Análisis del conocimiento histórico de la distribución geográfica y ecológica del loro frente blanca, Amazona albifrons, con evidencia de colonización reciente
Claudio Mota-Vargas a, Karla Patricia Parra-Noguez b y Octavio Rojas-Soto a, *
a Red de Biología Evolutiva, Laboratorio de Bioclimatología, Instituto de Ecología, A.C, Carretera antigua a Coatepec 351, El Haya, 91073 Xalapa, Veracruz, México
b Universidad Michoacana de San Nicolás de Hidalgo, Ciudad Universitaria, Francisco J. Múgica s/n., 58194 Morelia, Michoacán, México
*Autor para correspondencia: octavio.rojas@inecol.mx (O. Rojas-Soto)
Recibido: 17 mayo 2018; aceptado: 4 febrero 2020
Resumen
La distribución del loro frente blanca, Amazona albifrons, en México incluye la vertiente del Pacífico y del golfo, desde Sonora y el sur de Veracruz, respectivamente, y continúa hasta Costa Rica. Recientemente, se ha registrado más allá de su área de distribución histórica conocida, aparentemente por introducciones humanas, pero los registros en el centro de Veracruz parecen corresponder a una colonización reciente. En este trabajo analizamos la distribución geográfica de A. albifrons mediante el uso de literatura especializada, modelos de distribución de especies y trabajo de campo en Veracruz entre los años 2013-2016. Registramos las fechas de observación, la conducta reproductiva y sus hábitos alimenticios. A partir de esta información, delimitamos el área de distribución de la especie desde México hasta Panamá. Con base en los registros a lo largo del año durante los 4 años de trabajo de campo y la observación de conducta reproductiva durante el invierno y la primavera, consideramos que ha ampliado recientemente su distribución hacia el centro de Veracruz, que incluye ± 200 km al norte de la distribución histórica conocida en el este de México. Finalmente, analizamos e identificamos un total de 14 plantas de las que se alimenta en esta región.
Palabras clave: Área de distribución; Ampliación de distribución; Modelos de distribución de especies; Psittacidae; Recursos alimenticios
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Analysis of the historical knowledge on the geographic distribution of the White-fronted Parrot, Amazona albifrons, with evidence of recent colonization
Abstract
The distribution of the White-fronted Parrot, Amazona albifrons, in Mexico includes the Pacific and the gulf slopes, from southern Sonora and southern Veracruz, respectively, and continues to Costa Rica. Recently, it has been reported beyond the known historical distribution area, apparently due to human introductions, but the records found in central Veracruz seem to correspond to a recent natural colonization. In this study, we analyzed the geographic distribution of A. albifrons through information from specialized literature, the use of species distribution models, and field work in central Veracruz from 2013-2016. We recorded the observation dates, its reproductive behavior, and feeding habits. Based on this information, we delimited the range of the species from Mexico to Panama. Based on year-round records during the 4 years of fieldwork and observation of reproductive behavior during winter and spring, we consider that it has recently expanded its distribution to central Veracruz, which includes ± 200 km north of the known historical distribution in eastern Mexico. Finally, we analyzed and identified a total of 14 species of plants in this region from which the species feed.
Keywords: Distribution area; Distribution extension; Species distribution models; Psittacidae; Food resources
© 2020 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
Delimitar el área de distribución de las especies no es una tarea sencilla, ya que existen diferentes conceptos para definirla, así como distintos métodos para delimitarla (Antúnez y Mendoza, 1992; Mota-Vargas y Rojas-Soto, 2012; Rappoport, 1975). Sin embargo, su reconocimiento es necesario, ya que el área de distribución es la unidad fundamental de la biogeografía (Peterson et al., 2004), el contexto histórico y espacial de la sistemática (Morrone, 2001; Zunino y Zullini, 2003) y la línea base para cualquier enfoque en conservación (Anderson y Martínez-Meyer, 2004; Peterson, 2001).
Los registros de presencia de las especies son de natural importancia ya que se parte de ellos para construir y delimitar las áreas de distribución, independientemente del concepto y el método empleado (Peterson et al., 2011; Zunino y Zullini, 2003). No obstante, los registros de presencia no siempre representan novedades biogeográficas y/o ecológicas, que se traduzcan en un incremento en el conocimiento que tenemos sobre las distribuciones de las especies. Recientemente, Sánchez-González (2013) discutió cuándo el registro de una especie en particular debe ser considerado como un registro nuevo que puede ayudar a reconfigurar su distribución, o bien, reconocerlo solo como un registro esperado con base en su distribución histórica ya conocida. En ese trabajo se destacan diversos criterios para identificar como novedoso biogeográficamente a un registro, por ejemplo, cuando se registra al taxón por primera vez en una entidad política, se reporta al taxón en una región biótica distinta, o bien, cuando se confirma la presencia del taxón en un sitio en donde no hay datos publicados que lo confirmen.
Con respecto a este estudio de caso, el loro frente blanca, Amazona albifrons, habita en diferentes tipos de vegetación, como el bosque tropical caducifolio y subcaducifolio, el bosque nublado, el bosque de pino-encino, la vegetación riparia, los manglares, así como en áreas perturbadas que incluyen cultivos de café bajo sombra y zonas urbanas; se encuentra a una altitud de 1,800 m snm (González-García et al., 2014; Howell y Webb, 1995; Marín-Togo et al., 2012; Plasencia-Vázquez y Escalona-Segura, 2012). Esto sugiere que la especie es ecológicamente plástica ante diferentes tipos de ambientes; lo que, en conjunto con su capacidad de dispersión, le han permitido abarcar una región biogeográficamente compleja, como lo es la región Mesoamericana (Ríos-Muñoz, 2013; Ríos-Muñoz y Navarro-Sigüenza, 2009).
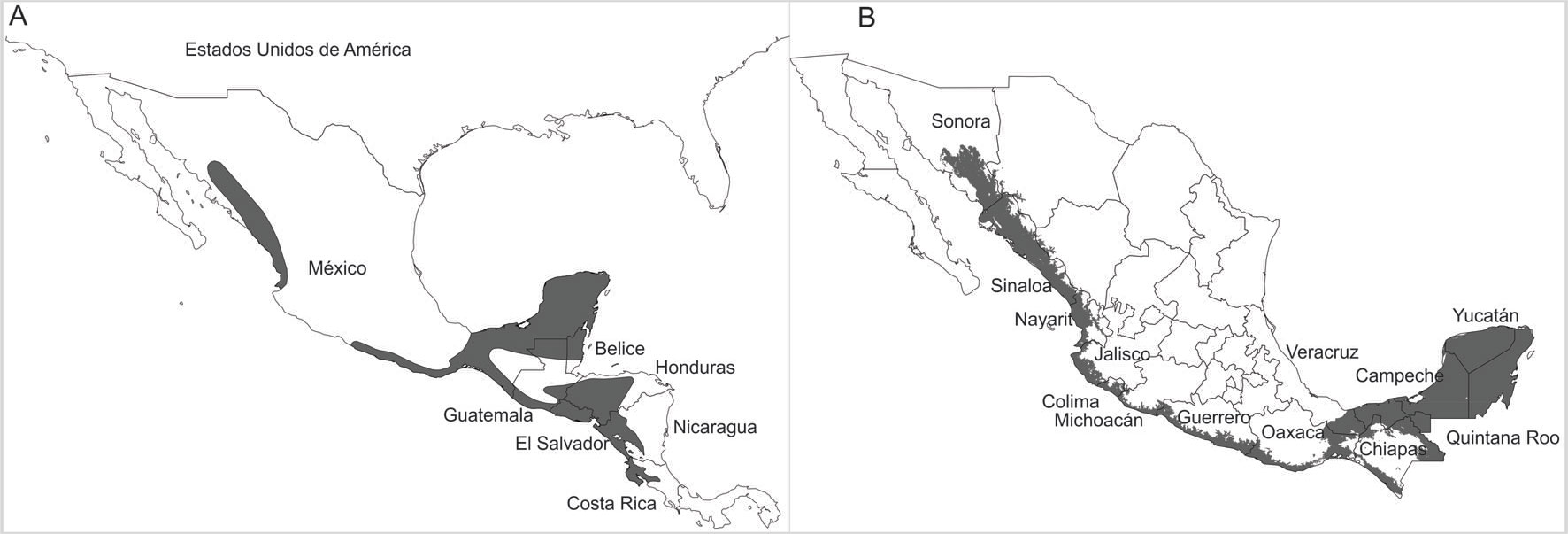
A la fecha, se han hecho diferentes propuestas para delimitar el área de distribución del loro frente blanca (e.g., Forshaw y Cooper, 1989; Howell y Webb, 1995; fig. 1A), mismas que en su mayoría han utilizado métodos geográficos convencionales para definirla (mano alzada: Rappoport, 1975; Zunino y Zullini, 2003). No obstante, existen herramientas como los modelos de distribución de especies (MDE), que permiten delimitar el área de distribución con base en algoritmos matemáticos correlativos, que combinan los registros geográficos con variables ambientales para obtener áreas de presencia potencial de las especies y que han mostrado resultados más satisfactorios en comparación con los métodos tradicionales, ya que además pueden ser estadísticamente validados (Mota-Vargas y Rojas-Soto, 2012; Peterson, 2001; Peterson et al., 2018; Phillips et al., 2006; Stockwell y Peters, 1999). De hecho, los MDE ya han sido utilizados recientemente por otros autores para delimitar el área de distribución del loro frente blanca (Monterrubio-Rico et al., 2016; Navarro-Sigüenza y Peterson, 2007; Ríos-Muñoz y Navarro-Sigüenza, 2009; fig. 1B). Sin embargo, estos trabajos se limitaron a describir su distribución en México y sin considerar aspectos metodológicos fundamentales dentro del MDE y actualmente establecidos, como el número de registros (Wisz et al., 2008), así como la necesidad de establecer “a priori” el área de accesibilidad y los ambientes que la especie ha podido explorar (Anderson y Raza, 2010; Barve et al., 2011); es decir, se requiere incluir la delimitación total del área de distribución conocida de la especie y contextualizar biogeográficamente los registros.
En el presente trabajo realizamos una revisión histórica de la información disponible en la literatura, en internet y a través de trabajo de campo; y describimos la acumulación temporal del conocimiento sobre la distribución geográfica del loro frente blanca. Este es un ejemplo en donde se pueden distinguir los registros biogeográficamente novedosos cuando: 1) corresponden con registros realizados en áreas fuera de su distribución histórica conocida, específicamente por falta de muestreo, 2) representan registros por introducción antropogénica (considerados como exóticos), y 3) los registros son recientes ampliaciones naturales de su distribución (i.e., colonizaciones). Estos últimos registros son de particular importancia porque podrían representar una evidencia de aclimatación de poblaciones silvestres ante los efectos de cambio climático o modificaciones antropogénicas. Además, discutimos algunos aspectos relacionados con la estacionalidad, conducta reproductiva y alimentación de las poblaciones que posible y recientemente colonizaron el centro de Veracruz.
Materiales y métodos
En este trabajo consideramos como “distribución geográfica conocida” a aquella área que es potencialmente ocupada por una especie y que está definida a partir de registros de presencia históricamente acumulados (fig. 2A). De aquí surge la importancia de los nuevos registros de una especie, que implica el poder distinguir, cuáles son los registros esperados, de los que resultan novedosos; donde los esperados son aquellos presentes dentro o muy cerca de la distribución histórica conocida y los novedosos son aquellos que se encuentran significativamente más allá de dicha distribución (Sánchez-González, 2013); estos últimos son los más importantes biogeográfica y ecológicamente, ya que aportan nueva información que modifica y enriquece el conocimiento de la distribución de una especie (fig. 2A, B).
Para llevar a cabo la compilación de los registros, se realizó una búsqueda exhaustiva de los registros históricos del loro frente blanca en literatura (Calvo y Blake, 1998; Chávez-León, 2007; De Labra et al., 2010; González-García, 2006; González-García et al., 2014; Jordano, 1983; Kantak, 1979; Monroy-Ojeda e Isern, 2013; Plasencia-Vázquez y Escalona-Segura, 2012; Robertson y Woolfenden, 1992; Ruelas-Inzunza y Aguilar-Rodríguez, 2010). Además, se revisaron bases de datos disponibles en línea, como la Infraestructura Global de Información sobre Biodiversidad (GBIF, 2017, por sus siglas en inglés), la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio, 2016) y el Laboratorio de Ornitología de la Universidad de Cornell (EBIRD, 2016). La búsqueda de los registros en las bases de datos se realizaron directamente con el nombre científico de la especie, Amazona albifrons, y se descargó toda la información tal cual se obtuvo. La curación y edición de la base incluyó la eliminación de los registros que no contaban con coordenadas, localidad y fecha, y sólo se dejaron registros únicos por fecha y localidad, eliminando todos aquellos dudosos. Posteriormente, se realizó una reconstrucción histórica del conocimiento de su distribución geográfica por décadas, a partir de la descripción de la especie hasta la actualidad; es decir, los registros se identificaron y se representaron de manera acumulativa, en mapas que abarcaron unidades temporales de 10 años (fig. 3).
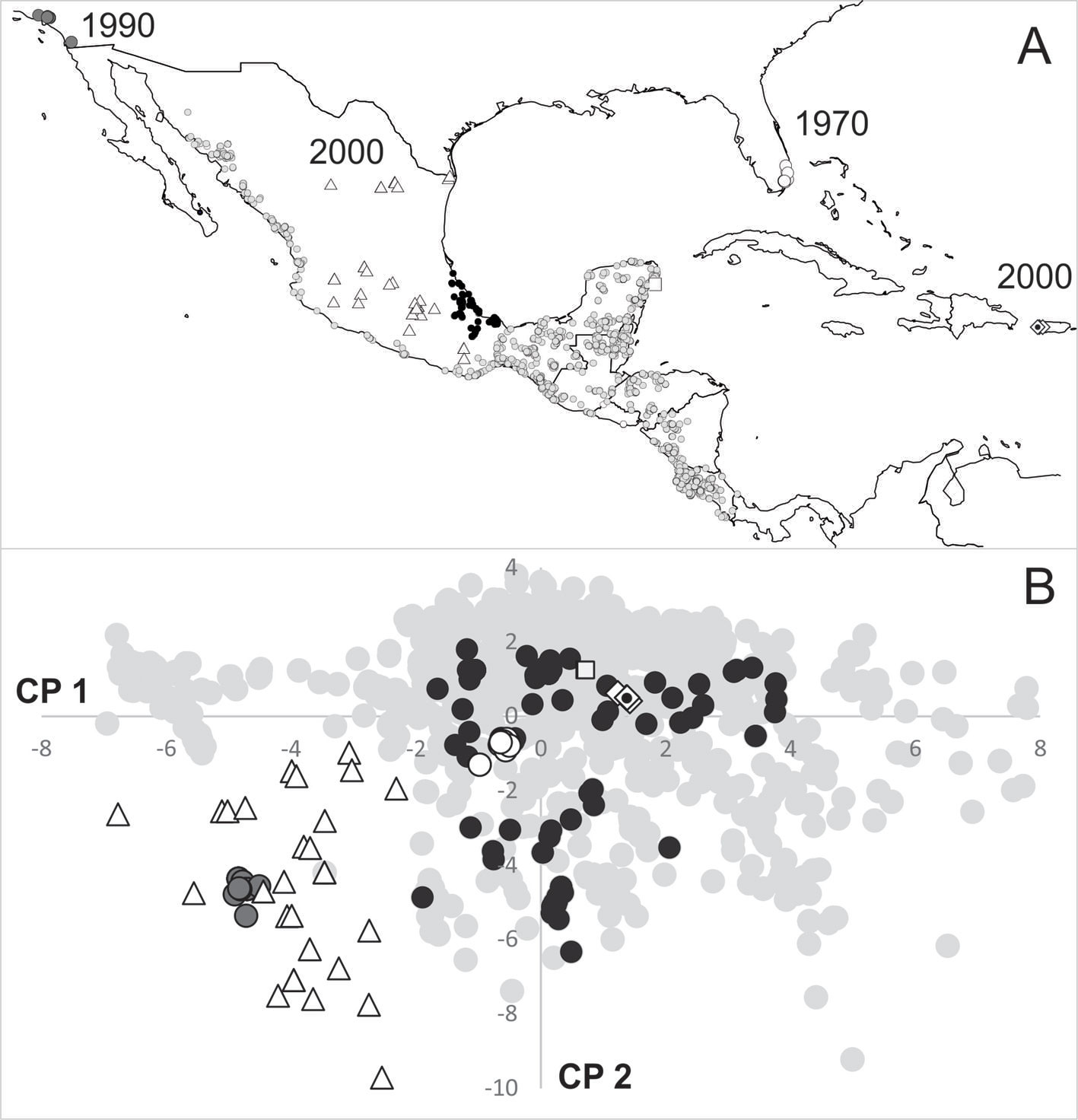
Uno de los criterios para evaluar si los registros novedosos (aquellos más allá de su distribución conocida) son ambientalmente importantes, es analizar si corresponden también con ambientes novedosos (fig. 2B) (Sánchez-González, 2013). Por eso se llevó a cabo una descripción del perfil bioclimático a partir de los registros totales compilados de la especie, en combinación con 19 variables bioclimáticas (tabla 1) obtenidas del proyecto Worldclim a una resolución espacial de 30 segundos (≈ 1 km²) (Hanna et al., 2016; Hijmans et al., 2005). Con ayuda del SIG (Arcview 3.3; ESRI, 1999) se extrajeron los valores de cada una de las variables climáticas correspondientes con cada registro de la especie y mediante un análisis de componentes principales (ACP) realizado en PAST (Hammer et al., 2001), se analizaron e identificaron a aquellos registros ambientalmente novedosos, con base en la distancia en el espacio ambiental que guardaron con relación a los registros históricamente conocidos (fig. 4).
Se llevó a cabo trabajo de campo para buscar a la especie en diversas localidades de los municipios de Xalapa, Coatepec y Xico, ubicados en el centro del estado de Veracruz (fig. 4). Con base en la cantidad de localidades de registro, la abundancia relativa de individuos, así como la existencia de conductas reproductivas, evaluamos si la especie ha colonizado esta zona en las últimas décadas, o bien, si sus registros en la región podrían ser accidentales. Se llevaron a cabo salidas de campo de un día a partir de agosto del año 2013, hasta agosto de 2016; con un promedio de 12 salidas por cada estación del año (≈ 3 salidas por mes). Cada visita incluía caminatas por la mañana (6:00 – 11:00 h) y por la tarde (17:00 – 19:30 h).

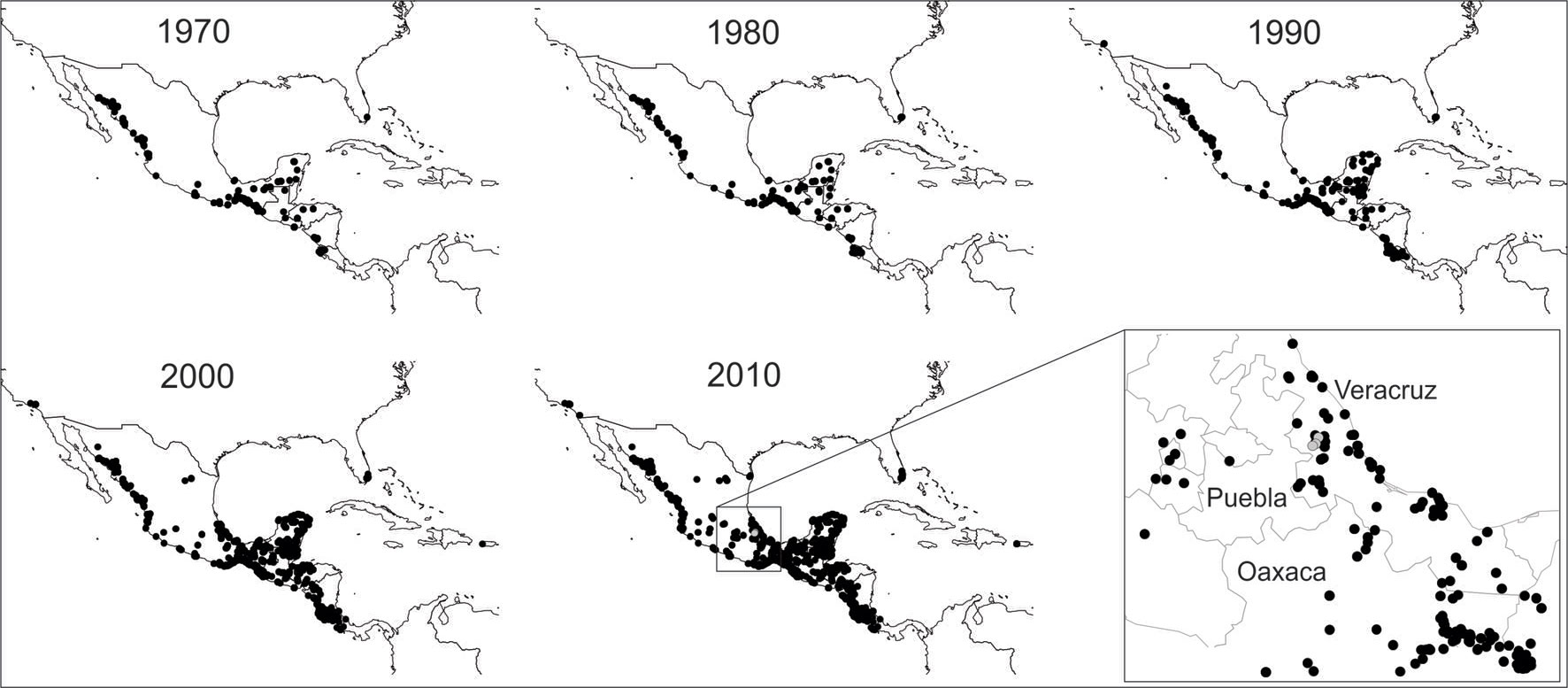
Tabla 1
Variables ambientales utilizadas para generar el perfil climático de A. albifrons y para la elaboración del MDE. Se resaltan en negritas los autovalores que más contribuyeron a cada uno de los 2 primeros componentes.
| Variable | PC 1 | PC 2 |
| 1 = Temperatura media anual | -0.095 | 0.415 |
| 2 = Temperatura promedio del día | -0.258 | -0.108 |
| 3 = Isotermalidad (2/7) | 0.200 | 0.054 |
| 4 = Estacionalidad de la temperatura (desviación estándar) | -0.294 | -0.086 |
| 5 = Temperatura máxima del mes más caluroso | -0.267 | 0.263 |
| 6 = Temperatura mínima del mes más frío | 0.142 | 0.375 |
| 7 = Intervalo de la temperatura anual (5-6) | -0.311 | -0.118 |
| 8 = Temperatura media del cuarto más húmedo | -0.218 | 0.317 |
| 9 = Temperatura media del cuarto más seco | -0.052 | 0.411 |
| 10 = Temperatura media del cuarto más caluroso | -0.215 | 0.338 |
| 11 = Temperatura media del cuarto más frío | 0.060 | 0.406 |
| 12 = Precipitación anual | 0.334 | 0.066 |
| 13 = Precipitación del mes más húmedo | 0.251 | 0.109 |
| 14 = Precipitación del mes más seco | 0.251 | -0.006 |
| 15 = Estacionalidad de la precipitación (coeficiente de variación) | -0.252 | 0.012 |
| 16 = Precipitación del cuarto más húmedo | 0.254 | 0.099 |
| 17 = Precipitación del cuarto más seco | 0.253 | 0.000 |
| 18 = Precipitación del cuarto más caluroso | 0.059 | 0.017 |
| 19 = Precipitación del cuarto más frío | 0.277 | 0.084 |
| % varianza | 35.068 | 28.323 |
| % varianza acumulada | 63.391 |
La identificación de los individuos de la especie se realizó con la ayuda de binoculares y guías de campo (Howell y Webb, 1995), así como por vocalizaciones (Xeno-canto, 2015). Durante las salidas se registró el número de individuos observados, si éstos se encontraban en pareja (ya que existe dimorfismo sexual; Forshaw y Cooper, 1989) y si mostraban conducta reproductiva; es decir, vuelos en pareja, acicalamiento mutuo o alimentación recíproca según lo descrito por Herrera y Díaz-Herrera (2007). Además, observamos y describimos las especies de plantas que estaban consumiendo para determinar si su dieta estaba potencialmente siendo modificada, al compararla con las especies de frutos ingeridos en otras localidades dentro de su distribución histórica conocida (Calvo y Blake, 1998; Gómez-de Silva et al., 2005; Herrera y Herrera-Díaz, 2007; Jordano, 1983; Kantak, 1979; Sánchez-Soto, 2016). Las especies vegetales fueron identificadas mediante la consulta de guías de campo (Benítez et al., 2004; Niembro et al., 2010; Pennington y Sarukhán, 1998) y visitas al Herbario del Instituto de Ecología, A. C. en Xalapa, Veracruz. También se tomó registro fotográfico de los frutos y semillas, y se consultó a expertos para la identificación de plantas de la región (ver agradecimientos).
Los modelos de distribución de especies (MDE) requieren de 2 tipos de datos de entrada, los registros de presencia de la especie y variables o coberturas ambientales; ambos obtenidos en el espacio geográfico. Los MDE se generaron con el algoritmo Maxent Ver. 3.4.1, que se basa en el principio de máxima entropía (Phillips et al., 2006, 2018), que es uno de los algoritmos ampliamente utilizados y que ha demostrado su buen desempeño (Elith et al., 2011). Maxent es un algoritmo correlativo que combina las 2 fuentes de información para reconstruir el nicho ecológico de la especie, que es posteriormente proyectado a la geografía, dando como resultado el modelo de distribución (Mateo et al., 2011; Peterson et al., 2011; Soberón et al., 2017).
Para caracterizar el nicho ecológico de la especie, se utilizaron las 19 coberturas ambientales usadas en los análisis del perfil climático, a las cuales se les aplicó un análisis de correlación de Pearson con el fin de identificar y eliminar a las altamente correlacionadas (> 0.8; Elith et al., 2010; Pearson et al., 1920). Las variables finales seleccionadas para crear los modelos fueron: 1) temperatura media anual, 2) temperatura promedio del día, 3) isotermalidad, 4) temperatura máxima del mes más caluroso, 5) intervalo de la temperatura anual, 6) temperatura media del trimestre más húmedo, 7) precipitación anual, 8) estacionalidad de la precipitación (coeficiente de variación), 9) precipitación del trimestre más húmedo, 10) precipitación del trimestre más seco, 11) precipitación del trimestre más caluroso y 12) precipitación del trimestre más frío.
Para la calibración de los MDE, se requiere definir “a priori” el área de accesibilidad o “M” (sensu Soberón y Peterson, 2005), la cual se delimitó considerando un buffer a partir de los registros pero con base en las provincias biogeográficas definidas por Löwenberg-Neto (2014) y Morrone (2014); es decir, se incluyeron aquellas provincias tanto con registros de presencia, como aquellas que estuviesen cercanas y contiguas a una distancia aproximada de 200 km, ya que potencialmente la especie podría tener acceso dada su capacidad de dispersión. Éstas incluyeron para el occidente, a la provincia de las Tierras Bajas del Pacífico y la Sierra Madre Occidental, para el oriente, a la Provincia Veracruzana y Sierra Madre Oriental, y al Eje Volcánico Transversal y el Balsas en México, hasta la provincia de Gautuso-Salamanca, en Panamá.
Existen métodos que permiten mejorar aparentemente la parametrización en Maxent, como ENMeval, que mejora estadísticamente las combinaciones en los parámetros (Muscarella et al., 2014), o el KuENM (Cobos et al., 2019) que incorpora además criterios biológicos; sin embargo, ambos métodos utilizan al criterio de información de Akaike (AIC), hecho que ha sido recientemente cuestionado (Velasco y González-Salazar, 2019). Por ello, la parametrización de Maxent en este estudio fue la proveniente por defecto, con un valor máximo de 1,000 iteraciones, un límite de convergencia de 0.00001 y el valor del factor de regularización en 1.
Se usó la salida logística para obtener un mapa digital con probabilidades de idoneidad continuas y con valores entre 0 y 1 (Phillips et al., 2006). Para transformar el mapa probabilístico a binario, considerando que los datos provienen de fuentes con distintos de grados de certeza, se estableció un umbral de 10% de omisión en los puntos de entrenamiento, es decir, se estableció como presente (1) a partir del valor de probabilidad correspondiente con la inclusión de 90% de los puntos de entrenamiento y como ausente (0), a los valores menores de ese umbral.
Maxent evalúa automáticamente el desempeño de los modelos a través del cálculo del área bajo la curva (AUC por sus siglas en inglés), de la curva ROC (receiver operating characteristic; Phillips et al., 2006). Sin embargo, debido a que diversos problemas han sido asociados a esta prueba, se usó una modificación llamada ROC parcial, la cual otorga un peso diferencial a los errores de omisión y comisión (Lobo et al., 2008; Peterson et al., 2008). Esta prueba se aplicó usando el programa “Tool for Partial-ROC V. 1.0.” (Barve, 2008) y se consideró un valor de error de omisión menor a 5%.
Resultados
La reconstrucción histórica de la distribución del loro frente blanca inició con la descripción de Amazona albifrons (Psittacus albifrons sensu Sparrman, 1788) y posterior reconocimiento de 3 subespecies: A. a. albifrons sensu Nelson (1899) distribuida desde Nayarit en México, hasta el suroeste de Guatemala; A. a. saltuensis (Nelson, 1899) distribuida desde el sur de Sonora hasta Sinaloa en México; y finalmente, A. a. nana (Miller, 1905) distribuida desde el sureste de Veracruz en México hasta el noroeste de Costa Rica. Desde inicios del siglo pasado, se fueron completando los registros y se fue delineando mediante “mano alzada” el área de su distribución (fig. 1A). A partir de 1970 comenzaron a obtenerse algunos registros geográficamente lejanos de su área de distribución histórica hasta entonces conocida; por ejemplo, en los estados de Florida (Robertson y Woolfenden, 1992; Stevenson y Anderson, 1994) y California (EBIRD, 2016), EUA; así como en Puerto Rico (EBIRD, 2016). En México a partir de 1990, se reportó como una especie exótica en la isla Cozumel, Quintana Roo (Plasencia-Vázquez y Escalona-Segura, 2012), y a partir de la década de los 2000, en algunos estados como Coahuila, Morelos, Ciudad de México (Gómez-de Silva et al., 2005), el norte de Michoacán (Chávez-León, 2007; figs. 3, 4A).
Para el caso específico de Veracruz, el cual es uno de los estados más conocidos del país en cuanto a avifauna se refiere (Navarro-Sigüenza et al., 2008, 2014; Rodríguez-Yáñez et al., 1994), los primeros registros del loro frente blanca provienen del sur desde la década de 1940 (Colección Nacional de Aves, Instituto de Biología de la UNAM, CNAV, No. Cat. 019275 y 019280). Sin embargo, no es sino a partir de la década de 1990 y hasta la actualidad, que se comenzaron a registrar localidades de presencia hacia el centro del estado (De Labra et al., 2010; EBIRD, 2016; González-García, 2006; González-García et al., 2014; Ruelas-Inzunza y Aguilar-Rodríguez, 2010; fig. 4) por lo que con base en el buen estado de conocimiento acumulado de la avifauna en el centro de Veracruz y los relativamente nuevos registros de la especie en la región, sugerimos que corresponden a una colonización reciente.
El análisis de la distribución en el espacio ambiental del loro frente blanca en términos de los 2 primeros componentes principales y cuyas variables más importantes fueron la temperatura y la precipitación media anual en ambos componentes (tabla 1), muestra que los registros novedosos geográficamente (la mayoría de ellos exóticos de acuerdo con EBIRD, 2016; Plasencia-Vázquez y Escalona-Segura, 2012; Robertson y Woolfenden, 1992; Stevenson y Anderson, 1994) no lo son ambientalmente (fig. 4). Por ejemplo, los registros de Florida, EUA (1970), Puerto Rico (2000) e isla Cozumel, México (1990), se encuentran fuera de la distribución geográfica conocida pero dentro de la distribución ambiental ya conocida (fig. 4). Por otro lado, existen registros exóticos (Chávez-León, 2007; EBIRD, 2016; Gómez-de Silva et al., 2005) tanto geográfica como ambientalmente, como los reportados en California, EUA (1990) y en el centro de México (década 2000; fig. 4). Cabe señalar que los nuevos registros encontrados en el centro de Veracruz, los cuales sugerimos que representan una colonización geográfica reciente, corresponden con las condiciones ambientales conocidas para la especie (fig. 3), por lo que resaltamos el hecho de que la colonización es en el espacio geográfico.
El trabajo de campo en las localidades de reciente registro en el centro de Veracruz permitió confirmar la presencia del loro frente blanca durante todo el año, ya que se obtuvo un promedio 88 registros durante el verano, 92 en otoño, 33 en invierno y 25 en primavera.
Esta cantidad de registros y su elevada abundancia relativa a lo largo de todo el año, nos permitió demostrar que es una especie residente que sugiere una reciente colonización en la región. Esta condición se vio reforzada por la frecuente observación, durante el invierno y la primavera, de conductas reproductivas en las parejas registradas. Por otro lado, se registró un total de 14 especies de plantas que son consumidas por el loro frente blanca en diferentes épocas del año en el centro de Veracruz (tabla 2), que, contrastando con la lista de especies consumidas en otras regiones, resulta en el registro de una ingesta de 13 nuevas especies de plantas; es decir, 93% distinta de la dieta previamente conocida.
El análisis de la prueba ROC parcial sobre modelo obtenido, mostró un valor de proporción de AUC de 1.66 (p ˂ 0.001) considerando que los valores van de 0 a 2 y donde 1 es lo esperado si el modelo fuese al azar; lo que sugiere un modelo con un muy buen desempeño. El mapa producto del modelo tiene una extensión de 714,089 pixeles (fig. 5) y describe macroclimáticamente que la distribución geográfica potencial del loro frente blanca incluye las tierras bajas de México, desde Sonora (en la vertiente del Pacífico) y Veracruz (en la vertiente del golfo), cubriendo prácticamente toda la península de Yucatán, hasta el centro-norte de Honduras; a partir de Nicaragua, su distribución se restringe solo a las tierras bajas de la vertiente de la costa del Pacífico hasta el oeste de Panamá (fig. 5).

Tabla 2
Lista de plantas consumidas por A. albifrons. Se presenta la familia, especie, la fuente bibliográfica de referencia, las especies registradas en este estudio (*) y la época en que se observó que la estaban consumiendo: Inv = invierno, Prim = primavera, Ver = verano y Oto = otoño.
| Familia | Especie | Fuente bibliográfica | Inv | Prim | Ver | Oto |
| Anacardiaceae | Manguifera indica | Gómez-de Silva et al. (2005) | ||||
| Anacardiaceae | Metopium browneii | Kantak (1979) | ||||
| Betulaceae | Carpinus caroliniana* | X | X | |||
| Boraginaceae | Ehretia tinifolia | Kantak (1979) | ||||
| Cactaceae | Pachycereus sp. | Gómez-de Silva et al. (2005) | ||||
| Cactaceae | Stenocereus sp. | Gómez-de Silva et al. (2005) | ||||
| Cannabaceae | Trema micrantha* | X | X | |||
| Fabaceae | Leucaena leucodephala* | X | ||||
| Fabaceae | Inga sp.* | Calvo y Blake (1998) | X | X | ||
| Euphorbiaceae | Jatropha curcas | Herrera y Herrera-Díaz (2007) | ||||
| Fabaceae | Acacia sp.* | X | ||||
| Fagaceae | Quercus sp.* | X | X | |||
| Ficus | Ficus sp. | Gómez-de Silva et al. (2005) | ||||
| Loranthaceae | Psittacanthus calyculatus* | X | X | |||
| Moraceae | Ficus continifolia | Jordano (1983) | ||||
| Myrtaceae | Eucalyptus* | X | ||||
| Myrtaceae | Syzygium paniculatum* | X | ||||
| Oxalidaceae | Averrhoa carambola | Sánchez-Soto (2016) | ||||
| Platanaceae | Platanus mexicana* | X | X | |||
| Poaceae | Zea maíz | Gómez-de Silva et al. (2005) | ||||
| Proteaceae | Grevillea robusta* | X | X | |||
| Rasaceae | Eriobotrya japónica* | X | ||||
| Sapindaceae | Talisia olivaeformis | Kantak 1979 | ||||
| Vitaceae | Vitis pepenoei* | X | X | |||
| Vitaceae | Vitis tiliifolia* | X |
Discusión
Cuando el conocimiento sobre la distribución de una especie se basa en décadas de acumulación de registros, los cuales comienzan a describir los mismos patrones geográficos de presencia después de un cierto período; se podría sugerir que el área de distribución de la especie en estudio está relativamente bien conocida (i.e., lo que aquí definimos como distribución histórica conocida). Con base en ello, son de particular notoriedad e importancia, todos aquellos nuevos registros que se realizan fuera de la distribución histórica conocida de una especie, ya que aportan información geográfica que permite sugerir que, a pesar de los esfuerzos históricos, no se tenía en realidad aún un buen conocimiento acerca de su distribución y podrían incluso representar el descubrimiento poblaciones no conocidas. En otro caso, quizá más probable, podría sugerir que la especie se ha enfrentado a modificaciones recientes en su distribución; es decir, potenciales procesos de invasión y/o de colonización, particularmente en aquellas especies cuya distribución histórica, había permanecido sin cambios a pesar de los esfuerzos en su búsqueda. Independientemente de cuál sea el caso, ambos ayudan a mejorar el entendimiento de la biología de la especie (i.e., nicho ecológico, hábitos reproductivos, dieta, etc.) y modificar las prioridades de conservación (Sánchez-González, 2013).
A partir del análisis temporal de la acumulación de los registros del loro frente blanca, se pudo realizar una reconstrucción del área de distribución histórica conocida de la especie y que podría reconocerse como completa a partir de la década de 1970, con base en la descripción de los mismos patrones geográficos de presencia. Este mismo análisis permitió distinguir a partir de esta década, aquellos registros geográficamente novedosos y que correspondieron con registros realizados en áreas fuera de su distribución histórica conocida; es decir, registros geográficamente periféricos que permitieron delimitar mejor el área de distribución del loro frente blanca, llenando los espacios geográficos y ambientales de su distribución (fig. 4).
Por otro lado, también se lograron identificar registros de introducción antropogénica (considerados como exóticos, aunque sus poblaciones se encuentran establecidas en la actualidad) en el centro y norte de México (Chávez-León, 2007; EBIRD, 2016; Gómez-de Silva et al., 2005), y en California, Florida y Puerto Rico (Graham, 1978; Pranty y Epps, 2002; Robertson y Woolfenden, 1992). Estos registros exóticos tienen la particularidad de que se encuentran asociados a centros urbanos y son discontinuos geográficamente respecto a la distribución histórica conocida de la especie; en algunos casos como en México, distantes a 200 km y en otros como EUA y Puerto Rico, a más de 1,000 km.
Se identificaron registros recientes cuyo análisis cronológico y de conocimiento histórico en el centro de Veracruz, sugiere una ampliación natural de su distribución (i.e., colonización) y que no corresponden con registros casuales con base en el trabajo de campo. Estos registros se han realizado en esta región apenas desde la década de 1990 hasta la actualidad (González-García, 2006; González-García et al., 2014; Ruelas-Inzunza y Aguilar-Rodríguez, 2010) y además se puede notar una continuidad geográfica de presencia de la especie desde el sur hasta el centro del estado de Veracruz y que forma parte de la provincia biogeográfica Golfo de México (Morrone, 2014). Aunque estos registros no representan novedades ambientales, sí son novedades geográficas; siendo ésto sobresaliente, ya que provienen de una región muy bien estudiada desde la llegada de los españoles hace más de 500 años (Navarro-Sigüenza et al., 2008, 2014). De hecho, en Veracruz se llevaron a cabo las primeras referencias de algunas especies de aves del país a partir de la llegada de los españoles y ya durante mediados el siglo XX, se publicaron listados relativamente completos en diversas localidades del centro del estado (Díaz-del Castillo, 2005; González-García y Terrazas, 1983; Navarro-Sigüenza et al., 1991).
Las propuestas de área de distribución de A. albifrons de Forshaw y Cooper (1989) y Howell y Webb (1995) que utilizaron el método de mano alzada (aproximación geográfica), difieren de las reconstruidas a partir del uso de MDE, que incluye una aproximación geográfica y ambiental; misma que ha demostrado presentar mejores resultados en comparación con métodos estrictamente geográficos (Mota-Vargas y Rojas-Soto, 2012). Las primeras delimitaciones del área de distribución de esta especie a partir de MDE estuvieron restringidas a México (Moterrubio-Rico, 2016; Navarro-Sigüenza y Peterson, 2007; Ríos-Muños y Navarro-Sigüenza, 2009); es decir, fueron hechas a partir de una representación parcial del nicho ecológico de la especie y considerando solo una parte del área de accesibilidad de la especie (M sensu Soberón y Peterson, 2005), lo que puede sesgar los resultados de los modelos realizados con Maxent, e incluso su validación (Barve et al., 2011). Sin embargo, excluyendo a los registros exóticos, como los de EUA, los del centro y norte de México y los de Puerto Rico, la reconstrucción llevada a cabo en el presente trabajo es una distribución histórica actualizada (desde México hasta Panamá) que incluyó la región centro del estado Veracruz; lo que implica la ampliación de su distribución aproximada de 200 km al norte de la distribución histórica conocida en la vertiente del golfo de México. Si bien no podemos negar la posibilidad de que los registros del centro de Veracruz hayan sido producto de una introducción antropogénica, como se ha documentado en otros sitios, en Veracruz existe una continuidad geográfica y ambiental con relación a la distribución histórica conocida, lo que nos permite sugerir una expansión natural reciente; a diferencia de los registros considerados como exóticos, cuya discontinuidad geográfica es evidente y su introducción antropogénica es más probable y en algunos casos documentada (Chávez-León, 2007; Gómez-de Silva et al., 2005; Graham, 1978; Pranty y Epps, 2002).
Con relación a su reproducción, se observó una conducta reproductiva en diferentes parejas durante el invierno y primavera, a pesar de no haberse detectado nidos durante el trabajo de campo, posiblemente porque son difíciles de observar. Sin embargo, su constante presencia a lo largo del año y su abundancia relativa, evidencian que las poblaciones se han mantenido en la región a lo largo de los 4 años que duró el trabajo de campo y probablemente desde sus primeros registros en la década de 1990 (González-García et al., 2014; Ruelas-Inzunza y Aguilar-Rodríguez, 2010).
Con respecto a los hábitos alimenticios, se detectó un total de 14 especies de plantas que fueron consumidas por el loro frente blanca; una de ellas, del género Inga, ya había sido previamente reportada por Calvo y Blake (1998), pero el resto de las especies no habían sido descritas como parte de su dieta. Esto demuestra que el loro frente blanca está haciendo uso de los recursos disponibles en la región y que ha facilitado su sobrevivencia. Un aspecto de primordial importancia a partir de la riqueza de frutos registrada y de la cual está haciendo uso, es que la especie ya está teniendo probablemente un papel ecológico importante en los bosques de la región, debido a los potenciales efectos en la dispersión y/o depredación de semillas, así como una potencial competencia con el loro corona blanca (Pionus senilis), ya que se observó forrajear en áreas donde también es común esta última especie; esto sin considerar otras potenciales interacciones ecológicas anteriormente inexistentes en la región previas a su expansión (colonización).
Finalmente, consideramos que existe la necesidad de continuar con los inventarios avifaunísticos y de la biodiversidad en general (Peterson et al., 1998, 2004; Remsen, 1995; Rojas-Soto y Oliveras de Ita, 2005; Vuilleumier, 1998); así como de incorporar otras fuentes de información recientes y cada vez más abundantes y accesibles, como las provenientes de la ciencia ciudadana (Conabio, 2016; EBIRD, 2016; Naturalista, 2018); ya que sólo a partir de la integración de toda la información disponible, es que podremos detectar cambios finos en las distribuciones actuales, particularmente ante los ya inminentes cambios presentes y futuros en los patrones de la distribución de la biodiversidad, debidos a la alteración de los ecosistemas por la modificación antropogénica y el cambio climático.
Agradecimientos
A Patricia Téllez-Mendoza y Stéphanie Baltazar-Hernández por su apoyo en el trabajo de campo; a Claudia Gallardo y Francisco Lorea por su apoyo en la identificación de especies de plantas. A Alejandro Salinas, Katherine Renton y Luis A. Sánchez por sus valiosos comentarios durante en la presentación de este trabajo en el XV CECAM 2017. A Alexander Peña, Felipe Toro, Karen Rojas y Keisy Parra por sus valiosos comentarios al manuscrito.
Referencias
Anderson, R. P. y Martínez-Meyer, E. (2004). Modeling species’ geographic distributions for preliminary conservation assessments: an implementation with the spiny pocket mice (Heteromys) of Ecuador. Biological Conservation, 116, 167–179. https://doi.org/10.1016/s0006-3207(03)00187-3
Anderson, R. P. y Raza, A. (2010). The effect of the extent of the study region on GIS models of species geographic distributions and estimates of niche evolution: preliminary tests with montane rodents (genus Nephelomys) in Venezuela. Journal of Biogeography, 37, 1378–1393. https://doi.org/10.1111/j.1365-2699.2010.02290.x
Antúnez, A. y Mendoza, M. (1992). Factores que determinan el área de distribución geográfica de las especies: conceptos, modelos y métodos de análisis. En J. M. Vargas, R. Real y A. Antúnez (Eds.), Objetivos y métodos biogeográficos aplicaciones en herpetologı́a (pp. 51–72). Madrid: Asociación Herpetológica Española.
Barve, N. (2008). Tool for Partial-ROC (Biodiversity Institute, Lawrence, KS), ver 1.0.
Barve, N., Barve, V., Jiménez-Valverde, A., Lira-Noriega, A., Maher, S. P., Peterson, A. T. et al. (2011). The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecological Modelling, 222, 1810–1819. https://doi.org/10.1016/j.ecolmodel.2011.02.011
Benítez, G., Pulido-Salas, M. T. y Equihua, M. (2004). Arboles multiusos nativos de Veracruz para reforestación, restauración y plantaciones. Xalapa: INECOL, A.C.
Calvo, L. y Blake, J. (1998). Bird diversity and abundance on two different shade coffee plantations in Guatemala. Bird Conservation International, 8, 297–308. https://doi.org/10.1017/s0959270900001945
Chávez-León, G. (2007). Riqueza de aves del parque nacional Barranca del Cupatitzio, Michoacán, México. Acta Zoológica Mexicana, 23, 11–29. https://doi.org/10.21829/azm.2007.232567
Cobos, M. E., Peterson, A. T., Barve, N. y Osorio-Olvera, L. (2019). Kuenm: an R package for detailed development of ecological niche models using Maxent. PeerJ, 7, e6281. https://doi.org/10.7717/peerj.6281
Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad). (2016). Base de datos digital de Amazona albifrons. Recuperado en enero 2016: http://www.enciclovida.mx
De Labra, E., Escalante, P., Monterrubio, T. C. y Coates-Estrada, R. (2010). Hábitat, abundancia y perspectivas en conservación de Psittacidos en la reserva de Los Tuxtlas, Veracruz, México. Ornitología Neotropical, 21, 599–610.
Díaz-del Castillo, B. (2005). Historia verdadera de la conquista de la Nueva España (manuscrito Guatemala), edición crítica de José Antonio Barbón Rodríguez. México D.F., Madrid: El Colegio de México/ Agencia Española de Cooperación Internacional.
EBIRD. (2016). Laboratorio de Ornitología de la Universidad de Cornell. Recuperado en enero 2016: https://ebird.org
Elith, J., Kearney, M. y Phillips, S. (2010). The art of modelling range-shifting species. Methods in Ecology and Evolution, 1, 330–342. https://doi.org/10.1111/j.2041-210x.2010.00036.x
Elith, J., Phillips, S. J., Hastie, T., Dudík, M., Chee, Y. E. y Yates, C. J. (2011). A statistical explanation of MaxEnt for ecologists. Diversity and distributions, 17, 43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x
ESRI (Environmental Systems Research Institute). (1999). ArcView GIS, V.3.2. CA, USA, ESRI Inc.
Forshaw, J. M. y Cooper, W. T. (1989). Parrots of the world. London: Blandford.
GBIF (Global Biodiversity Information Facility). (2017). Recuperado en enero 2017. http://doi.org/10.15468/dl.m8qops
Gómez-de Silva, H., Oliveras de Ita, A. y Medellín, R. A. (2005). Amazona albifrons. Vertebrados superiores exóticos en México: diversidad, distribución y efectos potenciales. México D.F.: INECOL, A.C./ UNAM. Bases de datos SNIB-Conabio. Proyecto U020.
González-García, F. (2006). Las aves. En P. Moreno-Casasola (Ed.), Entornos veracruzanos: la costa de La Mancha (pp. 423–447). Xalapa: INECOL, A.C.
González-García, F. y Terrazas, T. (1983). Guía las aves de Xalapa, Veracruz. Xalapa: Instituto Nacional de Investigaciones sobre Recursos Bióticos y Secretaría de Desarrollo Urbano y Ecología.
González-García, F., Straub, R., Lobato-García, J. A. y MacGregor-Fors, I. (2014). Birds of a neotropical green city: an up-to-date review of the avifauna of the city of Xalapa with additional unpublished records. Urban Ecosystem, 17, 991–1012. https://doi.org/10.1007/s11252-014-0370-3
Graham, C. L. G. (1978). Poxvirus infection in a spectacled amazon parrot (Amazona albifrons). American Association of Avian Pathologists, 22, 340–343. https://doi.org/10.2307/1589547
Hammer, Ø., Harper, D. A. y Ryan, P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4, 1–9.
Hanna, Z. R., Ortiz-Ramírez, M. F., Ríos-Muñoz, C. A., Cayetano-Rosas, H., Bowie, R. C. y Navarro-Sigüenza, A. G. (2016). Phylogenetic and morphologic evidence confirm the presence of a new montane cloud forest associated bird species in Mexico, the Mountain Elaenia (Elaenia frantzii; Aves: Passeriformes: Tyrannidae). PeerJ, 4, e1598. https://doi.org/10.7717/peerj.1598
Herrera, N. y Díaz-Herrera, A. (2007). Biología reproductiva de la cotorra frente blanca (Amazona albifrons) en Barra de Santiago, El Salvador. Sociedad Mesoamericana para la Biología y la Conservación, 11, 82–89.
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G. y Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, 1965–1978. https://doi.org/10.1002/joc.1276
Howell, S. N. G. y Webb, S. (1995). A guide to the Birds of Mexico and Northern Central America. Oxford: Oxford University Press.
Jordano, P. (1983). Fig-seed predation and dispersal by birds. Biotropica, 15, 38–41. https://doi.org/10.2307/2387996
Kantak, G. E. (1979). Observations on some fruit-eating birds in Mexico. Auk, 96, 183–86.
Lobo, J. M., Jiménez-Valverde, A. y Real, R. (2008). AUC: a misleading measure of the performance of predictive distribution models. Global Ecology and Biogeography, 17, 141–151. https://doi.org/10.1111/j.1466-8238.2007.00358.x
Löwenberg-Neto, P. (2014). Neotropical region: a shapefile of Morrone’s (2014) biogeographical regionalization. Zootaxa, 3802, 300. https://doi.org/10.11646/zootaxa.3802.2.12
Marín-Togo, M. C., Monterrubio-Rico, T. C., Renton, K., Rubio-Rocha, Y., Macías-Caballero, C., Ortega-Rodríguez, J. M. et al. (2012). Reduced current distribution of Psittacidae on the Mexican Pacific coast: potential impacts of habitat loss and capture for trade. Biodiversity and Conservation, 21, 451–473. https://doi.org/10.1007/s10531-011-0193-y
Mateo, R. G., Felicísimo, Á. M. y Muñoz, J. (2011). Modelos de distribución de especies: una revisión sintética. Revista chilena de historia natural, 84, 217–240. https://doi.org/10.4067/s0716-078×2011000200008
Miller, W. W. (1905). List of bird collected in southern Sinaloa, Mexico, by J. H. Batty, during 1903-1904. Bulleting American Museum of Natural History, 21, 339–369.
Monrroy-Ojeda, A. e Isern, S. G. (2013). Noteworthy bird records in Sontecomapan, Veracruz, México. Acta Zoológica Mexicana, 29, 666–676.
Monterrubio-Rico, T. C., Charre-Medellín, J. F., Pacheco-Figueroa, C., Arriaga-Weiss, S., Valdez-Leal, J. D., Cancino-Murillo, R. et al. (2016). Distribución potencial histórica y contemporánea de la familia Psittacidae en México. Revista Mexicana de Biodiversidad, 87, 1103–117. https://doi.org/10.1016/j.rmb.2016.06.004
Morrone, J. J. (2001). Sistemática, Biogeografía, Evolución: los patrones de la biodiversidad en espacio-tiempo. México D.F.: Universidad Nacional Autónoma de México.
Morrone, J. J. (2014). Biogeographical regionalization of the Neo-tropical region. Zootaxa. 3782, 1–110.
Mota-Vargas, C. y Rojas-Soto, O. R. (2012). The importance of defining the geographic distribution of species for conservation: The case of the Bearded Wood-Partridge. Journal for Nature Conservation, 20, 10–17. https://doi.org/10.1016/j.jnc.2011.07.002
Muscarella, R., Galante, P. J., Soley-Guardia, M., Boria, J. M., Kass, R. A., Uriarte, M. et al. (2014). ENMeval: an R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods in Ecology and Evolution, 5, 1198–1205. https://doi.org/10.1111/2041-210x.12261
Naturalista. (2018). Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Recuperado en enero 2018: http://www.naturalista.mx/
Navarro-Sigüenza, A. G. y Peterson, A. T. (2007). Amazona albifrons (loro frente-blanca) residencia permanente. Distribución potencial. Catálogo de metadatos geográficos. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Recuperado en enero 2017: http://www.conabio.gob.mx
Navarro-Sigüenza, A. G., Morales-Pérez, J. E. y Hernández-Baños, B. E. (1991). Aves de Teocelo. Teocelo, 7/8, 11–34.
Navarro-Sigüenza, A. G., Ortiz-Pulido, R. y Peterson, A. T. (2008). Un panorama breve de la historia de la Ornitología mexicana. Ornitología Neotropical, 19, 367–379.
Navarro-Sigüenza, A. G., Rebón-Gallardo, M. F., Gordillo-Martínez, A., Peterson, A. T., Berlanga-García, H. y Sánchez-González, L. A. (2014). Biodiversidad de aves en México. Revista Mexicana de Biodiversidad, 85, 476–495. https://doi.org/10.7550/rmb.41882
Nelson, E. W. (1899). Descriptions of new birds from northwestern Mexico. Proceedings of the Biological Society of Washington, 13, 25–31.
Niembro, A., Vázquez, M. y Sánchez, O. (2010). Árboles de Veracruz. 100 especies para la reforestación estratégica. México D.F.: Gobierno del Estado de Veracruz/ Secretaría de Educación de Veracruz/ Centro de Investigaciones Tropicales/ Universidad Veracruzana.
Pearson, K. (1920). Notes on the history of correlation. Biometrika, 13, 25–45. https://doi.org/10.1093/biomet/13.1.25
Pennington, T. D. y Sarukhán, J. (1998). Árboles tropicales de México. Manual para la identificación de las principales especies. México D.F.: Fondo de Cultura Económica.
Peterson, A. T. (2001). Predicting species’ geographic distributions based on ecological niche modelling. The Condor, 103, 599–605. https://doi.org/10.1650/0010-5422(2001)103[0599:psgdbo]2.0.co;2
Peterson, A. T., Navarro-Sigüenza, A. y Benítez-Díaz, H. (1998). The need for continued scientific collecting: a geographic analysis of Mexican bird specimens. Ibis, 140, 288–294. https://doi.org/10.1111/j.1474-919x.1998.tb04391.x
Peterson, A. T., Navarro-Sigüenza, A. y Scachetii-Pereira, R. (2004). Detecting errors in biodiversity data on collectors’ itineraries. Bulletin of the British Ornithological Club, 124, 143–151.
Peterson, A. T., Papes, M. y Soberón, J. (2008). Rethinking Receiver operating characteristic analysis applications in ecological niche modeling. Ecological Modelling, 213, 63–72. https://doi.org/10.1016/j.ecolmodel.2007.11.008
Peterson, A. T., Soberón, J., Pearson, R. G., Anderson, R. P., Martínez-Meyer, E., Nakamura, M. et al. (2011). Ecological niches and geographic distributions. Princeton, NJ: Princeton University Press.
Peterson, A. T., Navarro-Sigüenza, A. G. y Gordillo, A. (2018). Assumption-versus data-based approaches to summarizing species’ ranges. Conservation Biology, 32, 568–575. https://doi.org/10.1111/cobi.12801
Phillips, S. J., Dudík, M. y Schapire, R. E. (2018). Maxent software for modeling species niches and distributions, Version 3.4.1. Recuperado en enero 2018: http://biodiversityinformatics.amnh.org/open_source/maxent/
Phillips, S. J., Anderson, R. P. y Schapire, R. E. (2006). Maximum entropy modelling of species geographic distributions. Ecological Modelling, 190, 231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Plasencia-Vázquez, A. H. y Escalona-Segura, G. (2012). Nuevo registro del loro frente blanca (A. albifrons) para la Isla Cozumel, Quintana Roo, México. Huitzil, 13, 43–46.
Pranty, B. y Epps, S. (2002). Distribution, population, status, and documentation of exotic parrots in Broward County Florida. Florida Field Naturalist, 30, 111–131.
Rappoport, E. H. (1975). Aerografía: estrategias geográficas de las especies. México: Fondo de Cultura Económica.
Remsen, J. V. Jr. (1995). The importance of continued collecting of bird specimens to ornithology and bird conservation. Bird Conservation International, 5, 145–180. https://doi.org/10.1017/s095927090000099x
Ríos-Muñoz, C. A. (2013). ¿Es posible reconocer una unidad biótica entre América del Norte y del Sur? Revista Mexicana de Biodiversidad, 84, 1022–1030. https://doi.org/10.7550/rmb.34170
Ríos-Muñoz, C. A. y Navarro-Sigüenza. A. G. (2009). Efectos del cambio de uso de suelo en la disponibilidad hipotética de hábitat para los psitácidos de México. Ornitología Neotropical, 20, 491–509.
Robertson, W. B. y Woolfenden, G. E. (1992). Florida bird species: an annotated list. Gainesville: Florida Ornithological Society.
Rodríguez-Yáñez, C., Villalón, R. M. y Navarro-Sigüenza, A. G. (1994). Bibliografía de las aves de México (1825-1992). México D.F.: Publicaciones Especiales del Museo de Zoología, Núm. 8., UNAM.
Rojas-Soto, O. y Oliveras de Ita, A. (2005). Los inventarios avifaunísticos: reflexiones sobre su desarrollo en el neotrópico. Ornitología Neotropical, 16, 441–445.
Ruelas-Inzunza, E. y Aguilar-Rodríguez, S. H. (2010). La avifauna urbana del Parque Ecológico Macuiltépetl en Xalapa, Veracruz, México. Ornitología Neotropical, 21, 87–103.
Sánchez-González, L. A. (2013). Cuando un “nuevo registro” es realmente un nuevo registro: consideraciones para su publicación. Huitzil, 14, 17–21.
Sánchez-Soto, S. (2016). Informe de frutos de carambola dañados por Amazona albifrons Sparrman, en Tabasco, México. Agronomía Mesoamericana, 27, 415–419. https://doi.org/10.15517/am.v27i2.21282
Soberón, J. y Peterson, A. T. (2005). Interpretation of models of fundamental ecological niches and species’ distributional areas. Biodiversity Informatics, 2, 1–10. https://doi.org/10.17161/bi.v2i0.4
Soberón, J., Osorio-Olvera, L. y Peterson, T. (2017). Diferencias conceptuales entre modelación de nichos y modelación de áreas de distribución. Revista Mexicana de Biodiversidad, 88, 437–441. https://doi.org/10.1016/j.rmb.2017.03.011
Sparrman, A. (1788). Novas et selectas aves. Fasículo III. Estocolmo: Museum Carlsonianum.
Stevenson, H. M. y Anderson, B. H. (1994). The birdlife of Florida. Gainesville: University Press of Florida.
Stockwell, D. R. B. y Peters, D. P. (1999). The GARP modeling system: problems and solutions to automated spatial prediction. International Journal of Geographic Information Systems, 13, 143–158. https://doi.org/10.1080/136588199241391
Velasco, J. y González-Salazar, C. (2019). Akaike information criterion should not be a “test” of geographical prediction accuracy in ecological niche modelling. Ecological Informatics, 51, 25–32. https://doi.org/10.1016/j.ecoinf.2019.02.005
Vuilleumier, F. (1998). The need to collect birds in the Neotropics. Ornitología Neotropical, 9, 201–203.
Wisz, M. S., Hijmans, R. J., Li, J., Peterson, A. T., Graham, C. H., Guisan, A. et al. (2008). Effects of sample size on the performance of species distribution models. Diversity and Distributions, 14, 763–773. https://doi.org/10.1016/s0304-3800(01)00388-x
Xeno-canto. (2015). Compartiendo cantos de aves de todo el mundo. Recuperado en febrero 2015: http://www.xeno-canto.org
Zunino, M. y Zullini, A. (2003). Biogeografía, la dimensión espacial de la evolución. México D.F.: Fondo de Cultura Económica.


