Zooplankton biodiversity in tropical karst lakes of southeast Mexico, Chiapas
Rocío Fernández, Luis A. Oseguera, Javier Alcocer *
Grupo de Investigación en Limnología Tropical, Facultad de Estudios Superiores-Iztacala, Universidad Nacional Autónoma de México, Av. de los Barrios 1, Los Reyes Iztacala, 54090 Tlalnepantla, Estado de México, Mexico
*Corresponding author: jalcocer@unam.mx (J. Alcocer)
Received: 26 August 2019; accepted: 17 February 2020
Abstract
This study describes the biodiversity of the limnetic zooplankton communities of 18 tropical karst lakes of the Lagunas de Montebello National Park, Chiapas. Sampling was carried out in 2 contrasting climatic periods. A total of 59 zooplankton species were identified (25 Copepoda, 13 Cladocera and 21 Rotifera). Montebello’s global biodiversity is relatively high (∼11%) with reference to what is reported for the entire country. However, the species richness per lake varies from 4 to 24 species with an average of 10.2 ± 6.0. The most frequent species are the copepods Mastigodiaptomus maya and M. nesus, the cladocerans Diaphanosoma cf. birgei, Ceriodaphnia dubia and Eubosmina tubicen, and the rotiferan Keratella americana. Eutrophic lakes showed higher species richness and share more species in common than oligotrophic lakes; the species richness of the deep lakes is lower than that of the shallow, and that of the plateau doubles those of the mountain lakes. M. maya is the only species inhabiting all lake types. The high singularity of the zooplankton of each lake demonstrates the fragility of these ecosystems so pollution and/or eutrophication threaten the high regional zooplankton biodiversity.
Keywords: Species richness; Copepoda, Cladocera; Rotifera; Lagunas de Montebello National Park
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Biodiversidad de zooplankton en lagos tropicales kársticos del sureste de México, Chiapas
Resumen
Este estudio presenta la biodiversidad de las comunidades limnéticas de zooplancton de 18 lagos kársticos tropicales del Parque Nacional Lagunas de Montebello, Chiapas. Los muestreos se llevaron a cabo en 2 períodos climáticos contrastantes. Se identificaron un total de 59 especies de zooplankton (25 Copepoda, 13 Cladocera y 21 Rotifera). La biodiversidad global de Montebello es relativamente alta (∼11%) con referencia a lo reportado para todo el país. Sin embargo, la riqueza específica por lago varía de 4 a 24 especies con un promedio de 10.2 ± 6.0 especies. Las especies que se registraron en un mayor número de lagos fueron: los copépodos Mastigodiaptomus maya y M. nesus, los cladóceros Diaphanosoma cf birgei, Ceriodaphnia dubia y Eubosmina tubicen, y el rotífero Keratella americana. Los lagos eutróficos mostraron una mayor riqueza específica y comparten más especies en común que los lagos oligotróficos; la riqueza específica de los lagos profundos es menor a la de los someros y la de los lagos de planicie duplica a los de montaña. La elevada singularidad del zooplankton de cada lago evidencia la fragilidad de estos ecosistemas por lo que su contaminación y/o eutroficación amenaza la elevada biodiversidad regional de zooplankton.
Palabras clave: Riqueza específica; Copepoda, Cladocera; Rotifera; Parque Nacional Lagunas de Montebello
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
Previous taxonomic studies dealing with zooplankton allow recognizing the great biodiversity of Mexican inland waters. Up to date, 300 rotifer species have been recorded in Mexico, this is, ≈14% of the total rotifer species reported worldwide (García-Morales & Elías-Gutiérrez, 2013; Sarma & Elías-Gutiérrez, 1999). In addition, Elías-Gutiérrez et al. (2008) and Mercado-Salas and Suárez-Morales (2011) enumerated 150 species of cladocerans and 90 species of copepods in Mexico. Currently, a total of 540 zooplankton species have been reported from Mexican inland waters.
Taxonomic lists are not only useful in recording the biodiversity but also in recognizing zoogeographic patterns, and even relating species to water quality (Sládeček, 1983). Based on the presence or absence, frequency and density, and water quality, diverse indexes have been developed to use zooplankton species as bioindicators of trophic state, water pollution and other water characteristics from epicontinental aquatic ecosystems (Lougheed & Chow-Fraser, 2002).
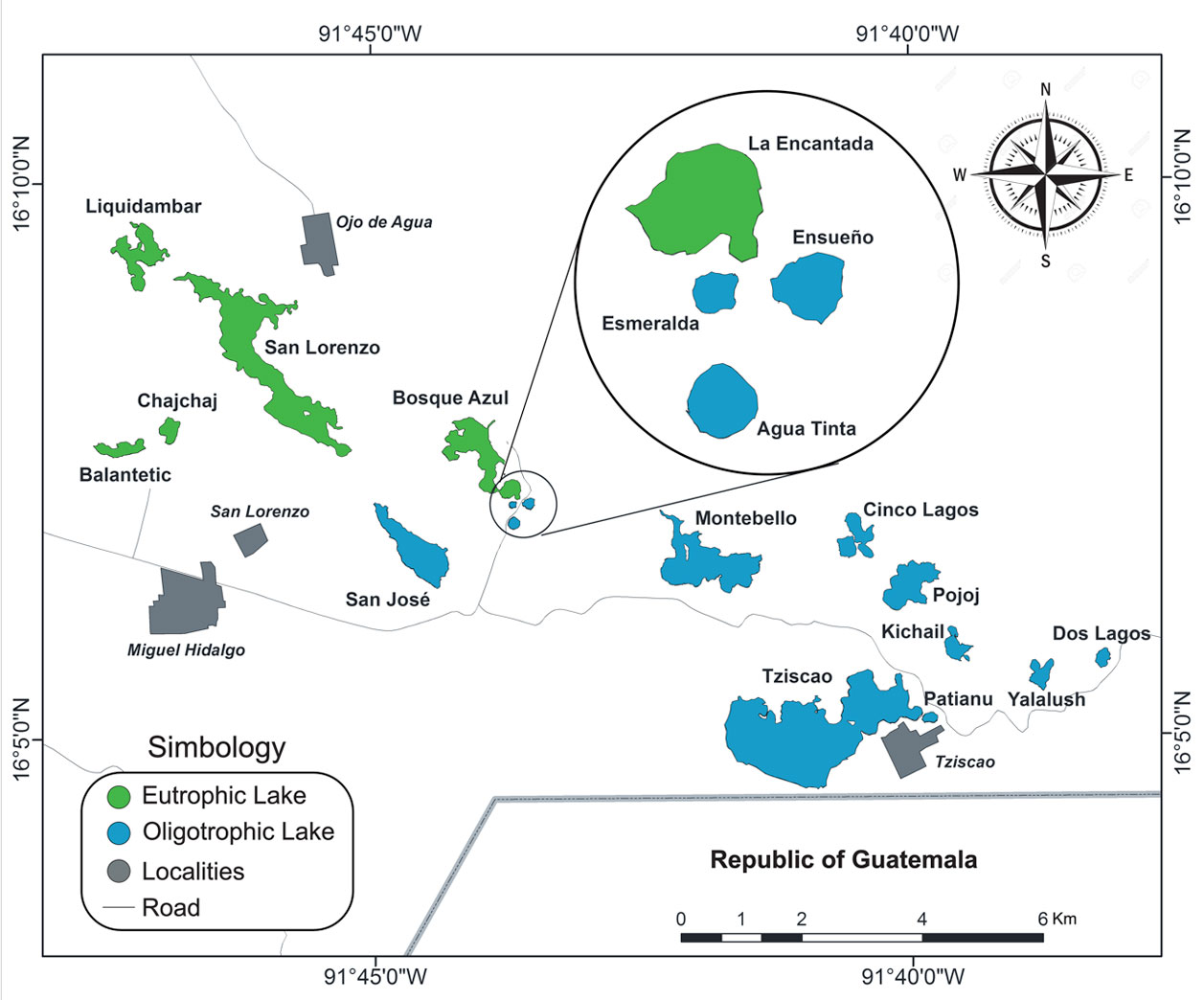
The heterogeneous orographic conformation of the Mexican territory shaped an ample diversity of inland (lotic and lentic) waters (Alcocer & Bernal-Brooks, 2010). Nonetheless, large lake-districts could be recognized in Mexico; of particular interest are those lakes originated by dissolution on the ample karst landscapes of the eastern and southeastern Mexican territories. Well-known examples in this area are the “cenotes” (sinkholes) of the Yucatán Peninsula and the Lagunas de Montebello National Park (LMNP) in Chiapas.
The LMNP includes more than 60 lakes with a broad array of bathymetric and morphometric characteristics (Alcocer et al., 2016, 2018). The lake diversity provides a unique opportunity to uncover the largely unknown inland aquatic biodiversity of Chiapas and Mexico. A previous study on the LMNP benthic invertebrate’s biodiversity (Cortés-Guzmán et al., 2019) showed a high regional biodiversity but at the same time a large singularity of the benthic invertebrate fauna from each lake, so we expect similar results for the zooplankton. The aim of the present study is to provide accurate data on the largely unknown biodiversity of the zooplankton communities of the LMNP lakes, Chiapas, Mexico. It should be noted that there are only 2 previous published studies concerning zooplankton of this area both describing new species: Eucyclops tziscao from lake Tziscao (Gutiérrez-Aguirre & Cervantes-Martínez, 2013) and Mastigodiaptomus suarezmorelesi from lake Montebello (Gutiérrez-Aguirre et al., 2013).
Materials and methods
The LMNP is in the southeastern region of Chiapas State, at the border with the Guatemala Republic (Fig. 1). Extreme coordinates of the LMNP are 16°04’40” – 16°10’2” N, 91°37’40” – 91°47’40” W (Conanp, 2007), with an altitude between 1,200 and 1,800 m asl (Durán-Calderón et al., 2014). It covers an area of 6,022 h and includes part of 2 municipalities, La Independencia and La Trinitaria, the latter encompassing 95% of the LMNP.
Climate in the LMNP is mostly C(fm) type (humid temperate with rains all year round), while in the extreme NW is A(cm) type (warm humid with abundant rains in summer and with absence of frosts in the coldest months) (Conanp, 2009). The average monthly temperature is 23.6°C with an annual rainfall of 1,862 mm (García, 2004).
The LMNP belongs to the Hydrological Region No. 30 Grijalva-Usumacinta (Conanp, 2007). The main watercourse is the Río Grande. Mesozoic to Lower Cretaceous series composes the limestone-type sedimentary class lithology. The lake district is karst landscape (solution lakes) with dolines, uvalas and poljes (Alcocer et al., 2016). There are more than 60 lakes (just in Mexico) along a NW-SE oriented fault system. Durán-Calderon et al. (2014) classified the lakes in 2 large groups: mountain lakes, mostly groundwater fed, and plateau lakes, largely fed by surface runoff (Río Grande).
We selected 18 water bodies along the fault system covering the entire lake district. The lakes included shallow and deep, oligotrophic and eutrophic, and mountain and plateau lakes (Table 1). Further geographical, bathymetric and morphometric information is provided elsewhere (Alcocer et al., 2016).
Two field campaigns were carried out in 2014, the first one in the cold and dry season (February) and the second in the warm and rainy season (October). Water samples were obtained with an UWITEC water sampler bottle at 2 (shallow lakes) or 5 (deep lakes) water depths, along the water column. Water samples were filtered in situ through a 54μm aperture mesh; retained organisms were fixed with 4% formaldehyde. We used specialized taxonomic keys to identify the rotifers (Koste, 1978, Sarma & Nandini, 2017), and cladocerans and copepods (Benzie, 2005; Elías-Gutiérrez et al. 2008; Korovchinsky, 1992, 2002; Mercado-Salas et al., 2012).
To verify the similarity of the taxonomic composition between lakes, a dendrogram was made based on the species presence-absence by means of the Jaccard index and the linkage between groups with SPSS v21.
Results
We identified 59 zooplankton taxa, 90% identified to species and 10% at genus level; from here on, all taxa are referred to as “species” (Table 2). Out of the 59, 47 species are new records for Chiapas, and 2 (Daphnia gessneri and D. hyalina) are new records for Mexico. In addition, we found 4 species of copepods (Arctodiaptomus dorsalis, Mesocyclops aspericornis, M. thermocyclopoides and Thermocyclops crassus) usually reported as exotic or invasive species (Elías-Gutiérrez et al., 2008; Suárez-Morales et al., 2011).
There are 25 copepods (42.4%), 13 cladoceran (22.0%) and 21 rotifer (35.6%) species. Among copepods, 7 are Calanoidea (Diaptomindae) and 18 Cyclopoida (Cyclopidae). There are 13 Cladocera species represented by 4 families, Sididae (1), Daphniidae (8), Moinidae (2) and Bosminidae (2). Finally, 21 species belong to Rotifera, subclass Monogononta, with 8 families.
Table 3 shows the distribution of the zooplankton species in the 18 lakes; out of the 59 species, 44% were restricted to a single lake, and 27% of the species were found in 2-3 lakes. Regarding Calanoidea, 2 species (Mastigodiaptomus maya and M. nessus) are present in most of the lakes, and the remaining 5 species were restricted to a single lake. 10 species of the Cyclopoidea are restricted to a single lake while 8 species were distributed in 2 to 6 lakes; Microcyclops ceibaensis was the most distributed species found in 6 lakes. Three cladocerans (Diaphanosma birgei, Ceriodaphnia dubia and Eubosmina tubicen) were observed in more than 9 lakes, 4 species were distributed between 2 to 5 lakes, and 46% of cladocerans (6 species) were restricted to only 1 lake. Only Keratella americana out of the 21 rotifer species was observed in 9 lakes, 3 rotifer species were present in 4 to 6 lakes, and the remaining 15 species were distributed in 1-3 lakes.
Table 1
List of lakes of the PNLM included in this study and their classification in the main categories. Depth and thermal type according to Alcocer et al. (2016, 2018), trophic state according to Vera-Franco et al. (2015), and geographic site type according to Durán-Calderon et al. (2014).
|
Lake |
Abr. |
Depth |
Thermal type |
Trophic state |
Site type |
|
Balantetic |
BL |
Shallow |
Warm polymictic |
Eutrophic |
Plateau |
|
Chajchaj |
CH |
Shallow |
Warm polymictic |
Eutrophic |
Plateau |
|
Liquidámbar |
LI |
Deep |
Warm monomictic |
Eutrophic |
Plateau |
|
San Lorenzo |
SL |
Deep |
Warm monomictic |
Eutrophic |
Plateau |
|
Bosque Azul |
BA |
Deep |
Warm monomictic |
Eutrophic |
Plateau |
|
La Encantada |
EC |
Deep |
Warm monomictic |
Eutrophic |
Plateau |
|
Esmeralda |
ES |
Shallow |
Warm polymictic |
Oligotrophic |
Mountain |
|
Ensueño |
EN |
Deep |
Warm monomictic |
Oligotrophic |
Mountain |
|
Agua Tinta |
AT |
Deep |
Warm monomictic |
Oligotrophic |
Mountain |
|
San José |
SJ |
Deep |
Warm monomictic |
Oligotrophic |
Plateau |
|
Montebello |
MB |
Deep |
Warm monomictic |
Oligotrophic |
Mountain |
|
Cinco Lagos |
CL |
Deep |
Warm monomictic |
Oligotrophic |
Mountain |
|
Pojoj |
PO |
Deep |
Warm monomictic |
Oligotrophic |
Mountain |
|
Kichail |
KI |
Deep |
Warm monomictic |
Oligotrophic |
Mountain |
|
Tziscao |
TZ |
Deep |
Warm monomictic |
Oligotrophic |
Mountain |
|
Patianú |
PA |
Deep |
Warm monomictic |
Oligotrophic |
Mountain |
|
Yalalush |
YA |
Shallow |
Warm polymictic |
Oligotrophic |
Mountain |
|
Dos Lagos |
DL |
Deep |
Warm monomictic |
Oligotrophic |
Mountain |
Zooplankton species richness (SZOO) ranged from 4 (6.8%) to 24 (40.7%) species with an average of 10.2 ± 6.0 species per lake. Six lakes (San Lorenzo, Chajchaj, Balantetic, Bosque Azul, La Encantada and Tziscao) out of 18 displayed high SZOO (12-24 species). On the contrary, 4 lakes (Esmeralda, Pojoj, Ensueño and Montebello) displayed the lowest SZOO values (< 7 species) (Fig. 2).
Copepods were found in the 18 lakes. The copepod species richness (SCOP) averaged 3.9 ± 2.5 species (1-9 species). The highest SCOP was found in Bosque Azul (9 species), and San Lorenzo and Tziscao (8 species each). The cladocerans species richness (SCLAD) averaged 2.8 ± 1.7 species (0-5 species). The highest SCLAD was found in Bosque Azul, La Encantada and Tziscao (5 species), and the lowest (0, absence) in Liquidambar and Chajchaj. Finally, the average rotifer species richness (SROT) was 3.5 ± 4.1 species (0-12 species). The highest rotifer species richness (SROT) was found in San Lorenzo (12 species), Chajchaj (11 species) and Balantetic (10 species), and the lowest (0, absence) in Ensueño, San José, Pojoj, Pataniu and Yalalush (Fig. 2)
The SCOP/SCLAD/SROT ratio of the Montebello lakes averages 1.4/1.0/1.2. The ratio was different in eutrophic lakes (1.6/1.0/3.3) where SROT augmented while SCOP diminished. In the oligotrophic lakes (1.2/1.0/0.4) the ratio inverted with SROT diminishing while SCOP augmenting. The ratio in plateau (1.3/1.0/2.1) and mountain lakes (1.2/1.0/0.4) was similar to the eutrophic and oligotrophic lakes, respectively.
There is a large singularity of zooplankton species in each lake. Although a single species could be found between 1 and up to 16 lakes (not a single species was found inhabiting the 18 or even 17 lakes), most species (76%) were found inhabiting between 1 and 3 lakes (Fig. 3). One species (Mastigodiaptomus maya) was found in 16 lakes, 2 species (M. nesus, Diaphanosoma cf. birgei) were found in 11 lakes, 1 species (Ceriodaphnia dubia) was found in 10 lakes, and 2 species (Eubosmina tubicen, Keratella americana) were found in 9 lakes. The other 53 species (≈ 90%) were found in less than 50% (< 9) of the lakes.
Table 2
Taxonomic list of the zooplankton species (Copepoda, Cladocera, Rotifera) recorded in the 18 lakes of the Lagunas de Montebello National Park, Chiapas.
Order Calanoida Sars, 1903
Family Diaptomidae Sars, 1903
Subfamily Diaptominae Kiefer, 1932
|
1. Arctodiaptomus dorsalis (Marsh, 1907) |
|
2. Leptodiaptomus cuauhtemoci (Osorio-Tafall, 1941) |
|
3. L. mexicanus (Marsh, 1929) |
|
4. Mastigodiaptomus maya Suárez-Morales y Elías-Gutierrez, 2000 |
|
5. M. nesus Bowman, 1986 |
|
6. M. suarezmoralesi Gutiérrez-Aguirre y Cervantes-Martínez, 2013 |
|
7. Mastigodiaptomus sp. 2 Light, 1939 |
|
Order Cyclopoida Burmeister, 1834 |
|
Family Cyclopidae Dana, 1846 |
|
Subfamily Cyclopinae Kiefer, 1927 |
|
8. Apocyclops panamensis (Marsh, 1913) |
|
9. Diacyclops puuc Fiers y Reid, 1996 |
|
10. Mesocyclops aspericornis (Daday, 1906) |
|
11. M. brasilianus Kiefer, 1933 |
|
12. M. edax (Forbes, 1981) |
|
13. M. longisetus (Thiébaud, 1912) |
|
14. M. pescei Petkovski, 1986 |
|
15. M. thermocyclopoides Harada, 1931 |
|
16. Microcyclops ceibaensis (Marsh, 1919) |
|
17. M. dubitabilis (Kiefer, 1934) |
|
18. Thermocyclops tenuis (Marsh, 1910) |
|
19. T. inversus Kiefer, 1936 |
|
20. T. crasssus (Fischer, 1853) |
|
Subfamily Eucyclopinae Kiefer, 1927 |
|
21. Ectocyclops sp. Brady, 1904 |
|
22. Eucyclops tziscao Mercado-Salas, 2013 |
|
23. Macrocyclops albidus (Jurine, 1820) |
|
24. M. fuscus (Jurine, 1820) |
|
25.Tropocyclops prasinus prasinus (Fischers, 1995) |
|
Order Cladocera Milne-Edwards, 1840 |
|
Family Sididae Baird, 1850 (emend. Sars, 1865) |
|
26. Diaphanosoma cf birgei Korineck, 1981 |
|
Family Daphniidae Straus, 1820 |
|
Subgenus Daphnia O.F. Müller, 1785 |
|
27. Daphnia ambigua Scourfield, 1947 |
|
28. D. cheraphila Hebert & Finston, 1996 |
|
29. D. cf galeata Sars, 1864 |
|
30. D. gessneri Herbst, 1967 |
|
31. D. cf hyalina Leydig, 1860 |
|
32. D. obtusa Kurz, 1875 |
|
33. Ceriodaphnia dubia Richard, 1894 |
|
34. C. lacustris Birge, 1893 |
|
Family Moinidae Goulden, 1968 |
|
35. Moina micrura Kurz, 1874 |
|
36. Moinodaphnia sp. Herrick, 1887 |
|
Family Bosminidae Baird, 1845 (emend. Sars, 1865) |
|
37. Eubosmina tubicen Brehm, 1953 |
|
38. Bosmina sp. Baird, 1845 |
|
Phylum Rotifera Cuvier, 1798 |
|
Subclase Monogononta Plate, 1889 |
|
Order Flosculariaceae Harring, 1913 |
|
Family Hexarthridae Bartos, 1959 |
|
39. Hexarthra intermedia (Wisniewski, 1929) |
|
Family Testudinellidae Harring, 1913 |
|
40. Testudinella patina (Hermann, 1783) |
|
41. Ptygura sp. Ehrenberg, 1832 |
|
Family Trochosphaeridae Harring, 1913 |
|
42. Filinia longiseta (Ehrenberg, 1834) |
|
43. F. terminalis (Plate, 1886) |
|
Order Ploima (Hudson & Gosse, 1886) |
|
Family Asplanchnidae Eckstein, 1883 |
|
44. Asplanchna girodi de Guerne, 1888 |
|
Family Brachionidae Ehrenberg, 1838 |
|
45.Brachionus angularis Gosse, 1851 |
|
46. B. bidentatus Anderson, 1889 |
|
47. B. calyciflorus Pallas, 1766 |
|
48. B. havanaensis Rousselet, 1911 |
|
49. B. rubens Ehrenberg, 1838 |
|
50. Keratella americana Carlin (1943) |
|
51. K. cochlearis (Gosse, 1851) |
|
52. K. tropica (Apstein, 1907) |
|
53. Plationus patulus (Müller, 1786) |
|
54. Platyas quadricornis (Ehrenberg, 1832) |
|
Family Lecanidae Remane, 1933 |
|
55. Lecane bulla (Gosse, 1851) |
|
56. L. papuana (Murray, 1913) |
|
Family Synchaetidae Hudson & Gosse, 1886 |
|
57. Polyarthra vulgaris Carlin, 1943 |
|
58. Synchaeta oblonga Ehrenberg, 1831 |
|
Family Trichotriidae Harring, 1913 |
|
59. Macrochaetus sp. Perty, 1850 |
SZOO was higher (× 2.3 times) in eutrophic lakes with 16.3 ± 6.3 species (7-24 species) than in oligotrophic lakes with 7.2 ± 2.7 taxa (4-14 species). Similar figures (× 2.1 times) were found for SZOO in plateau lakes with 15.0 ± 6.8 species (7-24 species) comparing to mountain lakes with 7.2 ± 2.8 (4-14 species). Differently, SZOO in deep lakes with 9.9 ± 6.1 species (4-24 species) was similar to shallow lakes with 11.3 ± 6.4 species (4-18 species). SZOO in deep eutrophic lakes with 16.3 ± 8.1 species (7-24 species) was similar to shallow eutrophic lakes with 16.5 ± 2.1 species (15-18 species). Finally, SZOO in deep oligotrophic lakes with 7.4 ± 2.7 taxa (4-14 species) was slightly higher than shallow oligotrophic lakes with 6.0 ± 2.8 species (4-8 species).
Table 3
Distribution of the zooplankton species in the 18 lakes of the Lagunas de Montebello National Park, Chiapas. (* = first record in Chiapas, ** = first record in Chiapas and reported as exotic/invasive, X = previously reported in Chiapas, *X = first record in Mexico).
|
Table 3 Continued |
||||||||||||||||||
|
Copepoda |
BL |
CH |
LI |
SL |
BA |
EC |
ES |
EN |
AT |
SJ |
MB |
CL |
PO |
KI |
TZ |
PA |
YA |
DL |
|
Copepoda |
BL |
CH |
LI |
SL |
BA |
EC |
ES |
EN |
AT |
SJ |
MB |
CL |
PO |
KI |
TZ |
PA |
YA |
DL |
|
Arctodiaptomus dorsalis |
** |
|||||||||||||||||
|
Leptodiaptomus cuauhtemoci |
* |
|||||||||||||||||
|
L. mexicanus |
* |
|||||||||||||||||
|
Mastigodiaptomus maya |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
||
|
M. nesus |
* |
* |
* |
* |
* |
* |
* |
* |
* |
* |
* |
|||||||
|
M. suarezmoralesi |
X |
|||||||||||||||||
|
Mastigodiaptomus sp. 2 |
* |
|||||||||||||||||
|
Apocyclops panamensis |
* |
* |
* |
* |
* |
|||||||||||||
|
Diacyclops puuc |
* |
* |
||||||||||||||||
|
Mesocyclops aspericornis |
** |
|||||||||||||||||
|
M. brasilianus |
X |
|||||||||||||||||
|
M. edax |
* |
* |
* |
|||||||||||||||
|
M. longisetus |
X |
|||||||||||||||||
|
M. pescei |
X |
|||||||||||||||||
|
M. thermocyclopoides |
** |
|||||||||||||||||
|
Microcyclops ceibaensis |
* |
* |
* |
* |
* |
* |
||||||||||||
|
M. dubitabilis |
* |
* |
||||||||||||||||
|
Thermocyclops tenuis |
* |
|||||||||||||||||
|
T. inversus |
* |
* |
* |
* |
* |
|||||||||||||
|
T. crasssus |
** |
|||||||||||||||||
|
Ectocyclops sp. |
* |
* |
||||||||||||||||
|
Eucyclops tziscao |
X |
X |
X |
|||||||||||||||
|
Macrocyclops albidus |
* |
|||||||||||||||||
|
M. fuscus |
* |
|||||||||||||||||
|
Tropocyclops prasinus |
* |
|||||||||||||||||
|
Subtotal |
2 |
7 |
1 |
8 |
9 |
3 |
2 |
4 |
3 |
3 |
2 |
2 |
2 |
3 |
7 |
3 |
7 |
2 |
|
Cladocera |
||||||||||||||||||
|
Diaphanosoma cf birgei |
* |
* |
* |
* |
* |
* |
* |
* |
* |
* |
* |
|||||||
|
Daphnia ambigua |
* |
* |
||||||||||||||||
|
D. cheraphila |
* |
|||||||||||||||||
|
D. cf galeata |
* |
* |
* |
|||||||||||||||
|
D. gessneri |
*X |
*X |
*X |
*X |
*X |
*X |
||||||||||||
|
D. cf hyalina |
*X |
|||||||||||||||||
|
D. obtusa |
* |
|||||||||||||||||
|
Ceriodaphnia dubia |
* |
* |
* |
* |
* |
* |
* |
* |
* |
* |
||||||||
|
C. lacustris |
* |
* |
* |
* |
||||||||||||||
|
Moina micrura |
* |
|||||||||||||||||
|
Moinodaphnia sp. |
* |
|||||||||||||||||
|
Eubosmina tubicen |
* |
* |
* |
* |
* |
* |
* |
* |
* |
|||||||||
|
Bosmina sp. |
* |
|||||||||||||||||
|
Subtotal |
0 |
4 |
0 |
3 |
6 |
5 |
1 |
1 |
3 |
4 |
2 |
2 |
2 |
4 |
5 |
4 |
2 |
4 |
|
Rotifera |
||||||||||||||||||
|
Hexarthra intermedia |
X |
X |
X |
X |
||||||||||||||
|
Testudinella patina |
* |
* |
* |
|||||||||||||||
|
Ptygura sp. |
* |
* |
* |
|||||||||||||||
|
Filinia longiseta |
* |
* |
* |
* |
* |
* |
||||||||||||
|
F. terminalis |
* |
* |
* |
|||||||||||||||
|
Asplanchna girodi |
* |
* |
* |
|||||||||||||||
|
Brachionus angularis |
* |
* |
* |
|||||||||||||||
|
B. bidentatus |
* |
|||||||||||||||||
|
B. calyciflorus |
* |
* |
* |
|||||||||||||||
|
B. havanaensis |
X |
X |
X |
X |
X |
X |
||||||||||||
|
B. rubens |
* |
* |
||||||||||||||||
|
Keratella americana |
* |
* |
* |
* |
* |
* |
* |
* |
* |
|||||||||
|
K. cochlearis |
* |
* |
* |
|||||||||||||||
|
K. tropica |
* |
* |
* |
|||||||||||||||
|
Plationus patulus |
* |
* |
* |
|||||||||||||||
|
Platyas quadricornis |
* |
|||||||||||||||||
|
Lecane bulla |
* |
* |
||||||||||||||||
|
L. papuana |
X |
X |
||||||||||||||||
|
Polyarthra vulgaris |
* |
|||||||||||||||||
|
Synchaeta oblonga |
* |
|||||||||||||||||
|
Macrochaetus sp. |
* |
|||||||||||||||||
|
Subtotal |
6 |
12 |
11 |
10 |
8 |
4 |
1 |
0 |
2 |
0 |
2 |
3 |
0 |
2 |
1 |
0 |
0 |
1 |
|
Total |
7 |
24 |
18 |
15 |
23 |
12 |
4 |
5 |
8 |
7 |
6 |
7 |
4 |
9 |
13 |
7 |
9 |
7 |

Figure 4 shows the groups of lakes formed by the cluster analysis based on species presence/absence of the Montebello lakes. The most similar lakes are: eutrophic San Lorenzo and Bosque Azul, and oligotrophic Pojoj, Patianú, and San José; the rest of the lakes turn out to be taxonomically different. Nonetheless and with a lower level of similarity, the eutrophic lakes clustered together, and the oligotrophic lakes formed another cluster. As previously explained, most species are restricted to a single or a few lakes instead of showing an ample distribution; this explains the low degree of similarity between the lakes.
Out of the 59 species registered for the Montebello lakes, 8 (2 copepods, 2 cladocerans and 4 rotifers) characterized (present in ≥ 65% of each lake type) the different lake types (Table 4). Six species characterized eutrophic lakes (1 copepod, 1 cladoceran, and 4 rotifers), while 3 species characterized oligotrophic lakes (2 copepods and 1 cladoceran); the only species in common for eutrophic and oligotrophic lakes was the copepod Mastigodiaptomus maya.
Seven species characterized plateau lakes (1 copepod, 2 cladocerans, and 4 rotifers), while 2 characterized mountain lakes (2 copepods); the only species in common for plateau and mountain lakes was, once again, the copepod Mastigodiaptomus maya. Finally, 2 species characterized deep lakes (1 copepod, and 1 cladoceran), while 1 species characterized shallow lakes (1 copepod); the only species in common in deep and shallow lakes was, one more, the copepod Mastigodiaptomus maya. In this way, the copepod Mastigodiaptomus maya was the only species common to all lake types. Not a single species of cladoceran or rotifer were common to all lake types.

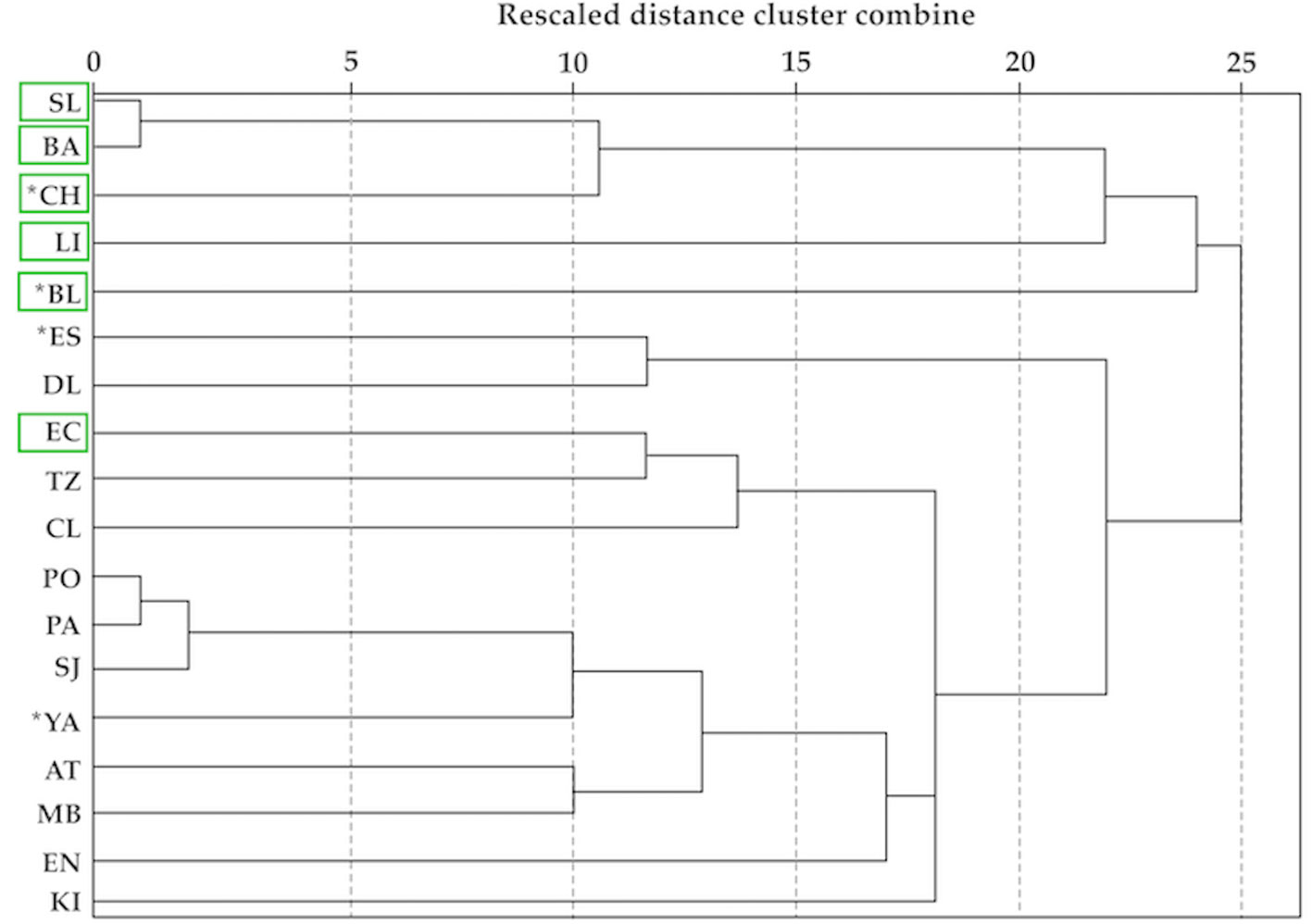
Table 4
Characteristic (most frequently found ≥ 65% of lakes) zooplankton species cataloged by lake type in the 18 lakes of the Lagunas de Montebello National Park, Chiapas. E = Eutrophic lakes, O = oligotrophic lakes, P = plateau lakes, M = mountain lakes, D = deep lakes, S = shallow lakes.
|
Species |
E |
O |
P |
M |
D |
S |
|
Copepoda |
||||||
|
Mastigodiaptomus maya |
X |
X |
X |
X |
X |
X |
|
Mastigodiaptomus nesus |
– |
X |
– |
X |
– |
– |
|
Subtotal |
1 |
2 |
1 |
2 |
1 |
1 |
|
Cladocera |
||||||
|
Diaphanosoma cf birgei |
– |
X |
X |
– |
X |
– |
|
Ceriodaphnia dubia |
X |
– |
X |
– |
– |
– |
|
Subtotal |
1 |
1 |
2 |
0 |
1 |
0 |
|
Rotifera |
||||||
|
Filinia longiseta |
X |
– |
X |
– |
– |
– |
|
Brachionus havanaensis |
X |
– |
– |
– |
– |
– |
|
Keratella americana |
X |
– |
X |
– |
– |
– |
|
Hexarthra intermedia |
X |
– |
X |
– |
– |
– |
|
Subtotal |
4 |
0 |
4 |
0 |
0 |
0 |
|
Total |
6 |
3 |
7 |
2 |
2 |
1 |
Discussion
The zooplankton species richness in the limnetic (pelagic) zone of the Montebello lakes is comparatively high (∼11% of the total SZOO reported from Mexico), including 27.7% copepods, 8.7% cladocerans, and 7.0% rotifers those numbers reported for Mexico (90 copepods, 150 cladocerans, and 300 rotifers). However, these figures will surely increase by incorporating the species inhabiting in the littoral zone that are generally more numerous than those dwelling in the limnetic zone.
It is generally accepted that oligotrophic lakes shelter a larger number of species compared with the eutrophic ones (e.g., Pinto-Coelho et al., 2005). This is in contrast with what we found in the Montebello lakes where SZOO was 43 species in eutrophic and 31 species in oligotrophic lakes, this is, approximately 1.4 times higher in eutrophic than in oligotrophic lakes. This is also opposite to Cortés-Guzmán et al. (2019), who found deep benthic macroinvertebrates of the Montebello lakes to be more diverse in oligotrophic than in eutrophic lakes.
Montebello eutrophic lakes share a higher number of species in common (60.5% of taxa in ≥ 2 lakes) than the oligotrophic lakes (48.4% of taxa in ≥ 2 lakes). Since eutrophic lakes are: 1) located at the plateau area, 2) fed largely by surface run-off and the Río Grande, and 3) artificially interconnected through channels, the zooplankton from one lake could easily disperse to others nearby. Also, the flooding events during the rainy season, characteristics of the tropical karst regions, interconnect the lakes otherwise separated on the surface, and then zooplankton could also be spread among adjacent lakes. Differently, mountain lakes are isolated and fed mostly by groundwater that limits zooplankton dispersal.
The zooplankton of eutrophic lakes comprises mainly cosmopolitan species, mostly rotifers (Beaver & Crisman, 1990; Jeppensen et al., 2011; Lougheed & Chow-Fraser, 2002; Sládeček, 1983). Differently, the zooplankton of oligotrophic lakes is composed mostly by copepods and cladocerans (~ 80%). Some taxa of the oligotrophic lakes indicate oligotrophic conditions such as the cladoceran Daphnia gessneri and the calanoid copepods (Branco et al., 2002; Cervantes-Martínez & Gutiérrez-Aguirre, 2015).
There are differences between the zooplankton of the Montebello lakes and that reported from other temperate and tropical lakes. Copepods showed the largest number of taxa (25) followed by rotifers (21) and cladocerans (13), while other studies (Dodson & Silva-Briano, 1996; Guevara et al., 2009; Papa & Zafaralla, 2011; Pinto-Coelho et al., 2005) report rotifers and cladocerans as the dominant groups. Diaphanosoma, Ceriodaphnia, Macrothix and Moina are frequently reported as dominant in tropical and subtropical lakes (Dodson & Frey, 1991; Sarma et al., 2005), while in Montebello the dominant gender is Daphnia with 6 species out of 13 cladocerans (46%).
Elías-Gutiérrez et al. (2008) and Suárez-Morales et al. (2011) reported the presence of 5 invasive or exotic copepods in Mexican inland waters, 4 of them were found in the Montebello lakes: Arctodiaptomus dorsalis (Yalalush), Mesocyclops aspericornis (Bosque Azul), M. thermoscyclopoides (San Lorenzo), and Thermocyclops crassus (Bosque Azul). Three exotic copepods are distributed in eutrophic lakes located in the plateau, and 1, A. dorsalis, was found in an oligotrophic, mountain lake.
Summarizing, the zooplankton species richness of the Montebello lakes with 59 species comprises 27.7%, 8.7%, and 7.0% of those species of copepods, cladocerans, and rotifers, respectively, reported for Mexico. Forty-eight out of the 59 species are new records for Chiapas, 2 species (Daphnia gessneri and D. hyalina) are new records for Mexico, and 4 species (Arctodiaptomus dorsalis, Mesocyclops aspericornis, M. thermocyclopoides and Thermocyclops crassus) are reported as exotic or invasive species.
Slightly less than half the species (26 out of 59, 44%) were found in a single lake, which stresses the singularity of the zooplankton community from each lake, making evident the fragility of these ecosystems. The disappearance of the fauna from a single lake means a high probability of its disappearance from the entire region. Not a single species was found in the 18 lakes; the copepod Mastigodiaptomus maya (but not other copepod, cladocerans, or rotifer) was found inhabiting eutrophic and oligotrophic, plateau and mountain, and deep and shallow lakes. Eutrophic lakes showed higher taxonomic richness compared to the oligotrophic lakes and share more species in common (60.5%), than the oligotrophic lakes (48.4%).
Acknowledgements
This research was granted by the Fondo Sectorial de Investigación y desarrollo sobre el Agua Conagua-Conacyt through the project 167603 and by DGAPA/UNAM through the Projects PAPIIT-IN219215 and PAPIIT-IV200319, and to DGAPA Programa de Becas Posdoctorales for the financial support to RF through a postdoctoral scholarship. We thank Mayra Carolina Reyes Santillan for the identification of cyclopoids copepods. René Morales Hernández and collaborators for support during the fieldwork. We also thank the Parque Nacional Lagunas de Montebello, Comisión Nacional de Áreas Naturales Protegidas (Conanp) (Jesús A. León and Roberto Castellanos), the local community, and the Comisariados Ejidales from Antelá, Cárdenas, Miguel Hidalgo, Ojo de Agua, and Tziscao for facilitating access to the lakes. To the Comité de Administración de Tziscao (Sergio Marcos and Miguel A. Tomas), Presidente del Comité de Turismo de Tziscao (Armando Hernández), Comisario Ejidal de Tziscao (Enrique M. Hernández), and personnel of the Villas Tziscao Hotel (Rosemberg F. Jorge, Juan G. Espinoza and Gemuel P. Hernández) for offering their support and facilities for this study.
References
Alcocer, J., & Bernal-Brooks, F. W. (2010). Limnology in Mexico. Hydrobiologia, 644, 1–54. https://doi.org/10.1007/s10750-010-0211-1
Alcocer, J., Merino-Ibarra, M., Oseguera, L. A., & Escolero, O. (2018). Anthropogenic impacts on tropical karst lakes: “Lagunas de Montebello”, Chiapas. Ecohydrology, 11, e2029. https://doi.org/10.1002/eco.2029
Alcocer, J., Oseguera, L. A., Sánchez, G., González, C. G., Martínez, J. R., & González, R. (2016). Bathymetric and morphometric surveys of the Montebello lakes, Chiapas. Journal of Limnology, 75, 56–65. https://doi.org/10.4081/jlimnol.2016.1343
Beaver, J. R., & Crisman, T. L. (1990). Use of microzooplankton as an early indicator of advancing cultural eutrophication. Internationale Vereinigung für theoretische und angewandte Limnologie: Verhandlungen, 24, 532–537. https://doi.org/10.1080/03680770.1989.11898794
Benzie, J. A. H. (2005). Cladocera: the genus Daphnia (including Daphniosis). Ghent, Leyden: Kenobi Productions & Backhuys Publishers.
Branco, C. W., Rocha, M. I. A., Pinto, G. F., Gômara, G. A., & Filippo, R. D. (2002). Limnological features of Funil Reservoir (RJ, Brazil) and indicator properties of rotifers and cladocerans of the zooplankton community. Lakes & Reservoirs: Research & Management, 7, 87–92. https://doi.org/10.1046/j.1440-169X.2002.00177.x
Cervantes-Martínez, A., & Gutiérrez-Aguirre, M. A. (2015). Physicochemistry and zooplankton of two karstic sinkholes in the Yucatán Peninsula, Mexico. Journal of Limnology, 74, 382–393.https://doi.org/10.4081/jlimnol.2014.976
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2007). Programa de conservación y manejo Parque Nacional Lagunas de Montebello. México D.F.: Secretaría de Medio Ambiente y Recursos Naturales.
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2009). Estudio para el monitoreo de calidad del agua de las lagunas en el Parque Nacional Lagunas de Montebello, Chiapas. México D.F.: Secretaría de Medio Ambiente y Recursos Naturales.
Cortés-Guzmán, D., Alcocer, J., & Oseguera, L. A. (2019). Benthic macroinvertebrate community diversity of Montebello Lakes, Chiapas. Revista Mexicana de Biodiversidad, 90, e902769. http://dx.doi.org/10.22201/ib.20078706e.2019.90.2769
Dodson, S. I., & Frey, D. G. (1991). Cladocera and other Branchiopods. In A. Covich, & J. Thorp (Eds.), Ecology and systematics of North American freshwater invertebrates. San Diego: Academic Press.
Dodson, S. I., & Silva-Briano, M. (1996). Crustacean zooplankton species richness and associations in reservoirs and ponds of Aguascalientes State, Mexico. Hydrobiologia, 325, 163–172.
Durán-Calderón, I., Escolero-Fuentes, O. A., Muñoz Salinas, E., Castillo-Rodríguez, M., & Silva-Romo, G. (2014). Cartografía geomorfológica a escala 1: 50,000 del Parque Nacional Lagunas de Montebello, Chiapas (México). Boletín de la Sociedad Geológica Mexicana, 66, 263–277.
Elías-Gutiérrez, M., Suárez-Morales, E., Gutiérrez-Aguirre, M. A., Silva-Briano, M., Granados-Ramírez, J. G., & Garfias-Espejo, T. (2008). Cladocera y Copepoda de las aguas continentales de México. Guía ilustrada. UNAM. México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ El Colegio de la Frontera Sur/ Consejo Nacional de Ciencia y Tecnología/ Secretaría de Medio Ambiente y Recursos Naturales.
García, E. (2004). Modificaciones al Sistema de clasificación climática de Köppen. Instituto de Geografía, UNAM. Ciudad de México: UNAM.
García-Morales, A. E., & Elías-Gutiérrez, M. (2013). DNA barcoding of freshwater Rotifera in Mexico: Evidence of cryptic speciation in common rotifers. Molecular Ecology Resources, 13, 1097–1107. https://doi.org/10.1111/1755-0998.12080
Guevara, G., Lozano, P., Reinoso, G., & Villa, F. (2009). Horizontal and seasonal patterns of tropical zooplankton from the eutrophic Prado Reservoir (Colombia). Limnologica, 39, 128–139. https://doi.org/10.1016/j.limno.2008.03.001
Gutiérrez-Aguirre, M. A., & Cervantes-Martínez, A. (2013). Diversity of freshwater copepods (Maxillopoda: Copepoda: Calanoida, Cyclopoida) from Chiapas, Mexico with a description of Mastigodiaptomus suarezmoralesi sp. nov. Journal of Natural History, 47, 479–498. https://doi.org/10.1080/00222933.2012.742587
Gutiérrez-Aguirre, M. A., Mercado-Salas, N. F., & Cervantes-Martínez, A. (2013). Description of Eucyclops tziscao sp. n., E. angeli sp. n., and a new record of E. festivus Lindberg, 1955 (Cyclopoida, Cyclopidae, Eucyclopinae) in Chiapas, Mexico. Zookeys, 351, 1–30. https://doi.org/10.3897/zookeys.351.5413
Jeppesen, E., Nõges, P., Davidson, T. A., Haberman, J., Nõges, T., Blank, K. et al. (2011). Zooplankton as indicators in lakes: a scientific-based plea for including zooplankton in the ecological quality assessment of lakes according to the European Water Framework Directive (WFD). Hydrobiologia, 676, 279. https://doi.org/10.1007/s10750-011-0831-0
Korovchinsky, N. M. (1992). Sididae & Holopediidae. Guides to the identification of the microinvertebrates of the continental waters of the World, 3. The Hague: Academic Publishing.
Korovchinsky, N. M. (2002). Description of two new species of Diaphanosoma Fischer, 1850 (Crustacea, Branchiopoda, Sididae) from the United States and Canada and species richness of the genus in North America. Hydrobiologia, 489, 45–54.
Koste, W. (1978). Rotatoria. Die Rädertiere Mitteleuropas, 2. Berlin: Gebrüder Borntraeger.
Lougheed, V. L., & Chow-Fraser, P. (2002). Development and use of a zooplankton index of wetland quality in the Laurentian Great Lakes basin. Ecological Applications, 12, 474–486. https://doi.org/10.1890/1051-0761(2002)012[0474:DAUOAZ]
Mercado-Salas, N. F., Pozo, C., Morrone, J. J., & Suárez-Morales, E. (2012). Distribution patterns of the American species of the freshwater genus Eucyclops (Copepoda: Cyclopoida). Journal of Crustacean Biology, 32, 457–464. https://doi.org/10.1163/193724012X626502
Mercado-Salas, N. F., & Suárez-Morales, E. (2011). Morfología, diversidad y distribución de los Cyclopoida (Copepoda) de zonas áridas del centro-norte de México. I. Cyclopinae. Hidrobiológica, 21, 1–25.
Papa, R. D. S., & Zafaralla, M. T. (2011). The composition, diversity and community dynamics of limnetic zooplankton in a tropical caldera lake (lake Taal, Philippines). Raffles Bulletin of Zoology, 59, 1–17.
Pinto-Coelho, R., Pinel-Alloul, B., Méthot, G., & Havens, K. E. (2005). Crustacean zooplankton in lakes and reservoirs of temperate and tropical regions: variation with trophic status. Canadian Journal of Fisheries and Aquatic Sciences, 62, 348–361. https://doi.org/10.1139/f04-178
Sarma, S. S. S., & Elías-Gutiérrez, M. (1999). Rotifers (Rotifera) from four natural water bodies of Central Mexico. Limnologica, 29, 475–483. https://doi.org/10.1016/S0075-9511(99)80054-1
Sarma, S. S. S., & Nandini, S. (2017). Rotíferos mexicanos (Rotifera). Manual de Enseñanza. Estado de México. Tlalnepantla, Edo. De México: UNAM.
Sarma, S. S. S., Nandini, S., & Gulati, R. D. (2005). Life history strategies of Cladocerans: comparisons of tropical and temperate taxa. In H. Segers, & K. Martens (Eds.), Aquatic Biodiversity II (pp. 315–333). Dordrecht: Springer.
Sládeček, V. (1983). Rotifers as indicators of water quality. Hydrobiologia, 100, 169–201.
Suárez-Morales, E., Gutiérrez-Aguirre, M. A., & Mendoza, F. (2011). The Afro-Asian cyclopoid Mesocyclops aspericornis (Crustacea: Copepoda) in eastern Mexico with comments on the distribution of exotic copepods. Revista Mexicana de Biodiversidad, 82, 109–115. http://dx.doi.org/10.22201/ib.20078706e.2011.1


