
María Razo-González a, *, Juan Márquez b, Gabriela Castaño-Meneses a
y Rodolfo Novelo-Gutiérrez c
a Universidad Nacional Autónoma de México, Facultad de Ciencias, Campus Juriquilla, Unidad Multidisciplinaria de Docencia e Investigación, Boulevard Juriquilla #3001, 76230 Juriquilla, Querétaro, México
b Universidad Autónoma del Estado de Hidalgo, Centro de Investigaciones Biológicas, Laboratorio de Sistemática Animal, Km 4.5, carretera Pachuca-Tulancingo s/n, Ciudad del Conocimiento, Col. Carboneras, 42184 Mineral de la Reforma, Hidalgo, México
c Instituto de Ecología, A.C., Red de Biodiversidad y Sistemática, Carretera Antigua a Coatepec 351, El Haya, 91073 Xalapa, Veracruz, México
*Autor para correspondencia: maria_razo@hotmail.com (M. Razo-González)
Recibido: 27 octubre 2020; aceptado: 17 febrero 2021
Resumen
Se identificaron los patrones de distribución de 120 especies de tricópteros de la Sierra de Juárez, Oaxaca, mediante un análisis de parsimonia de endemismos con eliminación progresiva de caracteres (PAE-PCE) y la búsqueda de trazos generalizados. El análisis se basó en los registros de distribución provenientes de colectas en campo y de literatura especializada, y las unidades de estudio fueron las provincias biogeográficas de México. Se identificaron 8 trazos generalizados que coinciden en lo general con los hallados en otros estudios, todos ellos incluyendo a la provincia Veracruzana. Los trazos generalizados se integraron por provincias pertenecientes a la Zona de Transición Mexicana y la región Neotropical. Los 8 trazos generalizados se interceptan o atraviesan la Sierra de Juárez, conformando un nodo panbiogeográfico notablemente más complejo respecto a lo reportado en estudios previos. Esto permite reconocer que su historia biogeográfica ha contribuido considerablemente en su alta biodiversidad, por lo que se considera necesario que sea un área natural protegida.
Palabras clave: Patrones de distribución; Provincias; Parsimonia; Trazos generalizados; Nodos
© 2021 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
The biogeographic complexity of the Sierra de Juarez, Oaxaca, Mexico, revealed through parsimony analysis of endemicity of caddisflies species (Insecta: Trichoptera)
Abstract
The distributional patterns of 120 species of caddisflies recorded in the Sierra de Juarez, Oaxaca, were analyzed by means of parsimony analysis of endemicity with progressive character elimination (PAE-PCE) to identify generalized tracks. The analysis was based on distribution records from field collections and specialized literature, and the study units were the Mexican biogeographic provinces. Eight generalized tracks were identified that generally coincide with those found in other studies, all of them including the Veracruzan province. Tracks include provinces belonging to the Mexican Transition Zone and the Neotropical region. The 8 generalized tracks intersect or cross the Sierra de Juárez, forming a panbiogeographic node that is notably more complex compared to that reported in previous studies. This allows us to recognize that its biogeographic history has contributed considerably to its high biodiversity, and it is necessary that it is a protected natural area.
Keywords: Distributional patterns; Provinces; Parsimony; Generalized tracks; Nodes
© 2021 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
Los Trichoptera constituyen el orden más diverso de insectos exclusivamente acuáticos y se distribuyen en todo el mundo, excepto en la Antártida (de Moor e Ivanov, 2008; Wiggins, 1996). La presencia de los tricópteros en el registro fósil data del período Triásico, por lo que su distribución y biogeografía reflejan los eventos del pasado, que han promovido la diversificación de la biota acuática de la Tierra (Holzenthal et al., 2007). Las larvas de los tricópteros son componentes esenciales de la dinámica trófica y del flujo de energía en los ecosistemas acuáticos (Mackay y Wiggins, 1979). Están considerados entre los organismos acuáticos más útiles e importantes como indicadores biológicos de la calidad del agua (Holzenthal et al., 2007; Houghton et al., 2011).
El conocimiento de los tricópteros en México aún es incompleto debido a la escasez de especialistas, por lo que hasta ahora se han registrado 488 especies, de las cuales 156 se distribuyen en el estado de Oaxaca y representan 31.9% de la diversidad del país (Razo-González, 2018). Los estudios de tricópteros mexicanos son básicamente revisiones taxonómicas, descripciones de especies nuevas y estudios ecológicos, donde solo 3 trabajos incorporan alguna metodología biogeográfica para analizar su distribución. Bueno-Soria y Barba-Álvarez (1992) realizaron la primera aproximación a las afinidades biogeográficas de algunos tricópteros de México, mediante una aportación al Atlas Nacional de Naturaleza de México. En dicho estudio, determinaron la afinidad de algunas familias y géneros de tricópteros con base en su distribución geográfica. En otro estudio, Bueno-Soria et al. (2005) catalogaron la diversidad de tricópteros del arroyo Las Flores en Tabasco, e indagaron sus afinidades biogeográficas. Posteriormente, Bueno-Soria et al. (2007) analizaron la distribución de los tricópteros de la Sierra Tarahumara para inferir sus posibles afinidades biogeográficas, mediante un análisis de parsimonia de endemismos, y determinaron que la fauna de dicha Sierra presenta afinidad primordialmente Neotropical.
La Sierra de Juárez es un macizo montañoso localizado en la zona centro-norte del estado de Oaxaca, que forma parte de la región terrestre prioritaria de las sierras del norte de Oaxaca-Mixe (Arriaga et al., 2000), que se caracteriza por una gran diversidad de ambientes y cuya importancia para la conservación se debe, principalmente, a su variedad de ecosistemas, la presencia de endemismos y su riqueza específica, entre otros aspectos. La vegetación predominante en la zona es el bosque de pino, pero también se presentan bosques mixtos, selva alta perennifolia y bosque mesófilo de montaña (Arriaga et al., 2000). Predomina el clima templado, aunque a escala local se presenta un mosaico de climas, que van desde semicálidos húmedos y subhúmedos, a semifríos y templados húmedos en las partes más altas (Trejo, 2004). La precipitación media anual oscila entre 800 y 2,000 mm y la temperatura media anual varía entre 12 y 18 °C.
La Sierra de Juárez forma parte de la Zona de Transición Mexicana, especialmente interesante por presentar una alta mezcla de elementos de afinidad neártica y neotropical (Bueno-Soria y Barba-Álvarez, 1992). De acuerdo con Morrone y Márquez (2008), en esta región convergen 3 provincias biogeográficas, por lo que se catalogó de manera preliminar como un nodo panbiogeográfico. Escalante et al. (2004, 2018), mediante el análisis de los patrones de distribución de taxones de mamíferos, identificaron varios nodos en los que se incluye el área de la Sierra de Juárez. En otro estudio, León-Paniagua y Morrone (2009) analizaron varios taxones de vertebrados mediante biogeografía cladística, encontrando que en las tierras altas de Oaxaca, en las que se incluye la Sierra de Juárez, convergen 2 componentes bióticos, y se sugiere la realización de más estudios en el área para comprender la diversificación biótica de esta zona. De acuerdo con la regionalización más reciente de México, las provincias que convergen en la Sierra de Juárez son la Cuenca del Balsas, en la vertiente occidental de la sierra, la Sierra Madre del Sur y la Veracruzana (Morrone, 2019). Morrone (2017), en su regionalización biogeográfica de la provincia de la Sierra Madre del Sur, identificó la existencia de 3 subprovincias: 1) la subprovincia Occidental a su vez se divide en los distritos Jaliscience y Jaliscience-Manantlán, 2) la subprovincia Central que comprende el distrito Michoacano, y 3) la subprovincia Oriental, la cual se divide en los distritos Guerrerense y Altiplanicie Oaxaqueña; en este último distrito, que corresponde a las Sierras Mixteca y Norte, se incluye la Sierra de Juárez. En este estudio dicha sierra no se cataloga como un nodo; sin embargo, el distrito en donde se ubica es el que concentra el mayor número de especies endémicas de toda la provincia (35 especies; Morrone, 2017).
Rosen (1988) propuso el análisis de parsimonia de endemismos como herramienta para la identificación de áreas de endemismo. De acuerdo con Morrone (1994), el análisis de parsimonia de endemismos (PAE, por sus siglas en inglés) identifica áreas de endemismo (análogas a taxones) con base en conjuntos de taxones compartidos (análogos a caracteres), de acuerdo con el cladograma más parsimonioso. El análisis parte de una matriz de datos de presencia/ausencia de un conjunto de localidades y de un grupo de taxones. Algunos autores han utilizado este método para evaluar las relaciones entre las provincias biogeográficas de México como áreas de endemismo predeterminadas (Morrone y Márquez, 2001; Morrone et al., 1999). El PAE con eliminación progresiva de caracteres (PAE-PCE por sus siglas en inglés), es una modificación propuesta por Luna et al. (2000) y García-Barros et al. (2002). El PAE-PCE es un proceso iterativo, en el que cada vez que se lleva a cabo un análisis de parsimonia y se obtiene un cladograma, las especies que sustentan los clados (sinapomorfías) se eliminan de la matriz para realizar un nuevo análisis. El procedimiento se repite hasta que no aparecen sinapomorfías en los cladogramas. El PAE-PCE ha sido utilizado como una herramienta para facilitar la identificación de trazos generalizados, pues cada nuevo análisis permite obtener clados alternativos que representan nuevos trazos generalizados (Luna et al., 2000). Debido a que el PAE puede considerarse como un método de anidamiento de trazos generalizados en un esquema jerárquico (Craw et al., 1999; Smith, 1992), éste ha sido aplicado en algunos estudios en México con el propósito de establecer áreas prioritarias de conservación y analizar los límites de la Zona de Transición Mexicana (Luna et al., 2000; Morrone y Márquez, 2001).
En este trabajo se examinan los patrones de distribución de los tricópteros de la Sierra de Juárez mediante un PAE-PCE para identificar trazos generalizados y evidenciar la complejidad biogeográfica de la zona. Con ello, se pretende comprender por qué es un área altamente biodiversa desde el punto de vista histórico, así como contar con argumentos adicionales a los ya documentados para considerarla como área natural protegida.
Materiales y métodos
La Sierra de Juárez es un complejo montañoso ubicado en el distrito Ixtlán de Juárez, que forma parte de la Sierra Madre de Oaxaca. Limita al norte por el río Santo Domingo, al oeste por el río Grande, al este por el río Cajonos y en el extremo sur converge con la Sierra Monteflor. Los municipios que pertenecen a esta sierra son: Ixtlán de Juárez, Natividad, San Juan Atepec, San Juan Evangelista Analco, San Juan Quiotepec, Capulálpam de Méndez, San Miguel Yotao, San Pablo Macuiltianguis, San Pedro Yaneri, San Pedro Yólox, Santa Ana Yareni, Santa Catarina Lachatao, Santa María Yavesía, Santiago Comaltepec, Santiago Laxopa y Santiago Xiacuí, los cuales abarcan una superficie de 1,700 km2 aproximadamente. La vegetación predominante en la Sierra de Juárez, son los bosques templados entre los que se pueden mencionar los bosques de pinos, de encinos y mixtos. La selva alta perennifolia también tiene una representatividad importante, seguida del bosque mesófilo de montaña, el bosque caducifolio, y en menor proporción el bosque de galería y pastizales (Arriaga et al., 2000; Torres-Colín, 2004).
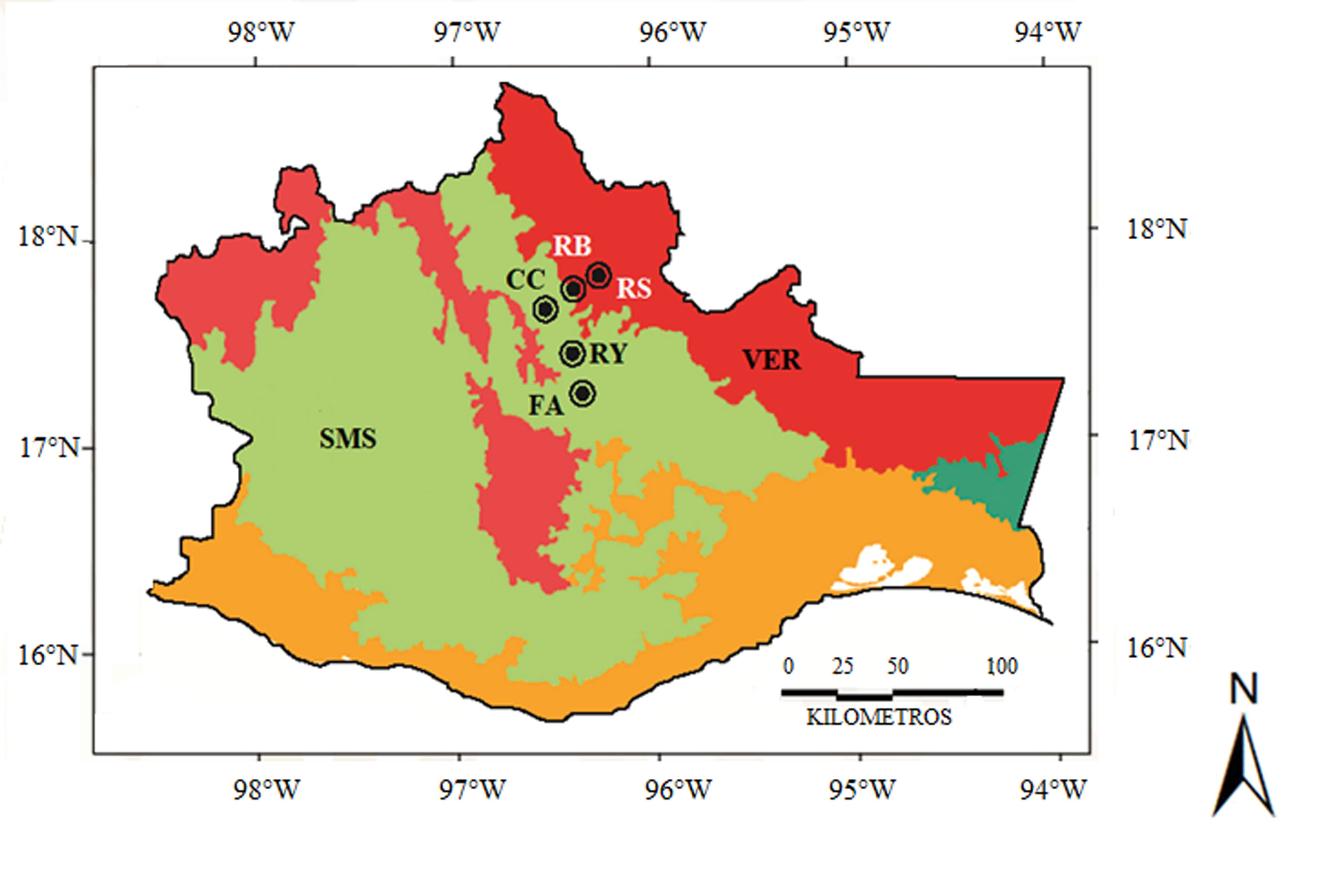
El estudio se basó en 120 especies de tricópteros colectadas en la Sierra de Juárez, Oaxaca. Las colectas se realizaron en 5 ríos distribuidos a lo largo de 1 gradiente de altitud que abarca aproximadamente 2,300 m (fig. 1, tabla 1). Debido a que los adultos de Trichoptera son de hábitos crepusculares-nocturnos y, además, son atraídos por la luz, la recolección se realizó mediante trampas de luz negra (Blahnik y Holzenthal, 2004). Las trampas de luz se colocaron en la ribera de cada 1 de los 5 ríos y estuvieron encendidas durante 4 horas a partir del crepúsculo. Los insectos recolectados se colocaron en frascos con alcohol al 96% para su preservación y posterior identificación. De manera complementaria, se usó una red entomológica en el día, durante una hora, con el propósito de capturar activamente a los tricópteros de hábitos diurnos como los de la familia Xiphocentronidae (Schmid, 1982). Para identificar las especies, las estructuras genitales de los machos adultos se procesaron de acuerdo a la técnica descrita por Blahnik y Holzenthal (2004). El material colectado fue depositado en la Colección Nacional de Insectos del Instituto de Biología de la UNAM.
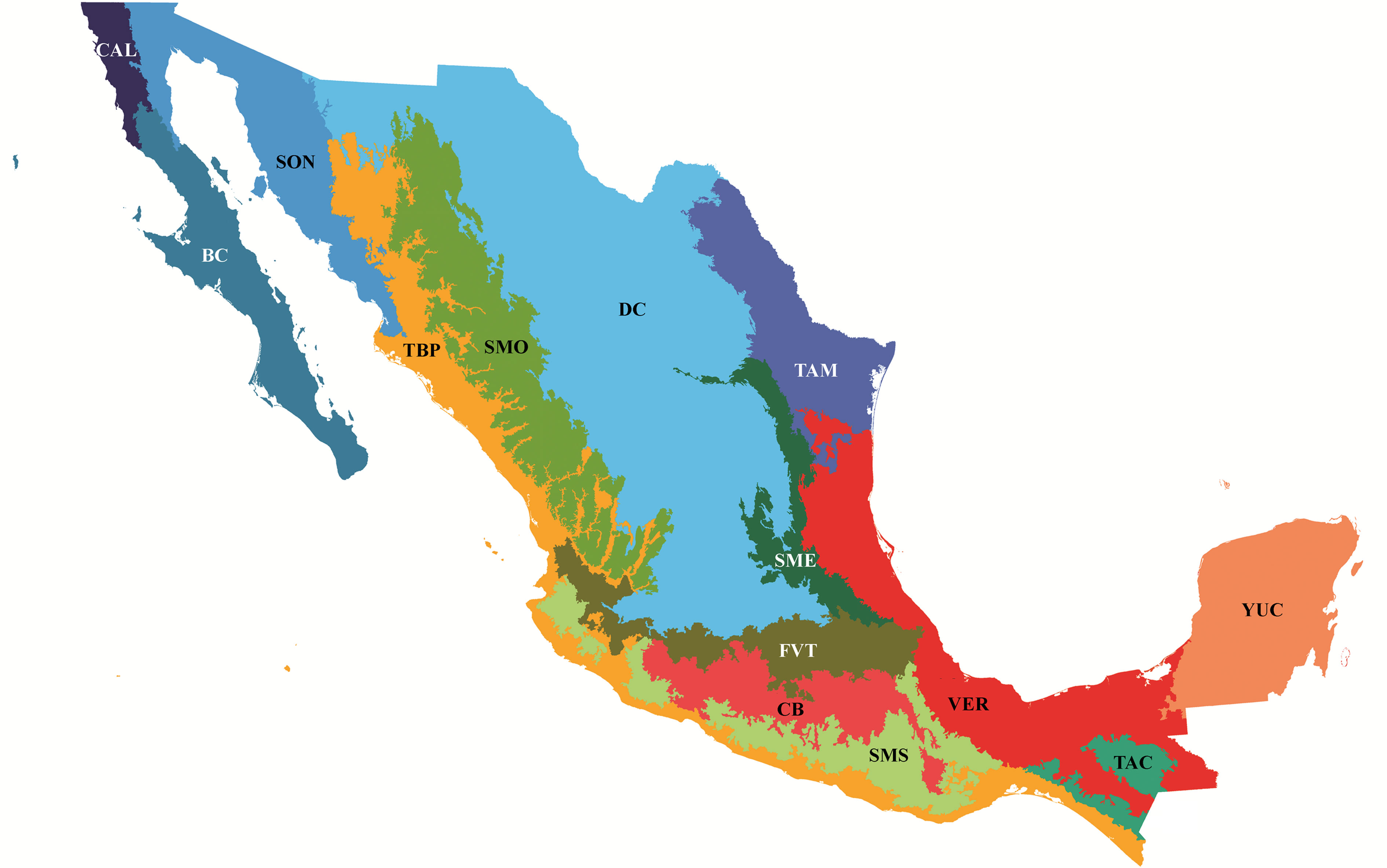
Para el análisis biogeográfico se tomaron en cuenta 946 registros de distribución geográfica, solo en el territorio mexicano, de las 120 especies de tricópteros reportadas para la Sierra de Juárez, incluyendo los registrados en campo y los obtenidos mediante la revisión exhaustiva de la literatura disponible (Banks, 1900, 1901; Barba-Álvarez y Bueno-Soria, 2005; Barba-Álvarez et al., 2002; Blahnik, 1998; Blahnik y Holzenthal, 2006, 2008, 2014; Bueno-Soria, 1981, 1983, 1984, 1985, 1986, 1990, 1999, 2002, 2009, 2010; Bueno-Soria y Barba-Álvarez, 2011, 2015; Bueno-Soria y Hamilton, 1986; Bueno-Soria y Holzenthal, 1986, 2004; Bueno-Soria y Santiago-Fragoso, 1996; Bueno-Soria et al., 2001, 2005; Denning, 1941, 1952, 1965, 1968; Denning y Blickle, 1979; Denning y Sykora, 1966; Flint, 1965, 1967, 1970, 1971, 1972, 1974, 1981, 1985, 1991, 1998; Flint y Bueno-Soria, 1977; Flint et al., 1987; Hagen, 1861; Harris y Contreras-Ramos, 1989; Holzenthal, 1988, 1995; Holzenthal y Andersen, 2004; Holzenthal y Harris, 1992; Johanson, 2002; Mosely, 1934, 1937, 1954; Moulton y Harris, 1997; Muñoz-Quesada y Holzenthal, 2015; Oláh y Johanson, 2011, 2012; Razo-González, 2018; Razo-González et al., 2020; Rojas-Ascencio et al. 2002; Ross, 1947, 1948, 1951, 1956, 1959; Ross y King, 1952; Schmid, 1982, 1989; Thompson y Holzenthal, 2015; Weaver, 1988). Dichos registros fueron asignados manualmente a las provincias biogeográficas propuestas por Morrone et al. (2017; fig. 2).
Se construyó una matriz básica de datos (tabla 2) donde las unidades de estudio fueron las provincias biogeográficas de México, codificando la presencia (1) o ausencia (0) de las especies (columnas) en cada una de las provincias (filas). Las provincias Californiana, Baja California, Sonora, Desierto Chihuahuense y Península de Yucatán se excluyeron del análisis por no presentar alguna especie o contar con 1 solo registro. Así, el número final de áreas de endemismo fue de 10, incluyendo un área hipotética codificada con ceros para enraizar el cladograma (Morrone, 1994). Se eliminaron las especies Phylloicus aeneus (Hagen, 1861), Mortoniella meralda (Mosely, 1954), Helicopsyche borealis (Hagen, 1861) y Chimarra mexicana (Banks, 1900) por presentar una distribución amplia en el país, en al menos 7 de las 10 provincias.
El análisis se llevó a cabo mediante una búsqueda heurística (múltiple TBR+TBR) en WinClada (Nixon, 2002) y NONA (Goloboff, 1999) con 1,000,000 como el número máximo de árboles retenidos por cada iteración del análisis, 100,000 réplicas y 100 árboles iniciales por réplica. Se obtuvo el árbol de consenso estricto de los árboles más parsimoniosos. Una vez obtenido este árbol, se implementó el análisis de parsimonia de endemismos con eliminación progresiva de caracteres (PAE-PCE) de acuerdo con Luna et al. (2000) y García-Barros et al. (2002), donde cada vez que se obtuvo un conjunto de cladogramas, se elaboró un árbol de consenso estricto, se eliminaron las sinapomorfías geográficas (especies) que definieron los trazos generalizados y se analizó nuevamente la matriz. Este proceso se realizó hasta no obtener sinapomorfías en los árboles generados. Se obtuvieron valores de soporte para cada análisis mediante pruebas de bootstrap con 2000 réplicas. Posteriormente, se analizaron los cladogramas y se utilizaron los siguientes criterios para la definición de los clados que se transformaron en trazos generalizados: 1) que los clados contaran con al menos 2 sinapomorfías geográficas, 2) cuando se presentó más de un posible trazo generalizado dentro de una rama interna del cladograma, se seleccionó aquel que presentó el mayor valor de bootstrap, y 3) cuando se presentaron 2 o más clados con 2 o más sinapomorfías en distintas ramas del cladograma, éstos se consideraron como trazos generalizados, independientemente de sus valores de bootstrap. Los trazos generalizados se dibujaron uniendo las provincias incluidas en el mismo clado por su distancia geográfica mínima. Dichos trazos son parciales debido a que en el análisis no se incluyeron las localidades fuera de México. Se revisó la distribución geográfica de las especies diagnósticas de cada trazo generalizado y cuando la distribución de al menos 2 de ellas se extiende hacia el sur del país, esta situación se hace notar con una flecha que apunta hacia esa dirección. Solo se describió el nodo ubicado en la Sierra de Juárez con base en la intersección o cruce de los 8 trazos generalizados, considerando que para definir otros nodos es necesaria la inclusión de más información, puesto que aquí no restringimos al análisis de las especies de la Sierra de Juárez.
Tabla 1
Sitios de estudio en la Sierra de Juárez, Oaxaca, México.
| Río | Altitud (m snm) | Vegetación | Clima | Provincia biogeográfica |
| Soyolapam | 136 | Selva alta perennifolia | Aw | Veracruzana |
| Bobo | 460 | Selva alta perennifolia | Af | Veracruzana |
| Yegu-Iag-Lagu | 1,320 | Bosque mesófilo | (A)c(m) | Sierra Madre del Sur |
| Flor de Azahar | 1,967 | Bosque encino-pino | Cw | Sierra Madre del Sur |
| Cascadas Comaltepec | 2,432 | Bosque pino | Cw | Sierra Madre del Sur |
Tabla 2
Matriz básica de datos de las especies de tricópteros de la Sierra de Juárez y su ubicación en provincias biogeográficas (X = raíz). Las columnas y filas han sido transpuestas por presentación. Abreviaturas de las provincias biogeográficas en el pie de la figura 2. Las especies están organizadas por familias y las familias se ordenan alfabéticamente.
| Provincias biogeográficas | ||||||||||
| Especies | X | TAM | SMO | SME | FVT | CB | SMS | TBP | VER | TAC |
| Calamocaratidae | ||||||||||
| Phylloicus gomezi | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Glossosomatidae | ||||||||||
| Culoptila jamapa | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 |
| Culoptila pararusia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Mortoniella falcicula | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Mortoniella mexicana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Protoptila bicornuta | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Protoptila ixtala | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 |
| Protoptila liqua | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Protoptila resolda | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Protoptila rota | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Protoptila spangleri | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Helicopsychidae | ||||||||||
| Cochliopyche vazquezae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Helicopsyche dampfi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| Helicopsyche planata | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 |
| Helicopsyche tuxtlensis | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Helicopsyche villegasi | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Hydrobiosidae | ||||||||||
| Atopsyche calopta | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| Atopsyche dampfi | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| Atopsyche erigia | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 1 |
| Atopsyche huenga | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 |
| Atopsyche japoda | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Atopsyche majada | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| Atopsyche pilcomayo | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Hydropsychidae | ||||||||||
| Cheumatopsyche gelita | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Diplectrona chiapensis | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 |
| Hydropsyche toschiae | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 |
| Leptonema albovirens | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Leptonema championi | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 |
| Leptonema crassum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| Leptonema pinotepa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| Leptonema s. mayanum | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 |
| Tabla 2. Continúa | ||||||||||
| Provincias biogeográficas | ||||||||||
| Especies | X | TAM | SMO | SME | FVT | CB | SMS | TBP | VER | TAC |
| Plectropsyche hoogstraali | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 0 |
| Smicridea bulara | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Smicridea dampfi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Smicridea dispar | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 |
| Smicridea dithyra | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 1 |
| Smicridea lacanha | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Smicridea radula | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 |
| Smicridea signata | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Smicridea soyatepecana | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Hydroptilidae | ||||||||||
| Anchitrichia spangleri | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| Byrsopteryx tabasquensis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Costatrichia lodora | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Hydroptila arctia | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 |
| Hydroptila denza | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 |
| Hydroptila mexicana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Hydroptila misolha | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Ithytrichia mexicana | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Leucotrichia imitator | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 |
| Leucotrichia melleopicta | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Mayatrichia rualda | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Metrichia circulatrix | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Metrichia crenula | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 |
| Metrichia yavesia | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Neotrichia exicoma | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Neotrichia xicana | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 |
| Neotrichia tuxtla | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Ochrotrichia buenoi | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Ochrotrichia ildria | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Ochrtorichia ixtlahuaca | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Ochrotrichia nicaragua | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Ochrotrichia pacifica | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| Ochrotrichia stylata | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Ochrotrichia yavesia | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Ochrotrichia yetla | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Oxyethira azteca | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 |
| Oxyethira desadora | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Oxyethira tica | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Tabla 2. Continúa | ||||||||||
| Provincias biogeográficas | ||||||||||
| Especies | X | TAM | SMO | SME | FVT | CB | SMS | TBP | VER | TAC |
| Rhyacopsyche mexicana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Zumatrichia filosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Zumatrichia multisetosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Leidostomatidae | ||||||||||
| Lepidostoma aztecum | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Lepidostoma bakeri | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 |
| Lepidostoma catarina | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Lepidostoma frontale | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 0 |
| Lepidostoma zapoteca | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Leptoceridae | ||||||||||
| Nectopsyche dorsalis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| Nectopsyche gemmoides | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 1 |
| Nectopsyche pavida | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Oecetis disjuncta | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| Oecetis metlacensis | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 |
| Oecetis silviae | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 |
| Oecetis sordida | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Triaenodes flintorum | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Triaenodes oaxacensis | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Odontoceridae | ||||||||||
| Marilia flexuosa | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Philopotamidae | ||||||||||
| Chimarra acuta | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 |
| Chimarra barrettae | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| Chimarra bicolor | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 |
| Chimarra butleri | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Chimarra dentosa | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 |
| Chimarra cornuta | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Chimarra laguna | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 |
| Chimarra oaxaca | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Chimarra ortiziana | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Chimarra pelaezi | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 |
| Chimarra ridleyi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Chimarra setosa | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Chimarra texana | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 |
| Wormaldia barbai | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 |
| Wormaldia noveloi | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Wormaldia palma | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Tabla 2. Continúa | ||||||||||
| Provincias biogeográficas | ||||||||||
| Especies | X | TAM | SMO | SME | FVT | CB | SMS | TBP | VER | TAC |
| Wormaldia planae | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 |
| Wormaldia tarasca | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 |
| Polycentropodidae | ||||||||||
| Cernotina calcea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Cernotina chiapaneca | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Limnephilus tulatus | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Polycentropus aliciae | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 |
| Polycentropus ariensis | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 |
| Polycentropus aztecus | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 |
| Polycentropus casicus | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| Polycentropus mexicanus | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 |
| Polycentropus mayanus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Polycentropus veracruzensis | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 |
| Polyplectropus carolae | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Polyplectropus charlesi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Xiphocentronidae | ||||||||||
| Cnodocentron trilineatum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Xiphocentron aureum | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 |
| Xiphocentron serestus | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 |
Resultados
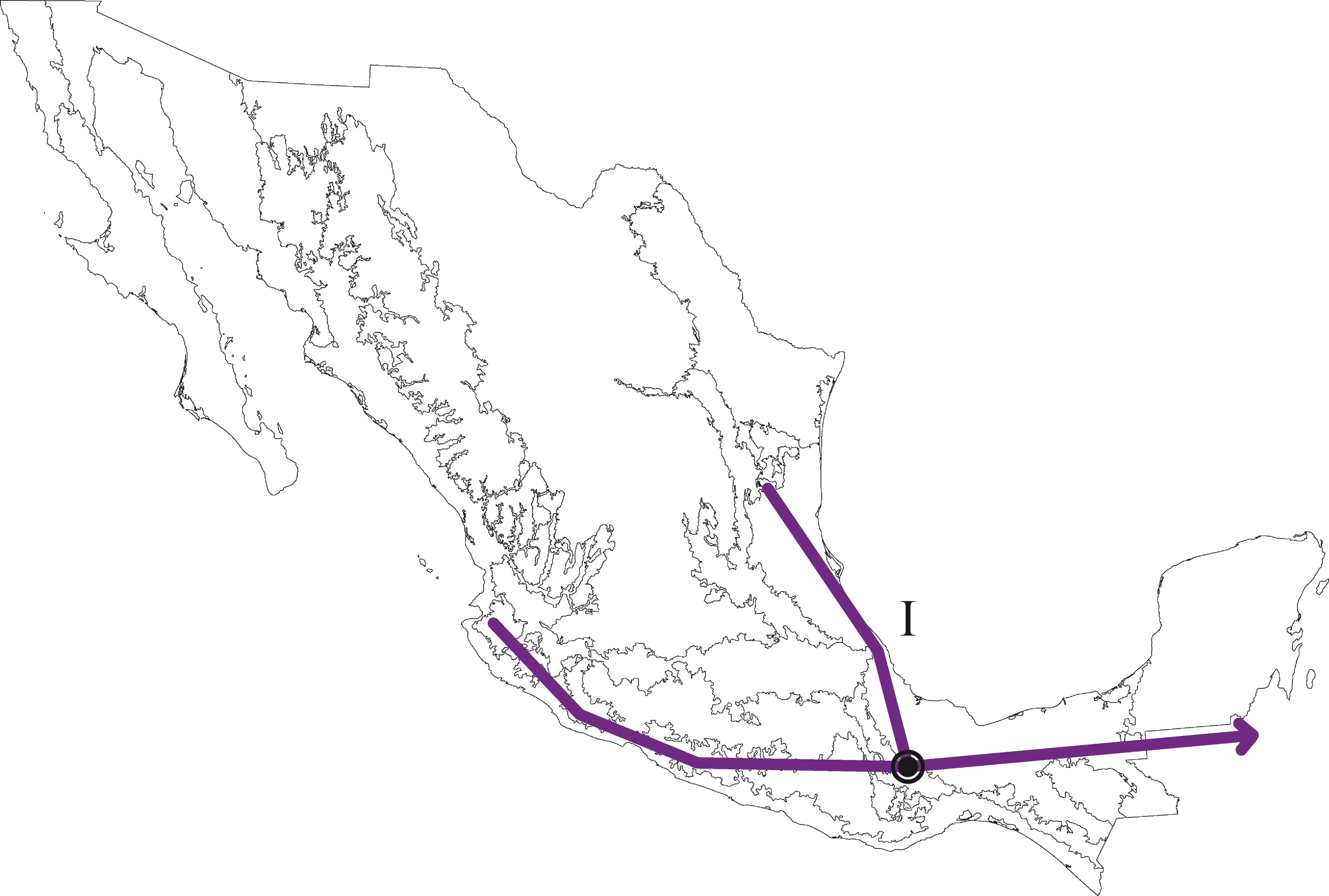
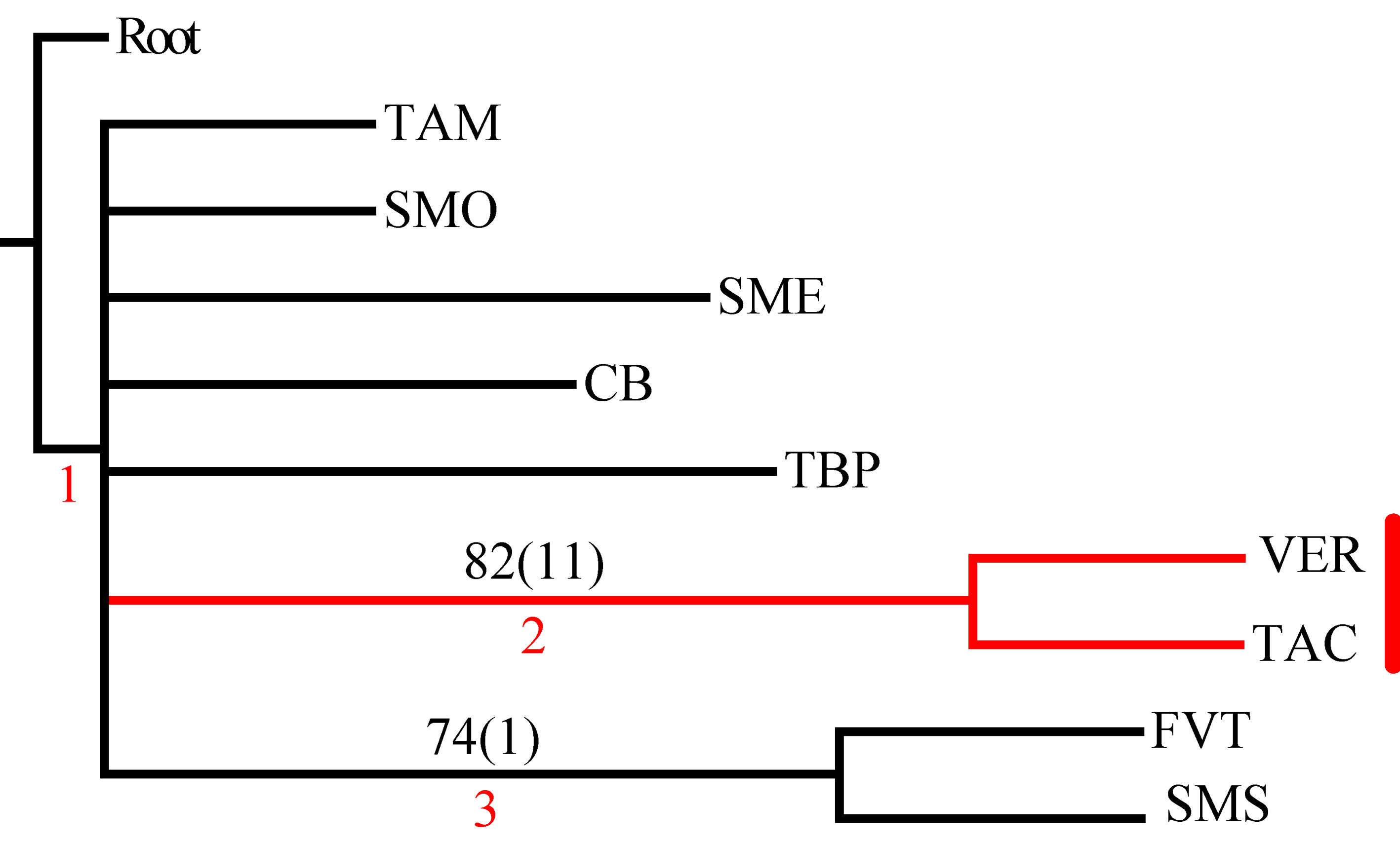
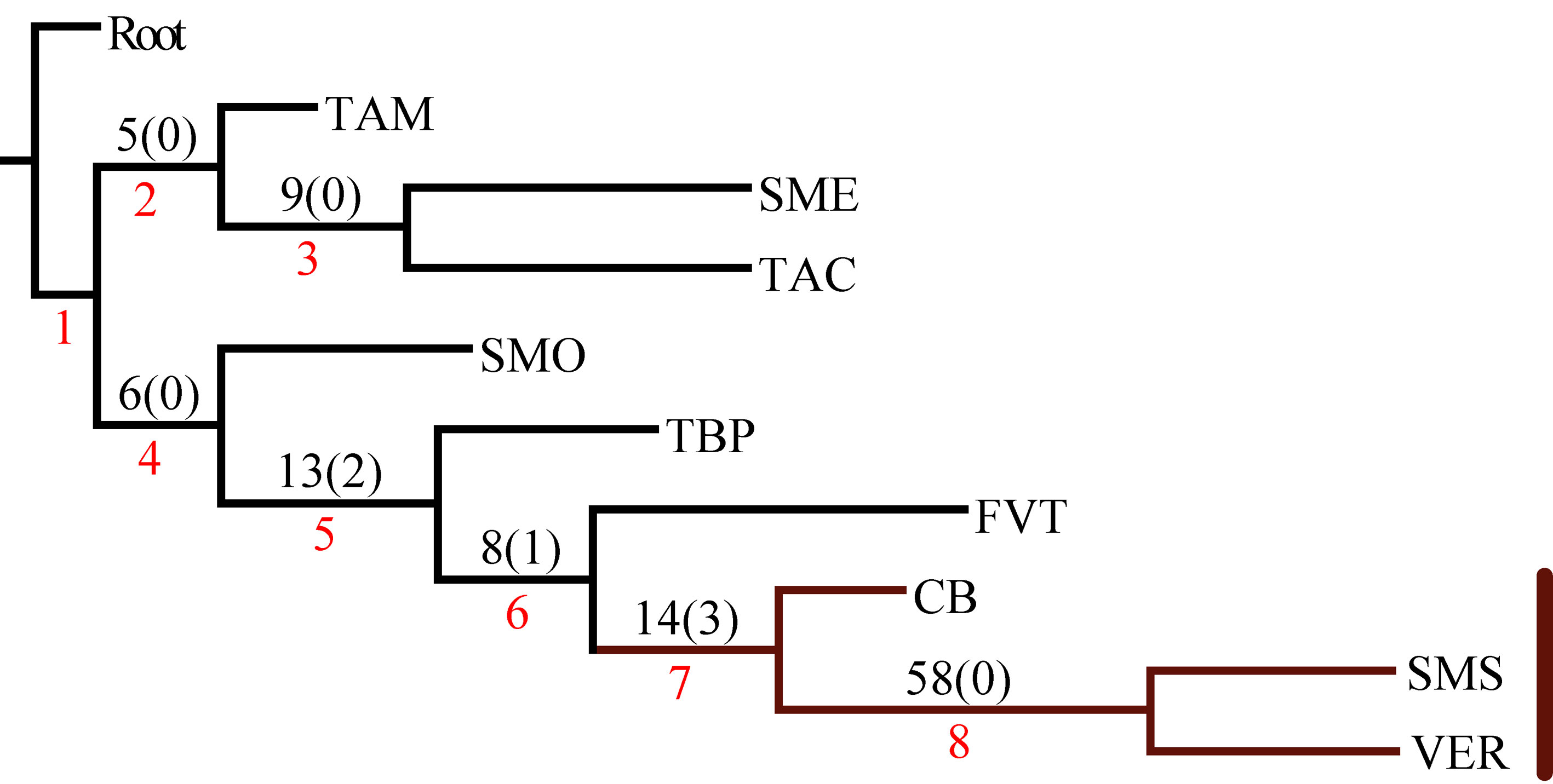
El análisis de la matriz original generó 2 árboles igualmente parsimoniosos con una longitud de 206 pasos, índice de consistencia de 0.57 e índice de retención de 0.44. El árbol de consenso estricto fue de 209 pasos, índice de consistencia de 0.56 e índice de retención de 0.42 (fig. 3). Los 2 cladogramas igualmente parsimoniosos fueron muy similares, y solo difirieron en la posición de las provincias de la Faja Volcánica Transmexicana y de las Tierras Altas de Chiapas, respecto del clado que contiene a las provincias de la Sierra Madre del Sur y Veracruzana. En el primer cladograma, la Faja Volcánica Transmexicana es el grupo hermano de dicho clado, soportado por las especies Hydropsyche toschiae (Denning, 1965) y Xiphocentron serestus Schmid, 1982, mientras que en el segundo cladograma las Tierras Altas de Chiapas ocupan esta posición, pero sin sinapomorfías. El cladograma de consenso estricto presenta 2 componentes principales, el primero incluye a las provincias de la Sierra Madre Occidental, la Cuenca del Balsas y las Tierras Bajas del Pacífico; el segundo componente incluye a la provincia de Tamaulipas como grupo hermano de la mayoría de las provincias montañosas junto con la Veracruzana. En este último, la provincia de la Sierra Madre Oriental aparece como grupo hermano de un clado con relaciones no resueltas, que incluye a la Faja Volcánica Transmexicana, las Tierras Altas de Chiapas y al clado de las provincias de la Sierra Madre del Sur y la Veracruzana, soportado por 11 especies (tabla 3). El clado que contiene a las provincias de la Sierra Madre del Sur y Veracruzana se seleccionó como el trazo generalizado I (fig. 4).
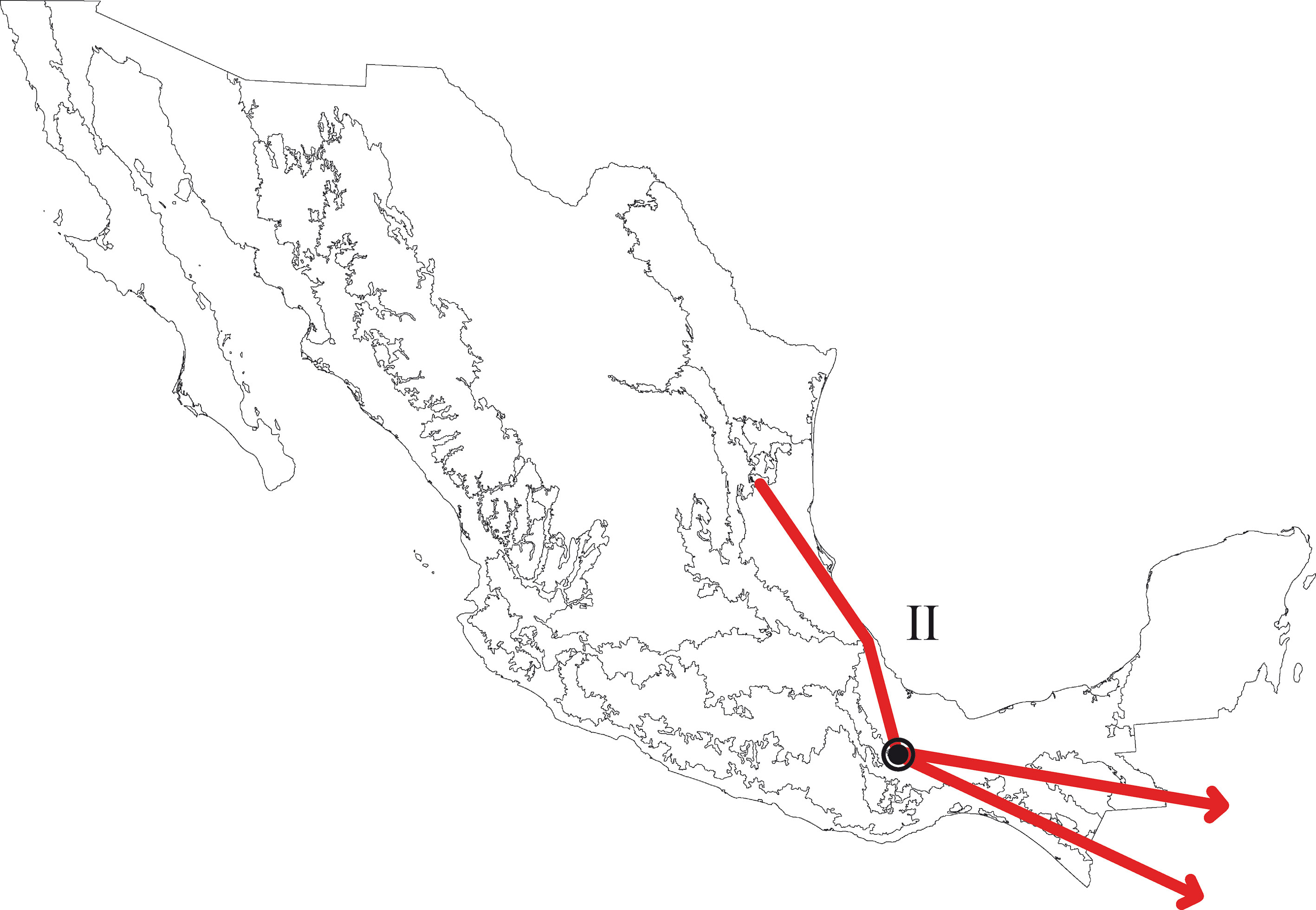
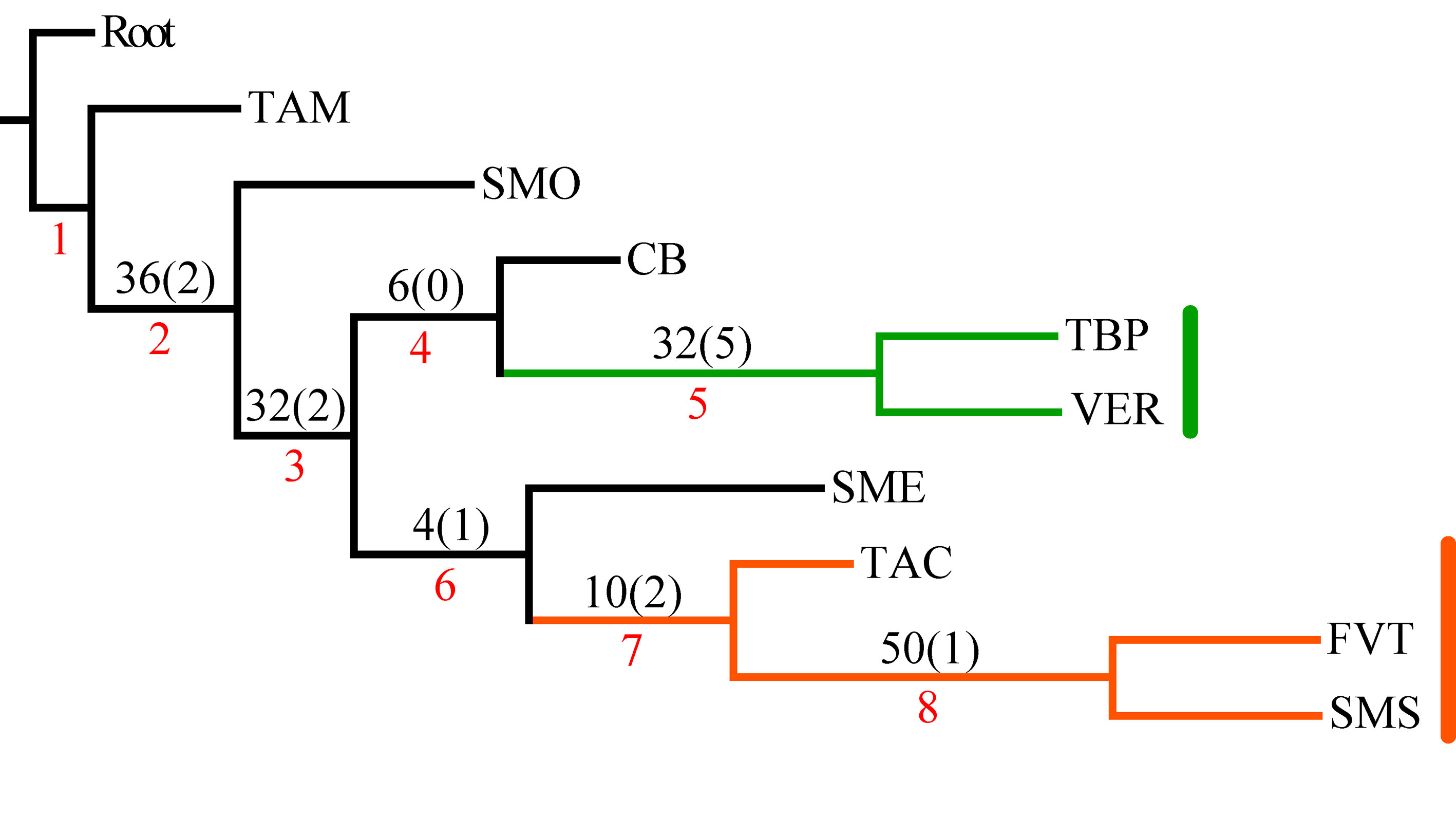
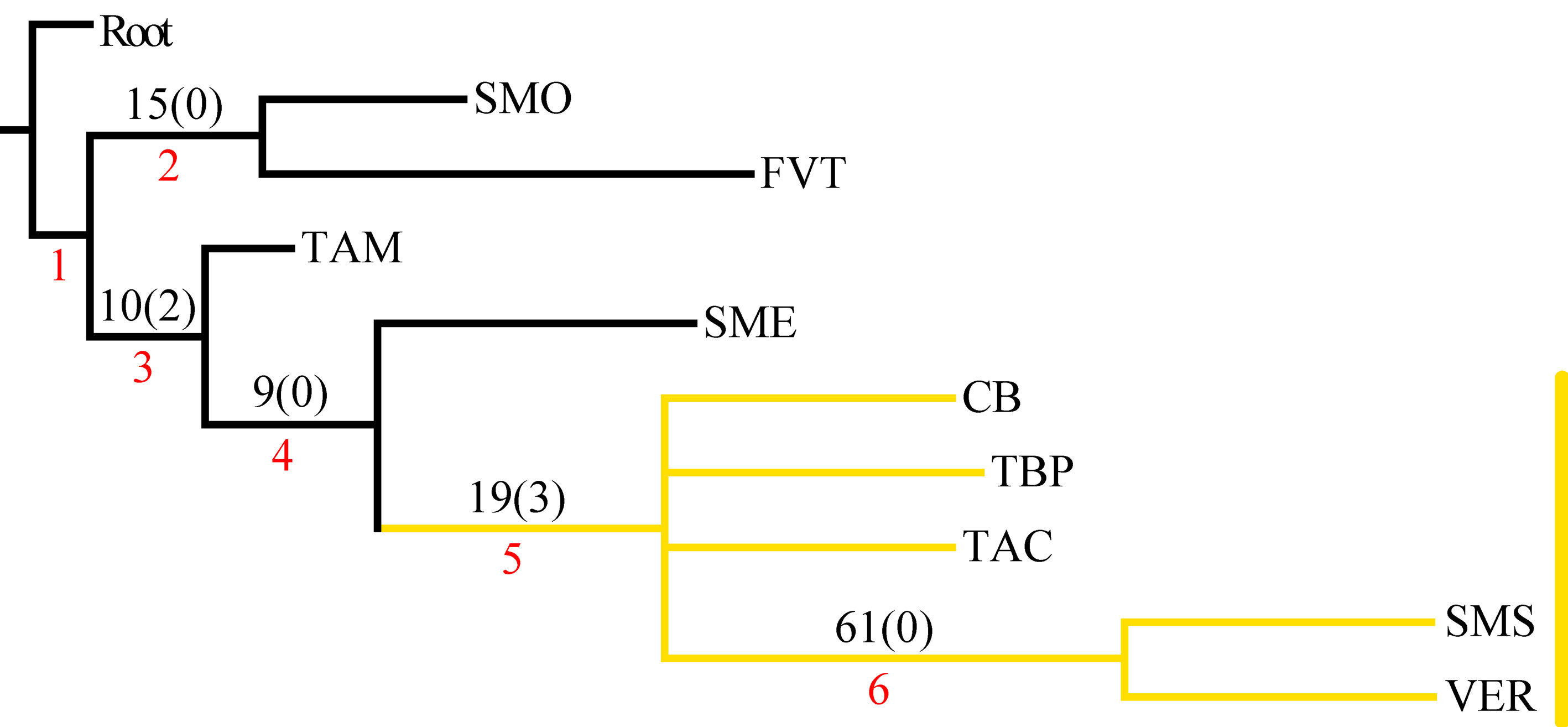
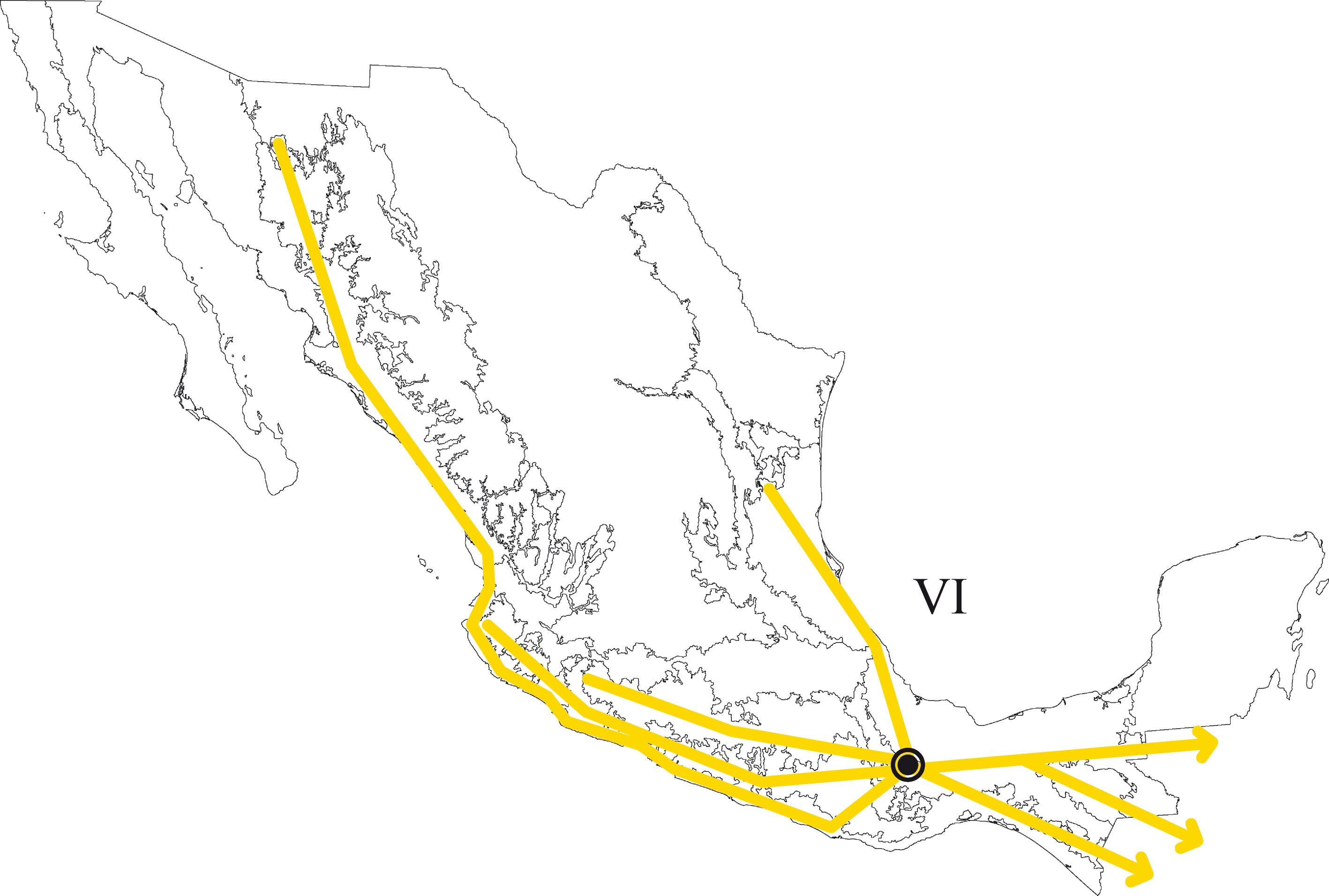
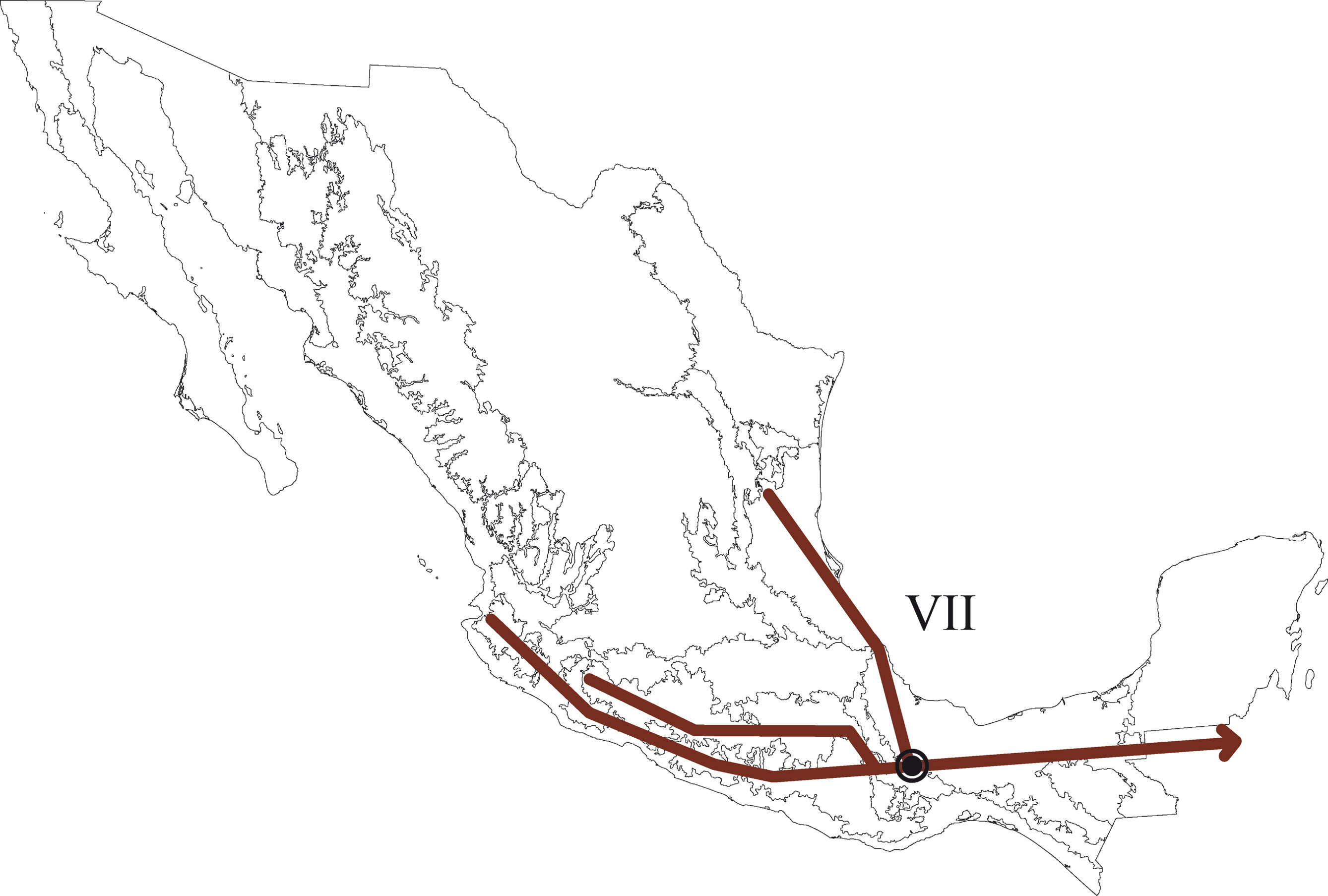
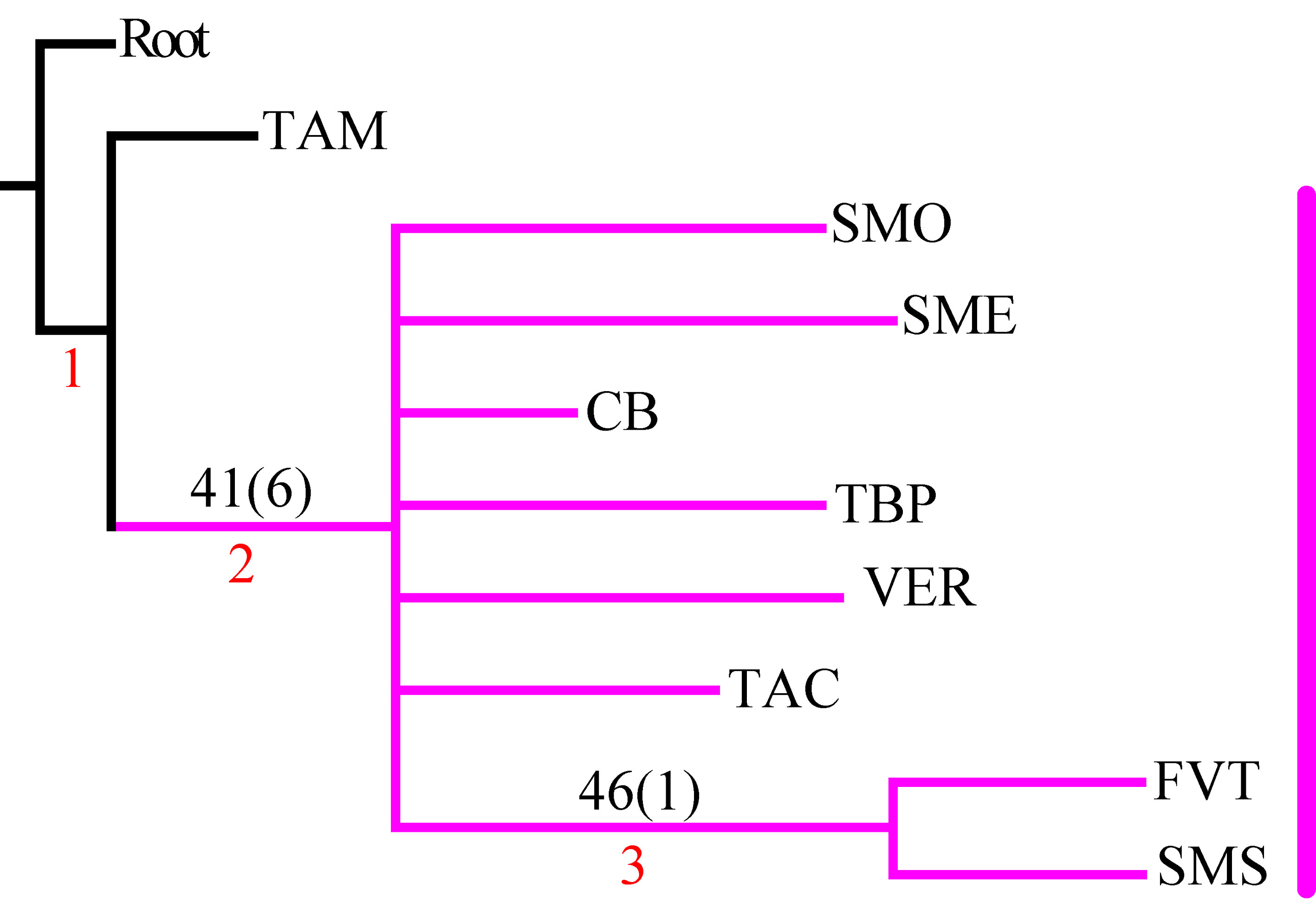
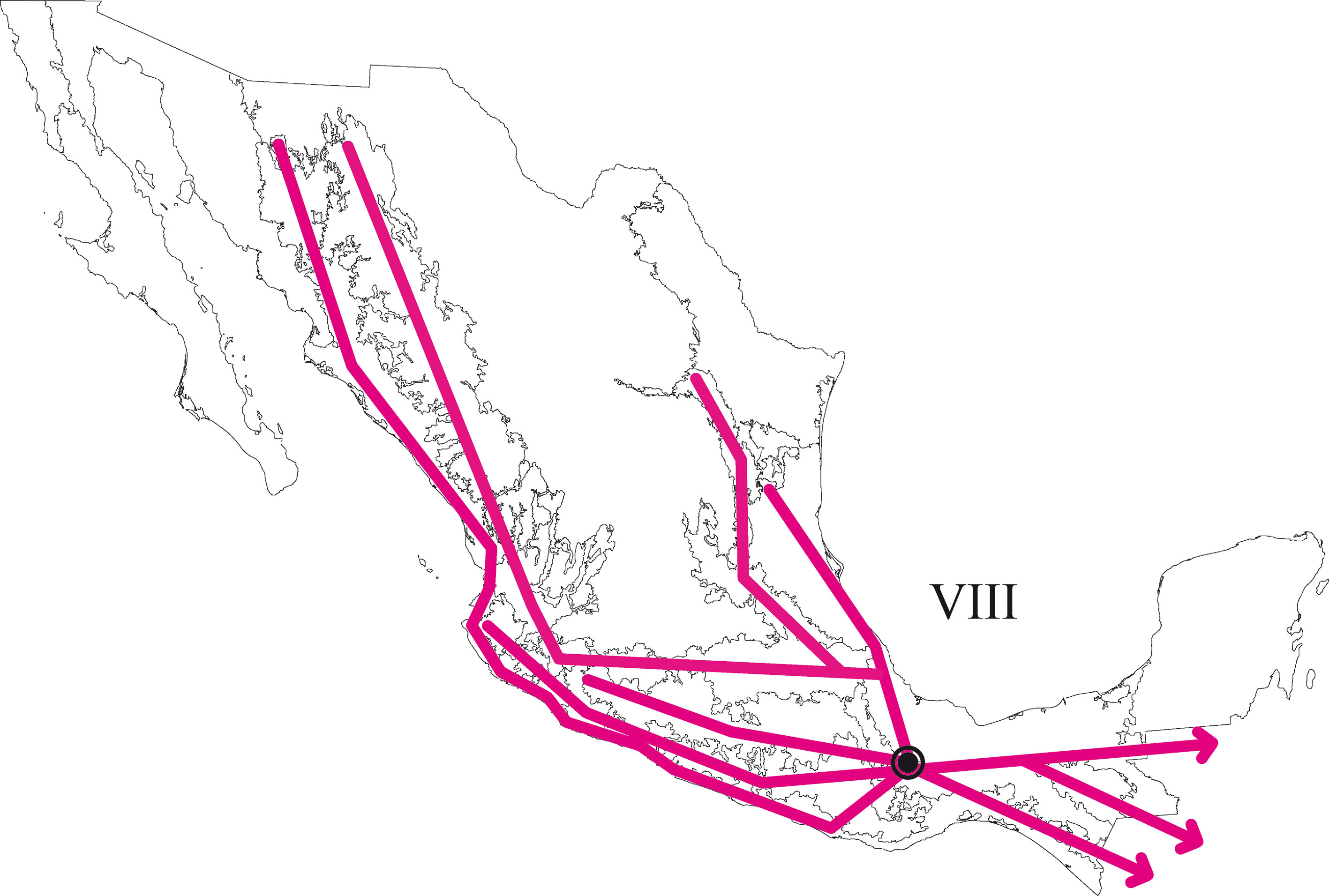
En el PAE-PCE, para el segundo análisis se eliminaron 11 especies de la matriz que resultaron sinapomorfías para el trazo generalizado I del primer análisis. Se obtuvieron 5 cladogramas igualmente parsimoniosos (L = 187, CI = 0.57, RI = 0.45) y en el cladograma de consenso estricto (L = 209, CI = 0.51, RI = 0.30; fig. 5), el clado que contiene a las provincias Veracruzana y Tierras Altas de Chiapas se representó como el trazo generalizado II (fig. 6). Del tercer análisis, se eliminaron 11 especies, se produjo un solo cladograma (L =172, CI = 0.56, RI = 0.44; fig. 7) donde el clado que contiene a las provincias Tierras Bajas del Pacífico y Veracruzana, y el clado que contiene a las provincias Tierras Altas de Chiapas, Faja Volcánica Transmexicana y Sierra Madre del Sur, se transformaron en los trazos generalizados III y IV (fig. 8). El cuarto análisis se realizó eliminando 7 especies, originó 4 cladogramas igualmente parsimoniosos (L = 160, CI = 0.56, RI = 0.44), en cuyo cladograma de consenso (L = 178, CI = 0.50, RI = 0.30; fig. 9), el clado mejor sustentado con 4 especies definió el trazo generalizado V que contiene a las provincias Sierra Madre Oriental, Faja Volcánica Transmexicana, Tierras Altas de Chiapas, Sierra Madre del Sur y Veracruzana (fig. 10). En un quinto análisis se eliminaron 4 especies, obteniendo así 3 árboles igualmente parsimoniosos (L = 149, CI = 0.57, RI = 0.44), en el cladograma de consenso estricto (L = 155, CI = 0.55, RI = 0.38; fig. 11), el clado que contiene a las provincias Cuenca del Balsas, Tierras Bajas del Pacífico, Tierras Altas de Chiapas, Sierra Madre del Sur y Veracruzana se transformó en el trazo generalizado VI (fig. 12). Para el sexto análisis se eliminaron 3 especies y se produjo un solo cladograma (L = 142, CI = 0.58, RI = 0.42) (fig. 13), donde el clado que abarca a las provincias Cuenca del Balsas, Sierra Madre del Sur y Veracruzana se representó como el trazo generalizado VII (fig. 14). El séptimo análisis se realizó después de eliminar 3 especies, se generaron 4 árboles igualmente parsimoniosos (L = 136, CI = 0.58, RI = 0.42) y se construyó el cladograma de consenso estricto (L = 156, CI = 0.51, RI = 0.21; fig. 15) en el que el clado que contiene a las provincias Sierra Madre Occidental, Sierra Madre Oriental, Cuenca del Balsas, Tierras Bajas del Pacífico, Veracruzana, Tierras Altas de Chiapas, Faja Volcánica Transmexicana y Sierra Madre del Sur, se interpretó como el trazo generalizado VIII (fig. 16).
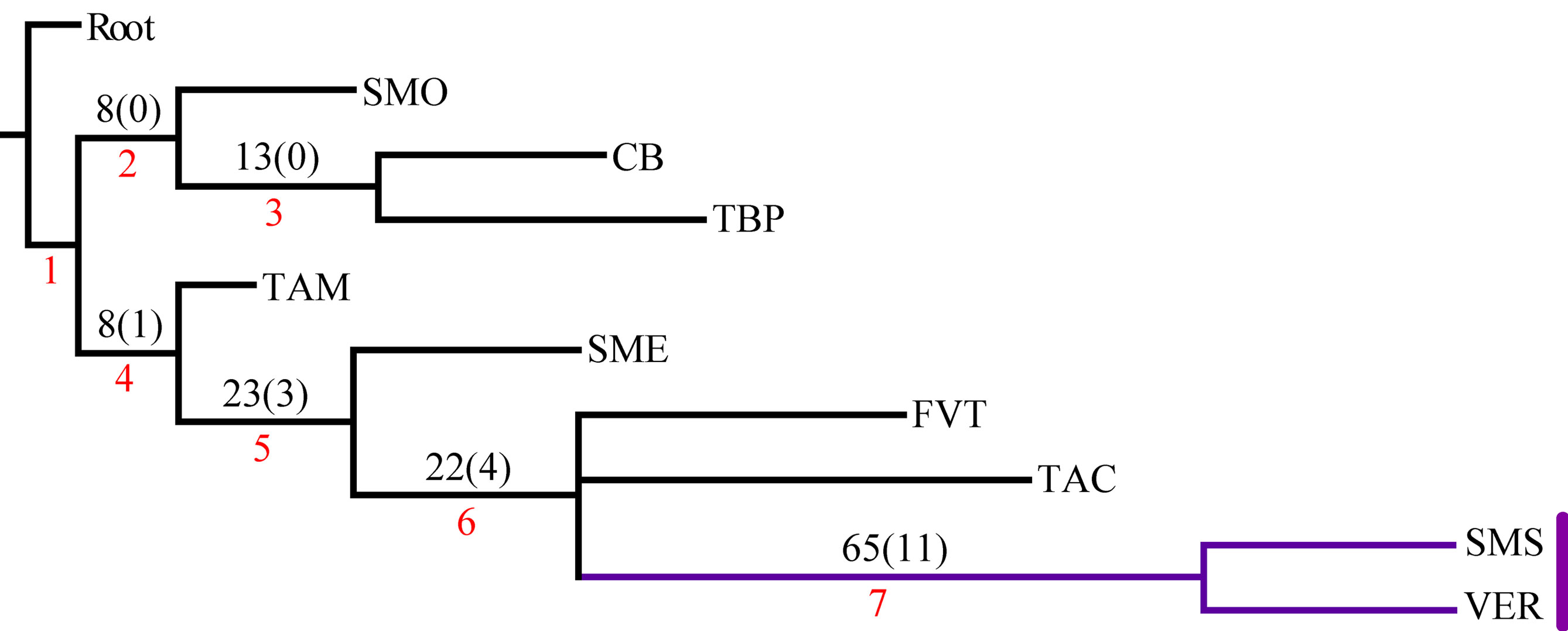
Se llevó a cabo un octavo análisis eliminando 6 especies, se produjeron 4 árboles igualmente parsimoniosos (L = 136, CI = 0.58, RI = 0.42) y se construyó un cladograma de consenso estricto (L = 120, CI = 0.61, RI = 0.40). Sin embargo, las relaciones mostradas fueron soportadas únicamente por homoplasias o por una sola sinapomorfía, de manera que ya no se reconocieron trazos generalizados en este último análisis. Las especies de tricópteros que definieron cada trazo generalizado se listan en la tabla 3.
Tabla 3
Trazos generalizados de Trichoptera de la Sierra de Juárez, Oaxaca, organizados por regiones y provincias, con las especies que los definen. Las abreviaturas de las provincias se especifican en la figura 2.
| Trazos generalizados | Regiones | Provincias | Especies |
| I | Zona de Transición Mexicana y Neotropical | SMS
VER |
Helicopsyche tuxtlensis, Atopsyche japoda, Smicridea soyatepecana, Ithytrichia mexicana, Mayatrichia rualda, Metrichia circulatrix, Neotrichia tuxtla, Ochrotrichia stylata, Triaenodes flintorum, Marilia flexuosa, Chimarra setosa |
| II | Neotropical | VER
TAC |
Protoptila liqua, P. resolda, P. rota, Cochliopsyche vazquezae, Smicridea lacanha, Costatrichia lodora, Hydroptila mexicana, H. misolha, Neotrichia exicoma, Zumatrichia filosa, Polycentropus mayanus |
| III | Zona de Transición Mexicana y Neotropical | TBP
VER |
Helicopsyche damfi, Leptonema pinotepa, Anchitrichia spangleri, Ochrotrichia pacifica, Nectopsyche dorsalis |
| IV | Zona de Transición Mexicana y Neotropical | TAC
VER SMS |
Lepidostoma bakeri, Polycentropus mexicanus |
| V | Zona de Transición Mexicana y Neotropical | SME
FVT TAC SMS VER |
Atopsyche damfi, Oxyethira azteca, Lepidostoma frontale, Chimarra barrettae |
| VI | Zona de Transición Mexicana y Neotropical | CB
TBP TAC VER SMS |
Protoptila ixtala, Leptonema simulans mayanum, Chimarra dentosa |
| VII | Zona de Transición Mexicana y Neotropical | CB
VER SMS |
Hydroptila arctia, Metrichia crenula, Wormaldia barbai |
| VIII | Zona de Transición Mexicana y Neotropical | SMO
SME CB TBP VER TAC FVT SMS |
Atopsyche calopta, A. majada, Plectropsyche hoogstraali, Chimarra acuta, C. bicolor, C. laguna |
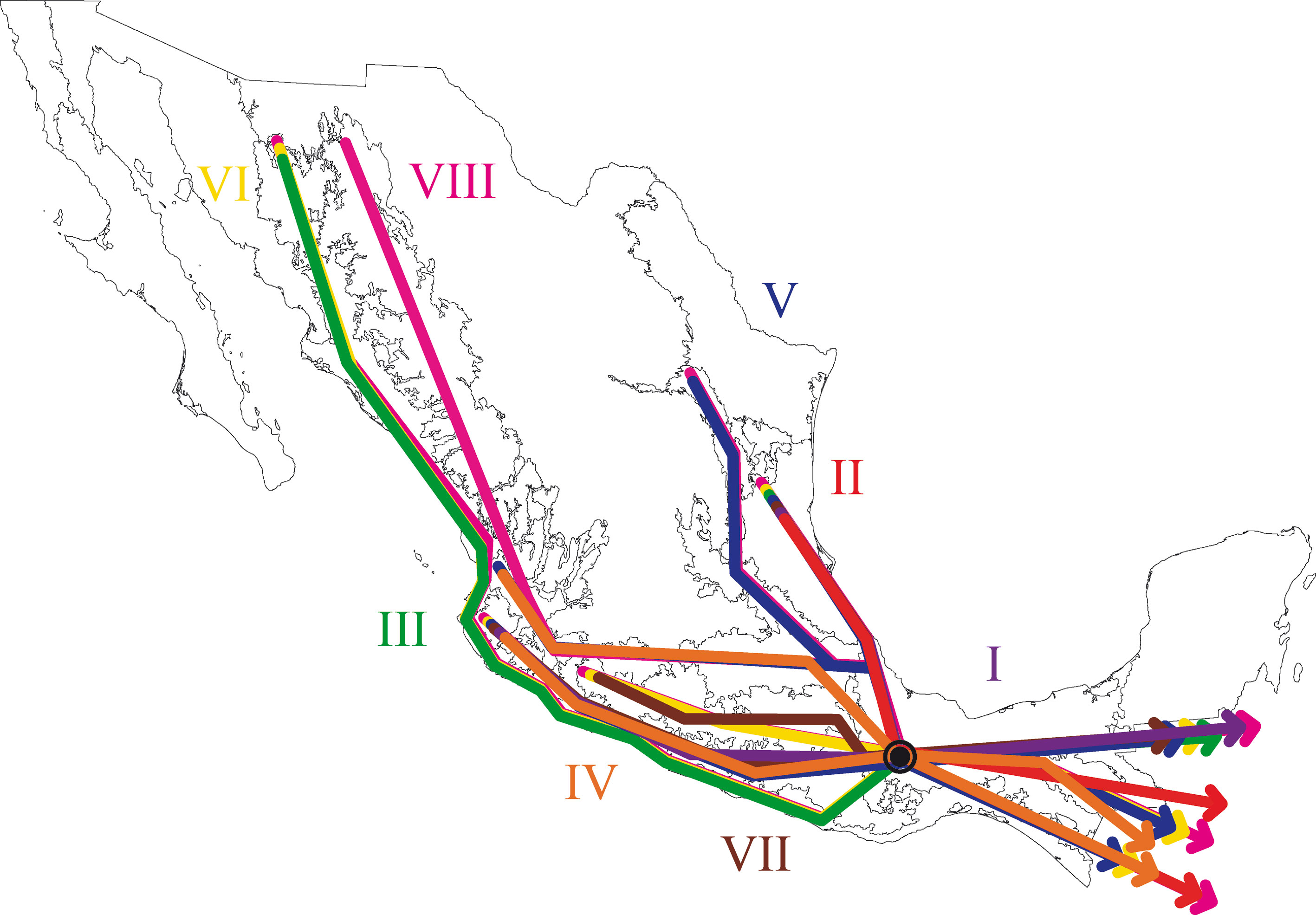
En este estudio se definió la Sierra de Juárez como un nodo panbiogeográfico por la superposición manual de los 8 trazos generalizados identificados (fig. 17).
De las 120 especies de tricópteros registradas, 47 presentan registros solo en el territorio mexicano, es decir, 73 extienden su distribución más allá de las fronteras geopolíticas, ya sea hacia el norte o hacia el sur. Las especies endémicas de la provincia de la Sierra Madre del Sur son: Phyllocus gomezi (Razo-González, 2018), Mortoniella falcicula (Blahnik y Holzenthal, 2008), Atopsyche pilcomayo (Schmid, 1989), Metrichia yavesia (Bueno-Soria, 2002), Ochrotrichia buenoi (Razo-González, 2018), O. yavesia (Bueno-Soria y Holzenthal, 2004), Lepidostoma catarina (Bueno-Soria, Santiago-Fragoso y Barba-Álvarez, 2001), L. zapoteca (Razo-González, 2018), Triaenodes oaxacensis (Holzenthal y Andersen, 2004), Chimarra oaxaca (Blahnik, 1998) y Wormaldia noveloi (Razo-González, 2018). Las especies endémicas de la provincia Veracruzana son Culoptila pararusia (Blahnik y Holzenthal, 2006), Mortoniella mexicana (Blahnik y Holzenthal, 2008), Protoptila spangleri (Flint, 1967), Smicridea dampfi (Flint, 1974), Byrsopteryx tabasquensis (Bueno-Soria, Santiago-Fragoso y Barba-Álvarez, 2001), Ochrotrichia yetla (Bueno-Soria, 2009), Chimarra cornuta (Ross, 1959), Cernotina chiapaneca (Bueno-Soria, 2010), Polyplectropus carolae (Bueno-Soria, 1990) y Cnodocentron trilineatum (Mosely, 1934). Las especies endémicas de la Sierra de Juárez son Phylloicus gomezi (Razo-González, 2018), Mortoniella falcicula (Blahnik y Holzenthal, 2008), Atopsyche pilcomayo (Schmid, 1989), Metrichia yavesia (Bueno-Soria, 2002), Ochrotrichia buenoi (Razo-González, 2018), O. yavesia (Bueno-Soria y Holzenthal, 2004), O. yetla (Bueno-Soria, 2009), Lepidostoma catarina (Bueno-Soria, Santiago-Fragoso y Barba-Álvarez, 2001), L. zapoteca (Razo-González, 2018), Chimarra oaxaca (Blahnik, 1998) y Wormaldia noveloi (Razo-González, 2018).



Discusión
Las 120 especies registradas en la Sierra de Juárez representan 24.6% de la riqueza nacional registrada, en una porción equivalente a 0.9% de la superficie del país. Los trazos generalizados y el nodo identificado en este trabajo fortalecen la hipótesis que la Sierra de Juárez es un área muy compleja biogeográficamente hablando, al ubicarse en una zona de transición entre varias provincias, dado que en estas zonas ocurren eventos de hibridación biótica causados por cambios ecológicos e históricos que permiten la combinación de diferentes componentes bióticos (Morrone, 2004a).
Cuando se comparan nuestros resultados con estudios previos que han examinado las relaciones entre las provincias biogeográficas mexicanas, algunas similitudes son evidentes. Morrone et al. (1999) y Contreras-Medina et al. (2007), con base en datos de distribución de aves, insectos y plantas, detectaron la segregación en 2 grupos principales, uno incluye a las provincias del norte del país y otro contiene a las provincias del centro y el sur de México. En este estudio tal segregación es evidente solo para el caso de la provincia de Tamaulipas, debido a que la mayoría de las provincias del norte del país cuentan con muy pocos o ningún registro de distribución de tricópteros. Respecto al segundo grupo, la posición de las provincias es variable, pero la relación entre la Sierra Madre del Sur y la Veracruzana se mantiene, como también se ha reportado en otros estudios (Corona y Morrone, 2005; Corona et al., 2007; García-Marmolejo et al., 2008; Morrone y Escalante, 2002; Morrone y Gutiérrez, 2005). De las especies de tricópteros que fungieron como sinapomorfías geográficas, 3 se distribuyen más allá de la frontera con los EUA, 16 son endémicas de México, y la gran mayoría presenta una distribución sureña, que en algunos casos se extiende hacia Centro y Sudamérica.
El análisis de la distribución geográfica de los tricópteros registrados en la Sierra de Juárez permitió identificar 8 trazos generalizados que ponen de manifiesto la naturaleza compleja de la región por la mezcla de especies que pertenecen a distintos componentes bióticos. En el trazo generalizado I se relacionan las provincias Veracruzana y de la Sierra Madre del Sur, patrón que se ha reconocido recurrentemente mediante el análisis de diversos taxones en otros estudios (Contreras-Medina et al., 2007; Corona y Morrone, 2005; Corona et al., 2007; García-Marmolejo et al., 2008; Morrone y Escalante, 2002; Morrone y Gutiérrez, 2005; Toledo et al., 2007). La relación entre la provincia de las Tierras Altas de Chiapas y la Veracruzana, observada en el trazo generalizado II, ha sido evidenciada en trabajos que incluyen el análisis de varios taxones (Echeverry y Morrone, 2013; Morrone et al., 1999), coleópteros (Morrone y Márquez, 2001; Toledo et al., 2007), mamíferos (Escalante et al., 2005) y plantas (Contreras-Medina et al., 2007; Cué-Bär et al., 2006). Este trazo está sustentado por una mezcla de especies endémicas y de especies que se distribuyen en México y Centroamérica, mostrando un patrón de distribución que, aparentemente, corresponde con el componente biótico Mesoamericano de Morrone y Márquez (2003). El trazo generalizado III, que relaciona a las provincias Tierras Bajas del Pacífico y Veracruzana, coincide con el trazo Sudamericano de Contreras-Medina y Eliosa-León (2001) mediante el análisis de taxones tanto animales como vegetales; también es semejante a los trazos 14 y 15 de Corona y Morrone (2005, coleópteros), al trazo V de García-Marmolejo et al. (2008, mamíferos), y al trazo II de Maya-Martínez et al. (2011, mariposas), los cuales también pertenecen al componente Mesoamericano (Morrone y Márquez, 2003). Las relaciones entre las provincias Tierras Altas de Chiapas, Faja Volcánica Transmexicana y Sierra Madre del Sur observadas en el trazo IV coinciden con el trazo “d” de Contreras-Medina et al. (2007); sin embargo, dicho trazo no incluye a las Tierras Altas de Chiapas. El trazo generalizado V, que relaciona a las provincias Sierra Madre Oriental, Faja Volcánica Transmexicana, Tierras Altas de Chiapas, Veracruzana y Sierra Madre del Sur, a pesar de presentar relaciones no resueltas, concuerda con el trazo generalizado 12 de Álvarez-Mondragón y Morrone (2004, aves); sin embargo, nuestro trazo incluye a las Tierras Altas de Chiapas en lugar de las Tierras Bajas del Pacífico. El trazo V también coincide con uno de los clados encontrados por Escalante et al. (2005) mediante el análisis de mamíferos y se ajusta al trazo 1 de Corona et al. (2007, bupréstidos), excepto que nuestro trazo también incluye a la Faja Volcánica Transmexicana. El trazo generalizado VI coincide con el trazo IV de García-Marmolejo et al. (2008) y con uno de los trazos encontrados por Echeverry y Morrone (2010), exceptuando que nuestro trazo no incluye a la provincia Faja Volcánica Transmexicana. El trazo generalizado VII, que asocia a la Cuenca del Balsas como grupo hermano del clado y que contiene a las provincias Sierra Madre del Sur y Veracruzana, representa un patrón que no se había registrado como tal en otros estudios. El trazo generalizado VIII es una muestra clara de la compleja historia biogeográfica de México, y particularmente de la Sierra de Juárez, al integrar a las provincias Sierra Madre Occidental, Sierra Madre Oriental, Cuenca del Balsas, Tierras Bajas del Pacífico, Veracruzana, Tierras Altas de Chiapas, Faja Volcánica Transmexicana y Sierra Madre del Sur. Dicho trazo coincide con trazo el IV de García-Marmolejo et al. (2008), excepto que nuestro trazo no incluye a la provincia de Yucatán.
En estudios precedentes, la Sierra Madre Oriental, que solo aparece en los trazos generalizados V y VIII, se relaciona frecuentemente con las provincias del norte del país y con la Faja Volcánica Transmexicana (Escalante et al., 2005; Morrone y Escalante, 2002; Morrone y Gutiérrez, 2005; Morrone y Márquez, 2001; Morrone et al., 1999). Las relaciones reflejadas en los resultados obtenidos aquí pueden estar influenciadas por la naturaleza de la Sierra Madre Oriental que mediante el análisis de plantas (Luna et al., 1999), hongos (Cifuentes et al., 2004), coleópteros (Márquez y Morrone, 2003) y helechos (Sanginés-Franco et al., 2011) se ha definido como un área no natural.
Todos los trazos identificados involucran provincias de transición y neotropicales que se corresponden con los componentes Mesoamericano y Mexicano de Montaña (Morrone y Márquez, 2003). El componente Mesoamericano incluye a las provincias de las Tierras Bajas del Pacífico, las Tierras Altas de Chiapas y la Veracruzana; el componente Mexicano de Montaña comprende las provincias de la Sierra Madre Occidental, la Sierra Madre Oriental, la Faja Volcánica Transmexicana, la Cuenca del Balsas y la Sierra Madre del Sur, que conforman la Zona de Transición Mexicana, y que se caracterizan por albergar una alta diversidad resultado de los procesos de especiación que se llevaron a cabo a partir de la hibridación de las regiones Neártica y Neotropical. Cabe señalar que en nuestro estudio solo se presentaron registros de distribución de tricópteros en la subprovincia Oriental de la Sierra Madre del Sur y que las especies endémicas de esta provincia pertenecen al distrito Altiplanicie Oaxaqueña, lo cual coincide con la riqueza alta y los numerosos endemismos reportados por Morrone (2017) para esta zona.
Los nodos son áreas complejas en donde 2 o más trazos generalizados se superponen o convergen, ellos se interpretan como zonas de convergencia tectónica y/o biótica (Morrone, 2004b). El nodo obtenido para los tricópteros en la Sierra de Juárez concuerda con los patrones de distribución observados en otros estudios. Coincide con el nodo Sierra de Juárez de Márquez y Morrone (2003, coleópteros estafilínidos), el nodo Oaxaca norte de Álvarez-Mondragón y Morrone (2004, aves), el nodo Sierra Madre de Oaxaca de Escalante et al. (2004, 2018, mamíferos), el nodo C de Corona y Morrone (2005, coleópteros bupréstidos), los nodos 2 y 3 de Márquez y Asiain (2006, estafilínidos) y el nodo 5 de González et al. (2017, hongos), poniendo en evidencia la mezcla de elementos pertenecientes a diferentes taxones y componentes bióticos, no solo en la dimensión horizontal (latitudinal) del espacio, sino también en la vertical (altitudinal) como se pudo observar en nuestro estudio.
Los trazos generalizados identificados en este trabajo mediante el análisis de los patrones de distribución de los tricópteros de la Sierra de Juárez han sido reconocidos previamente como áreas de endemismo aplicando el PAE con un enfoque panbiogeográfico, para diferentes taxones (Contreras-Medina et al., 2007; Echeverry y Morrone, 2010; Escalante et al., 2005; Toledo et al., 2007). Los resultados aquí reportados integran patrones encontrados en múltiples trabajos, con una amplia variedad de taxones; sin embargo, también se reconocieron nuevos patrones no reportados con anterioridad. Todo ello es evidencia de la enorme complejidad biogeográfica de la Sierra de Juárez que, a pesar de ser un área pequeña, en comparación con otras áreas, reviste una gran importancia tanto en términos ecológicos como evolutivos. Comparando los trazos generalizados obtenidos se observa que 4 de ellos relacionan a la provincia Veracruzana con la provincia de la Sierra Madre del Sur, esta relación se muestra especialmente estrecha en el trazo generalizado I, el cual está sustentado por la distribución congruente de 11 especies. En el caso del trazo generalizado II la provincia Veracruzana se relaciona cercanamente con las Tierras Altas de Chiapas. Lo anterior indica la gran importancia que la provincia Veracruzana tiene para la distribución de los tricópteros en México, ya que dicha provincia recorre buena parte del país en sentido norte-sur y entra en contacto con varias provincias biogeográficas en su límite occidental. Nuestro análisis fortalece la premisa de que las montañas de la Zona de Transición Mexicana forman parte de un área compleja que exhibe una riqueza alta y eventos de especiación que se han confirmado en diversos estudios con diferentes taxones tanto animales como vegetales. Asimismo, la identificación de un nodo en la Sierra de Juárez en donde se superponen y convergen múltiples trazos generalizados, no solo de tricópteros sino de otros muchos taxones, demuestra la evolución activa que ocurre en esta región y la importancia que las montañas de Oaxaca y Guerrero tienen para los eventos de vicarianza que han ocurrido en la fauna de linajes mesoamericanos (Halffter, 2003).
Para discernir los patrones de distribución (sensu Halffter, 2003) de los tricópteros, queda un largo camino por recorrer, ya que hasta el momento no hay una hipótesis estable de las relaciones filogenéticas entre los subórdenes y dentro de algunas superfamilias. Existen al menos 2 filogenias, que son las mejor soportadas, pero que discrepan significativamente en cuanto a la conformación de los subórdenes de Trichoptera (ver Malm et al., 2013; Thomas et al., 2020), lo cual puede ser una fuente importante de error al momento de estudiar la biogeografía histórica de los tricópteros. Otro factor a considerar es el vacío de información que existe en muchas áreas geográficas de México, por ejemplo, para las provincias Californiana y de Yucatán, donde no hay registros, debido probablemente al esfuerzo de muestreo desigual que se ha realizado hasta ahora, más que por la ausencia real de tricópteros en esas zonas, todo ello aunado a la carencia de especialistas y de investigaciones que aborden de manera sistemática la diversidad de tricópteros en el país.
En términos de la conservación, la Sierra de Juárez se incluye en la Región Terrestre Prioritaria para la Conservación Sierras del norte de Oaxaca-Mixe, la cual ostenta un alto valor para la conservación por los servicios ambientales que provee (Arriaga et al., 2000) y forma parte del Área de Importancia para la Conservación de las Aves Sierra Norte de Oaxaca. También cabe destacar el esfuerzo de varias comunidades indígenas en la creación de áreas destinadas voluntariamente a la conservación (ADVC), las cuales son áreas naturales de carácter federal certificadas por la Comisión Nacional de Áreas Naturales Protegidas y que son manejadas por las mismas comunidades. En la Sierra de Juárez se encuentran 14 ADVC, que abarcan 26.5% de su superficie y que en su mayoría se encuentran adyacentes, permitiendo la interconexión de un mosaico de diferentes tipos de vegetación (Conanp, 2020). En este contexto, consideramos que el nodo localizado en la Sierra de Juárez es particularmente importante para la conservación, debido a que presenta una alta riqueza de especies en diferentes grupos biológicos que pertenecen a diferentes componentes bióticos, y es necesaria la creación de áreas naturales protegidas de mayor calibre que las ADVC, como una Reserva de la Biosfera que abarquen esta región.
Agradecimientos
La primera autora agradece a José Antonio Gómez-Anaya (INECOL A.C.) por su apoyo durante el trabajo de campo; a José Arturo Casasola González (Universidad de la Sierra Juárez) por las facilidades para el trabajo de gabinete en el Instituto de Estudios Ambientales; a Joaquín Bueno-Soria y Rafael Barba Álvarez (Universidad Nacional Autónoma de México) por su asistencia para la identificación de Trichoptera. A las autoridades de Santiago Comaltepec, San Martín Soyolapam, San Juan Tepanzacoalco, La Esperanza, Santiago Cuasimulco, Santiago Laxopa y Santa Cartarina Lachatao por proporcionar acceso a sus tierras. Agradecemos los valiosos comentarios de dos revisores anónimos. Al Posgrado en Ciencias Biológicas de la Universidad Nacional Autónoma de México, por el apoyo financiero otorgado a MRG mediante una beca doctoral de Conacyt (326085).
Referencias
Álvarez-Mondragón, E. y Morrone, J. J. (2004). Propuesta de áreas para la conservación de aves de México, empleando herramientas panbiogeográficas e índices de complementariedad. Interciencia, 29, 112–120.
Arriaga, L., Espinoza-Rodríguez, J. M., Aguilar-Zúñiga, C., Martínez-Romero, E., Gómez-Mendoza, L. y Loa, E. (Coord.). (2000). Regiones Terrestres Prioritarias de México. Ciudad de México: Conabio. https://doi.org/10.5962/bhl.title.118644
Banks, N. (1900). New genera and species of Nearctic neuropteroid insects. Transactions of the American Entomological Society, 26, 239–259.
Banks, N. (1901). A list of neuropteroid insects from Mexico. Transactions of the American Entomological Society, 27, 361–372.
Barba-Álvarez, R. y Bueno-Soria, J. (2005). New species of the genus Polycentropus (Trichoptera: Polycentropodidae) from Mexico. Proceedings of the Entomological Society of Washington, 107, 663–670.
Barba-Álvarez, R., Bueno-Soria, J. y Santiago-Fragoso, S. (2002). The caddisfly fauna of the Temazcalapa river, Guerrero, México. En W. Mey (Ed.), Proceedings of the Xth International Symposium on Trichoptera (pp. 317–322). Potsdam, Alemania: The Caddis Press.
Blahnik, R. J. (1998). A revision of the Neotropical species of the genus Chimarra, subgenus Chimarra (Trichoptera: Philopotamidae). Memories of the American Entomological Institute, 59, 1–318.
Blahnik, R. J. y Holzenthal, R. W. (2004). Collection and curation of Trichoptera, with emphasis on pinned material. Nectopsyche, Neotropical Trichoptera Newsletter, 1, Suppl., 8–22.
Blahnik, R. J. y Holzenthal, R. W. (2006). Revision of the genus Culoptila (Trichoptera: Glossosomatidae). Zootaxa, 1233, 1–52. http://doi.org/10.5281/zenodo.172764
Blahnik, R. J. y Holzenthal, R. W. (2008). Revision of the Mexican and Central American species of Mortoniella (Trichoptera: Glossosomatidae: Protoptilinae). Zootaxa, 1711, 1–72. https://doi.org/10.11646/zootaxa.1711.1.1
Blahnik, R. J. y Holzenthal, R. W. (2014). Review and redescription of the species in the Oecetis avara group, with the description of 15 new species (Trichoptera, Leptoceridae). Zookeys, 376, 1–83. https://doi.org/10.3897/zookeys.376.6047
Bueno-Soria, J. (1981). Estudios en insectos acuáticos de México I. Trichoptera (Lepticeridae). Cinco nuevas especies de Oecetis McLachlan. Folia Entomológica Mexicana, 49, 103–120.
Bueno-Soria, J. (1983). Five new species of caddisflies (Trichoptera) from Mexico. Proceedings of the Entomological Society of Washington, 85, 450–455.
Bueno-Soria, J. (1984). Estudios en insectos acuáticos II: revisión para México y Centroamérica del género Hydroptila Dalman (Trichoptera: Hydroptilidae). Folia Entomológica Mexicana, 59, 79–138.
Bueno-Soria, J. (1985). Estudios en insectos acuáticos III: cinco nuevas especies de Chimarra Stephens (1829) de México y Centro América (Trichoptera: Philopotamidae). Folia Entomologica Mexicana, 63, 13–23.
Bueno-Soria, J. (1986). Estudios en insectos acuáticos VI: cinco nuevas especies de tricópteros de México y Costa Rica (Trichoptera: Hydropsychidae). Folia Entomologica Mexicana, 68, 53–65.
Bueno-Soria, J. (1990). Estudios en insectos acuáticos VIII. Revisión para México y Centroamérica del género Polyplectropus Ulmer (Trichoptera: Polycentropodidae). Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 61, 357–404.
Bueno-Soria, J. (1999). Studies in aquatic insects XV: new species of Neotrichia and first record of Oxyethira hilosa (Trichoptera: Hydroptilidae) from Mexico. Entomological News, 110, 113–117.
Bueno-Soria, J. (2002). The genus Metrichia Ross (Trichoptera: Hydroptilidae) from México. Transactions of the American Entomological Society, 128, 223–243.
Bueno-Soria, J. (2009). A review of the genus Ochrotrihcia Mosely (Trichoptera: Hydroptilidae) from Mexico and Central America. Transactions of the Entomological Society, 135, 59–160. https://doi.org/10.3157/061.135.0202
Bueno-Soria, J. (2010). Some new Trichoptera (Glossosomatidae, Hydroptilidae, Hydropsychidae and Polycentropodidae) from Mexico. Proceedings of the Entomological Society of Washington, 112, 22–31. https://doi.org/10.4289/0013-8797-112.1.22
Bueno-Soria, J. y Barba-Álvarez, R. (1992). Entomofauna (insectos acuáticos): biogeografía. México D.F.: Instituto de Geografía, Universidad Nacional Autónoma de México/ Atlas Nacional de México/ Naturaleza, IV.8.5.
Bueno-Soria J. y Barba-Álvarez, R. (2011). Trichoptera de Chiapas. En F. Álvarez (Ed.), Chiapas: estudios sobre su diversidad biológica (pp. 345–362). Ciudad de México: Instituto de Biología, UNAM.
Bueno-Soria, J. y Barba-Álvarez, R. (2015). New species of Plectropsyche Ross, 1947 (Trichoptera: Hydropsychidae: Hydropsychiane). Zootaxa, 4040, 421–432. https://doi.org/10.11646/zootaxa.4040.4.2
Bueno-Soria, J. y Hamilton, S.W. (1986). Estudios en insectos acuáticos VI: cinco especies nuevas de tricópteros de México (Trichoptera: Polycentropodidae; Hydroptilidae; Hydropsychidae). Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 57, 299–310.
Bueno-Soria, J. y Holzenthal, R. W. (1986). Estudios en insectos acuáticos V: Descripción de tres nuevas especies de tricópteros de México (Trichoptera: Philopotamidae). Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 57, 137–144.
Bueno-Soria, J. y Holzenthal, R. W. (2004). New species of the genus Ochrotrichia Mosely (Trichoptera: Hydroptilidae) from Mexico and Panama. Transactions of the American Entomological Society, 130, 245–269.
Bueno-Soria, J. y Santiago-Fragoso, S. (1996). Studies in aquatic insects X: Descriptions of five new species of the genus Culoptila Mosely (Trichoptera: Glossosomatidae) from Mexico. Proceedings of the Biological Society of Washington, 109, 446–452.
Bueno-Soria, J., Morrone, J. J. y Barba-Álvarez, R. (2005). Trichoptera of Arroyo Las Flores, Tabasco, Mexico, and their biogeographic affinities. En K. Tanida y A. Rossiter (Eds.), Proceedings of the XIth International Symposium on Trichop-
tera (pp. 73–76). Kanagawa, Japón: Tokai University Press.
Bueno-Soria, J., Morrone, J. J. y Barba-Álvarez, R. (2007). Trichoptera of the Sierra Tarahumara, Chihuahua, México. En J. Bueno-Soria, R. Barba-Álvarez y B. Armitage (Eds.), Proceedings of the XIIth International Symposium on Trichoptera (pp. 31–34). México D.F.: The Caddis Press.
Bueno-Soria, J., Santiago-Fragoso, S. y Barba-Álvarez, R. (2001). Studies in aquatic insects, XVIII: new species and new record of caddisflies (Trichoptera) from Mexico. Entomological News, 112, 145–158.
Cifuentes, J., Villegas, M., García-Sandoval, R., Vidal-Gaona, G., Sierra, S., Valenzuela-Garza, R. et al. (2004). Distribución de macromicetos: Una aproximación al análisis de áreas de endemismos. En I. Luna-Vega, J. J. Morrone y D. Espinosa (Eds.), Biogeografía de la Sierra Madre Oriental (pp. 355–374). México D.F.: Conabio/ UNAM.
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2020). Listado de áreas destinadas voluntariamente a la conservación. Recuperado el 19 agosto, 2020 de: https://advc.conanp.gob.mx/listado-de-advc/
Contreras-Medina, R. y Eliosa-León, H. (2001). Una visión panbiogeográfica preliminar de México. En J. Llorente-Bousquets y J. J. Morrone (eds.), Introducción a la biogeografía en Latinoamérica: teorías, conceptos, métodos y aplicaciones (pp. 197–211) México D.F.: Las Prensas de Ciencias, Universidad Nacional Autónoma de México.
Contreras-Medina, R., Luna, I. y Morrone, J. J. (2007). Application of parsimony analysis of endemicity to Mexican gymnosperm distributions: grid-cells, biogeographical provinces and track analysis. Biological Journal of the Linnean Society, 92, 405–417. https://doi.org/10.1111/j.1095-8312.2007.00844.x
Corona, A. Ma. y Morrone, J. J. (2005). Track analysis of the species of Lampetis (Spinthoptera) Casey, 1909 (Coleoptera: Buprestidae) in North America, Central America, and the West Indies. Caribbean Journal of Science, 41, 37–41.
Corona, A. M., Toledo, V. H. y Morrone, J. J. (2007). Does the Trans-Mexican Volcanic Belt represent a natural biogeographical unit? An analysis of the distributional patterns of Coleoptera. Journal of Biogeography, 34, 1008–1015. https://doi.org/10.1111/j.1365-2699.2006.01666.x
Craw, R. C., Grehan, J. R. y Heads, M. J. (1999). Panbiogeography: tracking the history of Life. New York: Oxford University Press.
Cué-Bär, E. M., Villaseñor, J. L., Morrone J. J. e Ibarra-Manríquez, G. (2006). Identifying priority areas for conservation in Mexican tropical deciduous forest based on tree species. Interciencia, 31, 712–719.
de Moor, F. C. e Ivanov, V. D. (2008). Global diversity of caddisflies (Trichoptera: Insecta) in freshwater. Hidrobiologia, 595, 393–407. https://doi.org/10.1007/s10750-007-9113-2
Denning, D. G. (1941). Descriptions of three new species of Mexican Chimarra (Trichoptera: Philopotamidae). Entomological News, 52, 82–85.
Denning, D. G. (1952). Descriptions of several new species of caddisflies. The Canadian Entomologist, 84, 17–22. https://doi.org/10.4039/Ent8417-1
Denning, D. G. (1965). New Hydropsychidae (Trichoptera). Journal of the Kansas Entomological Society, 38, 75–84.
Denning, D. G. (1968). New species and notes of western Trichoptera. Journal of the Kansas Entomological Society, 41, 63–69.
Denning, D. G. y Blickle, R. L. (1979). New species of Helicopsyche (Trichoptera: Helicopsychidae). Pan Pacific Entomologist, 55, 27–33.
Denning, D. G. y Sykora, J. (1966). New North American Trichoptera. The Canadian Entomologist, 98, 1219–1226. https://doi.org/104039/Ent981219-11
Echeverry, A. y Morrone, J. J. (2010). Parsimony analysis of endemicity as a panbiogeographical tool: an analysis of Caribbean plant taxa. Biological Journal of the Linnean Society, 101, 961–976. https://doi.org/10.1111/j.1095-8312.2010.01535.x
Echeverry, A. y Morrone, J. J. (2013). Generalized tracks, area cladograms and tectonics in the Caribbean. Journal of Biogeography, 40, 1619–1637. http://doi.org/10.1111/jbi.12117
Escalante, T., Noguera-Urbano, E. A. y Corona, W. (2018). Track analysis of the Nearctic region: identifying complex areas with mammals. Journal of Zoological Systematics and Evolutionary Research, 56, 466–477. https://doi.org/10.1111/jzs.12211
Escalante, T., Rodríguez, G. y Morrone, J. J. (2004). The diversification of Nearctic mammals in the Mexican transition zone. Biological Journal of the Linnean Society, 83, 327–339. https://doi.org/10.1111/j.1095-8312.2004.00386.x
Escalante, T., Rodríguez, G. y Morrone, J. J. (2005). Las provincias biogeográficas del Componente Mexicano de Montaña desde la perspectiva de los mamíferos continentales. Revista Mexicana de Biodiversidad, 76, 199–205. http://dx.doi.org/10.22201/ib.20078706e.2005.002.304
Flint, O. S. Jr. (1965). New species of Trichoptera from the United States. Proceedings of the Entomological Society of Washington, 67, 168–176.
Flint, O. S. Jr. (1967). Studies of Neotropical caddisflies, IV: new species from Mexico and Central America. Proceedings of the United States National Museum, 123, 1–24. https://doi.org/10.5479/si.00963801.123-3608.1
Flint, O. S. Jr. (1970). Studies of Neotropical caddisflies X: Leucotrichia and related genera from North and Central America (Trichoptera: Hydroptilidae). Smithsonian Contributions to Zoology, 60, 1–64. https://doi.org/10.5479/si.00810282.60
Flint, O. S. Jr. (1971). Studies of Neotropical caddisflies, XI: the genus Rhyacopsyche in Central America (Hydroptilidae). Proceedings of the Biological Society of Washington, 46, 515–526.
Flint, O. S. Jr. (1972). Studies of Neotropical caddisflies, XIII: the genus Ochrotrichia from Mexico and Central America (Trichoptera: Hydroptilidae). Smithsonian Contributions to Zoology, 118, 1–28. https://doi.org/10.5479/si.00810282.118
Flint, O. S. Jr. (1974). Studies of Neotropical caddisflies XVII: the genus Smicridea from North and Central America (Trichoptera: Hydropsychidae). Smithsonian Contributions to Zoology, 167, 1–65. https://doi.org/10.5479/si.00810282.167
Flint, O. S. Jr. (1981). Studies of Neotropical caddisflies, XXVIII: the Trichoptera of the Río Limón Basin, Venezuela. Smithsonian Contributions to Zoology, 330, 1–61. https://doi.org/10.5479/si.00810282.330
Flint, O. S. Jr. (1985). Studies of Neotropical caddisflies, XXXVI: the genus Cochliopsyche in Middle America (Trichoptera: Helicopsychidae). Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 56, 213–216.
Flint, O.S. Jr. (1991). Studies of Neotropical caddisflies, XLV: the taxonomy, phenology and faunistics of the Trichoptera of Antioquia, Colombia. Smithsonian Contributions to Zoology, 520, 1–113. https://doi.org/10.5479/si.00810282.520
Flint, O. S. Jr. (1998). Studies of Neotropical caddisflies, LIII: a taxonomic revision of the subgenus Curgia of the genus Chimarra (Trichoptera: Philopotamidae). Smithsonian Contributions to Zoology, 594, 1–131. https://doi.org/10.5479/si.00810282.594
Flint, O. S. Jr. y Bueno-Soria, J. (1977). Studies of Neotropical caddisflies, XXI: the genus Lepidostoma (Trichoptera: Lepidostomatidae). Proceedings of the Entomological Society of Washington, 90, 375–387.
Flint, O. S. Jr., McAlpine, J. F. y Ross, H. H. (1987). A revision of the genus Leptonema Guérin (Trichoptera: Hydropschidae: Macronematinae). Smithsonian Contributions to Zoology, 450, 1–193. https://doi.org/10.5479/si.00810282.450
García-Barros, E., Guerrea, P., Luciáñez, M. J., Cano, J. M., Munguira, M. L., Moreno, J. C. et al. (2002). Parsimony analysis of endemicity and its application to animal and plant geographical distributions in the Ibero-Balearic region (western Mediterranean). Journal of Biogeography, 29, 109–124. https://doi.org/10.1046/j.1365-2699.2002.00653.x
García-Marmolejo, G., Escalante, T. y Morrone, J. J. (2008). Establecimiento de prioridades para la conservación de mamíferos terrestres neotropicales de México. Mastozoología Neotropical, 15, 41–65.
Goloboff, P. 1999. NONA, versión 2.0. Publicado por el autor, Tucumán.
González, A., Contreras, R., Espinosa, D. y Luna, I. (2017). Track analysis of the order Gomphales (Fungi: Basidiomycota) in Mexico. Phytotaxa, 316, 22–38. https://doi.org/10.11646/phytotaxa.316.1.2
Hagen, H. (1861). Synopsis of the Neuroptera of North America with a list of the South American species, Smithsonian Institution Miscellaneous Collections, 4, 1–347. https://do.ogr/10.5962/bhl.title.60275
Halffter, G. (2003). Biogeografía de la entomofauna de montaña de México y América Central. En J. J. Morrone y Llorente, J. (Eds.), Una perspectiva latinoamericana de la biogeografía (pp. 87–97). Ciudad de México: Las Prensas de Ciencias, Universidad Nacional Autónoma de México.
Harris, S. C. y Contreras-Ramos, A. (1989). Ithytrichia mexicana (Trichoptera: Hydroptilidae), a new species of caddisfly from Mexico. Entomological News, 100, 176–178.
Holzenthal, R. W. (1988). Catálogo sistemático de los tricópteros de Costa Rica. Brenesia, 29, 51–83.
Holzenthal, R. W. (1995). The caddisfly genus Nectopsyche: new gemma group species from Costa Rica and the Neotropics (Trichoptera: Leptoceridae). Journal of the North American Benthological Society, 14, 61–83. https://doi.org/10.2307/1467725
Holzenthal, R. W. y Andersen, T. (2004). The caddisfly genus Triaenodes in the Neotropics (Trichoptera: Leptoceridae). Zootaxa, 511, 1–80. https://doi.org/10.11646/zootaxa.511.1.1
Holzenthal, R. W. y Harris, S. C. (1992). Hydroptilidae (Trichoptera) of Costa Rica: the genus Oxyethira Eaton. Journal of the New York Entomological Society, 100, 155–177.
Holzenthal, R. W., Blahnik, R. J., Prather, A. L. y Kjer, K. M. (2007). Order Trichoptera Kirby, 1813 (Insecta), caddisflies. Zootaxa, 1668, 639–698. https://doi.org/10.11646/zootaxa.1668.1.29
Houghton, D.C., Berry, E.A., Gilchrist, A., Thompson, J. y Nussbaum, M. A. (2011). Biological changes along the continuum of an agricultural stream: influence of a small terrestrial preserve and use of adult caddisflies in biomonitoring. Journal of Freshwater Ecology, 26, 381–397. https://doi.org/10.1080/02705060.2011.563513
Johanson, K. A. (2002). Systematic revision of American Helicopsyche of the subgenus Feropsyche (Trichoptera: Helicopsychidae). Entomologica Scandinavia, 60, 1–144.
León-Paniagua, L. y Morrone, J. J. (2009). Do the Oaxacan Highlands represent a natural biotic unit? A cladistic biogeographical test based on vertebrate taxa. Journal of Biogeography, 36, 1939–1944. https://doi.org/10.1111/j.1365-2699-2009-02134.x
Luna, I., Alcántara, O., Espinosa, D. y Morrone, J. J. (1999). Historical relationship of the Mexican cloud forests; a preliminary vicariance model applying Parsimony Analysis of Endemicity to vascular plant taxa. Journal of Biogeography, 26, 1299–1305. https://doi.org/10.1046/j.1365-2699.199.00361.x
Luna, I., Alcántara, O., Morrone, J. J. y Espinosa, D. (2000). Track analysis and conservation priorities in the cloud forests of Hidalgo, Mexico. Diversity and Distributions, 6, 137–143. https://doi.org/10.1046/j.1472-4642-2000.00079.x
Mackay, R. J. y Wiggins, G.B. (1979). Ecological diversity in Trichoptera, Annual Review of Entomology, 24, 185–208. https://doi.org/10.1146/annurev.en.24.010179.001153
Malm, T., Johanson, K. A. y Wahlberg, N. (2013). The evolutionary history of Trichoptera (Insecta): a case of successful adaptation to life in freshwater. Systematic Entomology, 38, 459–473. https://doi.org/10.1111/syen.12016
Márquez, J. y Asiain, J. (2006). Patrones de distribución de la familia Staphylinidae (Coleoptera). En J. J. Morrone y J. Llorente-Bousquets (Eds.), Componentes bióticos principales de la entomofauna mexicana (pp. 157–236). Ciudad de México: Las Prensas de Ciencias, Universidad Nacional Autónoma de México.
Márquez, J. y Morrone, J. J. (2003). Análisis panbiogeográfico de las especies de Heterolinus y Homalolinus (Coleoptera: Staphylinidae: Xantholinini). Acta Zoológica Mexicana, 90, 15–25.
Maya-Martínez, A., Schmitter-Soto, J. J. y Pozo, C. (2011). Panbiogeography of the Yucatan península based on Charaxinae (Lepidoptera: Nymphalidae). Florida Entomologist, 94, 527–533.
Morrone, J. J. (1994). On the identification of areas of endemism. Systematic Biology, 43, 438–441. https://doi.org/10.1093/sysbio/43.3.438
Morrone, J. J. (2004a). Panbiogeografía, componentes bióticos y zonas de transición. Revista Brasileira de Entomologia, 48, 149–162. https://doi.org/10.1590/S0085-56262004000200001
Morrone, J. J. (2004b). Homología biogeográfica. Las coordenadas espaciales de la vida. Cuadernos del Instituto de Biología 37. México D.F.: Instituto de Biología, UNAM.
Morrone, J. J. (2017). Biogeographic regionalization of the Sierra Madre del Sur province, Mexico. Revista Mexicana de Biodiversidad, 88, 710–714. https://doi.org/10.1016/j.rmb.2017.07.012
Morrone, J. J. (2019). Regionalización biogeográfica y evolución biótica de México: encrucijada de la biodiversidad del Nuevo Mundo. Revista Mexicana de Biodiversidad, 90, 1–68. https://doi.org/10.22201/ib.20078706e.2019.90.2980
Morrone, J. J. y Escalante, T. (2002). Parsimony analysis of endemicity (PAE) of Mexican terrestrial mammals at different area units: when size matters. Journal of Biogeography, 29, 1095–1104. https://doi.org/10.1046/j.1365-2699.2002.00753.x
Morrone, J. J. y Gutiérrez, A. (2005). Do fleas (Insecta: Siphonaptera) parallel their mammal host diversification in the Mexican transition zone? Journal of Biogeography, 32, 1315–1325. https://doi.org/10.1111/j.1365-2699.2005.01285.x
Morrone, J.J. y Márquez, J. (2001). Halffter’s Mexican transition zone, beetle generalized tracks, and geographical homology. Journal of Biogeography 28, 635–650. https://doi.org/101046/j.1365-2699.2001.00571.x
Morrone, J. J. y Márquez, J. (2003). Aproximación a un Atlas Biogeográfico Mexicano: Componentes bióticos principales y provincias biogeográficas. En J. J. Morrone y J. Llorente-Bousquets (Eds.), Una perspectiva latinoamericana de la biogeografía (pp. 217–220). Ciudad de México: Las Prensas de Ciencias, Universidad Nacional Autónoma de México.
Morrone, J. J. y Márquez, J. (2008). Biodiversity of Mexican terrestrial arthropods (Arachnida and Hexapoda): a biogeographical puzzle. Acta Zoológica Mexicana, 24, 15–41. https://doi.org/10.21829/azm.2008.241613
Morrone, J. J., Escalante, T. y Rodríguez-Tapia, G. 2017. Mexican biogeographic provinces: map and shapefiles. Zootaxa, 4277, 277–279. https://doi.org/10.11646/zootaxa.4277.2.8
Morrone, J. J., Espinosa, D., Aguilar, C. y Llorente, J. (1999). Preliminary classification of the Mexican biogeographic provinces: a parsimony analysis of endemicity based on plant, insect, and bird taxa. Southwestern Naturalist, 44, 507–514. https://doi.org/10.2307/3672351
Mosely, M. E. (1934). New exotic Hydroptilidae. Transactions of the Royal Entomological Society of London, 82, 137–163. https://doi.org/10.1111/j.1365-2311.1934.tb00031.x
Mosely, M. E. (1937). Mexican Hydroptilidae (Trichoptera). Transactions of the Royal Entomological Society of London, 86, 151–190. https://doi.org/10.1111/j.1365-2311.1937.tb00242.x
Mosely, M. E. (1954). The Protoptila group of the Glossosomatinae (Trichoptera: Rhyacophilidae). Bulletin of the British Museum (Natural History). Entomology, 3, 317–346. https://doi.org/10.5962/bhl.part.1056
Moulton, S. R. y Harris, S. C. (1997). New species southwestern Nearctic microcaddisflies (Trichoptera: Hydroptilidae). Proceedings of the Entomological Society of Washington, 99, 494–501.
Muñoz-Quesada, J. F. y Holzenthal, R. W. (2015). Revision of the Neotropical species of the caddisfly genus Wormaldia McLachlan (Trichoptera: Philopotamidae). Zootaxa, 3998, 1–138. https://doi.org/10.11646/zootaxa.3998.1.1
Nixon K. C. (2002). WinClada, v 1.00.08. University of Ithaca, New York.
Oláh, J. y Johanson, K. A. (2011). New Neotropical Hydroptilidae (Trichoptera). Annales Historico-Naturales Musei Nationalis Hungarici, 103, 117–255.
Oláh, J. y Johanson, K. A. (2012). Description of new species in the Leucotrichiinaetribe (Trichoptera: Hydroptilidae). Annales Historico-Naturales Musei Nationalis Hungarici, 104, 131–213. https://doi.org/10.1155/2012/916718
Razo-González, M. (2018). Caddisflies (Insecta: Trichoptera) from Santa Catarina Lachatao, Oaxaca, México: New species, new geographical records, and checklist. Zootaxa, 4388, 22–40. https://doi.org/10.11646/zootaxa.4388.1.2
Razo-González, M., Cataño-Meneses, G., Novelo-Gutiérrez, R. y Márquez, J. (2020). Preliminary evaluation of the nocturnal flight of caddisflies (Insecta: Trichoptera) in a temperate forest in Oaxaca, Mexico. Aquatic Insects, 41, 339–355. https://doi.org./10.1080/01650424.2020.1797818
Rojas-Ascencio, A., Bueno-Soria, J. y Gaviño-Rojas, R. (2002). Trichoptera from Arroyo Colorado, Municipality of Temascaltepec, State of Mexico, Mexico. Nova Supplementa Entomologica (Proceedings of the 10th International Symposium on Trichoptera), 15, 375–378.
Rosen, B. R. (1988). From fossils to earth history: applied historical biogeography. En A. A. Myers y P. S. Giller (Eds.), Analytical biogeography, Vol. 1 (pp. 437–481). Londres: Chapman y Hall.
Ross, H. H. (1947). Descriptions and records of North American Trichoptera, with synoptic notes. Transactions of the American Entomological Society, 73, 125–168.
Ross, H. H. (1948). Notes and descriptions of Nearctic Hydroptilidae (Trichoptera). Journal of the Washington Academy of Sciences, 38, 201–206.
Ross, H. H. (1951). The Trichoptera of Lower California. Proceedings of the California Academy of Sciences, 27, 65–76.
Ross, H. H. (1956). Evolution and classification of the mountain caddisflies. Urbana, Illinois: University of Illinois Press.
Ross, H. H. (1959). New species of Chimarra from Mexico and Central America (Trichoptera: Philopotamidae). Entomological News, 70, 169–178.
Ross, H. H. y King, E.W. (1952). Biogeographic and taxonomic studies in Atopsyche (Trichoptera, Rhyacophilidae). Annals of the Entomological Society of America, 45, 177–204. https://doi.org/10.1093/aesa/45.2.177
Sanginés-Franco, C., Luna-Vega, I., Alcántara, O. y Contreras-Medina, R. (2011). Distributional patterns and biogeographic analysis of ferns in the Sierra Madre Oriental, Mexico. American Fern Journal, 101, 81-104. https://doi.org/10.1640/0002-8444-101.2.81
Schmid, F. (1982). La famille des Xiphocentronides (Trichoptera:Annulipalpia). Memories de la Societé Entomologique du Canada, 121, 1–127. https://doi.org/10.4039/entm114122fv
Schmid, F. (1989). Les Hydrobiosides (Trichoptera, Annulipalpia). Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, 59, 1–154.
Smith, A. B. (1992). Echinoid distribution in the Cenomanian: an analytical study in biogeography. Palaeogeography, Palaeoclimatology, Palaeoecology, 93, 263–276. https://doi.org/10.1016/0031-0182(92)90086-k
Thomas, J. A., Frandsen, P. B., Prendini, E., Zhou, X. y Holzenthal, R. W. (2020). A multigene phylogeny and timeline for Trichoptera (Insecta). Systematic Entomology, 45, 670–686. https://doi.org/1.1111/syen.12422
Thompson, R. E. y Holzenthal, R. W. (2015). A revision of the Neotropical caddisfly genus Leucotrichia Mosely, 1934 (Hydroptilidae: Leucotrichiinae). Zookeys, 499, 1–100. https://doi.org/10.3897/zookeys.499.8360
Toledo, V. H., Corona, A. M. y Morrone, J. J. (2007). Track analysis of the Mexican species of Cerambycidae (Insecta, Coleoptera). Revista Brasileira de Entomologia, 5, 131–137. https://doi.org/10.1590/S0085-56262007000200002
Torres-Colín, R. (2004). Tipos de vegetación. En A. J. García-Mendoza, M. J. Ordoñez y M. Briones-Salas (Eds.), Biodiversidad de Oaxaca (pp. 105–117). Ciudad de México: Instituto de Biología, Universidad Nacional Autónoma de México/ Fondo Oaxaqueño para la Conservación de la Naturaleza/ World Wildlife Fund.
Trejo, I. (2004). Clima. En A. J. García-Mendoza, Ordoñez, M. J. y Briones-Salas, M. (Eds.), Biodiversidad de Oaxaca (pp. 67–85). Ciudad de México, México: Instituto de Biología, Universidad Nacional Autónoma de México-Fondo Oaxaqueño para la Conservación de la Naturaleza-World Wildlife Fund.
Weaver, J. S., III. (1988). A synopsis of the North American Lepidostomatidae (Trichoptera). Contributions of the American Entomological Institute, 24, 1–141.
Wiggins, G. B. (1996). Larvae of the North American Caddisfly genera (Trichoptera). Toronto, Canadá: University of Toronto Press.


