalizar el mismo (agosto 2014-agosto 2015).
La emergencia de las plántulas se analizó con un modelo de regresión logística (Zar, 1988) mediante el programa SAS. La dinámica de supervivencia se analizó con una curva de supervivencia mediante análisis LIFEREG (SAS, 2015). En ambos análisis se emplearon paraje, microhábitat y tratamiento pregerminativo como las variables explicativas.
Resultados
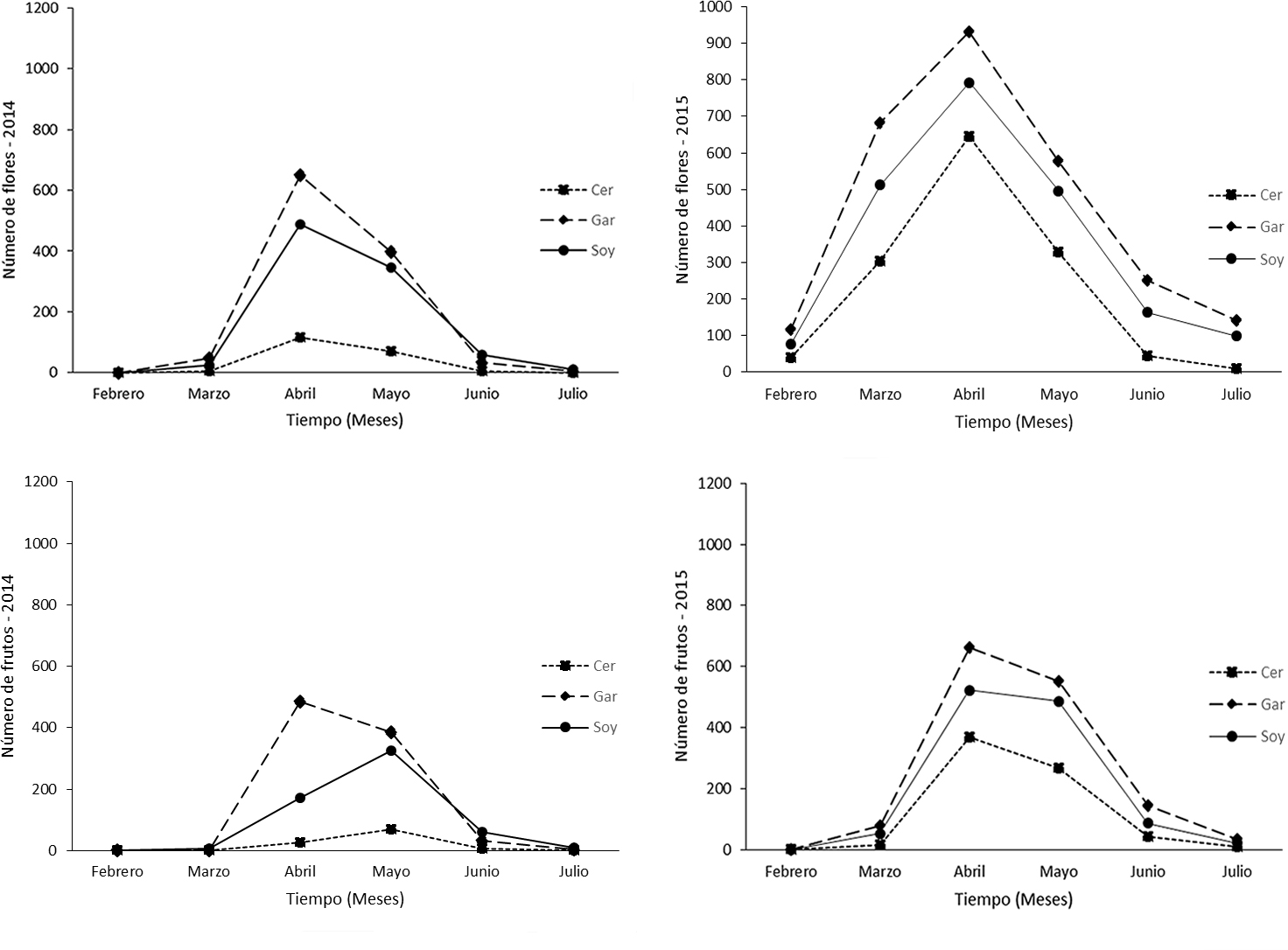
El número total de individuos reproductivos por paraje fue de 16 para El Cerrito, 36 para Garambullos y 25 para Soyate. Para el año 2014, la distribución temporal de la abundancia de estructuras reproductivas en cada fenofase, fue dependiente del paraje (floración: χ2 = 32.97, g.l. = 10, p ≤ 0.0001; fructificación: χ2 = 122.84, g.l. = 10, p ≤ 0.0001). Lo mismo ocurrió para el año 2015 (floración: χ2 = 172.70, g.l. = 10, p ≤ 0.0001; fructificación: χ2 = 39.36, g.l. = 10, p ≤ 0.0001). En los 2 años de observación, el paraje Garambullos presentó mayor acumulación de flores y frutos, seguido del paraje Soyate y El Cerrito.
Para el paraje El Cerrito, la distribución temporal de la abundancia de estructuras reproductivas en cada fenofase, fue diferente y mayor en el año 2015 con respecto al año 2014 (floración: χ2 = 62.27, g.l. = 5, p ≤ 0.0001; fructificación: χ2 = 44.74, g.l. = 5, p ≤ 0.0001). Lo mismo ocurrió para los parajes Garambullos y Soyate (floración: χ2 = 581.97, g.l. = 5, p ≤ 0.0001; fructificación: χ2 = 112.90, g.l. = 5, p ≤ 0.0001; floración: χ2 = 371.20, g.l. = 5, p ≤ 0.0001; fructificación: χ2 = 68.59, g.l. = 5, p ≤ 0.0001, respectivamente). En el año 2014, la producción de flores y frutos para El Cerrito, fue de 200.8 y 102.5, respectivamente, y en el año 2015 fue de 1,369 y 704. Para Garambullos, fue de 1,130.8 flores y 910.8 frutos en el 2014, y de 2,697 y 1,472 en el 2015. Para Soyate la producción de flores y frutos en 2014 fue de 926.6 y 574.1, mientras que en el año 2015, fue de 2,140 y 1,168 respectivamente.
El valor máximo del número de flores en los años 2014 y 2015 para los 3 parajes de estudio, se presentó en el mes de abril. El valor máximo del número de frutos para el año 2014, también se presentó en abril para el paraje Garambullos, y en mayo para los parajes El Cerrito y Soyate. Para el año 2015, la producción máxima de frutos ocurrió en abril para los 3 parajes de estudio (fig. 2).
El número promedio de semillas por fruto colectados en el año 2014 fue diferente entre los parajes Garambullos y Soyate (t de Student = 3.025; g.l. = 42; p = 0.004), los cuales presentaron un promedio (± e. e) de semillas por fruto de 1,050.8 (± 60.15) y 681.5 (± 49.31), respectivamente. En el año 2015, el promedio de semillas por fruto también presentó diferencias estadísticas significativas entre parajes (F = 18; g.l. = 2, 27; p < 0.0001), siendo mayor para el paraje Garambullos (1286 ± 94.77) (Tukey; p < 0.05) en comparación con los otros 2 parajes; Soyate (851.1 ± 106.69) y El Cerrito (515.3 ± 67.52).
La comparación del número promedio de semillas por fruto en el año 2014 y 2015 de acuerdo al paraje, no presentó diferencia estadística significativa para el paraje de Garambullos (t de Student = 1.91; g.l. = 43; p = 0.063), ni para el paraje Soyate (t de Student = 1.39; g.l. = 17; p = 0.1827).
La viabilidad de las semillas colectadas en el año 2014, fue mayor en el paraje Garambullos en comparación con Soyate (χ2 de Wald = 29.81; g.l. = 1; p < 0.0001), con porcentajes de viabilidad del 44.66 y 14.66%, respectivamente. Para las semillas colectadas en el año 2015, la viabilidad también presentó diferencias estadísticas significativas entre parajes (χ2 de Wald = 77.40; g.l. = 2; p < 0.0001). Las semillas de El Cerrito y Garambullos tuvieron porcentajes de viabilidad de 71.33 y 76.66%, respectivamente, mientras que las del paraje Soyate tuvieron un 28.66%.
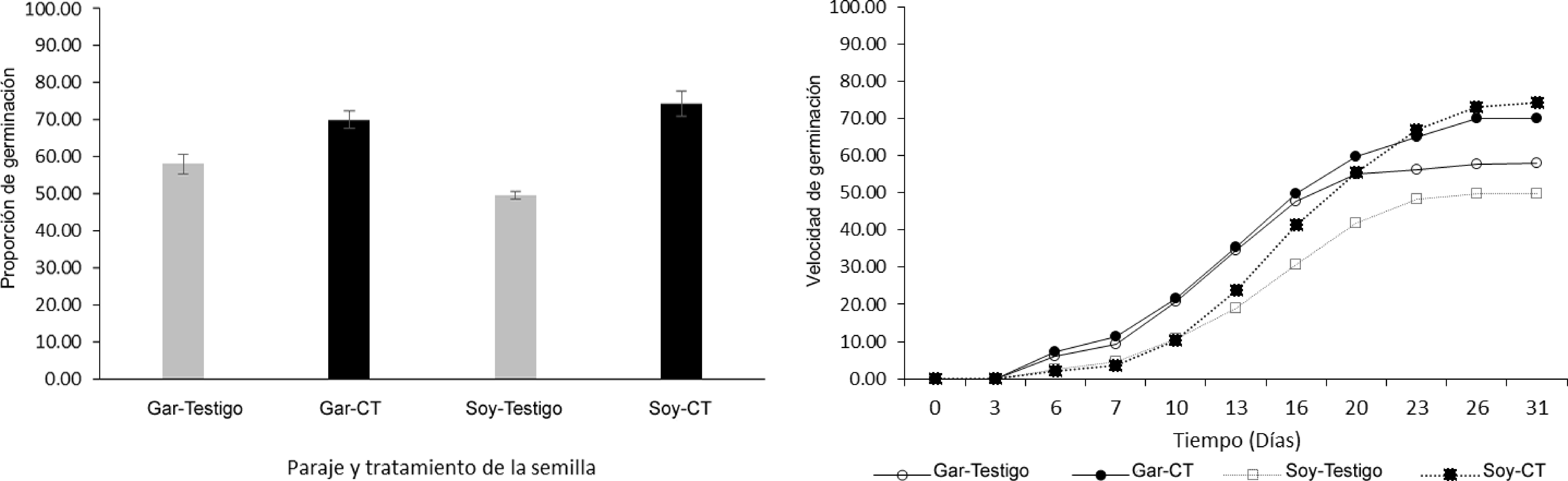
La germinación inicio a los 6 días después de la siembra. Las semillas procedentes del paraje Garambullos (semillas con y sin tratamiento pregerminativo), así como la semilla procedente de Soyate (con tratamiento pregerminativo), alcanzaron el 50% de germinación a los 20 días del experimento (fig. 3). La variable que fue significativa en relación con la germinación fue el tratamiento pregerminativo de las semillas (β = 0.799; χ2 de Wald = 42; g.l. = 1; p < 0.0001). Los porcentajes finales de germinación fueron mayores en semillas tratadas con hipoclorito de sodio en ambos parajes (Garambullos: 70%, Soyate: 74.33%), en comparación con las que no recibieron tratamiento (Garambullos: 58%, Soyate: 49.67%). La dinámica de germinación de las semillas de S. queretaroensis fue diferente entre tratamiento (χ2 = 32.27; g.l. = 3; p < 0.0001).
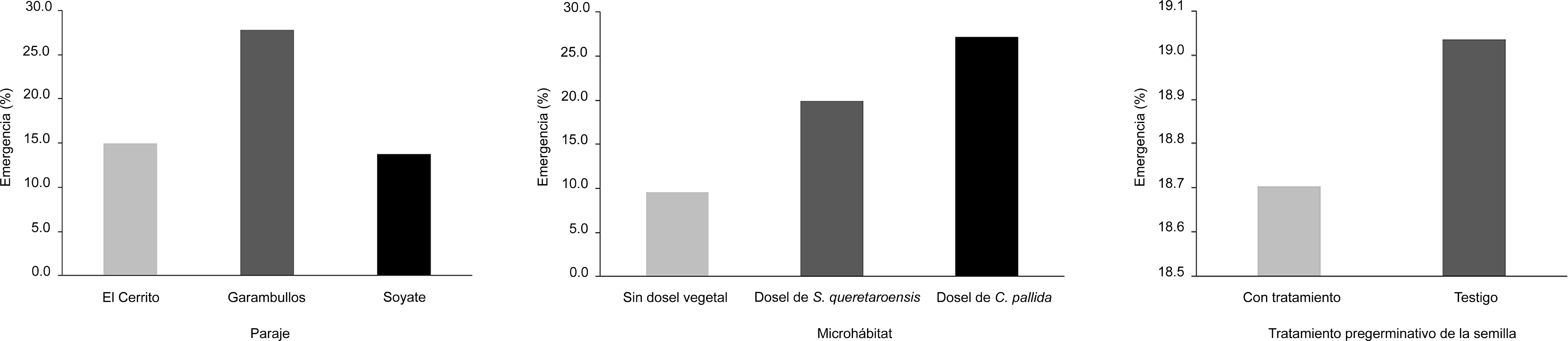
La emergencia de las plántulas inicio en agosto de 2014 y concluyó en diciembre del mismo año. El periodo en el cual se presentó la máxima emergencia de plántulas fue entre agosto y octubre. El mayor porcentaje de emergencia se presentó en septiembre. De acuerdo al paraje, Garambullos tuvo el mayor porcentaje de emergencia (27.8%), seguido del paraje El Cerrito (15%) y Soyate (13.8%). De acuerdo al microhábitat, el mayor porcentaje de emergencia ocurrió bajo el dosel de C. pallida (27.2%), seguido de bajo el dosel de S. queretaroensis (19.9%) y en espacios sin dosel vegetal (9.6%). En relación con el tratamiento pregerminativo de las semillas, los mayores valores de emergencia se tuvieron en semillas que no recibieron tratamiento (19.0%), en comparación con semillas tratadas (18.7%). De acuerdo con el modelo de regresión logística, las variables que fueron significativas en torno a los porcentajes finales de emergencia en campo fueron; el paraje y microhábitat (p < 0.0001). Para el caso de la variable paraje, en Garambullos se presentó un incremento positivo significativo en la emergencia (β = 0.9156; χ2 = 108.356; g.l. = 1; p < 0.0001). Para la variable microhábitat, el dosel de S. queretaroensis (β = 0.874; χ2 = 75.358; g.l. = 1; p < 0.0001) y el dosel de C. pallida (β = 1.295; χ2 = 176.86; g.l. = 1; p < 0.0001) incrementaron de manera significativa la emergencia de las plántulas (fig. 4).
Los porcentajes finales de supervivencia fueron de 2.8% para Garambullos, 4.8% para El Cerrito y 5.2% para Soyate. De acuerdo al microhábitat, bajo el dosel de C. pallida se tuvo una supervivencia del 2.9%, bajo el dosel de S. queretaroensis fue del 6.7% y en espacios sin cubierta vegetal fue de 1.2%. Se tuvo supervivencia del 5.5% para plántulas cuyas semillas fueron pretratadas y 2.3% de supervivencia en plántulas originadas de semillas sin tratar.
De acuerdo con el análisis de la dinámica de supervivencia con el procedimiento LIFEREG, el modelo indicó que las 3 variables (paraje, microhábitat y tratamiento de la semilla) tuvieron efectos significativos en los tiempos de supervivencia (p ≤ 0.0193). En el primer caso, en el paraje El Cerrito se presentó un incremento significativo en el tiempo de supervivencia (β = 0.3667; χ2 = 8.37; g.l. = 1; p = 0.0038). Para el caso del microhábitat, el dosel de S. queretaroensis tuvo efecto significativo en el tiempo de supervivencia de las plántulas (β = 0.2907; χ2 = 7.11; g.l. = 1; p = 0.0077). Para el caso del tratamiento pregerminativo, las semillas previamente tratadas con hipoclorito de sodio tuvieron un efecto positivo significativo en el tiempo de supervivencia (β = 0.2484; χ2 = 5.48; g.l. = 1; p = 0.0193).
Discusión
La diferencia en la temporalidad de la floración y la fructificación de S. queretaroensis en los distintos parajes de estudio en el año 2014, en el cual, el paraje Garambullos presentó mayor producción de estructuras reproductivas, seguido del Soyate y por último El Cerrito, podría estar relacionado de manera simple con el número de individuos reproductivos presentes en cada lugar; en donde ciertamente en el paraje Garambullos se registró el mayor número, seguido de Soyate, y con menor número El Cerrito. Este mismo patrón de abundancia de estructuras reproductivas se tuvo en el año 2015.
Otro posible factor en torno a la diferencia en la distribución temporal de la abundancia de estructuras reproductivas, puede estar relacionado con la exposición del relieve propio de cada paraje. Socolowski et al. (2010) encontró para Cereus pernambucensis Lem. que la luz y la temperatura representaban 2 aspectos importantes en la fructificación, así como la importancia del fotoequilibrio del fitocromo para el desarrollo y la germinación de las semillas. Reubicando esta información a la situación de campo, la orientación este del paraje Garambullos en comparación con la orientación oeste del Soyate, puede ser lo que le confiere ventajas al tener mayor acumulación de horas luz. Misma situación podría significar ventaja para el paraje El Cerrito (con orientación sur), pero a diferencia de Garambullos, tiene menor número de individuos reproductivos, lo que estaría probablemente influyendo en un menor número de estructuras reproductivas. En este mismo sentido, se ha documentado para otras especies diferentes a cactáceas, que la exposición de las flores podría tener efectos positivos en la polinización, ya que al elevarse la temperatura interna de estas, pueden atraer a más visitantes florales (Dafni, 1994).
La diferencia en la intensidad de la floración y fructificación de cada paraje en el año 2014 en comparación con el año 2015, podría estar determinado por el régimen de lluvia, ya que la precipitación fue mayor en el año 2015 con respecto al 2014 (Conagua-SMN, 2016). Bustamante y Búrquez (2005) comentaron que la fenología reproductiva de S. queretaroensis parece estar ligada en parte, al régimen de lluvia. Barbault (1982) mencionó que las especies vegetales que habitan en espacios desérticos suelen ajustar su etapa reproductiva al período húmedo. Lomelí (1991) refirió que entre los factores externos que afectan a la reproducción de S. queretaroensis, están los climáticos (temperatura y precipitación) y los edáficos.
Respecto a la diferencia en el número promedio de semillas por fruto entre parajes dentro de cada año de observación, podría ser resultado de diferencias en la orientación del relieve, sin embargo, se carece de información para aseverar lo antes expuesto.
Los resultados en torno a la viabilidad de las semillas sugieren que el paraje de estudio influyo de manera significativa, siendo mayor la viabilidad de semillas provenientes de los parajes con mayor exposición solar. Esta diferencia en viabilidad de semillas de acuerdo al paraje también podría obedecer a factores exógenos, como la exposición del relieve que influye en la expresión de las fenofases, sin embargo, se carece de información suficiente para aseverar lo anterior, por lo que se sugiere realizar investigación en torno a esta temática.
Se ha demostrado que el hipoclorito de sodio tiene propiedades oxidativas (Barnett, 1976). La inmersión de semillas de Agave parrasana en este compuesto al 3% fue benéfico para la germinación (Santacruz et al., 1999); lo mismo fue demostrado para Agave victoriae-reginae (Martínez et al., 2003). Mateos (2004), refiere que el hipoclorito de sodio en combinación con agua, desprende oxigeno naciente (O2) y éste a su vez oxida la materia orgánica, lo que podría incrementar la permeabilidad de la testa y la entrada de agua y oxígeno al embrión, elementos esenciales para que una semilla germine. En este sentido, la diferencia significativa en torno a la germinación de las semillas de S. queretaroensis tratadas con hipoclorito de sodio en comparación con las semillas que no recibieron tratamiento pregerminativo, pudo deberse a la oxidación de la testa; sin embargo, se recomienda realizar futuras investigaciones en torno al tema.
En cuanto a los porcentajes finales de germinación, Pimienta (1999b) reportó valores superiores al 90% para semillas de S. queretaroensis bajo condiciones de temperatura promedio de 22º C (± 1.5), flujo fotosintético de fotones 59.4 (± 15.1 μmol m-2 s-1) y una humedad relativa de 59.3 % (± 12.4). Pimienta y Nobel (1995) y Muñoz-Urias et al. (2000), refirieron que las semillas de S. queretaroensis son fotoblásticas. Mismo argumento ha sido referido para otras especies similares al pitayo (Martínez-Holguín, 1983; Rojas-Aréchiga et al., 2001). Contrastando nuestros resultados (semillas con tratamiento: Garambullos = 70%, Soyate = 74.33%; semillas sin tratamiento: Garambullos = 58%, Soyate = 49.67%), la diferencia en comparación con los resultados expuestos por Pimienta (1999b) pueden deberse a que el experimento de germinación se realizó en ausencia de fotoperiodo.
Referente a la emergencia del pitayo en campo, el paraje donde se establecieron las unidades experimentales influyó de manera significativa, en el paraje Garambullos se tuvo el mayor porcentaje de emergencia de la especie, lo que coincide en que dicho sitio tuvo la mayor acumulación de estructuras reproductivas, el mayor número promedio de semillas por fruto y la mayor proporción de semillas viables. Sin embargo, la supervivencia de plántulas se incrementó de manera significativa en el paraje El Cerrito, ubicado en exposición sur. En torno a este resultado, Méndez et al. (2006) no encontraron diferencias significativas en torno a la supervivencia de Pterocereus gaumeri (Britton & Rose) T. MacDoug. & Miranda, de acuerdo con su orientación con respecto a la planta nodriza (N, S, E, O). Por el contrario, Valiente-Banuet y Escurra (1991) refirieron un patrón significativo en torno a la supervivencia de Neobuxbaumia tetetzo (Weber ex K. Schum.) Backeb., del lado norte de la planta nodriza. Así mismo, Mata-González et al. (2002) indicaron la relación de la exposición sur con la mayor densidad y riqueza de especies de suculentas. Es posible que la mayor radiación que se tiene en la exposición sur del paraje El Cerrito, favoreciera al establecimiento de S. queretaroensis.
Considerando los tipos de microhábitat, se observó que bajo el dosel de C. pallida y de S. queretaroensis se incrementó significativamente la emergencia de las plántulas. Pareciera que S. queretaroensis es una especie que requiere condiciones microambientales específicas durante la etapa de plántula para emerger, condiciones que muy probablemente se asocian con el sombreado y protección que le brindaron C. pallida y el dosel de la propia especie, en comparación con los espacios desprovistos de vegetación. Resultados similares encontraron Yeaton y Romero-Manzanares (1986) quienes refirieron que Opuntia streptacantha Lem., en el altiplano zacatecano, así como (Valiente-Banuet y Ezcurra, 1991) Neobuxbaumia tetetzo, en el valle de Tehuacán, presentaban un mayor porcentaje de establecimiento de plántulas bajo el dosel de plantas nodriza, en comparación con los espacios sin dosel. Así mismo, Castillo y Valiente-Banuet (2010) reportaron la importancia de las plantas nodriza en la dinámica poblacional de Neobuxbaumia mezcalaensis (Bravo) Backeb. en la localidad de San Juan Raya, Puebla y García y Mandujano (2010) también lo refirieron para Lophophora williamsii (Lem. ex Salm-Dyck) J.M. Coult. Ramos (2007) encontró resultados similares para Mammillaria dixanthocentron Backeb. ex Mottram. Otras investigaciones subrayan de manera general que la emergencia y establecimiento de plántulas de diversas especies de cactáceas es beneficiada por la sombra de las especies con las cuales se asocian (López y Valdivia, 2007; Méndez et al., 2006; Valiente-Banuet y Ezcurra, 1991), disminuyendo así la evapotranspiración y amortiguando la temperatura (Godínez-Álvarez et al., 2003).
Agradecimientos
Los autores agradecen a las personas y autoridades de la comunidad de San José de Cosalima, que hicieron posible el acceso y acompañamiento para el desarrollo del trabajo en campo.
Referencias
Allison, P. A. (1995). Survival analysis using the SAS® System: a practical guide. Cary, North Carolina: SAS Institute Inc.
Arreola-Nava, H. J. y Terrazas, T. (2003). Especies de Stenocereus con aréolas morenas: clave y descripciones. Acta Botanica Mexicana, 64, 1–18.
Arriaga, C. L., Espinoza, R. J. M., Aguilar, Z. C., Martínez, R. E., Gómez, M. L. y Loa, L. E. (2000). Cerro Viejo-sierra de Chapala. Regiones terrestres prioritarias de México. Escala de trabajo 1:1 000 000. México, D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Arriaga-Ruiz, M. C., Neri-Luna, C., Pimienta, E. y Sánchez-Martínez, J. (2015). El fruto del pitayo silvestre (Stenocereus queretaroensis (Weber) Buxbaum), una alternativa alimenticia, nutricional y socioeconómica en época de estiaje. Ciencias Naturales y Agropecuarias, 2, 362–367.
Balleza, J. J. y Villaseñor, J. L. (2002). La familia Asteraceae en el estado de Zacatecas (México). Acta Botanica Mexicana, 59, 5–69.
Barbault, R. (1982). La noción de estrategia de adaptación en ecología. Ciencia y Desarrollo, 45, 114–122.
Barnett, J. P. (1976). Sterilizing southern pine seeds with hydrogen peroxide. Tree Planters Notes, 27, 17–19.
Barros, J. R. y Killian, S. E. (2013). Efecto de diferentes concentraciones de hipoclorito de sodio en la germinación de la semilla de Podocarpus parlatorei Pilg. Biología en Agronomía, 3, 55–58.
Bowers, J. (1998). Reproductive potential and minimum reproductive size of Ferocactus wislizeni (Cactaceae). Desert Plants, 14, 3–7.
Bustamante, E. y Búrquez, A. (2005). Fenología y biología reproductiva de las cactáceas columnares. Cactáceas y Suculentas Mexicanas, 50, 68–88.
Cabrera, L. J. A. y Gómez, S. M. (2005). Análisis florístico de La Cañada, Querétaro, México. Boletín de la Sociedad Botánica de México, 77, 35–50.
Casas, A. (2002). Uso y manejo de cactáceas columnares mesoamericanas. Biodiversitas, 40, 18–23.
Castillo, J. P. y Valiente-Banuet, A. (2010). Species-specificity of nurse plants for the establishment, survivorship, and growth of a columnar cactus. American Journal of Botany, 97, 1289–1295.
Conagua-SMN (Comisión Nacional del Agua-Servicio Meteorológico Nacional). (2016). Normales Climatológicas. Estación: 00032013- El Chique.
Dafni, A. (1994). Note on side advertisement in flowers. Functional Ecology, 8, 136–138.
Ferrer, M., Durán, R., Méndez, M., Dorantes, A. y Dzib, G. (2011). Dinámica poblacional de genets y ramets de Mammillaria gaumeri cactácea endémica de Yucatán. Boletín de la Sociedad Botánica de México, 89, 83–105.
Figueroa-Castro, D. M. y Valverde, P. L. (2011). Flower orientation in Pachycereus weberi (Cactaceae): effects on ovule production, seed production and seed weight. Journal of Arid Environments, 75, 1214–1217.
Gaol, M. L. y Fox, J. E. D. (2002). Reproductive potential of Acacia species in the central wheatbelt: variation between years. Conservation Science Western Australia Journal, 4, 147–157.
García, E. (2004). Modificaciones al Sistema de Clasificación Climática de Köppen (para adaptarlo a las condiciones de la República Mexicana), 5ª edición. México D.F.: Instituto de Geografía, Universidad Nacional Autónoma de México.
García, N. O. y Mandujano, M. C. (2010). Patrón de distribución espacial y nodricismo del peyote (Lophophora williamsii) en Cuatrociénegas, México. Cactáceas y Suculentas Mexicanas, 55, 36–55.
Godínez-Álvarez, H., Valverde, T. y Ortega-Baes, P. (2003). Demographic trends in Cactaceae. The Botanical Review, 69, 173–203.
González, M. F. (2012). Las zonas áridas y semiáridas de México y su vegetación. México D.F.: Instituto Nacional de Ecología, Secretaría de Medio Ambiente y Recursos Naturales.
Granados, S. D., Mercado, B. A. y López, G. F. (1999). Las pitayas de México. Ciencia y Desarrollo, 145, 58–67.
Hernández, R. D. (2002). Frugivoría en Stenocereus queretaroensis (Ceber) Buxbaum por aves y mamíferos y su contribución en la dispersión de semillas (Tesis). CUCBA, Universidad de Guadalajara. Zapopan, Jalisco, México.
Ibarra-Cerdeña, C. N., Íñiguez-Dávalos, L. I. y Sánchez-Cordero, V. (2005). Pollination ecology of Stenocereus queretaroensis (Cactaceae), a chiropterophilous columnar cactus in a tropical dry forest of Mexico. American Journal of Botany, 92, 503–509.
Inegi (Instituto Nacional de Estadística, Geografía e Informática). (2004). Guía para la interpretación de cartografía. Aguascalientes: Inegi.
Inegi (Instituto Nacional de Estadística, Geografía e Informática). (2010). Compendio de información geográfica municipal 2010-Tabasco-Zacatecas. Aguascalientes: Inegi.
Jiménez, C. L. (2011). Las cactáceas mexicanas y los riesgos que enfrentan. Revista Digital Universitaria, 12, 1–23.
León-de la Luz, J. L. y Domínguez, R. C. (1991). Evaluación de la reproducción por semilla de la pitaya agria (Stenocereus gummosus) en Baja California Sur, México. Acta Botanica Mexicana, 14, 75–87.
Lomelí, M. E. (1991). Demografía reproductiva y fenología floral en pitayo (Stenocereus queretaroensis, (Weber) Buxbaum) (Tesis). Facultad de Ciencias Biológicas, Universidad de Guadalajara, Jalisco, México.
López, R. y Valdivia, S. (2007). The importance of shrub cover for four cactus species differing in growth form in an Andean semi-desert. Journal of Vegetation Science, 18, 263–270.
MacDonald, J. H. (2014). Handbook of biolological statistics. Baltimore, Maryland: Sparky House Publishing.
Marshall, C. T., Kjesbu. O. S., Yaragina, N. A., Solemdal, P. y Ulltang, O. (1998). Is spawner biomass a sensitive measure of the reproductive and recruitment potential of northeast Arctic code?. Canadian Journal of Fisheries and Aquatic Sciences, 55, 1766–1783.
Martínez, P. A., Ortega, L. P., Chávez, V. M. y Bye, R. (2003). Somatic embryogenesis and organogenesis of Agave victoriae-reginae: considerations for its conservation. Plant Cell, Tissue and Organ Culture, 74, 135–142.
Martínez-Holguín, E. (1983). Germinación de semillas de Stenocereus griseus (Haw.) Buxbaum, pitayo de mayo. Cactáceas y Suculentas Mexicanas, 28, 51–57.
Martínez-Ramos, M., Arroyo-Cosultchil, G., Mandujano, C. M. y Golubov, J. (2016). Dinámica poblacional de Mammillaria humboldtii una cactácea endémica de Hidalgo, México. Botanical Sciences, 94, 199–208.
Mata-González, R., Pieper, R. D. y Cárdenas, M. M. (2002). Vegetation patterns as affected by aspect and elevation in small desert mountains. The Southwestern Naturalist, 47, 440–448.
Mateos F. P. (2004). Control de las poblaciones microbianas: esterilización y desinfección. Departamento de Microbiología y Genética. Universidad de Salamanca. Salamanca, España. http://webcd.usal.es/Web/educativo/micro2/tema08.html
Méndez, M., Dorantes, A., Dzib, G., Argáez, J. y Durán, R. (2006). Germinación y establecimiento de plántulas de Pterocereus gaumeri, una cactácea columnar, rara y endémica de Yucatán, México. Boletín de la Sociedad Botánica de México, 79, 33–41.
Muñoz-Urías, A., Huerta, F. M., González, D. y Ramos, J. J. (2000). Influencia de la luz en la germinación de semillas de pitayo (Stenocereus queretaroensis (Weber) Buxbaum). Memorias del Simposio Internacional sobre el Cultivo y Aprovechamiento de la Pitaya (Stenocereus) y la Pitahaya (Hylocereus y Selenicereus). 10-23 de mayo, 2000, Guadalajara, Jalisco, México.
Nassar, J. M. y Emaldi, U. (2008). Fenología reproductiva y capacidad de regeneración de dos cardones, Stenocereus griseus (Haw.) Buxb. y Cereus repandus (L.) Mill. (Cactaceae). Acta Botanica Venezuelica, 31, 495–528.
Pimienta, E. (1999a). El pitayo en Jalisco y especies afines en México. Guadalajara: Universidad de Guadalajara-Fundación Produce Jalisco, A.C.
Pimienta, E. (1999b). Productividad y ecofisiología del pitayo de Querétaro (Stenocereus queretaroensis (Weber) Buxbaum) (Tesis). CUCBA, Universidad de Guadalajara. Zapopan, Jalisco, México.
Pimienta, E. y Nobel, P. S. (1995). Reproductive characteristics of pitayo (Stenocereus queretaroensis) and their relationships with soluble sugars and irrigation. Journal of the American Society of Horticultural Science, 120, 1082–1086.
R-Development Core Team. (2015). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Ramos, L. (2007). Estudio poblacional de Mammillaria dixanthocentron Becket. ex Mitran en el Valle de Cuicatlán, Oaxaca (Tesis de maestría). Instituto Politécnico Nacional. Oaxaca, México.
Rivas, M. y Barilani, A. (2004). Diversidad, potencial productivo y reproductivo de los palmares de Butia capitata (Mart.) Becc. de Uruguay. Agrociencia, 8, 11–20.
Rojas-Aréchiga, M., Casas, A. y Vázquez-Yanes, C. (2001). Seed germination of wild and cultivated Stenocereus stellatus (Cactaceae) from the Tehuacán-Cuicatlán Valley, Central Mexico. Journal of Arid Environments, 49, 279–287.
Rojas-Aréchiga, M. y Vázquez-Yanes, C. (2000). Cactus seed germination: a review. Journal of Arid Environments, 44, 85–104.
Rzedowski, J. (1978). Vegetación de México. México D.F.: Limusa.
Salcedo, P. E. (1991). Aspectos taxonómicos y etnobotánicos del pitayo Stenocereus queretaroensis (Web) Buxb. en el municipio de Techaluta, Jalisco (Tesis). Facultad de Agronomía, Universidad de Guadalajara. Guadalajara, Jalisco, México.
Santacruz, R. F., Gutiérrez, P. H. y Rodríguez, G. B. (1999). Efficient in vitro propagation of Agave parrasana Berger. Plant Cell, Tissue and Organ Culture (Netherlands), 56, 163–167.
SAS (Statistical Analysis Software) (2002). SAS Institute Inc. OnlineDoc® 9.1. 3. Cary, North Carolina, http://support.sas.com/documentation/onlinedoc/91pdf/index_913.html; última consulta: 16.IV.2016.
SAS (Statistical Analysis Software). (2015). SAS Institute Inc. The LIFEREG Procedure. In SAS/STAT® 14.1 User’s Guide. Cary, NC. p. 4998-5116.
Socolowski, F., Mascia, D. C. V., Simao, E. y Takaki, M. (2010). Influence of light and temperature on seed germination of Cereus pernambucensis Lemaire (Cactaceae). Biota Neotropical, 10, 53–56.
Tremblay, R. (2006). The effect of population structure, plant size, herbivory and reproductive potential on effective population size in the temperate epiphytic orchid, Sarcochilus australis. Cunninghamia, 9, 529–535.
Trippel, E. A. (1999). Estimation of stock reproductive potential: history and challenges for Canadian Atlantic gadoid stock assessments. Journal of Northwest Atlantic Fishery Science, 25, 61–81.
Valiente-Banuet, A. y Ezcurra, E. (1991). Shade as a cause of the association between the cactus Neobuxbaumia tetetzo and the nurse plant Mimosa luisana in the Tehuacán Valley, Mexico. Journal of Ecology, 79, 961–971.
Yeaton, R. I. y Romero-Manzanares, A. (1986). Organization of vegetation mosaics in the Acacia schaffneri–Opuntia streptacantha association southern Chihuahuan Desert, Mexico. Journal of Ecology, 65, 586–595.
Zar, J. H. (1988). Statistical tables and graphs interpolation. Upper Saddle River, New Jersey: Prentice-Hall.


