La macroecología en México: historia, avances y perspectivas
Pilar Rodríguez(a)⁎ ✉ , Fabricio Villalobos(b), Alejandro Sánchez-Barradas(b), María Eugenia Correa-Cano(c)
Resumen
La macroecología surgió a finales de la década de 1980 y se definió como un programa de investigación enfocado a describir y entender los patrones de biodiversidad en escalas espaciales y temporales amplias. En esta revisión identificamos: a) las principales aportaciones teórico-conceptuales y prácticas, desarrolladas por autores mexicanos; b) los vacíos de conocimiento y desafíos teórico-metodológicos, y c) perspectivas en este campo de estudio en México. A través de búsquedas en la base de datos de artículos científicos registrados en la Web of Science y de búsquedas dirigidas, encontramos 163 artículos, la mayoría publicados en la última década. Un elevado porcentaje de los trabajos se desarrollaron a una escala regional (37%) y nacional (31%) y se concentraron en el medio terrestre (90%). Los grupos biológicos más estudiados fueron los mamíferos (33%), las plantas vasculares (21%) y las aves (17%). Los temas más estudiados en México fueron los patrones de diversidad, aspectos del nicho ecológico, el análisis de la diversidad a distintas escalas (alfa, beta y gamma) y el área de distribución geográfica de las especies. Es importante incorporar aspectos de abundancia y de diversidad filogenética y funcional que permitan incrementar la capacidad explicativa y predictiva de la macroecología.
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Palabras clave:
Abundancia de especies; Área de distribución; Diversidad alfa; Diversidad beta; Escalas; Nicho ecológico; Procesos; Tamaño corporal;
Macroecology in Mexico: history, progress and perspectives
Abstract
Macroecology emerged at the end of the 1980’s as a synthetic discipline aimed at describing and understanding biodiversity patterns at large spatial and temporal scales. Here, we review the state of the art of macroecology in Mexico by identifying: (i) the main theoretical and applied contributions made by Mexican researchers; (ii) knowledge gaps and theoretic-methodological challenges, and (iii) perspectives of this research field in Mexico. After conducting systematic searches in the Web of Science database and targeted specific topic searches we found 163 articles published by Mexican scientists, most of them in the last decade. Thirty seven percent of these papers were focused at a regional scale and 31% at a national scale, and most of them (90%) corresponded to terrestrial habitats. Mammals, plants and birds were the groups better represented (33%; 21%, and 17%, respectively). The main contributions were developed in the study of diversity patterns, ecological niche, analyses of diversity at different scales (alpha, beta and gamma diversity), and the distributional area. It is important to incorporate subjects such as abundance, phylogenetic and functional diversity and transform the discipline into a more predictive one able to contribute in solving environmental issues at national and global scales.
Keywords:
Abundance of species; Geographical range size; Alpha diversity; Beta diversity; Scales; Ecological niche; Processes; Body size;
Introducción
Entender las causas que explican la gran variedad de especies y su distribución en la Tierra es una de las preguntas centrales de la ecología y la biogeografía. Dichas causas son múltiples y pueden interactuar entre sí de manera compleja, generando diferentes respuestas expresadas en la organización de las comunidades ( Brown, 1995; Lawton, 1999 ). Considerando explícitamente esta naturaleza compleja de la biodiversidad, surge a finales de la década de los años ochenta del siglo xx un enfoque integrativo llamado «macroecología», que busca describir y entender los patrones de biodiversidad en amplias escalas espaciales y temporales ( Brown y Maurer, 1989 ). La macroecología puede definirse como un programa de investigación enfocado en las propiedades estadísticas que emergen de sistemas ecológicos complejos ( Marquet, 2009).El término «macroecología» aparece por primera vez en la literatura científica en 1971 ( Sarmiento y Monasterio, 1971; «descubierto» por Marquet, 2009 ), haciendo referencia a patrones de vegetación en escalas espaciales amplias. Las bases de la macroecología pueden encontrarse desde los trabajos de los naturalistas del siglo xvii (e.g., Johann Reinhold Foster) y siglo xix (e.g., Alexander von Humboldt), hasta los trabajos de zoogeografía y fitogeografía de los años 50 y 60 del siglo xx , centrados en entender la distribución de los organismos en el espacio geográfico. También se encuentran en la geografía ecológica de MacArthur (1972) (en Brown, 1999 ), que trataba de explicar en términos climáticos o ecológicos la distribución de los seres vivos. En el contexto latinoamericano destraca el aporte de Rapoport (1975) , cuyo libro Areografía: estrategias geográficas de las especies (que aparece publicado en inglés en 1982; Rapoport, 1982 ) constituyó una pieza clave en el desarrollo de la macroecología ( Brown, Stevens y Kaufman, 1996; Stevens, 1989 ).
La formalización de la macroecología propuesta por Brown y Maurer (1987, 1989) se centró en la integración de la diversidad, la abundancia y la distribución de las especies bajo una investigación estadística, no experimental, enfocada en la identificación de regularidades (i.e., patrones) que emergen al estudiar conjuntos de especies ( Brown, 1995; Marquet, 2009 ). Una vez establecidas sus bases formales y la lógica detrás de su enfoque ( Blackburn, 2004 ), la macroecología pasó de ser una propuesta integradora a ser una disciplina relevante en la investigación ecológica, como lo confirman la cantidad de revistas especializadas en el tema y su impacto ( Beck et al., 2012 ). Asimismo, las herramientas conceptuales y metodológicas se han refinado y expandido ( Beck et al., 2012; Keith et al., 2012 ). Como resultado, la macroecología ahora incluye enfoques de modelación estocástica y mecanística (e.g., modelos computacionales de simulación orientados a patrones y basados en procesos), además de su enfoque descriptivo y correlativo inicial ( Gotelli et al., 2009; Villalobos y Rangel, 2014 ).
El inicio de la macroecología en México está marcado por 3 hechos. En primer lugar, la estancia de E. H. Rapoport en nuestro país a mediados de la década de 1970, lugar donde es publicado su libro, que contiene ideas fundamentales acerca de la distribución de las especies ( Rapoport, 1975 ). En segundo lugar, los trabajos que se iniciaron en la década de 1980, centrados en el estudio de los patrones de riqueza de especies ( Ezcurra y Equihua, 1984; citas que se encuentra en Ramamoorthy, Bye, Lot y Fa, 1993 ). Si bien estos trabajos no fueron realizados utilizando un enfoque macroecológico propiamente dicho, constituyen una de las bases de los estudios que se realizaron posteriormente. En tercer lugar, en esa época, con la llegada al país de científicos mexicanos que hicieron sus estudios de posgrado en el extranjero se implementaron enfoques y aproximaciones al estudio de los patrones de diversidad provenientes de la macroecología ( Arita, Figueroa, Frisch, Rodríguez y Santos-del Prado, 1997; Ceballos y Navarro, 1991; Mourelle y Ezcurra, 1997a, 1997b ). La macroecología desarrollada en México también tiene aportes provenientes de la biogeografía. Por ejemplo, G. Halffter ha sido un impulsor de la idea de estudiar patrones geográficos de diversidad y de separar los componentes de la diversidad en alfa (riqueza local de especies), beta (diferencia en la composición de especies entre localidades) y gamma (riqueza de especies de una región), un enfoque que aplica a distintas escalas espaciales y que está estrechamente relacionado con la conservación de la biodiversidad ( Halffter y Moreno, 2005).
Un factor que también ha sido importante en el desarrollo de la macroecología en México es la disponibilidad de datos e información que existen para el país, una infraestructura poco frecuente en el mundo ( Sarukhán y Jiménez, 2016 ). Con la creación de la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio) en 1992 se dio impulso a la integración de datos primarios (registros de colectas) en bases de datos estandarizadas y se apoyaron proyectos para generar información a partir de estos datos primarios, como son las áreas de distribución de las especies. Los datos primarios, accesibles a través de la Conabio, la Unidad de Informática para la Biodiversidad (Unibio) y la Global Biodiversity Information Facility (GBIF), así como las áreas de distribución generadas a partir de estos datos primarios, han permitido la realización de numerosos estudios macroecológicos en México ( Koleff et al., 2008; Peterson et al., 1993; Peterson, Soberón y Sánchez-Cordero, 1999 ).
A pesar de la relevancia teórica y aplicada que tiene la macroecología en temas de biodiversidad y conservación y del desarrollo que ha tenido en las últimas 4 décadas, no existe un trabajo que sintetice el aporte de México a esta disciplina. En este trabajo revisamos cuáles son los temas que se han estudiado en México, identificamos las principales aportaciones teórico-conceptuales y prácticas desarrolladas por autores mexicanos, identificamos los vacíos importantes y concluimos discutiendo los desafíos teórico-metodológicos de la macroecología en el país.
La macroecología en México: algunas estadísticas generales
Para evaluar el desarrollo de la macroecología en México, en relación con el contexto global de esta disciplina, en primer lugar realizamos una búsqueda de publicaciones generadas a nivel mundial. Para determinar el número de publicaciones por año se siguió la propuesta de Beck et al. (2012) , usando un periodo de búsqueda desde 1989 (cuando aparece el primer artículo formal de macroecología; Brown y Maurer, 1989 ) hasta el 11 de julio de 2016. Utilizamos la base de datos de artículos científicos de Web of Science ( https://webofknowledge.com; consultada 11 Jul 2016; ver Material suplementario 1 ) y se encontraron 1,516 artículos.
Para determinar el número de publicaciones generadas por autores mexicanos, inicialmente se siguió la propuesta de Beck et al. (2012) . Sin embargo, debido a que la búsqueda arrojó únicamente 27 artículos, se realizaron búsquedas por temas más específicos (tamaño corporal, regla de Rapoport, etc.). Con este procedimiento, y utilizando distintas combinaciones de criterios (ver Material suplementario 1 ), encontramos 70 artículos; por lo tanto, decidimos complementar la búsqueda realizada en la base de datos Web of Science con búsquedas dirigidas, por ejemplo, utilizando el nombre de investigadores mexicanos que sabemos que trabajan en los distintos temas de macroecología. En total se encontraron 163 trabajos realizados por autores mexicanos.
A diferencia del aumento gradual en el número de artículos que se observó a nivel mundial, en México ha habido una gran variación temporal, como por ejemplo la baja significativa en el número de artículos publicados en el año 2010. Tanto a nivel mundial como en México, un porcentaje muy elevado de los artículos (96 y 88%, respectivamente) fueron publicados a partir del año 2000 ( fig. 1 ). Los grupos biológicos más estudiados en México fueron los mamíferos (n = 58 estudios, 33%), seguido de las plantas vasculares (n = 37, 21%) y de las aves (n = 31, 17%). A nivel mundial, los grupos más estudiados son los mismos, pero la proporción de trabajos para cada grupo fue distinta: plantas (24%), aves (19%) y mamíferos (17%) ( fig. 2 ). En relación con el tipo de hábitat, los estudios en México se concentran notoriamente en el medio terrestre (90%), una tendencia distinta a la que se encontró a nivel mundial, donde los estudios se enfocan en el medio terrestre y marino (56 y 35%, respectivamente) ( fig. 3 ). La escala geográfica que preferentemente abarcan los estudios en México es la regional (i.e., Baja California, desierto de Chihuahua) (n = 56, 37%), seguida de la escala a nivel del país (n = 48, 31%). Una proporción importante de los estudios abarcan una escala mundial (n = 14, 9%) y una proporción muy similar abarca América del Norte y Centroamérica (n = 13, 9%) (fig. 4 ). Finalmente, los temas preferentemente estudiados en México son el análisis de los patrones geográficos de diversidad, los estudios relacionados con el nicho ecológico de las especies, el estudio de los componentes alfa, beta y gamma y los análisis relacionados con el área de distribución de las especies ( fig. 5).
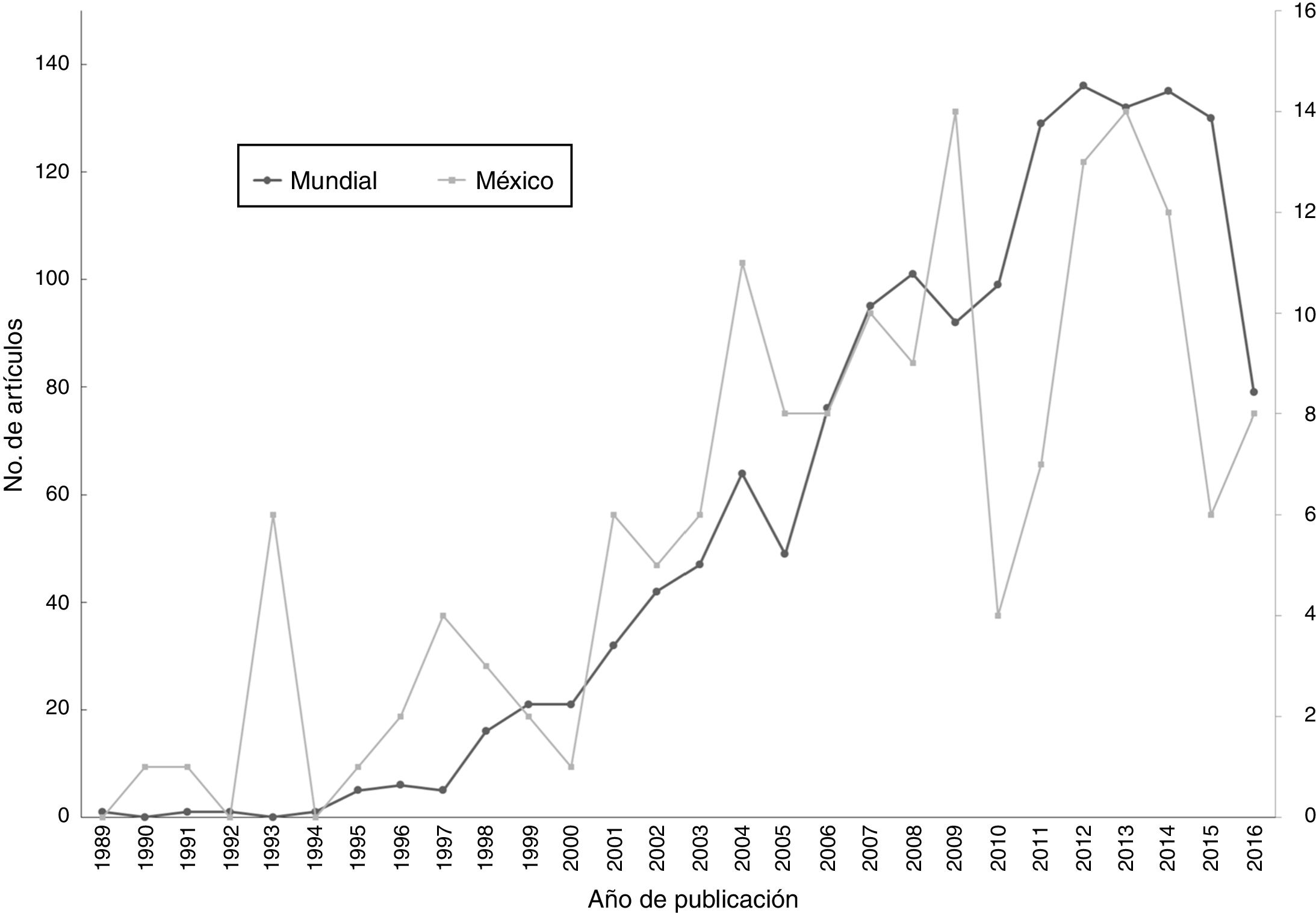
Figura 1
Número total de artículos científicos sobre macroecología producidos entre 1989 y 2016, resultado de la búsqueda realizada a nivel mundial (n = 1,516), y resultados de la búsqueda realizada para México (es decir, en los que al menos un autor es de nacionalidad mexicana) (n = 163). Se utilizaron dos escalas distintas con la idea de comparar las trayectorias.
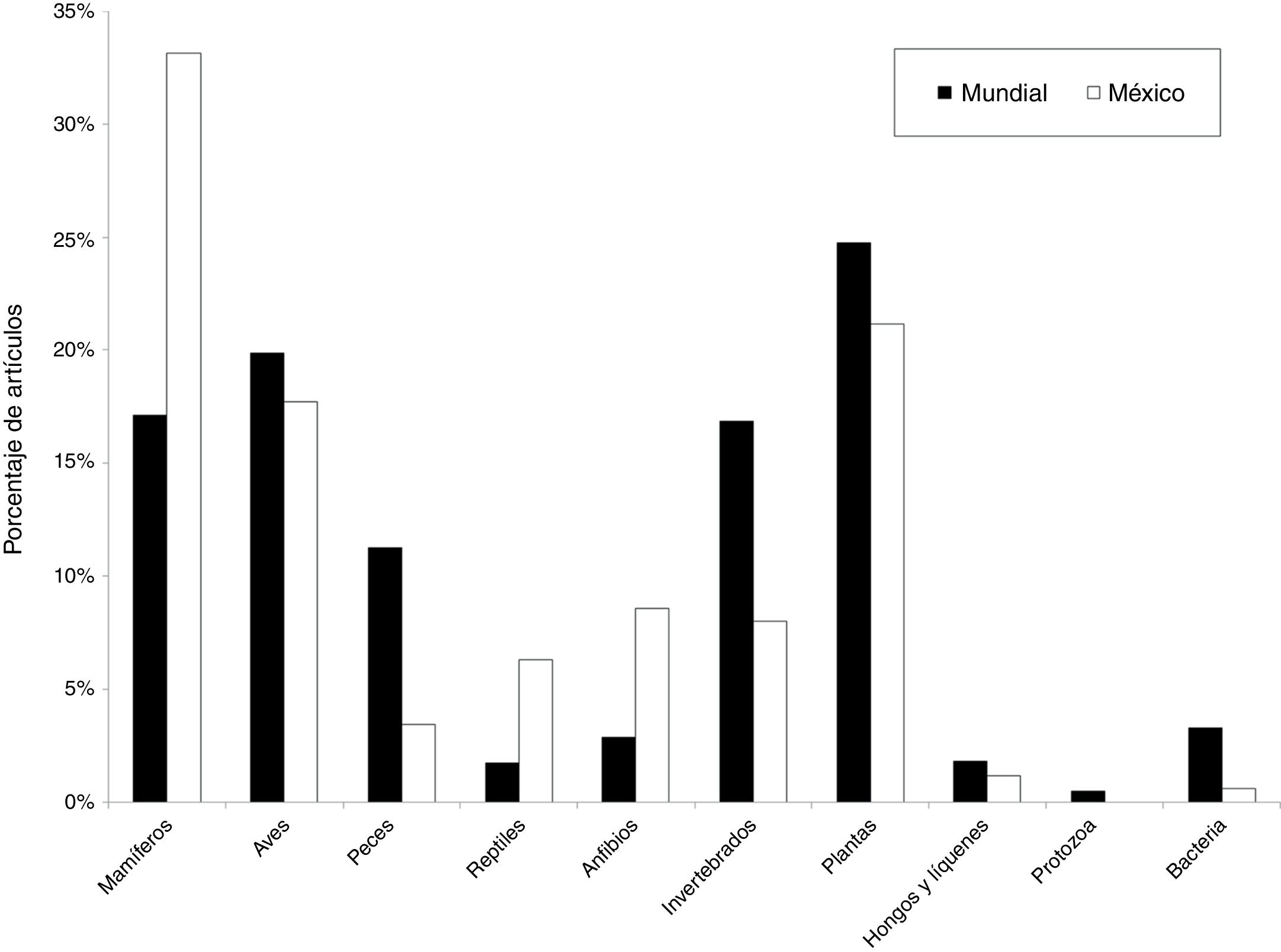
Figura 2
Porcentaje del total de artículos científicos producidos entre 1989 y 2016 por grupo taxonómico a nivel mundial (n = 1,516) y para México (es decir, en los que al menos un autor es de nacionalidad mexicana) (n = 163).
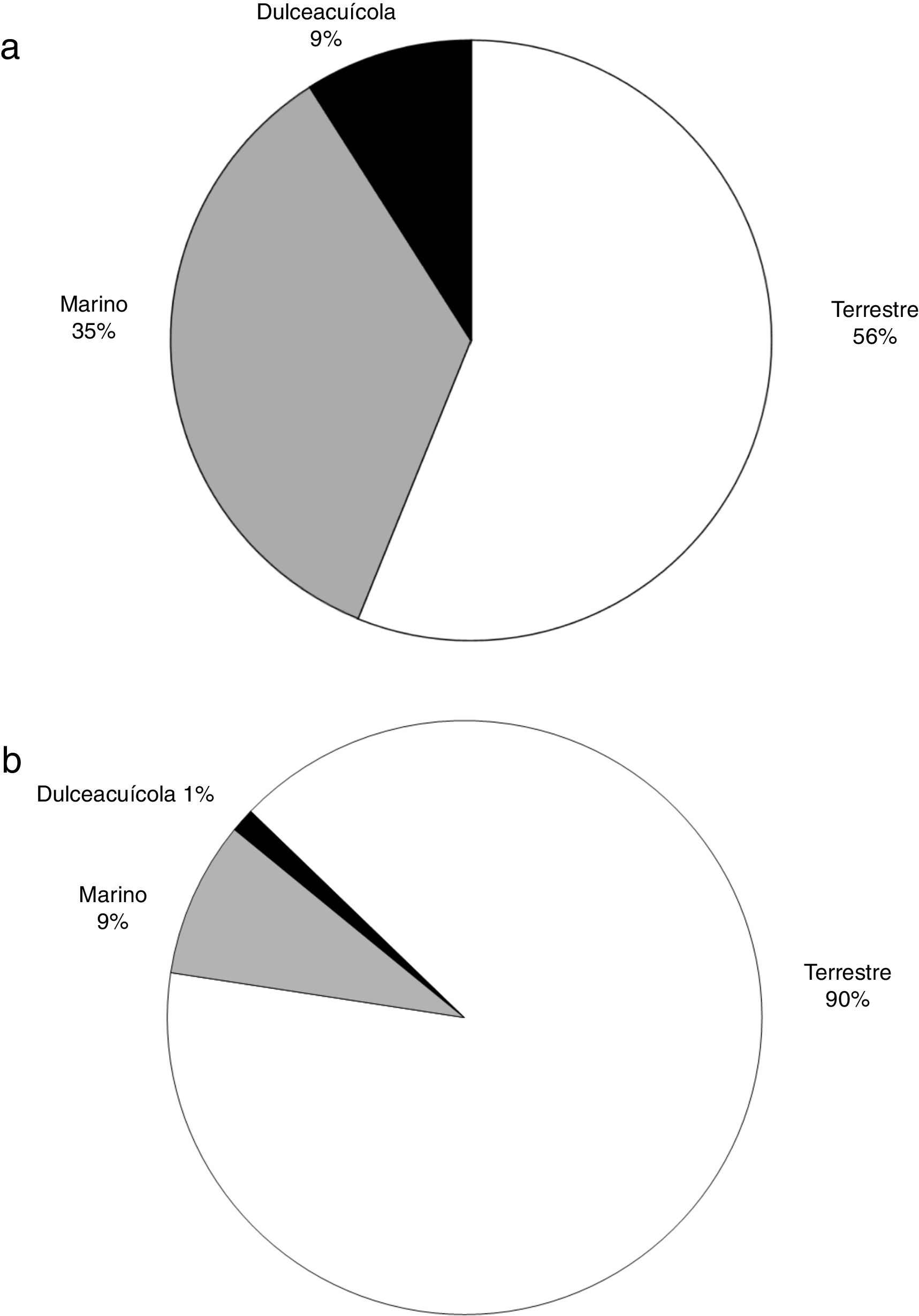
Figura 3
Porcentaje del total de artículos científicos producidos entre 1989 y 2016 por tipo de hábitat a nivel mundial (a) (n = 1,516) y para México (es decir, en los que al menos un autor es de nacionalidad mexicana) (b) (n = 163).
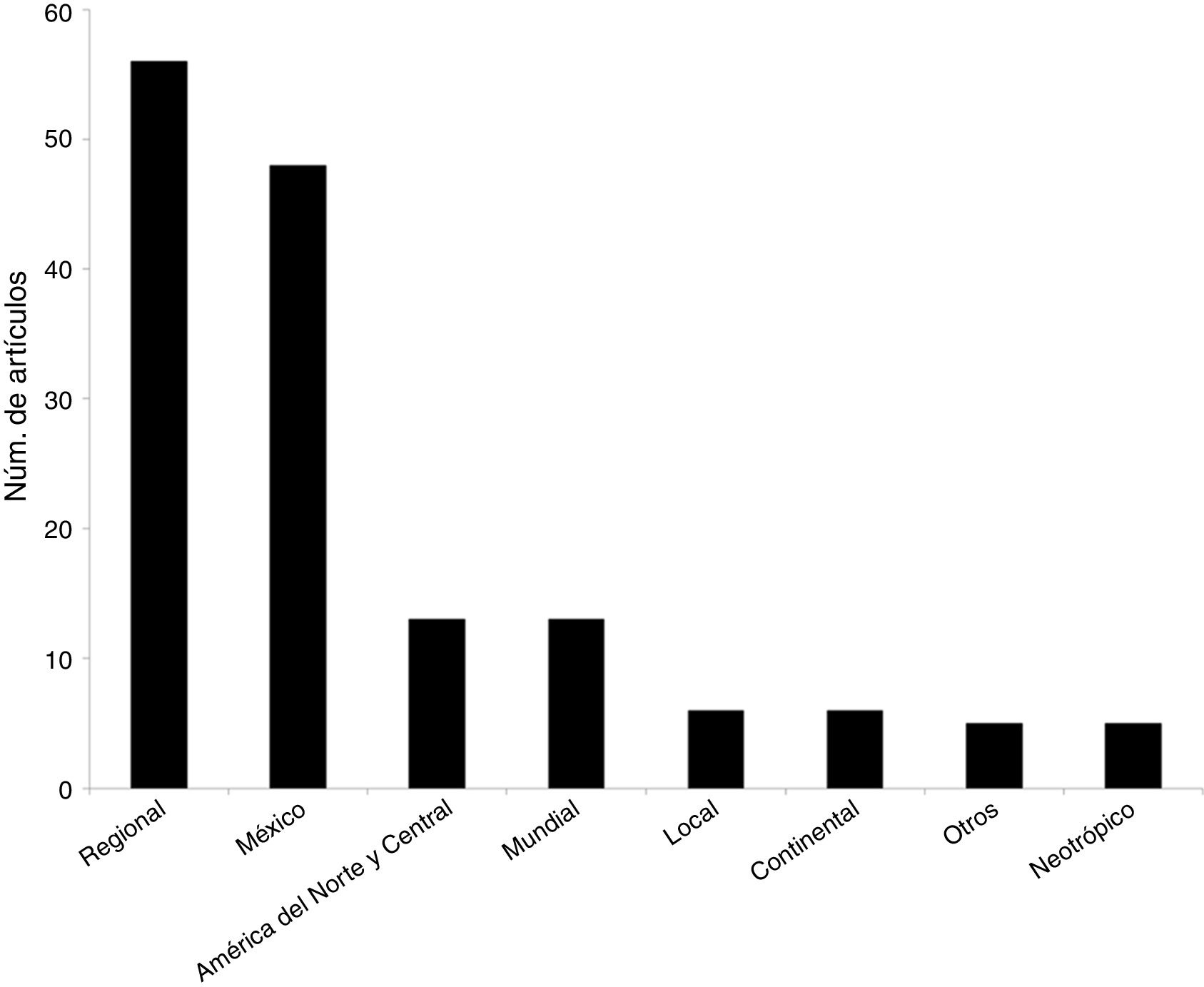
Figura 4
Número de artículos científicos producidos entre 1989 y 2016 por categorías de extensión geográfica, basado en los artículos que se registraron para México (es decir, en los que al menos un autor es de nacionalidad mexicana) (n = 163).
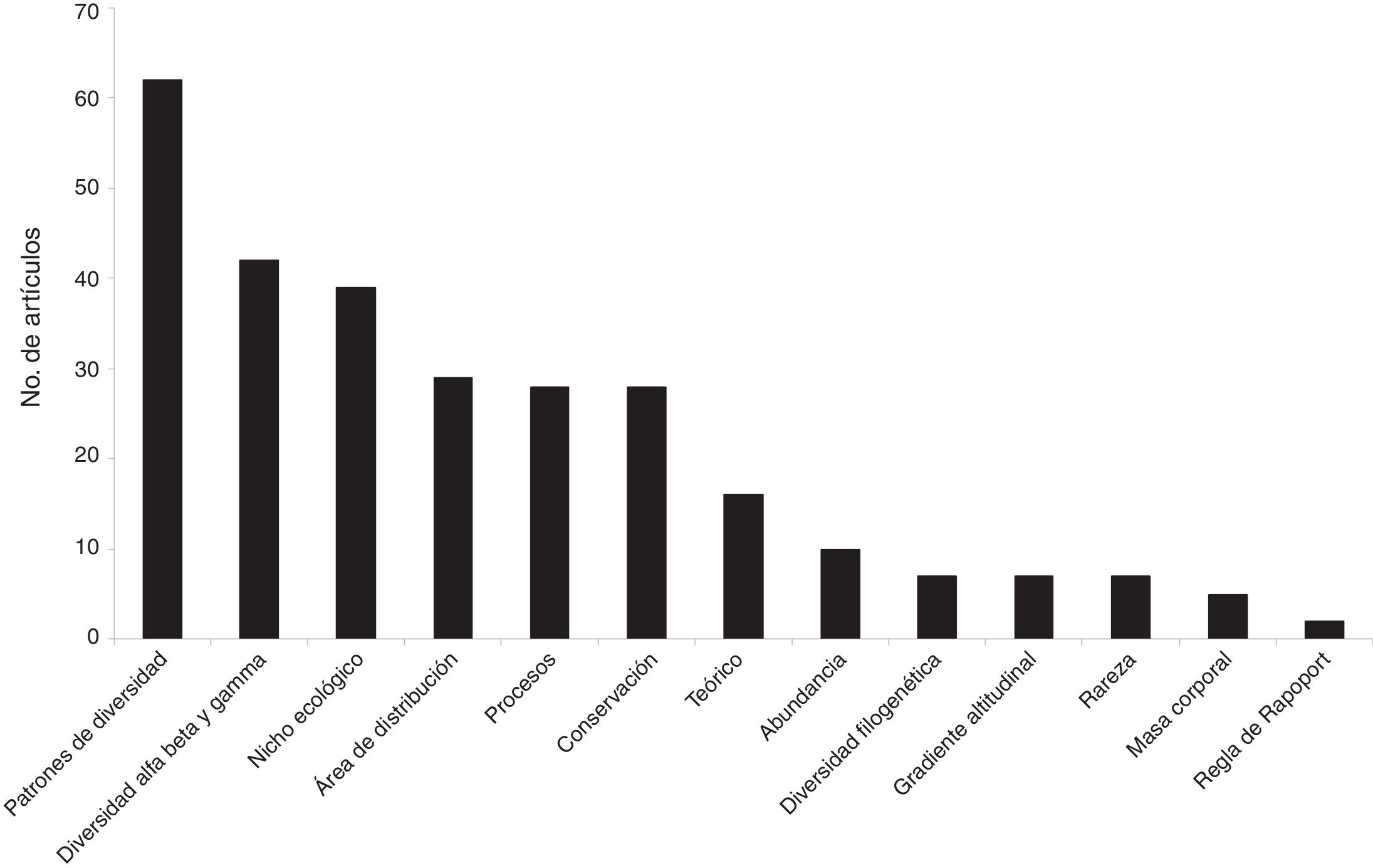
Figura 5
Número de artículos científicos producidos entre 1989 y 2016 por tema macroecológico, basado en los artículos que se registraron para México (es decir, en los que al menos un autor es de nacionalidad mexicana) (n = 163).
Los resultados de las búsquedas utilizando Web of Science reflejan parcialmente el trabajo desarrollado por macroecológos mexicanos. Muchos de los trabajos no fueron detectados en estas búsquedas, aun cuando probamos distintos criterios. Además, una parte de los trabajos que se desarrollan en México se encuentra en artículos no indexados y en la literatura «gris» (e.g., tesis, libros, reportes técnicos, etc.). Sin embargo, consideramos que una búsqueda más exhaustiva en Web of Science y en la literatura gris no modificará sustancialmente las tendencias encontradas en esta revisión.
Aportaciones teórico-conceptuales y aplicadas de la macroecología en México
En general, la macroecología desarrollada por científicos mexicanos ha realizado contribuciones en la mayoría de los grandes temas macroecológicos que, siguiendo a Marquet (2009) , serían: los patrones espaciales de diversidad, la distribución de frecuencias de atributos ecológicos (tamaños de área de distribución, tamaños corporales, etc.), las relaciones o covariación entre dichos atributos ecológicos (e.g., tamaño corporal vs tamaño de área de distribución), la dinámica espacial (reglas ecogeográficas) y temporal (dinámica) de los patrones macroecológicos. Además, incluimos otros temas de macroecología en los que los científicos mexicanos también han contribuido, específicamente la teoría macroecológica, la macroecología evolutiva y la teoría de nicho ecológico. Estos temas se describen y ejemplifican a continuación.
Patrones espaciales de diversidad
El análisis de los patrones espaciales de diversidad es uno de los temas fundamentales de la macroecología ( Brown, 2014; Rosenzweig, 1995 ). En nuestro país, este tema ha sido el más estudiado ( fig. 5 ) y los análisis se han centrado en el gradiente latitudinal de riqueza de especies, en los patrones de diversidad alfa, beta y gamma, y en menor medida en el gradiente altitudinal.
El análisis del gradiente latitudinal de riqueza de especies, que generalmente documenta un aumento del número de especies desde las zonas frías y templadas hacia las tropicales ( Brown, 2014; Rosenzweig, 1995 ), ha sido abordado por diferentes grupos de trabajo en México. Los resultados obtenidos en distintos estudios ratifican la existencia de este gradiente a nivel continental y nacional y han permitido conocer particularidades para distintos grupos taxonómicos. Ejemplos de estos trabajos son los desarrollados para los vertebrados ( Koleff et al., 2008 ), mamíferos terrestres y voladores ( Ceballos y Navarro, 1991; Ceballos y Rodríguez, 1993 ), anfibios y reptiles ( Flores-Villela, 1993; Ochoa-Ochoa, Rodríguez, Mora, Flores-Villela y Whittaker, 2012; Parra-Olea, Flores-Villela y Mendoza-Almeralla, 2014; Pineda y Lobo, 2009 ), aves ( Navarro-Sigu¿enza, Gordillo-Martínez y Peterson, 2009 ). También se han realizado estudios para algunos grupos de plantas (e.g., Golicher, Cayuela, Alkemade, González-Espinosa y Ramírez-Marcial, 2007; González-Espinosa, Rey-Benayas, Ramírez-Marcial, Huston y Golicher, 2004; González-Espinosa, Ramírez-Marcial, Méndez-Dewar, Galindo-Jaimes y Golicher, 2005; Pavón, Hernández-Trejo y Rico-Gray, 2000; Villaseñor, 2003 ). En el estudio del gradiente latitudinal de riqueza de especies existen trabajos interesantes en macroecología marina, por ejemplo, los estudios con invertebrados marinos ( Escobar-Briones, Gaytán-Caballero y Legendre, 2008 ) y mamíferos marinos ( Pompa, Ehrlich y Ceballos, 2011; Rosales-Nanduca, Gerrodette, Urbán, Cárdenas-Hinojosa y Medrano-González, 2011 ), por mencionar algunos.
En general, los estudios mencionados han sido de índole descriptiva y han permitido inferir algunas de las causas asociadas a la estructura y a la variación geográfica en las comunidades. Por ello, un reto futuro importante es el de avanzar en el entendimiento de los procesos que expliquen los patrones identificados, utilizando las nuevas aproximaciones de la macroecología (ver la sección «Macroecología evolutiva»).
En México se ha analizado también el gradiente altitudinal de riqueza de especies que, en términos generales, muestra un máximo número de especies en altitudes intermedias ( Mena y Vázquez-Domínguez, 2005; Navarro y León-Paniagua, 1995; Sánchez-Cordero, 2001 ). El número de trabajos identificados en este tema resultó bajo ( fig. 5 ) y, en general, estos trabajos han confirmado el patrón de mayor riqueza y recambio de especies en altitudes intermedias, aunque con algunas excepciones, donde se ha observado que la riqueza se incrementa con la altitud ( Navarro y León-Paniagua, 1995 ). Esta aparente discrepancia es común en los estudios de gradientes altitudinales y parece estar relacionada con la amplitud de la variación ambiental que fue cubierta en los gradientes estudiados ( McCain, 2007 ). De igual forma que en el tema del gradiente latitudinal de riqueza de especies, un reto importante en el estudio de gradientes altitudinales es el de avanzar hacia el entendimiento de los procesos que subyacen a los patrones altitudinales documentados.
Además de describir la estructura espacial de la biodiversidad, desde los primeros trabajos macroecológicos realizados por autores mexicanos se ha puesto énfasis en las aplicaciones de estos estudios a la conservación de la biodiversidad ( fig. 5). Por ejemplo, Ceballos, Rodríguez y Medellín (1998) analizan el patrón espacial de riqueza de especies separando el patrón espacial de las especies raras, de las endémicas y de las especies amenazadas. Encontraron que las regiones en las que se concentra la mayor riqueza de especies son distintas a las regiones en las que se concentran las especies raras, las endémicas o las que están amenazadas, mostrando que las estrategias de conservación deben ser distintas, dependiendo del grupo de especies que se desee conservar. Estudios en los que se analizan los patrones de los mamíferos a nivel mundial confirman el patrón encontrado para los mamíferos de México ( Ceballos, 2007; Ceballos y Brown, 1995; Ceballos y Ehrlich, 2006 ), y de igual forma, resultados semejantes han sido obtenidos para la avifauna mexicana ( Escalante, Navarro-Sigüenza y Peterson, 1993 ) y para las plantas de Baja de California ( Riemann y Ezcurra, 2005 ), por mencionar algunos casos.
Otro tema que destaca es el estudio de los patrones de la diversidad a distintas escalas espaciales ( fig. 5). La propuesta de Whittaker (1960, 1972) de dividir la diversidad en sus componentes alfa, beta y gamma provee un marco conceptual idóneo para el estudio da la diversidad en un país megadiverso ( Arita, 1997; Arita y León-Paniagua, 1993; Halffter, Soberón, Koleff y Melic, 2005; Moreno y Halffter, 2001; Sarukhán, Soberón y Larson-Guerra, 1996 ). En estudios desarrollados para los vertebrados, principalmente mamíferos ( Arita, 1997; Arita y Rodríguez, 2002; Rodríguez y Arita, 2004; Rodríguez, Soberón y Arita, 2003 ), anfibios y reptiles (Ochoa-Ochoa et al., 2012 ), se ha puesto a prueba la hipótesis propuesta por Arita y León-Paniagua (1993) , quienes plantearon que la gran diversidad de especies a nivel país se debe a la combinación de una diversidad alfa relativamente moderada y a una diversidad beta elevada, esta última resultado de la gran heterogeneidad ambiental existente en el país. También se han llevado a cabo algunos estudios para entender los patrones de diversidad de especies de cactáceas del país ( Goettsch y Hernández, 2006; Golubov, Mandujano y Mandujano, 2005 ), cuyos resultados abonan la idea de que la alta diversidad beta es un patrón común a muchos grupos biológicos ( Koleff et al., 2008).
Los estudios referentes a diversidad alfa y beta a escala local y regional son numerosos (e.g., García, Solano-Rodríguez y Flores-Villela, 2007; Lira-Noriega, Soberón, Navarro-Sigüenza, Nakazawa y Peterson, 2007; Pineda y Halffter, 2004; Pineda, Moreno, Escobar y Halffter, 2005 ). Un estudio representativo es aquel que compara la diversidad beta de distintos grupos taxonómicos en el istmo de Tehuantepec ( Calderón-Patrón, Moreno, Pineda-López, Sánchez-Rojas y Zuria, 2013 ). A esta escala de análisis se han puesto a prueba hipótesis que proponen explicar los patrones de diversidad beta de distintos grupos biológicos, como por ejemplo la capacidad de dispersión de las especies. De igual forma, estudios realizados con plantas de selvas estacionalmente secas de la zona de Nizanda, Oaxaca (e.g., Gallardo-Cruz, Meave, Pérez-García y Hernández-Stefanoni, 2010 ), y algunas zonas de la península de Yucatán ( López-Martínez et al., 2013 ) han permitido evaluar la contribución de distintas variables climáticas y topográficas en los patrones de diversidad beta.
Los estudios de la diversidad alfa, beta y gamma también tienen aplicaciones para la conservación de la biodiversidad. Por ejemplo, una propuesta relativamente reciente consistió en utilizar información acerca de la diversidad beta para definir «reservas archipiélagos», un sistema de áreas protegidas en el que se maximizaría el número de especies a conservar ( Halffter, 2007).
Dentro del tema de diversidad beta destacan varios aportes teóricos y metodológicos de importancia a nivel mundial ( fig. 5 ). Por ejemplo, el trabajo desarrollado por Koleff, Gaston y Lennon (2003) , en el que se analiza el desempeño de numerosas medidas para cuantificar la diversidad beta, es sin lugar a dudas un referente en este tema. Más recientemente, Soberón y Cavner (2015) demostraron que es posible derivar muchos de los índices que se usan para describir los patrones geográficos de biodiversidad, incluyendo los de diversidad beta, analizando matrices de incidencias de especies, construidas registrando la presencia (1) o ausencia (0) de cada especie en un conjunto de celdas geográficas; este análisis evalúa el traslape de las áreas de distribución geográfica de las especies. Otros trabajos que representan un aporte importante en este tema son aquellos en los que se analizó la diversidad beta a distintas escalas espaciales, destacando un estudio con aves ( Lennon, Koleff, Greenwood y Gaston, 2001 ) y otro con mamíferos no voladores ( Arita y Rodríguez, 2002; Ochoa-Ochoa et al., 2014 ). A grandes rasgos, en estos trabajos se demuestra que la magnitud de la diversidad beta es dependiente de la escala de análisis ( Moreno y Rodríguez, 2010, 2011 ).
Distribución de frecuencias de atributos ecológicos
Estudiar las distribuciones de frecuencias de atributos ecológicos, además de describir la variación en los valores de dichos atributos, puede informar acerca de los procesos causales de dichas distribuciones ( Gaston, 1990 ). Uno de los atributos más estudiados al respecto ha sido el tamaño del área de distribución de las especies ( fig. 5 ). En México, el primer trabajo que abordó este tema fue el de Arita et al. (1997) para mamíferos terrestres. Este trabajo corroboró el patrón macroecológico descrito anteriormente por Gaston (1990) : las áreas de distribución tienden a ser mayoritariamente pequeñas, con algunas áreas de tamaño mediano y muy pocas de tamaños grandes. Los resultados de este estudio tienen implicaciones directas para la conservación. Por ejemplo, un criterio para definir un área a conservar puede ser la identificación de regiones que concentren la mayor cantidad de especies con áreas de distribución restringidas ( Navarro-Sigu¿enza et al., 2011; Vázquez, Rodríguez y Arita, 2008; Vázquez y Valenzuela-Galván, 2009 ).
En una publicación (Koleff et al., 2008) que forma parte de la obra Capital Natural de México , publicada por la Conabio, se analizaron los patrones de área de distribución de 4 grupos de vertebrados terrestres de México (anfibios, aves, mamíferos y reptiles). Además de que se ratificó la existencia de un mayor número de especies con área de distribución pequeña y pocas especies con área de distribución grande, se distinguieron particularidades para cada grupo estudiado. Por ejemplo, las regiones del país en las que se concentran las especies con áreas de distribución pequeñas son diferentes para cada uno de estos grupos. En términos de conservación, a grandes rasgos, este resultado indica que se requeriría considerar distintas regiones del país para conservar diferentes grupos de vertebrados terrestres ( Koleff et al., 2008).
Un estudio pionero, que evaluó la importancia relativa de especies que difieren en la amplitud de su distribución geográfica al patrón espacial de riqueza de especies, mostró que en México las especies con distribución amplia contribuyen en mayor medida a dichos patrones que las especies con distribución restringida ( Vázquez y Gaston, 2004 ). Trabajos posteriores han profundizado en este tema, demostrando que la relación positiva entre riqueza de especies y el número de especies con distribución amplia es preponderante, considerando tanto la cohesión de las áreas de distribución (i.e., conformación espacial continua) como la variación del área de distribución geográfica de las especies ( Arita y Rodríguez-Tapia, 2009).
Covariación de atributos ecológicos
Evaluar la variación entre 2 (o más) atributos ecológicos entre diferentes especies ha sido uno de los temas centrales de la macroecología desde su formalización como disciplina científica ( Brown y Maurer, 1989 ). La lógica detrás de este análisis se basa en que los mecanismos ecológicos y evolutivos que determinan los niveles actuales de biodiversidad deberían verse reflejados en los patrones de covariación espacial de tales atributos ecológicos ( Brown y Maurer, 1987).
Uno de los patrones de covariación más estudiados corresponde a la relación entre el tamaño (o masa) corporal y atributos ecológicos tales como la abundancia poblacional y el área de distribución geográfica. En 2 trabajos pioneros a nivel global, desarrollados por un investigador mexicano (H. Arita) ( fig. 5 ), se evaluó la relación entre el tamaño corporal y la «rareza» (medida como densidad poblacional baja o distribución geográfica restringida), gremios alimentarios e historia (e.g., relaciones entre especies; Arita, Robinson y Redford, 1990; Arita, 1993 ) de especies de mamíferos neotropicales. Estos estudios mostraron que la relación entre las 2 variables de «rareza», área de distribución y densidad poblacional está modulada por el tamaño corporal, con relaciones negativas entre estas 2 variables en grupos de especies con mayor variación en tamaño corporal. Efectivamente, dicha relación negativa fue encontrada en los mamíferos neotropicales asociados a bosques ( Arita et al., 1990 ) pero no para los murciélagos neotropicales ( Arita, 1993 ), los cuales muestran una variación en tamaño corporal menor a la de los mamíferos no voladores.
Estudios más recientes proponen un enfoque integrador y simple que considera simultáneamente 3 de los principales atributos de la biodiversidad: riqueza de especies, rareza de las especies ( Villalobos, Dobrovolsk, Provete y Gouveia, 2013; Villalobos, Lira-Noriega, Soberón y Arita, 2013 ) e información evolutiva (Mendoza y Arita, 2014 ). Con la utilización de información macroecológica básica (i.e., matriz de incidencias de las especies) es posible identificar regiones ricas en especies raras (hotspots) y regiones pobres en dichas especies ( coldspots; Villalobos, Dobrovolsk et al., 2013 ). Dicho enfoque puede ser útil cuando se cuenta con poca información acerca de la distribución de las especies (e.g., mapas de distribución a escala geográfica en lugar de registros puntuales a escala local) y como una primera descripción rápida de los patrones de diversidad y distribución de la biota de una región.
La covariación entre el tamaño del área de distribución y la abundancia local también se da a nivel de especies individuales. Por ejemplo, en biogeografía (disciplina fundacional de la macroecología) se postula que hay una relación positiva entre la abundancia de individuos y la distancia del centro de la distribución geográfica de la especie. Al respecto, Martínez-Meyer, Díaz-Porras, Peterson y Yáñez-Arenas (2012) propusieron una aproximación novedosa en la que se incorpora la teoría de nicho, encontrando que la variación en la abundancia está más relacionada con las preferencias ambientales de las especies (i.e., su nicho ecológico) que con variables como la distancia a un centro geográfico ( Osorio-Olvera, Falconi y Soberón, 2016; Yáñez-Arenas, Martínez-Meyer, Mandujano y Rojas-Soto, 2012 .
Dinámica espacial y temporal de patrones macroecológicos
Los atributos ecológicos (e.g., tamaño corporal, área de distribución, historia de vida, etc.) pueden variar tanto en el espacio como en el tiempo. En la dimensión espacial, la variación de estos atributos está relacionada con la ubicación de las especies en el espacio geográfico o a lo largo de gradientes ambientales. Las variaciones geográficas de los atributos ecológicos de las especies son conocidas como «reglas ecogeográficas» ( Gaston, Chown y Evans, 2008 ). Dos de estas reglas han sido estudiadas en macroecología: la regla de Bergmann, correspondiente al tamaño corporal (a mayor temperatura, menor tamaño corporal; Blackburn, Gaston y Loder, 1999; Olalla-Tárraga, Bini, Diniz-Filho y Rodríguez, 2010 ), y la de Rapoport, correspondiente al tamaño del área de distribución de las especies (a mayor latitud, mayor tamaño del área de distribución; Hawkins y Diniz-Filho, 2006; Stevens, 1989 ). Los trabajos de macroecólogos mexicanos en estos temas son escasos ( fig. 5 ), aunque en lo que corresponde a la regla de Rapoport se han realizado contribuciones interesantes, enfocadas a entender la integración de diferentes patrones interconectados, incluyendo el gradiente latitudinal de riqueza de especies, la diversidad beta, la variación del tamaño de área de distribución de las especies y la regla de Rapoport. Por ejemplo, Arita, Rodríguez y Vázquez-Domínguez (2005) evaluaron la hipótesis de que el gradiente latitudinal de riqueza de especies es el resultado de la tendencia a que el tamaño de las áreas de distribución de las especies disminuye conforme nos acercamos a los trópicos (regla de Rapoport). Llevando a cabo comparaciones de los patrones observados contra los esperados bajo un modelo nulo (i.e., expectativa teórica en ausencia de un proceso de interés), demostraron la interconexión entre los patrones antes mencionados y la validez de la regla de Rapoport para los mamíferos terrestres de América del Norte.
En relación con la dimensión temporal de los patrones macroecológicos, son pocos los estudios realizados por macroecológos mexicanos. Uno de los primeros estudios en considerar explícitamente la dimensión temporal bajo un enfoque teórico fue el de Arita y Vázquez-Domínguez (2008) . En dicho estudio se propone un modelo nulo para evaluar si los trópicos son cuna (i.e., generadores) o museo (i.e., acumuladores) de la diversidad de especies. Con este enfoque original, los autores evaluaron la influencia relativa de procesos históricos como especiación, extinción de especies y dinámica de cambio de las áreas de distribución. Los resultados de este estudio sugieren que el gradiente latitudinal de la riqueza de especies es el resultado de balances entre altas tasas de especiación y extinción en los trópicos, generando tanto una cuna como un museo de diversidad en esa región. En un estudio más reciente se evaluó la dinámica temporal de los patrones de coexistencia geográfica de mamíferos extintos y se confirmó el efecto que tienen los factores históricos, tanto evolutivos (especiación y extinción de especies) como cambios climáticos de largo plazo, en dichos patrones de coexistencia ( Villalobos, Dobrovolsk et al., 2013 ). Con la continua generación y disponibilidad de datos relacionados con la historia de las especies (e.g., relaciones filogenéticas, datos fósiles, modelos climáticos), la evaluación de la dinámica temporal de los patrones macroecológicos seguramente será un tema de investigación importante en los próximos años, tal como lo muestran algunos trabajos recientes ( Saupe et al., 2015; Qiao, Saupe, Soberón, Peterson y Myers, 2016 ; ver más adelante la sección «Macroecología evolutiva»).
Teoría macroecológica
Los estudios macroecológicos realizados por autores mexicanos no solo han contribuido a la descripción de patrones y a la evaluación de hipótesis macroecológicas, sino también al desarrollo de la teoría macroecológica ( Material suplementario 2) (fig. 5 ). Un aporte importante es el de Arita, Christen, Rodríguez y Soberón (2012) . En este trabajo se propone una nueva manera de analizar la riqueza y el área de distribución de las especies de manera simultánea, identificando las relaciones biológicas y las restricciones analíticas entre estas 2 variables. Este trabajo ha sido la base para nuevos desarrollos teóricos ( Arita, 2015; Borregaard y Rahbek, 2010; Christen y Soberón, 2009; Soberón y Cavner, 2015; Villalobos y Arita, 2010; Villalobos, Lira-Noriega, Soberón y Arita, 2014 ) y aplicados ( Mendoza y Arita, 2014; Soberón y Ceballos, 2011; Villalobos, Dobrovolsk et al., 2013; Villalobos, Lira-Noriega et al., 2013 ). Una conclusión relevante de este aporte teórico es que los patrones de biodiversidad pueden ser descritos usando únicamente algunos parámetros interrelacionados, basados en las relaciones matemáticas entre diversidad y distribución de especies, evitando la proliferación de nuevos índices y facilitando la unificación de diferentes conceptos de biodiversidad bajo un mismo marco teórico ( Arita, Christen, Rodríguez y Soberón, 2008 ). En los próximos años se revelarán los alcances de dicho marco y sus posibilidades de contribuir al desarrollo de la teoría macroecológica ( Soberón, 2015).
Macroecología evolutiva
Recientemente, la macroecología se ha beneficiado de la rápida generación y disponibilidad de información acerca de la historia de las especies (p.ej., registro fósil, relaciones filogenéticas) y del planeta (p.ej., modelos climáticos). Esto ha permitido investigar con mayor profundidad los componentes históricos de los patrones de biodiversidad. Una nueva línea de investigación, llamada «macroecología evolutiva» ( Diniz-Filho, Gouveia y Lima-Ribeiro, 2013; Villalobos, Carotenuto, Raia y Diniz-Filho, 2016 ), considera de manera explícita, tanto analítica como conceptualmente, la historia de las especies y del planeta en la evaluación de tales patrones. Los autores mexicanos han comenzado a contribuir con esta nueva línea de investigación incorporando la dimensión temporal en sus estudios ( fig. 5). Por ejemplo, Velasco et al. (2016) (ver también Saupe et al., 2015, y Qiao et al., 2016 ) utilizaron información sobre la distribución geográfica, las preferencias ambientales y las relaciones filogenéticas de las especies para determinar la influencia del nicho ecológico en la diversificación evolutiva de un clado. Del mismo modo, Villalobos, Carotenuto et al. (2016) y Villalobos, Cianciaruso, Olalla-Tárraga, Rangel y Diniz-Filho (2016) emplearon datos de fósiles y de especies actuales, junto con información y análisis evolutivos, con el fin de entender las causas de los patrones de coexistencia geográfica entre especies, los cuales subyacen a los gradientes geográficos de biodiversidad. La macroecología evolutiva es sin duda una de las líneas de investigación más prometedoras para los años venideros ( Raia y Fortelius, 2017).
Teoría de nicho ecológico
Otra línea sobresaliente en la que macroecólogos mexicanos han hecho contribuciones importantes es en la teoría de nicho ecológico, particularmente en lo referente a la relación entre el nicho ecológico y el área de distribución geográfica de las especies, materia prima de los análisis macroecológicos ( fig. 5 ). Los aportes en este tema van desde trabajos teóricos ( Peterson y Soberón, 2012; Soberón, 2007 ) hasta el análisis de patrones de diversidad ( Pérez-Moreno, Martínez-Meyer, Soberón Mainero y Rojas-Soto, 2016; Peterson et al., 2002; Pineda y Lobo, 2009 ). Por un lado, el valor teórico de estos estudios radica en su aporte al entendimiento de la relación entre el nicho ecológico de las especies y la expresión geográfica del mismo (i.e., el área de distribución de las especies). Por otro lado, los trabajos empíricos han mostrado que tener mejores modelos de distribución de especies, basados en la teoría de nicho, permite realizar análisis cada vez más refinados acerca de patrones de diversidad ( Koleff et al., 2008; Peterson et al., 2002 ), adentrarse en los procesos que explican tales patrones (como, por ejemplo, entender los patrones migratorios de grupos de especies en México; Pérez-Moreno et al., 2016 ) e incluso realizar análisis aplicados a la conservación de la biodiversidad ( Ortega-Huerta y Peterson, 2001).
Vacíos de conocimiento y desafíos teórico-metodológicos
Si bien hay temas de la macroecología en los que ha habido un aporte significativo de los autores mexicanos, también hay temas que han sido escasamente desarrollados o que definitivamente no se han abordado en México (al menos hasta donde tenemos conocimiento). A continuación describiremos 5 de ellos.
En primer lugar, la investigación macroecológica en México ha tendido a ser principalmente descriptiva. Muchos de los trabajos han analizado patrones geográficos de diversidad, pero son pocos los que han asociado procesos o mecanismos que expliquen estos patrones. Algunos ejemplos de estudios en los que sí se busca comprender las causas que determinan las distribuciones geográficas de las especies han considerado modelos de procesos explícitos, como la dispersión y las tasas demográficas de las especies ( Lira-Noriega, Soberón y Miller, 2013; Osorio-Olvera et al., 2016 ), o bien el efecto del clima y la herencia de preferencias ambientales (i.e., conservación de nicho) sobre los patrones de codistribución entre especies y codiversidad entre sitios ( Villalobos et al., 2014 ). Estos trabajos pueden identificarse con el tema «Procesos» en el Material suplementario 2 (fig. 5).
En segundo lugar, las herramientas estadísticas propias de la macroecología no se emplean con frecuencia. Por ejemplo, son pocos los estudios en los que se utilizan modelos nulos con los cuales contrastar los resultados empíricos (e.g., Arita et al., 2005; Arita y Vázquez-Domínguez, 2008 ), aunque esta tendencia se está modificando en las nuevas generaciones de macroecólogos mexicanos ( Trejo-Barocio y Arita, 2013; Villalobos et al., 2014 ). La capacidad de cómputo con que se cuenta actualmente, así como el desarrollo de algoritmos eficientes para manipular grandes cantidades de datos ( Vilela y Villalobos, 2015 ), indican que los modelos de simulación para entender los patrones macroecológicos se aplicarán más frecuentemente en el futuro próximo.
En tercer lugar, existen temas que han sido escasamente desarrollados, como por ejemplo los patrones de distribución geográfica de la abundancia y de la masa corporal de las especies ( fig. 5 ), dos de los atributos más estudiados en macroecología. La vinculación entre un enfoque de nicho ecológico y la macroecología representa un gran potencial para avanzar en la búsqueda de procesos y mecanismos que expliquen los patrones de abundancia y distribución de las especies ( Martínez-Meyer et al., 2012; Yáñez-Arenas et al., 2012 ). En relación con el patrón de masa corporal, un trabajo notable lo constituye el desarrollado por Arita y Figueroa (1999) , inspirado en el clásico trabajo de Brown y Nicoletto (1991) . En este estudio se utiliza la masa corporal como una medida de diversidad complementaria a la riqueza de especies; además, la visualización en el espacio geográfico de la diversidad de masas corporales constituye uno de los primeros ejercicios de este tipo a nivel mundial.
En cuarto lugar, aún son escasos los estudios en los que se incorporan aspectos filogenéticos y evolutivos, así como atributos funcionales en los estudios macroecológicos ( fig. 5 ). A nivel mundial, en los estudios macroecológicos se utiliza cada vez más este tipo de información y se hacen análisis más sofisticados. Esta tendencia está comenzando a plasmarse en trabajos realizados por investigadores mexicanos (ver ejemplos en la sección «Macroecología evolutiva»).
En quinto lugar, los estudios se han centrado en pocos grupos taxonómicos. La mayoría de los estudios se enfocan en vertebrados terrestres, algunos en mamíferos marinos, y en ciertos grupos de plantas (principalmente cactáceas; fig. 2 ). Algunas excepciones importantes las constituyen los estudios en los que se analiza la microdiversidad de suelos con un enfoque de dividir la diversidad en sus componentes alfa, beta y gamma ( Noguez et al., 2005 ), y algunos trabajos con parásitos de helmintos de agua dulce ( Garrido-Olvera, Arita y Pérez-Ponce de León, 2012; Quiroz-Martínez y Salgado-Maldonado, 2013 ), por mencionar algunos (fig. 2).
Conclusiones y perspectivas de la macroecología en México
La macroecología a nivel mundial ha tenido un gran desarrollo desde su formalización como disciplina hasta nuestros días ( Beck et al., 2012 ). Es una disciplina que se ha consolidado, y cada vez cuenta con más grupos de investigación alrededor del planeta que interactúan entre ellos, cada vez con más frecuencia. Por otro lado, en los últimos años ha habido un espectacular avance en la generación y disponibilidad de información como, por ejemplo, de filogenias moleculares para diversos grupos taxonómicos, de bases de datos climáticos y de distribución geográfica de alta resolución a escala planetaria. Aunado a esto, el mayor poder computacional y los nuevos enfoques analíticos ( Vilela y Villalobos, 2015 ) están contribuyendo a impulsar el desarrollo de este campo. En esta nueva etapa de la macroecología se han re-analizado los patrones macroecológicos clásicos utilizando datos de mayor resolución y nuevos métodos (i.e., McKnight et al., 2007 ), se han planteado nuevas preguntas y se avanza hacia la síntesis de distintos patrones, así como hacia el entendimiento de los procesos y mecanismos que subyacen a dichos patrones ( Brown, 2014; Gotelli et al., 2009 ).
¿Cómo se inserta la macroecología mexicana en este escenario mundial? Después de la revisión de las fortalezas y debilidades de esta ciencia en el país, hemos detectado varias oportunidades y perspectivas para el desarrollo de esta disciplina en México:
- El potencial para la investigación macroecológica (empírica y teórica) con el que se cuenta en México en la actualidad está mediado directamente por los casi 10 Gigabytes de datos primarios sobre la biodiversidad disponibles en el país, una cantidad que crece día con día, y las herramientas bioinformáticas que están en continuo desarrollo por mexicanos y extranjeros. La exitosa utilización que se ha hecho en las últimas dos décadas de estos datos para desarrollar modelos de distribución de especies y, por lo tanto, de patrones de biodiversidad a escalas biogeográficas, más el potencial de correlacionar esta información con bases de datos de biomasa, de niveles tróficos, de funciones ecológicas, de interacciones bióticas, entre otras, constituye un gran potencial para el desarrollo de la macroecología en México.
- La macroecología mexicana debe pasar de ser una disciplina descriptiva a una disciplina que avance hacia el entendimiento de los procesos y mecanismos que expliquen los patrones de biodiversidad.
- Deben impulsarse temas en los que se integre información a distintas escalas espaciales y temporales, incorporando diferentes tipos de datos (e.g., distribución y abundancia) y facetas poco exploradas de la biodiversidad (e.g., diversidad filogenética y funcional), para avanzar hacia una síntesis acerca de la variación de la biodiversidad a diferentes escalas espaciotemporales.
- Es importante romper el fuerte sesgo taxonómico que existe en los trabajos macroecológicos y ampliar los análisis a diferentes grupos taxonómicos. Esto implica tanto la generación de datos nuevos con suficiente cobertura espacial (e.g., para todo el país) y resolución (e.g., taxonómica, geográfica, etc.), así como mejorar la disponibilidad de esos datos para su uso, con la aplicación de tecnologías computacionales para compartir datos. Cabe resaltar que la generación y compilación de datos debe ir de la mano con generación de teoría y la puesta a prueba de diferentes predicciones macroecológicas.
- Se deben fortalecer y multiplicar los grupos de trabajo en México que se dedican directamente a realizar una labor de investigación en macroecología. Es importante también establecer colaboraciones internacionales permanentes con grupos de trabajo enfocados en aspectos macroecológicos, donde los estudiantes e investigadores posdoctorados puedan realizar estancias de investigación y trabajo en los países convenio. Esto fortalecería los grupos actuales y ayudaría a la diversificación de temas de estudio con enfoque macroecológico.
- En el Padrón Nacional de Posgrados de Calidad de Conacyt no existe ningún programa de posgrado en macroecología, aunque en los programas de posgrado de algunas instituciones (p.ej., en el Colegio de la Frontera SUR [ECOSUR] y en el INECOL) se imparten cursos específicos del tema o este se incluye como parte de cursos de disciplinas estrechamente asociadas (e.g., ecología de comunidades). Sería muy importante incentivar la oferta de cursos de macroecología a nivel de posgrado para generar el interés de los alumnos y formar nuevos investigadores en esta disciplina.
- La investigación macroecológica requiere habilidades como el manejo de grandes volúmenes de información. Por ello, el currículum de los programas de posgrado de ecología y otras áreas afines a la macroecología del país deben reforzarse con cursos de análisis espacial, de minería de datos, de sistemas de información geográfica y de informática. Adicionalmente, la inversión en la capacidad de almacenamiento de datos es de suma importancia para el desarrollo de esta y otras disciplinas en ecología.
- Los resultados de análisis macroecológicos pueden proveer información primaria para la planeación de la conservación, tal como se ha ejemplificado en esta revisión. Sin embargo, aún es necesario que la biología de la conservación y la macroecología se entrelacen y retroalimenten de mejor manera.
La macroecología es probablemente la disciplina de la ecología con menor representación en México. Esto se ve reflejado tanto en términos de número de grupos de trabajo e investigadores activos, así como en la formación de estudiantes de posgrado y profesionistas en el área. La macroecología en México no ha alcanzado un nivel de madurez ni tiene una masa crítica importante aún; sin embargo, el potencial que tiene para generar información básica y aplicada que permita que el país haga frente a retos tan importantes como la pérdida de la biodiversidad y el cambio climático, entre otros, la convierte en una disciplina estratégica para el desarrollo científico y el desarrollo de políticas públicas sobre conservación en México. La relación directa entre la macroecología y la conservación biológica puede fomentar aún más la integración entre los diferentes sectores de la sociedad, desde los académicos y las instituciones educativas hasta los ciudadanos y las oficinas de gobierno.
Agradecimientos
Al comité organizador de este número especial, por la invitación a participar. A Héctor Arita por los comentarios que hizo al manuscrito pero sobre todo por sus enseñanzas y su ejemplo. Al Dr. Jorge Soberón y a la Dra. Claudia Moreno por sus críticas y sugerencias que sin duda mejoraron el escrito. A Eduardo H. Rapoport, por su inspirador trabajo y como un pequeño homenaje póstumo a su gran labor. Su libro Areografía: Estrategias geográficas de las especies contiene originales ideas y conceptos que constituyen una de las bases del desarrollo de la macroecología.
Anexo A
Material suplementario
Referencias
Arita, 1993
H.T. Arita
Rarity in Neotropical bats: correlations with phylogeny, diet, and body mass
Ecological Applications
3
1993
506-17
Arita, 1997
H.T. Arita
The non-volant mammal fauna of Mexico: species richness in a megadiverse country
Biodiversity and Conservation
6
1997
787-95
Arita, 2015
H.T. Arita
Multisite and multispecies measures of overlap, co-occurrence, and co-diversity
Ecography
40
2015
709-18
Arita et al., 2008
H.T. Arita
- Christen
- Rodríguez
- Soberón
Species diversity and distribution in presence-absence matrices: mathematical relationships and biological implications
American Naturalist
172
2008
519-32
Arita et al., 2012
H.T. Arita
- Christen
- Rodríguez
- Soberón
The presence-absence matrix reloaded: the use and interpretation of range-diversity plots
Global Ecology and Biogeography
21
2012
282-92
Arita y Figueroa, 1999
H.T. Arita
- Figueroa
Geographic patterns of body-mass diversity in Mexican mammals
Oikos
85
1999
310-9
Arita et al., 1997
H.T. Arita
- Figueroa
- Frisch
- Rodríguez
- Santos-del Prado
Geographical range size and the conservation of Mexican mammals
Conservation Biology
11
1997
92-100
Arita y León-Paniagua, 1993
H.T. Arita
- León-Paniagua
Diversidad de mamíferos terrestres
Ciencias [Núm. Especial]
7
1993
13-22
Arita et al., 1990
H.T. Arita
J.G. Robinson
K.H. Redford
Rarity in neotropical forest mammals and its ecological correlates
Conservation Biology
4
1990
181-92
Arita y Rodríguez, 2002
H.T. Arita
- Rodríguez
Geographic range, turnover rate and the scaling of species diversity
Ecography
25
2002
541-53
Arita y Rodríguez-Tapia, 2009
H.T. Arita
- Rodríguez-Tapia
Contribution of restricted and widespread species to diversity: the effect of range cohesion
Ecography
32
2009
210-4
Arita et al., 2005
H.T. Arita
- Rodríguez
- Vázquez-Domínguez
Continental and regional range sizes of North American mammals: Rapoport’s rule in null and real worlds
Journal of Biogeography
32
2005
961
Arita y Vázquez-Domínguez, 2008
H.T. Arita
- Vázquez-Domínguez
The tropics: cradle, museum or casino? A dynamic null model for latitudinal gradients of species diversity
Ecology Letters
11
2008
653-63
Beck et al., 2012
- Beck
- Ballesteros-Mejia
C.M. Buchmann
- Dengler
S.A. Fritz
- Gruber
What’s on the horizon for macroecology?
Ecography
35
2012
673-83
Blackburn, 2004
T.M. Blackburn
Method in macroecology
Basic and Applied Ecology
5
2004
401-12
Blackburn et al., 1999
T.M. Blackburn
K.J. Gaston
- Loder
Geographic gradients in body size: a clarification of Bergmann’s rule
Diversity and Distributions
5
1999
165-74
Borregaard y Rahbek, 2010
M.K. Borregaard
- Rahbek
Dispersion fields, diversity fields and null models: uniting range sizes and species richness
Ecography
33
2010
402-7
Brown, 1995
J.H. Brown
Macroecology
1995
Brown, 1999
J.H. Brown
The legacy of Robert Macarthur: from geographical ecology to macroecology
Journal of Mammalogy
80
1999
333-44
Brown, 2014
J.H. Brown
Why are there so many species in the tropics?
Journal of Biogeography
41
2014
8-22
Brown y Maurer, 1987
J.H. Brown
B.A. Maurer
Evolution of species assemblages: effects of energetic constraints and species dynamics on the diversification of North American avifauna
American Naturalist
130
1987
1-17
Brown y Maurer, 1989
J.H. Brown
B.A. Maurer
Macroecology: the division of food and space among species on continents
Science
243
1989
1145-50
Brown y Nicoletto, 1991
J.H. Brown
P.F. Nicoletto
Spatial scaling of species composition: body masses of North American land mammals
American Naturalist
138
1991
1478-512
Brown et al., 1996
J.H. Brown
G.C. Stevens
D.M. Kaufman
The geographic range: size, shape, boundaries, and internal structure
Annual Review of Ecology and Systematics
27
1996
597-623
Calderón-Patrón et al., 2013
- Calderón-Patrón
C.E. Moreno
- Pineda-López
- Sánchez-Rojas
- Zuria
Vertebrate dissimilarity due to turnover and richness differences in a highly beta-diverse region: the role of spatial grain size, dispersal ability and distance
Plos One
8
2013
e82905
Ceballos, 2007
- Ceballos
Conservation priorities for mammals in megadiverse Mexico: the efficiency of reserve networks
Ecological Applications
17
2007
569-78
Ceballos y Brown, 1995
- Ceballos
J.H. Brown
Global patterns of mammalian diversity, endemism, and endangerment
Conservation Biology
9
1995
559-68
Ceballos y Ehrlich, 2006
- Ceballos
P.R. Ehrlich
Global mammal distributions, biodiversity hotspots, and conservation
Proceedings of the National Academy of Sciences
103
2006
19374-9
Ceballos y Navarro, 1991
- Ceballos
- Navarro
Diversity and conservation of Mexican mammals
Latin American mammalogy: history, diversity and conservation
University of Oklahoma Press
Norman, Oklahoma
1991
167-98
Ceballos y Rodríguez, 1993
- Ceballos
- Rodríguez
Diversidad y conservación de mamíferos de México. II. Patrones de endemicidad de mamíferos mexicanos
Avances en el estudio de los mamíferos de México
Asociación Mexicana de Matozoología, A.C.
México D.F.
1993
87-108
Ceballos et al., 1998
- Ceballos
- Rodríguez
R.A. Medellín
Assessing conservation priorities in megadiverse Mexico: mammalian diversity, endemicity, and endangerment
Ecological Applications
8
1998
8-17
Christen y Soberón, 2009
- Christen
- Soberón
Anidamiento y los análisis Rq y Qr en PAMs
Miscelánea Matemática
49
2009
51-61
Diniz-Filho et al., 2013
J.A.F. Diniz-Filho
S.F. Gouveia
M.S. Lima-Ribeiro
Evolutionary macroecology
Frontiers of Biogeography
5
2013
195-203
Escalante et al., 1993
- Escalante
A.G. Navarro-Sigüenza
A.T. Peterson
A geographic, historical, and ecological analysis of avian diversity in Mexico
Biological diversity of Mexico: origins and distribution
Oxford University Press
New York
1993
281-307
Escobar-Briones et al., 2008
E.G. Escobar-Briones
- Gaytán-Caballero
- Legendre
Epibenthic megacrustaceans from the continental margin, slope and abyssal plain of the Southwestern Gulf of Mexico: factors responsible for variability in species composition and diversity
Deep-Sea Research
55
2008
2667-78
Ezcurra y Equihua, 1984
- Ezcurra
- Equihua
Métodos cuantitativos en la biogeografía
1984
Flores-Villela, 1993
- Flores-Villela
Herpetofauna of Mexico: distribution and endemism
Biological diversity of Mexico: origins and distribution
Oxford University Press
Oxford
1993
243-80
Gallardo-Cruz et al., 2010
J.A. Gallardo-Cruz
J.A. Meave
E.A. Pérez-García
J.L. Hernández-Stefanoni
Spatial structure of plant communities in a complex tropical landscape: implications for β-diversity
Community Ecology
11
2010
202-10
García et al., 2007
- García
- Solano-Rodríguez
- Flores-Villela
Patterns of alpha, beta and gamma diversity for the herpetofaun of pacific lowlands and adjacent interior valleys of Mexico
Animal Biodiversity and Conservation
30
2007
169-77
Garrido-Olvera et al., 2012
- Garrido-Olvera
H.T. Arita
- Pérez-Ponce de León
The influence of host ecology and biogeography on the helminth species richness of freshwater fishes in Mexico
Parasitology
139
2012
1652-65
Gaston, 1990
K.J. Gaston
Patterns in the geographical ranges of species
Biological Review
65
1990
105-29
Gaston et al., 2008
K.J. Gaston
S.L. Chown
K.L. Evans
Ecogeographical rules: elements of a synthesis
Journal of Biogeography
35
2008
483-500
Goettsch y Hernández, 2006
- Goettsch
H.M. Hernández
Beta diversity and similarity among cactus assemblages in the Chihuahuan Desert
Journal of Arid Environments
65
2006
513-28
Golicher et al., 2007
D.J. Golicher
- Cayuela
J.R.M. Alkemade
- González-Espinosa
- Ramírez-Marcial
Applying climatically associated species pools to the modelling of compositional change in tropical montane forests
Global Ecology and Biogeography
17
2007
262-73
Golubov et al., 2005
- Golubov
M.C. Mandujano
- Mandujano
Diversidad alfa y beta en Opuntia y Agave
Sobre diversidad biológica: el significado de las diversidades alfa, beta y gamma
Sociedad Entomológica Aragonesa, Conabio, Diversitas, Conacyt
Zaragoza
2005
209-19
González-Espinosa et al., 2004
- González-Espinosa
J.M. Rey-Benayas
- Ramírez-Marcial
M.A. Huston
- Golicher
Tree diversity in the Northern Neotropics: regional patterns in highly diverse Chiapas, Mexico
Ecography
27
2004
741-56
González-Espinosa et al., 2005
- González-Espinosa
- Ramírez-Marcial
- Méndez-Dewar
- Galindo-Jaimes
- Golicher
Riqueza de especies de árboles en Chiapas: variación espacial y dimensiones ambientales asociadas al nivel regional
Diversidad biológica en Chiapas
Plaza y Valdés, ECOSUR, COCyTECH
México, D.F.
2005
81-125
Gotelli et al., 2009
N.J. Gotelli
M.J. Anderson
H.T. Arita
- Chao
R.K. Colwell
S.R. Connolly
Patterns and causes of species richness: a general simulation model for macroecology
Ecology Letters
12
2009
873-86
Halffter, 2007
- Halffter
Reservas archipiélago: un nuevo tipo de área protegida
Hacia una cultura de conservación de la diversidad biológica, Vol. 6
Monografías Tercer Milenio, S.E.A.
Zaragoza
2007
281-6
Halffter y Moreno, 2005
- Halffter
- Moreno
Significado biológico de las diversidades alfa, beta y gamma
Sobre diversidad biológica: el significado de las diversidades alfa, beta y gamma
Sociedad Entomológica Aragonesa, Conabio, Diversitas, Conacyt
Zaragoza
2005
5-15
Halffter et al., 2005
- Halffter
- Soberón
- Koleff
- Melic
Sobre diversidad biológica: el significado de las diversidades alfa, beta y gamma
2005
Hawkins y Diniz-Filho, 2006
B.A. Hawkins
J.A.F. Diniz-Filho
Beyond Rapoport’s rule: evaluating range size patterns of New World birds in a two-dimensional framework
Global Ecology and Biogeography
15
2006
461-9
Keith et al., 2012
S.A. Keith
T.J. Webb
- Böhning-Gaese
S.R. Connolly
N.K. Dulvy
- Eigenbrod
What is macroecology?
Biology Letters
8
2012
904-6
Koleff et al., 2003
- Koleff
K.J. Gaston
J.J. Lennon
Measuring beta diversity for presence-absence data
Journal of Animal Ecology
72
2003
367-82
Koleff et al., 2008
- Koleff
- Soberón
H.T. Arita
- Dávila
- Flores-Villela
- Golubov
Capital natural de México. Vol. 1. Conocimiento actual de la biodiversidad
2008
323-64
Lawton, 1999
J.H. Lawton
Are there general laws in ecology?
Oikos
84
1999
177-92
Lennon et al., 2001
J.J. Lennon
- Koleff
J.J.D. Greenwood
K.J. Gaston
The geographical structure of British bird distributions: diversity, spatial turnover and scale
Journal of Animal Ecology
70
2001
966-79
Lira-Noriega et al., 2013
- Lira-Noriega
- Soberón
C.P. Miller
Process-based and correlative modeling of desert mistletoe distribution: a multiscalar approach
Ecosphere
4
2013
1-23
Lira-Noriega et al., 2007
- Lira-Noriega
- Soberón
A.G. Navarro-Sigüenza
- Nakazawa
A.T. Peterson
Scale-dependency of diversity components estimated from primary biodiversity data and distribution maps
Diversity and Distributions
13
2007
185-95
López-Martínez et al., 2013
J.O. López-Martínez
- Sanaphre-Villanueva
J.M. Dupuy
J.L. Hernández-Stefanoni
J.A. Meave
J.A. Gallardo-Cruz
β-diversity of functional groups of woody plants in a tropical dry forest in Yucatan
Plos One
8
2013
e73660
MacArthur, 1972
R.H. MacArthur
Geographical ecology: patterns in the distribution of species
1972
Marquet, 2009
P.A. Marquet
Macroecological perspectives on communities and ecosystems
The Princeton guide to ecology
Princeton University Press
Princeton
2009
386-94
Martínez-Meyer et al., 2012
- Martínez-Meyer
- Díaz-Porras
A.T. Peterson
- Yáñez-Arenas
Ecological niche structure and rangewide abundance patterns of species
Biology Letters
9
2012
20120637
McCain, 2007
C.M. McCain
Could temperature and water availability drive elevational species richness patterns? A global case study for bats
Global Ecology and Biogeography
16
2007
1-13
McKnight et al., 2007
- McKnight
P.S. White
R.I. McDonald
J.F. Lamoreaux
- Sechrest
R.S. Ridgely
Putting beta diversity on the map: broad-scale congruence and coincidence in the extremes
Plos Biol
5
2007
e272
Mena y Vázquez-Domínguez, 2005
J.L. Mena
- Vázquez-Domínguez
Species turnover on elevational gradients in small rodents
Global Ecology and Biogeography
14
2005
539-47
Mendoza y Arita, 2014
A.M. Mendoza
H.T. Arita
Priority setting by sites and by species using rarity, richness and phylogenetic diversity: the case of neotropical glassfrogs (Anura: Centrolenidae)
Biodiversity and Conservation
23
2014
909-26
Moreno y Halffter, 2001
C.E. Moreno
- Halffter
Spatial and temporal analysis of α β and γ diversities of bats in a fragmented landscape
Biodiversity and Conservation
10
2001
367-82
Moreno y Rodríguez, 2010
C.E. Moreno
- Rodríguez
A consistent terminology for quantifying species diversity?
Oecologia
163
2010
279-82
Moreno y Rodríguez, 2011
C.E. Moreno
- Rodríguez
Commentary: Do we have a consistent terminology for species diversity? Back to basics and toward a unifying framework
Oecologia
167
2011
889-92
Mourelle y Ezcurra, 1997a
- Mourelle
- Ezcurra
Raporport’s rule: a comparative analysis between South and North American columnar cacti
American Naturalist
150
1997
131-42
Mourelle y Ezcurra, 1997b
- Mourelle
- Ezcurra
Differentiation diversity of Argentine cacti and its relationship to environmental factors
Journal of Vegetation Science
8
1997
547-58
Navarro y León-Paniagua, 1995
D.L. Navarro
- León-Paniagua
Community structure of bats along an altitudinal gradient in tropical eastern Mexico
Revista Mexicana de Mastozoología
1
1995
9-21
Navarro-Sigu¿enza et al., 2009
A.G. Navarro-Sigu¿enza
- Gordillo-Martínez
A.T. Peterson
Mapeando la diversidad de las aves de México
TIP Revista Especializada en Ciencias Químico-Biolo¿gicas
12
2009
91-5
Navarro-Sigu¿enza et al., 2011
A.G. Navarro-Sigu¿enza
- Lira-Noriega
M.C. Arizmendi
- Berlanga
- Koleff
- Garci¿a-Moreno
Áreas de conservacio¿n para las aves de México: integrando criterios de priorizacio¿n
Planeacio¿n para la conservacio¿n de la biodiversidad terrestre en México: retos en un pai¿s megadiverso
Conabio-Conanp
México D.F.
2011
109-29
Noguez et al., 2005
A.M. Noguez
H.T. Arita
A.E. Escalante
L.J. Forney
- García-Oliva
- Souza
Microbial macroecology: highly structured prokaryotic soil assemblages in a tropical deciduous forest
Global Ecology and Biogeography
14
2005
241-8
Ochoa-Ochoa et al., 2014
L.M. Ochoa-Ochoa
- Munguía
- Lira-Noriega
- Sánchez-Cordero
- Flores-Villela
- Navarro-Sigüenza
Spatial scale and β-diversity of terrestrial vertebrates in Mexico
Revista Mexicana de Biodiversidad
85
2014
918-30
Ochoa-Ochoa et al., 2012
- Ochoa-Ochoa
- Rodríguez
- Mora
- Flores-Villela
- Whittaker
Climate change and amphibian diversity patterns in Mexico
Biological Conservation
150
2012
94-102
Olalla-Tárraga et al., 2010
M.A. Olalla-Tárraga
L.M. Bini
J.A.F. Diniz-Filho
M.A. Rodríguez
Cross-species and assemblage-based approaches to Bergmann’s rule and the biogeography of body size in Plethodon salamanders of eastern North America
Ecography
33
2010
362-8
Ortega-Huerta y Peterson, 2001
- Ortega-Huerta
- Peterson
Modelling spatial patterns of biodiversity for conservation prioritization in north-eastern Mexico
Diversity and Distributions
10
2001
39-54
Osorio-Olvera et al., 2016
L.A. Osorio-Olvera
- Falconi
- Soberón
Sobre la relación entre idoneidad del hábitat y la abundancia poblacional bajo diferentes escenarios de dispersión
Revista Mexicana de Biodiversidad
87
2016
1080-8
Qiao et al., 2016
- Qiao
E.E. Saupe
- Soberón
A.T. Peterson
C.E. Myers
Impacts of niche breadth and dispersal ability on macroevolutionary patterns
The American Naturalist
188
2016
149-62
Quiroz-Martínez y Salgado-Maldonado, 2013
- Quiroz-Martínez
- Salgado-Maldonado
Taxonomic distinctness and richness of helminth parasite assemblages of freshwater fishes in Mexican hydrological basins
Plos One
8
2013
e74419
Parra-Olea et al., 2014
- Parra-Olea
- Flores-Villela
- Mendoza-Almeralla
Biodiversity of amphibians in Mexico
Revista Mexicana de Biodiversidad
85
Supl
2014
460-6
Pavón et al., 2000
N.P. Pavón
- Hernández-Trejo
- Rico-Gray
Distribution of plant life forms along an altitudinal gradient in the semi-arid valley of Zapotitlán, Mexico
Journal of Vegetation Science
11
2000
39-42
Pérez-Moreno et al., 2016
- Pérez-Moreno
- Martínez-Meyer
- Soberón-Mainero
- Rojas-Soto
Climatic patterns in the establishment of wintering areas by North American migratory birds
Ecology and Evolution
6
2016
2022-33
Peterson et al., 1993
A.T. Peterson
O.A. Flores-Villela
- León-Paniagua
J.E. Llorente-Bousquets
M.A. Luis-Martínez
A.G. Navarro-Siguenza
Conservation priorities in Mexico: moving up in the world
Biodiversity Letters
1
1993
33-8
Peterson et al., 2002
A.T. Peterson
M.A. Ortega-Huerta
- Sánchez-Cordero
- Soberón
R.H. Buddemeier
D.R. Stockwell
Future projections for Mexican faunas under global climate change scenarios
Nature
416
2002
626-9
Peterson y Soberón, 2012
A.T. Peterson
- Soberón
Species distribution modeling and ecological niche modeling: getting the concepts right
Naturaleza y Conservacao
10
2012
102-7
Peterson et al., 1999
A.T. Peterson
- Soberón
- Sánchez-Cordero
Conservatism of ecological niches in evolutionary time
Science
285
1999
1265-7
Pineda y Halffter, 2004
- Pineda
- Halffter
Species diversity and habitat fragmentation: frogs in a tropical montane landscape in Mexico
Biological Conservation
117
2004
499-508
Pineda y Lobo, 2009
- Pineda
J.M. Lobo
Assessing the accuracy of species distribution models to predict amphibian species richness patterns
Journal of Animal Ecology
78
2009
182-90
Pineda et al., 2005
- Pineda
C.E. Moreno
- Escobar
- Halffter
Frog, bat, and dung beetle diversity in the cloud forest and coffee agroecosystems of Veracruz, Mexico
Conservation Biology
19
2005
400-10
Pompa et al., 2011
- Pompa
P.R. Ehrlich
- Ceballos
Global distribution and conservation of marine mammals
PNAS
108
2011
13600-5
Raia y Fortelius, 2017
- Raia
- Fortelius
Introduction to evolutionary macroecology
Evolutionary Ecology Research
18
2017
1-6
Ramamoorthy et al., 1993
T.P. Ramamoorthy
- Bye
- Lot
- Fa
Biological diversity of Mexico: origins and distribution
1993
Rapoport, 1975
E.H. Rapoport
Areografía: estrategias geográficas de las especies
1975
Rapoport, 1982
E.H. Rapoport
Areography: geographical strategies of species
1982
Riemann y Ezcurra, 2005
- Riemann
- Ezcurra
Plant endemism and natural protected areas in the peninsula of Baja California, Mexico
Biological Conservation
122
2005
141-50
Rodríguez y Arita, 2004
- Rodríguez
H.T. Arita
Beta diversity and latitude in North American mammals: testing the hypothesis of covariation
Ecography
27
2004
547-56
Rodríguez et al., 2003
- Rodríguez
- Soberón
H.T. Arita
El componente beta de la diversidad de mamíferos de México
Acta Zoológica Mexicana
89
2003
1-19
Rosales-Nanduca et al., 2011
- Rosales-Nanduca
- Gerrodette
- Urbán-R
- Cárdenas-Hinojosa
- Medrano-González
Macroecology of marine mammal species in the Mexican Pacific Ocean: diversity and distribution
Marine Ecology Progress Series
431
2011
281-91
Rosenzweig, 1995
M.L. Rosenzweig
Species diversity in space and time
1995
Sánchez-Cordero, 2001
- Sánchez-Cordero
Elevation gradients of diversity for rodents and bats in Oaxaca, Mexico
Global Ecology and Biogeography
10
2001
63-76
Sarmiento y Monasterio, 1971
- Sarmiento
- Monasterio
Ecología de las sabanas de América tropical. I. Análisis macroecológico de los Llanos de Calabozo, Venezuela
Cuadernos Geográficos
4
1971
1-126
Sarukhán y Jiménez, 2016
- Sarukhán
- Jiménez
Generating intelligence for decision making and sustainable use of natural capital in Mexico
Sustainability Science
19
2016
153-9
Sarukhán et al., 1996
- Sarukhán
- Soberón
- Larson-Guerra
Biological conservation in a high betadiversity country
Biodiversity, science and development: towards a new partnership
CABI/UNESCO
Wallingford
1996
246-63
Saupe et al., 2015
E.E. Saupe
- Qiao
J.R. Hendricks
R.W. Portell
S.J. Hunter
- Soberón
Niche breadth and geographic range size as determinants of species survival on geological time scales
Global Ecology and Biogeography
24
2015
1159-69
Soberón, 2007
- Soberón
Grinnellian and Eltonian niches and geographic distributions of species
Ecology Letters
10
2007
1115-23
Soberón, 2015
- Soberón
Pairwise versus presence-absence approaches for analyzing biodiversity patterns
Journal of Biogeography
42
2015
807-8
Soberón y Cavner, 2015
- Soberón
- Cavner
Indices of biodiversity pattern based on presence-absence matrices: a GIS implementation
Biodiversity Informatics
10
2015
22-34
Soberón y Ceballos, 2011
- Soberón
- Ceballos
Species richness and range size of the terrestrial mammals of the World: biological signal within mathematical constraints
Plos One
6
2011
e19359
Stevens, 1989
G.C. Stevens
The latitudinal gradient in geographical range: how so many species coexist in the tropics
American Naturalist
133
1989
240-56
Trejo-Barocio y Arita, 2013
- Trejo-Barocio
H.T. Arita
The co-occurrence of species and the co-diversity of sites in neutral models of biodiversity
Plos One
8
2013
e79918
Vázquez y Gaston, 2004
L.B. Vázquez
K.J. Gaston
Rarity, commonness, and patterns of species richness: the mammals of Mexico
Global Ecology and Biogeography
13
2004
535-42
Vázquez et al., 2008
L.B. Vázquez
- Rodríguez
H.T. Arita
Conservation planning in a subdivided world
Biodiversity and Conservation
17
2008
1367-77
Vázquez y Valenzuela-Galván, 2009
L.B. Vázquez
- Valenzuela-Galván
¿Qué tan bien representados están los mamíferos mexicanos en la red federal de áreas naturales protegidas del país?
Revista Mexicana de Biodiversidad
80
2009
249-58
Velasco et al., 2016
J.A. Velasco
- Martínez-Meyer
- Flores-Villela
- García
A.C. Algar
- Köhler
Climatic niche attributes and diversification in Anolis lizards
Journal of Biogeography
43
2016
134-44
Vilela y Villalobos, 2015
- Vilela
- Villalobos
letsR: a new R package for data handling and analysis in macroecology
Methods in Ecology and Evolution
6
2015
1229-34
Villalobos y Arita, 2010
- Villalobos
H.T. Arita
The diversity field of New World leaf-nosed bats (Phyllostomidae)
Global Ecology and Biogeography
19
2010
200-11
Villalobos et al., 2016
- Villalobos
- Carotenuto
- Raia
J.A.F. Diniz-Filho
Phylogenetic fields through time: temporal dynamics of geographic co-occurrence and phylogenetic structure within species ranges
Philosophical Transactions of the Royal Society B
371
2016
20150220
Villalobos et al., 2016
- Villalobos
M.V. Cianciaruso
M.A. Olalla-Tárraga
T.F. Rangel
J.A.F. Diniz-Filho
Global patterns of mammalian co-occurrence: phylogenetic and body size structure within species ranges
Journal of Biogeography
44
2016
136-46
Villalobos, Dobrovolsk et al., 2013
- Villalobos
- Dobrovolsk
D.B. Provete
S.F. Gouveia
Is rich and rare the common share? Describing biodiversity patterns to inform conservation practices for South American anurans
Plos One
8
2013
e56073
Villalobos, Lira-Noriega et al., 2013
- Villalobos
- Lira-Noriega
- Soberón
H.T. Arita
Range-diversity plots for conservation planning: using richness and rarity in priority setting, the case of Mexican avifauna
Biological Conservation
158
2013
313-20
Villalobos et al., 2014
- Villalobos
- Lira-Noriega
- Soberón
H.T. Arita
Co-diversity and co-distribution in phyllostomid bats: evaluating the relative roles of climate and niche conservatism
Basic and Applied Ecology
15
2014
85-91
Villalobos y Rangel, 2014
- Villalobos
T.F. Rangel
Geographic patterns of biodiversity: macroecological approaches for a complex phenomenon
Frontiers in ecology, evolution and complexity
CopIt-arXives
Ciudad de México
2014
1-11
Villaseñor, 2003
J.L. Villaseñor
Diversidad y distribución de las Magnoliophyta de México
Interciencia
28
2003
160-7
Whittaker, 1960
R.H. Whittaker
Vegetation of the Siskiyo Mountains, Oregon and California
Ecological Monograph
30
1960
279-338
Whittaker, 1972
R.H. Whittaker
Evolution and measurement of species diversity
Taxon
21
1972
213-51
Yáñez-Arenas et al., 2012
- Yáñez-Arenas
- Martínez-Meyer
- Mandujano
- Rojas-Soto
Modelling geographic patterns of population density of the white-tailed deer in central Mexico by implementing ecological niche theory
Oikos
121
2012
2081-9
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.
*
Autor para correspondencia.
Copyright © 2017 Universidad Nacional Autónoma de México, Instituto de Biología.