Influence of natural tail streamer asymmetry and tail length on annual apparent survival of adult Magnificent Frigatebird (Fregata magnificens)
Mónica González-Jaramillo a, José Luis Rangel-Salazar b, Horacio de la Cueva c, *
a Calle 5, int. 14, Samulá, 24090, San Francisco de Campeche, Campeche, Mexico
b Departamento de Ecología y Sistemática Terrestres, El Colegio de la Frontera Sur, Apartado postal 63, 29290 San Cristóbal de las Casas, Chiapas, Mexico
c Departamento de Biología de la Conservación, Centro de Investigación Científica y Educación Superior de Ensenada, Carretera Ensenada-Tijuana Núm. 3918, Zona Playitas, Apartado postal 360, 22860 Ensenada, Baja California, Mexico
*Corresponding author: cuevas@cicese.mx (H. de la Cueva)
Abstract
Flight traits mediated by symmetry should be constrained by mechanical or aerodynamic effects. Natural asymmetry and external tail feather length could have unfavorable effects on aerodynamic performance and affect the survival of highly aerial birds. The Magnificent Frigatebird, Fregata magnificens, is a long-lived, highly aerial seabird with a deeply forked tail, and an extraordinary division of labor between sexes. Its flight pattern is soaring and dynamic, it also has fast active flight and acrobatic maneuvers. We explore the possible effect of tail streamer asymmetry and tail length on annual apparent survival of Magnificent Frigatebirds by using 6 years (2000-2005) of capture-mark-resighting of live-encounter data analysis. Male frigatebirds showed significant differences between left and right tail streamer length, and tail asymmetry differences between sexes. Models did not find a clear effect of tail streamer asymmetry and tail length, as individual covariates, on annual apparent survival by sex. Natural variation in a secondary sexual trait like those explored here likely does not affect apparent survival. Life-history strategies such as flight pattern, breeding behavior, and tail moult may constrain any negative effects. Longer mark-resighting studies and additional modelling could answer conclusively the association between survival and flight traits in Magnificent Frigatebirds.
Keywords:
Aerodynamic cost; External rectrices; Seabird; Survival modelling
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Influencia de la asimetría natural en las rectrices y la longitud de la cola en la supervivencia aparente anual de la Fragata Magnífica
(Fregata magnificens) adultas
Resumen
Los caracteres de vuelo mediados por asimetría deben restringirse por efectos aerodinámicos. La asimetría natural y longitud de la cola pueden tener efectos desfavorables en el desempeño aerodinámico afectando negativamente la supervivencia de aves extremadamente aéreas. La Fragata Magnífica (Fregata magnificens) es un ave marina longeva, extremadamente aérea, con cola profundamente bifurcada y una extraordinaria división de labores entre sexos. Sus tipos de vuelo son: remonte o remonte dinámico, y vuelo rápido activo con maniobras acrobáticas. Exploramos el posible efecto de la asimetría natural y la longitud de la cola en la supervivencia aparente anual de la Fragata Magnífica, analizando 6 años (2000-2006) de datos de captura-marcaje-reavistamiento. Las fragatas mostraron diferencias significativas entre la longitud de las rectrices exteriores derecha e izquierda y diferencias de la asimetría de la cola entre sexos. Los modelos no detectaron un efecto claro de estas covariables individuales en la supervivencia aparente anual por sexo. Las estrategias de historia de vida como el tipo de vuelo, comportamiento reproductivo y muda de la cola, podrían restringir un efecto negativo en la supervivencia aparente. Estudios más largos de captura-reavistamiento y modelos adicionales podrían responder conclusivamente la asociación entre supervivencia y caracteres de vuelo en las Fragatas Magníficas.
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Palabras clave:
Costo aerodinámico; Rectrices exteriores; Ave marina; Modelación de supervivencia
Introduction
In consecutive monogamous species, those that may change partners from year to year, such as the Magnificent Frigatebird (Fregata magnificens), secondary sexual characters comprise an important mate selection feature. In male frigatebirds, seasonal throat sacs and a deeply forked tail are such characters. Tail length, including feather asymmetry in a deeply forked tail, could be unfavorable for survival over long time periods due to flight aerodynamic costs, which could affect body condition (Cuervo et al., 2002). Birds with tail streamer asymmetry have to overcome 2 important disadvantages: 1) the aerodynamic cost of being asymmetrical (Norberg, 1990; Thomas, 1993a), which reduces maneuverability and increases flight costs (Balmford et al., 1993; Evans, 1998; Møller, 1991; Thomas, 1993a, b), and 2) the unfavorable effect on mate selection, since tail streamer asymmetry can be used by potential partners as a quality indicator (Møller, 1990, 1992; Møller & Höglund, 1991). Costs of tail streamer asymmetry on aerodynamics (Evans, 1998, 1999; Norberg, 1994), effects on mate selection (Møller, 1991, 1992), and parental care performance (the cooperative parent behavior that increases the offspring fitness through the investment to rearing the chick (Osorno & Székely, 2004), such as differences in reproductive success between symmetric and asymmetric birds, have been assessed usually for one breeding season (González-Jaramillo & de la Cueva 2010, 2012).
Natural bird tail streamer asymmetry can occur due to asynchronous moult, wear, or breakage (González-Jaramillo & de la Cueva, 2010). Fluctuating asymmetry (FA, asymmetric development of a body trait; Møller & Höglund, 1991) theory, based on the simultaneous growth of feathers (Møller & Höuglund, 1991), is not applicable in all species if their tail moult is consecutive (e.g., Magni-ficent Frigatebird, Fregata magnificens; Madsen, 2005).
Manipulation of asymmetry and tail length in birds with deeply forked tails, e.g., Barn Swallows (Hirundo rustica; Møller, 1990, 1991), and with a long tail streamer, e.g., Red-billed streamertails (Trochilus polytmus; Evans et al., 1994), showed that birds with long and symmetric tails have a better flight performance than those with short and asymmetric tails. Thus, the aerodynamic cost of tail streamer asymmetry is significant only when tail length is considered (Evans et al., 1994). In birds with long tails, e.g., widowbirds, males with short tails increase their average rate of flight display, with energetic costs and female choice consequences (Pryke & Andersson, 2005), whereas males with long tails decrease it (Andersson, 1982).
Other birds that also use visual clues to regulate social behaviors, e.g., the European starlings (Sturnus vulgaris), cannot detect asymmetries < 1.25%, which are usually developed in secondary sexual characters mainly by fluctuating asymmetry (Swaddle, 1999). Thus, asymmetry cannot be used as an indicator of quality if it is not detected.
The Magnificent Frigatebird, Fregata magnificens (hereafter frigatebird), is a neotropical suliform seabird with reversed sexual size dimorphism (Osorno, 1996). Male frigatebirds have significantly longer external rectrices than females (González-Jaramillo & de la Cueva, 2012). Frigatebirds show female mate choice (Madsen, 2005), and a remarkable division of labor between sexes during chick rearing. Males desert the nest after 3 months of biparental care, and then the female continues rearing the chick after fledging up to 11 months (Osorno & Székely, 2004). In this species, natural tail streamer asymmetry has no evident effect on mate selection, or on biparental care performance within a breeding season (González-Jaramillo & de la Cueva, 2010).
Frigatebirds are highly aerial, characterized by a long, deeply forked tail (Diamond & Schreiber, 2002), capable of remaining airborne for long periods, and soaring over long distances (Pennycuick, 1983; Weimerskirch et al., 2003, 2006). They also show a highly acrobatic and fast flight, flapping their wings up to 3.21 Hz during active flight (Shnell, 1974). Theoretical flight models predict that the frigatebirds’ natural tail streamer asymmetry may affect flight performance, adding aerodynamic and compensation costs during soaring, thus predicting different aerodynamic performance by sex (González-Jaramillo & de la Cueva, 2012). Increased flight costs during the breeding season could have an effect on survival over a longer time period since tail streamer asymmetry has to be compensated and overcome by asymmetric birds in order to compete with symmetric individuals.
We could not find studies of the possible effect of asymmetry and length of tail streamers on birds’ survival across multiple breeding seasons. Using mark-resighting data, we assess if tail streamer asymmetry and tail length influenced apparent survival in either sex. Sexual dimorphism, breeding behavior, and differences in aerodynamic performance could influence survival in these frigatebirds. If tail streamer asymmetry and tail length have flight performance costs (Evans, 1999; Thomas, 1993a, b), these costs could in turn affect survival probabilities of frigatebirds during the following breeding seasons. Thus, models of annual apparent survival that include tail streamer asymmetry and tail length as individual covariables will show a better support than models of apparent survival without them.
Materials and methods
This study was conducted on Isabel Island, off the Central Mexican Pacific Coast (21º51’ N, 105º55’ W). Isabel Island is a Mexican National Park and is considered an “Área de Importancia para la Conservación de Aves en México” (AICA), the Mexican equivalent of North American Important Bird Areas (IBAs). The island has a breeding colony of 15,000-16,000 Magnificent Frigatebird individuals (Madsen et al., 2007), and several thousand pairs of another 8 seabird species (Drummond et al., 2000).
Eighty one adult nesting frigatebirds (30 female and 51 male) without broken external tail feathers were marked and released during the breeding season of 2000, then resighted the following 5 breeding seasons (2001-2005; Table 1). Nesting frigatebirds were caught randomly by hand at night. A random number series was selected from a random number table. We stood at the edge of the colony and started counting bushes to find the first random number. We followed the random sequence from there counting left and right as we moved through the colony. After catching frigatebirds, we temporarily removed them from the colony, and marked them using yellow canvas wing tags with black numbering placed on the left wing, 20.5 × 8 cm and 10.83 g (approximately 1% of the weight of the lightest individuals), and ringed with numbering metal rings placed on the left tarsus as described by Osorno (1996, 1999).
Given the fragility of these birds, extra care was taken whilst measuring. To minimize measurement error, all measures were taken at least twice or until constant measures with less than 1mm difference were obtained by the same person during the length of the study and registered only once concordant measurements had been obtained (González-Jaramillo & de la Cueva, 2007, 2010, 2012; González-Jaramillo et al., 2010; Madsen, 2005; Madsen et al., 2007). It is known that errors of < 1% between measurements do not alter significantly aerodynamic calculation costs (Blake et al., 1990; González-Jaramillo & de la Cueva, 2012), or natural asymmetry estimations (González-Jaramillo & de la Cueva, 2007, 2010; González-Jaramillo et al., 2010). Repeated measures were not taken between years, since frigatebirds are difficult to recapture in consecutive occasions. This difficulty in recapture motivated the capture- marking- resighting method used here.
Table 1
Adult male and female nesting frigatebrids captured and marked during the 2000 breeding season and resighted during the 2001-2005 breeding seasons. Isabel Island, Nayarit, Mexico.
|
Breeding season |
||||||
|
Sex |
Captured and marked (n) |
Resighted (n) |
||||
|
Breeding season |
2000 |
2001 |
2002 |
2003 |
2004 |
2005 |
|
Females |
30 |
14 |
10 |
9 |
8 |
14 |
|
Males |
51 |
38 |
28 |
22 |
16 |
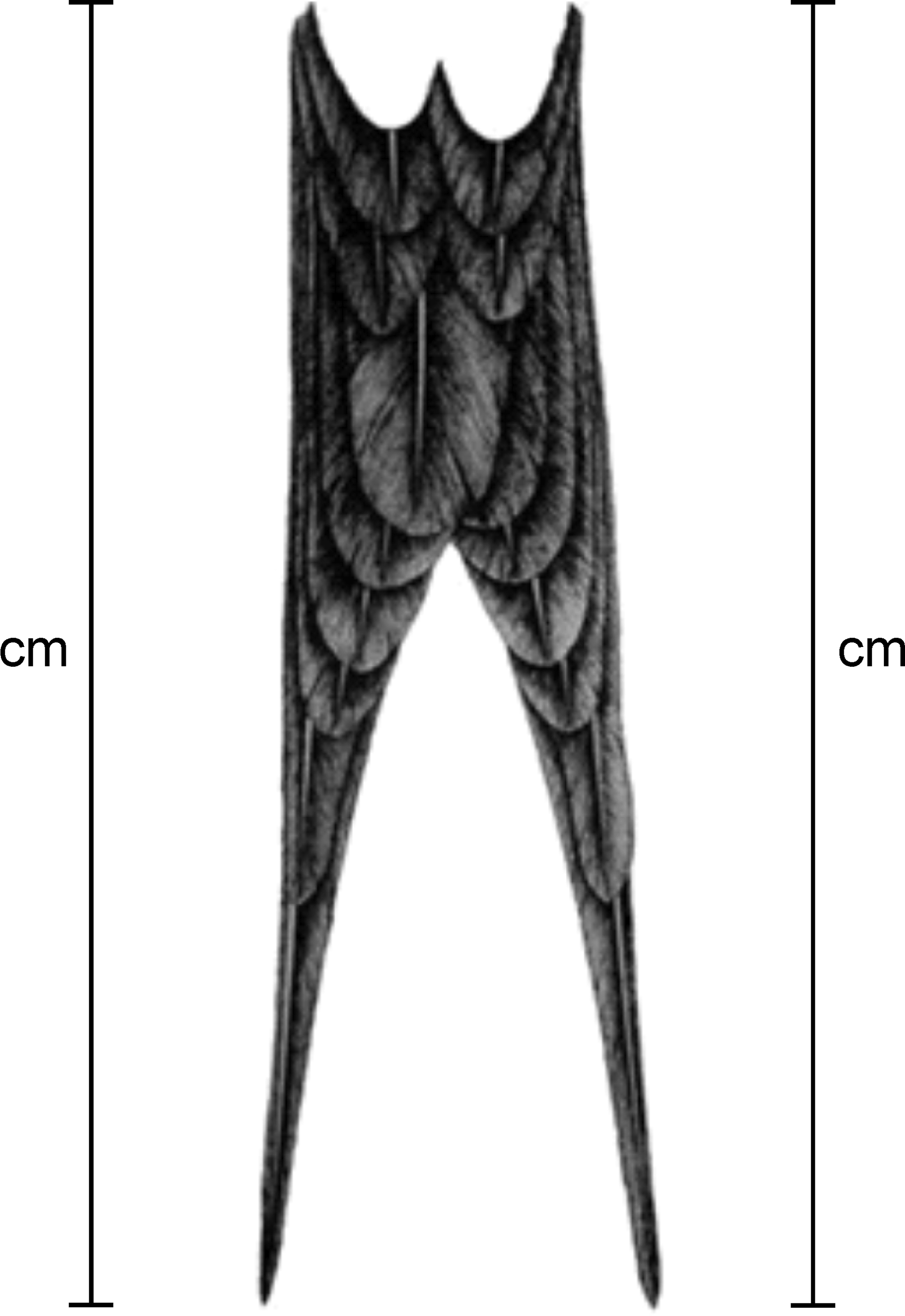
We measured (± 1mm) frigatebirds as follows: 1) tail streamer length, length of right and left tail streamer rectrices, from the quill to the tip of the feather cuvis (Fig. 1); 2) tail length, as the mean of left and right tail streamer feathers measures. Tail streamer asymmetry: 3) absolute tail asymmetry, the numerical difference between right and left tail streamer feathers, and 4) relative degree of asymmetry, i.e., measure 3) divided by measure 2) (Møller & Höglund, 1991; Palmer & Strobeck, 1986). Lastly, to explore differences of tail streamer traits (tail streamer length, tail length, and tail streamer asymmetry) between sexes, t-tests of 2 means were performed for each measurement: right and left tail streamer, tail length, and absolute and relative tail streamer asymmetry. We also measured differences in tail streamer length within sex with a paired t-test. To assess the possible association between tail streamer relative asymmetry and tail length of all frigatebirds sampled (n = 81) a Spearman rank correlation (r) analysis was performed.
Frigatebird encounter histories were coded as either sighted during each field season (1) or not detected (0). Mark-resighting models (Lebreton et al., 1992), using Program MARK 7.2 (White & Burnham, 1999), were used to estimate apparent survival (ϕ ; probability that an individual survives between 2 sample periods and returns to the study area if it is alive and does not emigrate permanently), and encounter rates (p; probability that an individual is available for encounter and detected in the study area if it is alive).
The coded encounter histories of each male and female frigatebird were associated with absolute tail streamer asymmetry (ts), and tail length (tl) at first capture, taken as continuous values, and used as individual covariates (ind cov). Given that asymmetry is dynamic and changes consecutively after a given moult, i.e., an asymmetric male will always be asymmetric, but the asymmetry won’t be the same from year to year, we cannot use a time since marking (TSM) global model in which a possible transition from asymmetric to symmetric tail can be considered.
Because fluctuating asymmetry theory is not applicable in this species in which male tail moult (n = 197) is continuous with a general turn-over rate of change of 0.31 of the tail feathers per year in all frigatebird ages (Madsen, 2005), we assumed that the individual covariate of asymmetry is fixed each breeding season over the lifetime of individuals. Therefore, we constrained estimates of annual apparent survival (ϕ), and encounter rates (p) to be a linear function of those values.
Because the aerodynamic effect of tail streamer asymmetry in maneuvering is associated with tail length (Evans et al., 1994), under the hypothesis that there is likely a normalizing selection on tail length (tl), such that in relation to some ideal tail length, having either a small or long tail could be a handicap, we tested a quadratic model (tl2) (Cooch & White, 2014).
Since no method exists to test for goodness of fit of models with individual covariates, we selected a global model {ϕ (s × t), p (s × t)} in which frigatebird apparent survival (ϕ), and encounter rates (p) depend on sex (s), and time (t). We then performed a logistic regression analysis to quantify the median variance inflation factor. The median ĉ-value was used in all the other models, including those with individual covariates, as recommended by Cooch and White (2014). If 1 ≤ ĉ ≤ 3 the global model is acceptable (Burnham & Anderson 2002). Overdispersion was corrected by using the median ĉ-value to improve parameter estimations (Cooch & White, 2014). Thus, in order to assess the effect of covariates, tail symmetry, and tail length, our starting global model {ϕ (s× t), p (s × t)} was compared vs. one model including the individual covariates: tail streamer asymmetry (ts) and tail length (tl): {ϕ (s×t) ts tl, p (s×t)}. A set of 11 candidate models was selected, including a quadratic model ({ϕ (s) ts tl2, p (s)}).
The logit link function was used in all of our design matrices. Individual covariates were not standardized since all models were constructed with a common intercept for male and female frigatebirds and different slope parameters. The design matrix was standardized by performing a logistic regression analysis to quantify the median variance inflation factor (ĉ; Cooch & White, 2014).
Constraints to the global model were first applied to encounter rate (p) to find the best fit (Hagen et al., 2005; Sandercock et al., 2005), and then to apparent survival (ϕ). Model selection was based on the difference in QAICc values between models (ΔQAICc), the best fit model had a ΔQAICc= 0, and the other models were considered equally parsimonious if ΔQAICc ≤ 2 (Burnham &Anderson, 1998). The quasi-Akaike Information Criterion (QAICc = -(Dev/ ĉ) + 2K + 2K (K+1)/(n-K-1), where Dev is the model deviance, K is the number of parameters, and n is sample size, adjusts the lack of fit between general and saturated models by adjusting the likelihood term, including the measure ĉ (Burnham & Anderson 1998). Ratios between Akaike weights of 2 candidate models (wi w-1j) were used to quantify the relative degree that a particular model was supported by the data relative to another model (Sandercock et al., 2005). Lastly, to estimate apparent survival (ϕ) and resighting (p) parameters, the model averaging procedure of Program MARK 7.2 (White & Burnham, 1999) was performed and mean individual covariate values were reported.
Results
Average feather and tail lengths were longer in females than in males; in both sexes left feathers were longer than right feathers (Table 2). Nevertheless, differences between right and left feather lengths were significant in females (t29 = 2.08, p < 0.05) but not in males (t50 = 0.73, p = 0.47). Absolute and relative tail asymmetries were greater in males than in females (Table 2). The Spearman rank correlation analysis showed a significant negative correlation between tail streamer asymmetry and tail length (r81 = -0.558, p < 0.05; Fig. 2).
Table 2
Comparison of tail measures (mean ± SD) of adult nesting male and female Magnificent Frigatebirds captured and marked during the 2000 breeding season. Isabel Island, Nayarit, Mexico.
|
Tail streamer length (cm) |
Tail length |
Tail streamer asymmetry |
|||
|
Sex |
Right |
Left |
(cm) |
Absolute (cm) |
Relative |
|
Males (n = 51) |
42.64 ± 8.12 |
43.48 ± 6.37 |
43.06 ± 6.03 |
3.74 ± 7.36 |
0.12 ± 0.30 |
|
Females (n = 30) |
44.77 ± 6.55 |
47.33 ± 1.99 |
46.00 ± 3.62 |
2.88 ± 6.25 |
0.07 ± 0.17 |
|
t-test p |
1.22 0.22 |
3.12 < 0.05 |
2.42 < 0.05 |
0.53 0.59 |
0.72 0.47 |
Our global model {ϕ (s× t), p (s×t)} showed an acceptable fit to male and female frigatebirds live encounter data (median ĉ = 1.96, model 8, Table 2). There was no effect of individual covariates, tail streamer asymmetry (ts), and tail length (tl) on apparent survival (ϕ) of male and female frigatebirds (s) through time (t) (ΔQAICc ≈ 30, w2 < 0.01, model 11, Table 3). The best supported model was that in which there was neither individual covariable, nor time effects on parameter estimations (ϕ and p), but there was a sex effect (model 1, Table 3), followed by the model including time and sex effect interaction on apparent survival (ϕ), but a constant encounter rate (p) through time (model 2, ΔQAICc = 2.17, w9 w-15 = 2.96, Table 3).
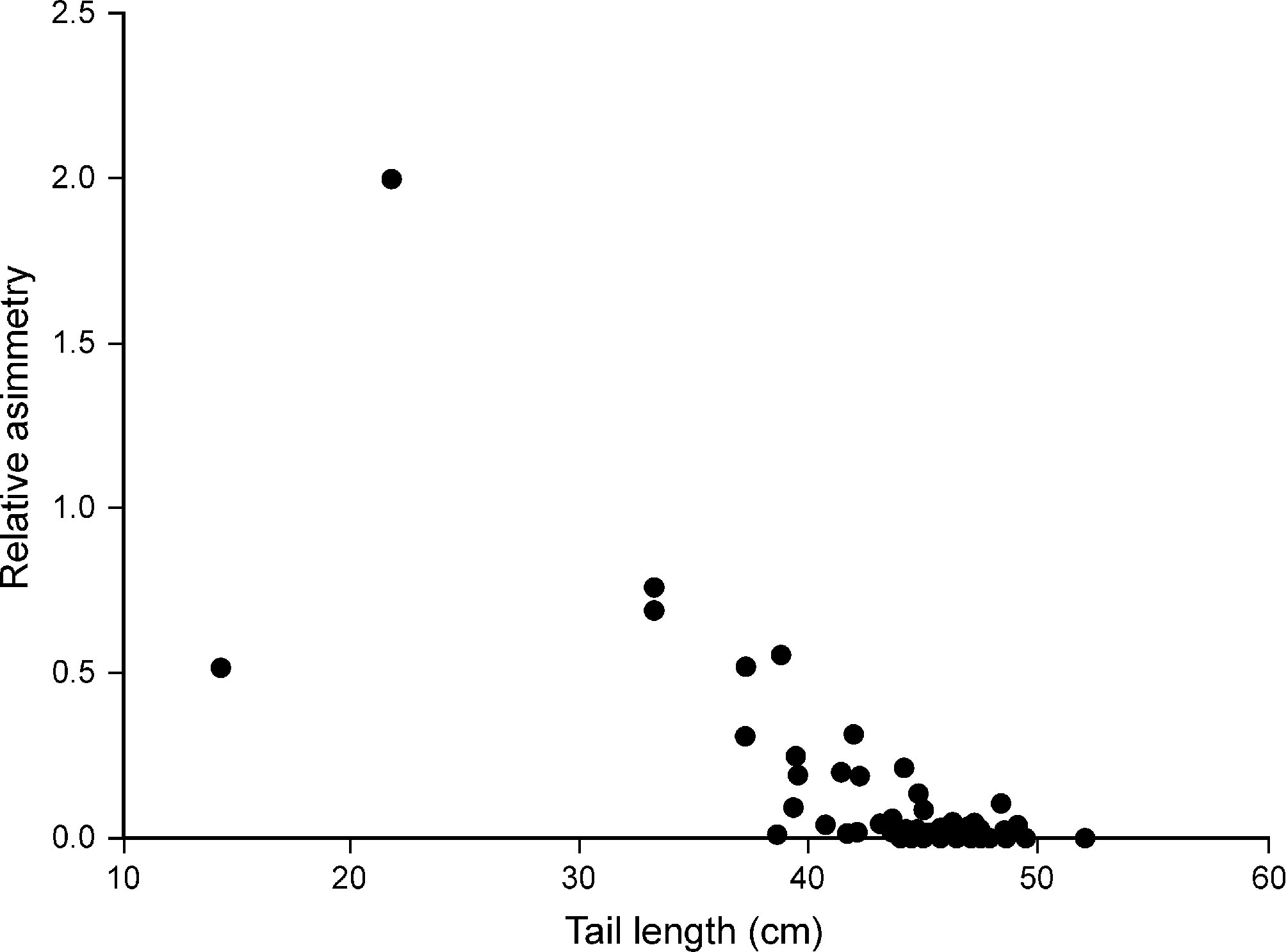
Table 3
Mark-capture-resighting models used to estimate apparent survival rates of adult male and female nesting Magnificent Frigatebirds during the 2000-2005 breeding seasons. Isabel Island, Nayarit, Mexico. Model structure includes: encounter rate (p) and apparent survival (ϕ). Model effects include: sex (s), time-dependence (t), absolute tail streamer asymmetry (ts), tail length (tl), and tail length2 (tl2, to test for normalizing selection effect). Model constraints include main effects (+), and factorial models with interactions (×).
|
Model statistics |
Model description |
|||||
|
Models |
K |
QDev |
QAICc |
ΔQAICc |
wi |
|
|
1. {ϕ (s), p (s)} |
4 |
238.46 |
246.63 |
0.00 |
0.46 |
ϕ and p only sex effect, time constant |
|
2. {ϕ (s×t), p (s)} |
9 |
229.97 |
248.81 |
2.17 |
0.15 |
ϕ sex and time interaction, p only sex effect |
|
3. {ϕ (s) ts tl, p (s)} |
6 |
236.73 |
249.11 |
2.48 |
0.14 |
ϕ sex effect and ind cov (ts, tl), p only sex effect |
|
4. {ϕ (s) ts tl2, p (s)} |
6 |
236.85 |
249.24 |
2.60 |
0.12 |
ϕ sex effect and ind cov (ts, tl2), p only sex effect |
|
5. {ϕ (s+t), p (s)} |
7 |
234.98 |
249.49 |
2.86 |
0.11 |
ϕ sex and time effects, p only sex effect |
|
6. {ϕ (s+t) ts tl, p(s)} |
9 |
233.71 |
252.54 |
5.91 |
0.03 |
ϕsex and time effects and ind cov (ts, tl), p only sex effect |
|
7. {ϕ (s×t), p (s+t)} |
15 |
224.52 |
256.88 |
10.17 |
< 0.01 |
ϕ and p, sex and time effects |
|
8. Global model {ϕ (s×t), p (s×t)} |
18 |
224.50 |
263.80 |
17.17 |
< 0.01 |
ϕ and p, sex and time interaction |
|
9. {(s×t) ts tl, p(s)} |
15 |
232.34 |
264.62 |
17.99 |
< 0.01 |
ϕ sex and time interaction and ind cov (ts, tl) effect, p only sex effect |
|
10.{(s×t) ts tl, p(s+t)} |
21 |
224.41 |
270.94 |
24.31 |
< 0.01 |
ϕ sex and time interaction and ind cov (ts, tl) effects, p sex and time effects |
|
11.{ (s×t) ts tl, p(s×t)} |
24 |
222.33 |
276.30 |
29.67 |
0.00 |
ϕ sex and time interaction and ind cov (ts, tl), p sex and time interaction |
Models including tail streamer asymmetry (ts) and tail length (tl) effects (model 3, Table 3), as well as the one including normalizing selection effect (quadratic model, model 4, Table 3) in parameter estimations (ϕ and p), showed less support than the best supported model without individual covariate effects (ΔQAICc = 2.47, w9 w-110 = 3.45, Model 3; ΔQAICc = 2.60, w9 w-111 = 3.67, model 4; Table 3). The model in which time (without interaction) on apparent survival was considered, but resighting rate depended only on sex effect (model 5, Table 3) and the model in which individual covariate effects were included (model 6, Table 2), showed even less support (ΔQAICc = 2.86 and ΔQAICc = 5.91, respectively; Table 2). The rest of our candidate models (models 7, 8, 9, 10, and 11; Table 3) showed no support (10 < ΔQAICc < 30, Table 3) and were excluded from our model inference.
Model averaging results showed variation of estimated apparent survival (ϕˆ ) among years (2000-2005), as well as a constant estimated resighting rate of male and female frigatebirds (pm = 0.77 ± 0.15, pf = 0.61 ± 0.25, Table 4).
Table 4
Estimated apparent annual survival (ϕˆ ), encounter rates (p), and the mean values (x) of male (ϕˆm, pm) and female (ϕˆf, pf) Magnificent Frigatebirds breeding at Isabel Island, Nayarit, Mexico during the 2000-2005 breeding seasons. Mean individual covariate values ± SE are reported. Parameters calculated by model averaging based on Akaike weights across the set of candidate models.
|
Interval |
ϕˆm |
ϕˆf |
pm |
pf |
|
2000 – 2001 |
0.86 ± 0.06 |
0.83 ± 0.10 |
0.77 ± 0.06 |
0.61 ± 0.09 |
|
2001 – 2002 |
0.84 ± 0.06 |
0.86 ± 0.08 |
0.77 ± 0.06 |
0.61 ± 0.09 |
|
2002 – 2003 |
0.84 ± 0.07 |
0.88 ± 0.08 |
0.77 ± 0.06 |
0.61 ± 0.09 |
|
2003 – 2004 |
0.83 ± 0.08 |
0.88 ± 0.08 |
0.77 ± 0.06 |
0.61 ± 0.09 |
|
2004 – 2005 |
0.89 ± 0.50 |
0.91 ± 0.95 |
0.77 ± 0.05 |
0.61 ± 0.93 |
|
x |
0.85 ± 0.15 |
0.87 ± 0.25 |
0.77± 0.06 |
0.61 ± 0.25 |
During the first time interval (2000-2001) males had a higher estimated apparent survival (ϕˆ ) and apparent resighting rate (p) than females (ϕˆm > ϕˆf, Table 4). The opposite occurred during the rest of the time (2001-2005), in which males had a smaller estimated apparent survival than females (ϕˆm < f, Table 4), with a clear increase in the last time interval (2004-2005, Table 4). In this analysis males showed a smaller estimated apparent survival and standard error than females (ϕˆm = 0.85 ± 0.15 vs ϕˆf = 0.87 ± 0.25).
Discussion
Length of left and right tail streamers differed between the sexes, and external tail feathers were more symmetric in females than in males. Nevertheless, our modeling did not show evidence of a clear effect of tail streamer asymmetry and tail length individual covariables on annual apparent survival of adult nesting frigatebirds during the length of the study.
The difference between left and right tail streamer length in adult frigatebirds, significant in females, could be an adaptation to soaring flight, as has been suggested for other flight structures such as wings (Hedenström, 1993; Norberg, 1990; Sapir et al., 2011). Some marine birds, including frigatebirds, fly over the seas by extracting energy from thermals and other wind movements during soaring flight (Hedenström, 1993) and dynamic soaring (Cone, 1964; Rayleigh, 1883), in which bird wings are widely opened, so the bird can ascend slowly by circling in convective thermals (Sapir et al., 2011) or near the water surface wind gradient (Cone, 1964; Rayleigh, 1883). During slow flight, the tail is widely spread, increasing the lift generated by the wings. As tail spread and tail length increase, the effect of tail asymmetry on flight performance increases too given, that drag is proportional to air speed and the exposed surface area of the feather (Thomas, 1993b). In species like frigatebirds where soaring flight is dominant, wing adaptations are expected because of different conditions of thermals and bird flight performance (Hedenström, 1993). Thus, once a species has developed a flight mode, some adaptations or variations in other flight structures, such as tails, could be expected (i.e., continuous tail moulting, which has been identified in other soaring species, e.g., vultures; Houston, 1975).
We found that both feather and tail lengths are greater in female frigatebirds. Morphological differences between sexes have also been identified in wings (González-Jaramillo & de la Cueva, 2012), and it has been suggested that they are associated with selection pressures due to dissimilar aerodynamic performance during breeding seasons, which are in turn related to dissimilar roles in biparental care performance (Osorno & Székely, 2004; González-Jaramillo & de la Cueva, 2010).
The best-supported model, without tail streamer asymmetry and tail length as individual covariates, was as parsimonious as the model including them. Thus, there is no evidence of an effect of these covariables on apparent survival during our study. Nevertheless, there is a possibility that our candidate sets of models did not include one that could clearly detect the suggested effect of tail asymmetry and apparent survival in frigatebirds, although we followed suggestions for model selection (Burnham & Anderson, 2002), including theoretical support, exploratory data analysis, the a priori formulation of the global model and candidate models considering relevant effects (i.e., sex; González-Jaramillo & de la Cueva, 2007), and the balance between our biological question and data.
The length of the study and model selection may have an influence on our results, considering the time length of our study (2000-2005 breeding seasons) compared to the known frigatebird life span, > 40 years (Madsen et al., 2007), and the possible effect of age on annual apparent survival of frigatebirds (González-Jaramillo et al., 2010), which we could not include in our candidate set models (Table 3).
Furthermore, moulting and breeding involve trade-offs of energetic costs needed to survive. Since asymmetry is more important in outer than in central tail feathers (Thomas, 1993b), a symmetrical moult implies that birds minimize the asymmetry effect of those feathers that are functionally important during flight performance (Arroyo et al., 2004). In vagile seabirds with soaring flight such as frigatebirds, moulting and breeding are more likely to overlap due to their particular life history (Bridge, 2006). Given that the strategy of continuous moult pattern in frigatebirds, avoiding tail gaps during the breeding season, and their ability to compensate for tail streamer asymmetry (González-Jaramillo & de la Cueva, 2012) could minimize the energetic cost on flight performance (Thomas, 1993b), it is possible that tail streamer asymmetry and tail length have no influence on energetic costs and, thus, neither on apparent survival of frigatebirds.
We suspect that flight performance strategies used by frigatebirds, as well as continuous moult, and division of labor between sexes during the breeding season, may constrain tail streamer asymmetry and tail length effects on annual apparent survival rates. Nevertheless, we highly recommend carrying out modelling with a longer time series of live encounter data > 6 years length to elucidate conclusively the question of survival and asymmetry in this highly aerial and long-lived seabird species.
Acknowledgments
We are grateful to B. Sandercock, and G. Fernandez for valuable comments and helpful suggestions on the manuscript. We thank J. L. Osorno, V. Madsen, the students at the Laboratorio de Conducta Animal, Instituto de Ecología, UNAM, and volunteers O. Espinoza and M. Richard. The administrators of Isabel Island National Park (Conanp) provided logistics and facilities. Fishermen of San Blas, Nayarit, helped us in uncountable ways. The authors do not have a conflict of interest to declare.
References
Andersson, M. (1982). Sexual selection, natural selection and quality advertisement. Biological Journal of the Linnean Society, 17, 375–393.
Arroyo, B., Mínguez, E., Palomares, L., & Pinilla, J. (2004). The timing and pattern of moult of flight feathers of European Storm-petrel Hydrobates pelagicus in Atlantic and Mediterranean breeding areas, Ardeola, 51, 365–373.
Balmford, A., Jones, I. L., & Thomas, A. L. R. (1993). On avian asymmetry: selection for symmetrical tails and wings in birds. Proceedings of the Royal Society of London Series B, 252, 245–251.
Blake, R.W., Kolotylo, R., & de la Cueva, H. (1990). Flight speeds of the barns swallow (Hirundo rustica). Canadian Journal of Zoology, 68, 1–5.
Bridge, E. S. (2006). Influences of morphology and behavior on wing-moult strategies in seabirds. Marine Ornithology, 34, 7–19.
Burnham, K. P., & Anderson, D. R. (1998). Model selection and inference: a practical information-theoretic approach. New York: Springer-Verlag.
Burnham, K. P., & Anderson, D. R. (2002). Model selection and multimodel inference, a practical information-theoretic approach, 2nd Ed. Berlin: Springer.
Cone, Jr., C. D. (1964). A mathematiccal analysis of the dynamic soaring flight of the albatross with ecological interpetations. Gloucester Point, VA: Virginia Institute of Marine Sciences Special Scientific Report. No. 50.
Cooch, E., & White, G. (2014). Program MARK, a gentle introduction. 13th Edition. Retrieved on 1/10/ 2014, from: http://www.phidot.org/software/mark/docs/book/
Cuervo, J. J., Møller, A. P., & de Lope, F. (2002). Experimental manipulation of tail length in female Barn Swallows (Hirundo rustica) affects their future reproductive success. Behavioural Ecology, 14, 451–456.
Diamond, A.W., & Schreiber, E. A. (2002). Magnificent Frigatebird (Fregata magnificens). In A. Poole (Ed.), The birds of North America online. Ithaca: Cornell Lab of Ornithology. Retrieved from : http://bna.birds.cornell.edu/bna/species/601
Drummond, H., Macías, C., Valiente-Baunet, A., & Osorno, J. L. (2000). Isla Isabel. In M. C. Arizmendi, & L. Márquez-Valdemar (Eds.), Áreas de importancia para la conservación de las aves en México (p. 41). México D.F.: CIPAMEX/ Conabio/ FMCN/ CCA.
Evans, M. R. (1998). Selection on swallow tail streamers. Nature, 394, 233–234.
Evans, M. R. (1999). Length of tail streamers in Barn Swallows. Nature, 397, 115.
Evans, M. R., Martins, T. L. F., & Haley, M. (1994). The asymmetrical cost of tail elongation in Red-billed streamertails. Proceedings of the Royal Society of London Series B, 256, 97–103.
González-Jaramillo, M., & de la Cueva, H. (2007). Apparent survival of adult Magnificent Frigatebirds in the breeding colony of Isla Isabel, Mexico. Waterbirds, 30, 73–79.
González-Jaramillo, M., & de la Cueva, H. (2010). Natural tail streamer asymmetry in male Magnificent Frigatebirds Fregata magnificens: influence on mate selection and male parental care performance. Marine Ornitholology, 38, 85–90.
González-Jaramillo, M., & de la Cueva, H. (2012). Influencia de la asimetría de las rectrices exteriores en la eficiencia de vuelo del rabihorcado (Fregata magnificens). Huitzil, 13, 6–16.
González-Jaramillo, M., Rangel-Salazar, J. L., & de la Cueva, H. (2010). Annual apparent survival rates of immature Magnificent Frigatebirds in a large breeding colony in Western Mexico. Waterbirds, 33, 518-526.
Hagen, C. A., Pitman, J. C., Sandercock, B. K., Robel, R. J., & Applegate, R. D. (2005). Age-specific variation on apparent survival rates of male Lesser Prairie-Chickens. Condor, 107, 78–86.
Hedenström, A. (1993). Migration by soaring or flapping flight in birds: the relative importance of energy cost and speed. Philosophical Transactions of the Royal Society of London Series B, 342, 353–361.
Houston, D. C. (1975). The moult of the white-backed and Rüpell´s vultures Gyps africanus and G. rueppelli. Ibis, 117, 474–488.
Kochan, J. B. (1996). Birds, wings and tails. Pennsylvania: Stackpole Books.
Lebreton, J. D., Burnham, K. P., Clobert, J., & Anderson, D. R. (1992). Modeling survival and testing biological hypotheses using marked animals: a unified approach with case studies. Ecological Monographs, 62, 67–118.
Madsen, V. (2005). Female mate choice in the Magnificent Frigatebird (Fregata magnificens) (Ph.D. Thesis). Instituto de Ecología, Universidad Nacional Autónoma de México. México D.F.
Madsen, V., Dabelsteen,T., Osorio, D., & Osorno, J. L. (2007). Morphology and ornamentation in Male Magnificent Frigatebirds: variation with age class and mating status. American Naturalist, 169, S93–S111.
Møller, A. P. (1990). Fluctuating asymmetry in male sexual ornaments may reliably reveal male quality. Animal Behaviour, 40, 1185–1187.
Møller, A. P. (1991). Sexual ornament size and the cost of fluctuating asymmetry. Proceedings of the Royal Society of London Series B, 234, 59–62.
Møller, A. P. (1992). Female swallow preference for symmetrical male sexual ornaments. Nature, 375, 238–240.
Møller, A. P., & Höglund, J. (1991). Patterns of fluctuating asymmetry in avian feather ornaments: implications for models of sexual selection. Proceedings of the Royal Society of London Series B, 245, 1–5.
Norberg, R. A. (1994). Swallow tail streamer is a mechanical device for self-deflection of tail leading edge, enhancing aerodynamic efficiency and flight manoeuvrability. Proceedings of the Royal Society of London Series B, 257, 227–233.
Norberg, U. M. (1990). Vertebrate flight. Berlin: Springer-Verlag.
Osorno, J. L. (1996). Evolution of breeding behavior in the Magnificent Frigatebird: copulatory pattern and parental investment (Ph.D.Thesis). University of Florida. Gainesville, Florida, USA.
Osorno, J. L. (1999). Offspring desertion in the Magnificent Frigatebrid: are males facing a trade- off between current and future reproduction? Journal of Avian Biology, 30, 335–341.
Osorno, J. L., & Székely, T. (2004). Sexual conflict and parental care in Magnificent Frigatebirds: full compensation by deserted females. Animal Behaviour, 68, 337–342.
Palmer, A. R., & Strobeck, C. (1986). Fluctuating asymmetry: measurement, analysis, pattern. Annual Review of Ecology and Systematics, 17, 391–421.
Pennycuick, C. J. (1983). Thermal soaring compared in three dissimilar tropical bird species, Fregata magnificens, Pelecanus occidentalis, and Coragyps atratus. Journal of Experimental Biology, 102, 307–325.
Pryke, S. R., & Andersson, S. (2005). Experimental evidence for female choice and energetic costs of male tail elongation in Red-collared Widowbirds. Biological Journal of the Linnean Society, 86, 35–43.
Rayleigh, L. (1883). The soaring flight of birds. Nature, 27, 534–535.
Sandercock, B. K., Székely, T., & Kosztolányi, A. (2005). The effects of age and sex on the apparent survival of Kentish Plovers breeding in southern Turkey. Condor, 107, 582–595.
Sapir, N., Horvitz, N., Wikelski, M., Avissar, R., Mahrer, Y., & Nathan, R. (2011). Migration by soaring or flapping: numerical atmospheric simulations reveal that turbulence kinetic energy dictates bee-eater flight mode. Philosophical Transactions of the Royal Society of London Series B, 278, 3380–3386.
Shnell, G. D. (1974). Flight speeds and wingbeat frequencies of the Magnificent Frigatebird. Auk, 91, 564–570.
Swaddle, J. P. (1999). Limits to length asymmetry detection in Starlings: implications for biological signaling. Philosophical Transactions of the Royal Society of London Series B, 266, 1299–1303.
Thomas, A. L. R. (1993a). On the aerodynamics of birds’ tails. Philosophical Transactions of the Royal Society of London Series B, 340, 361–380.
Thomas, A. L. R. (1993b). The aerodynamic costs of asymmetry in the wings and tail of birds: asymmetric birds can’t fly round tight corners. Proceedings of the Royal Society of London Series B, 254, 181–189.
Weimerskirch, H., Chastel, O., Barbraud, C., & Tostain, O. (2003). Frigatebirds ride high on thermals. Nature, 421, 333–334.
Weimerskirch, H., Le Corre, M., Marsac, F., Barbraud, C., Tostain, O., & Chastel, O. (2006). Postbreeding movements of frigatebirds tracked with satellite telemetry. Condor, 108, 220–225.
White, G. C., & Burnham, K. P. (1999). Program MARK: survival estimation from populations of marked animals. Bird Study, 46, S120–S139.


