
Judith A. Sánchez-Ledesma a, Roberto Garibay-Orijel b, *, Gonzalo Guevara-Guerrero c, Verónica Ávila-Rodríguez d y Jesús G. Arreola-Ávila a
a Universidad Autónoma Chapingo, Unidad Regional Universitaria de Zonas Áridas, Km. 40.5 Carretera Gómez Palacio-Chihuahua, 35230 Bermejillo, Durango, México
b Universidad Nacional Autónoma de México, Instituto de Biología, Circuito exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, México
c Instituto Tecnológico de Cd. Victoria, Av. Portes Gil 1301 Pte., 87010 Cd. Victoria, Tamaulipas, México
d Universidad Juárez del Estado de Durango, Facultad de Ciencias Biológicas, 35010 Gómez Palacio, Durango, México
*Autor para correspondencia: rgaribay@ib.unam.mx (R. Garibay-Orijel)
Recibido: 15 junio 2021; aceptado: 23 agosto 2022
Resumen
Los agroecosistemas, como los cultivos de Carya illinoinensis (nogal pecanero), representan una actividad económica importante y de constante crecimiento en La Comarca Lagunera, México. La mayor parte de los macromicetos que se asocian con el nogal pecanero se distribuyen en los primeros horizontes del suelo y la rizosfera de las plantas. Para recolectar los macromicetos asociados al nogal se realizaron recorridos 2 veces por semana, de mayo a octubre de 2018, cubriendo en total 25 ha de una huerta en La Comarca Lagunera, México. A los hongos recolectados se les secuenció la región ITS del ADN ribosomal para corroborar su identificación taxonómica por medio de análisis filogenéticos de máxima verosimilitud y bayesianos. Se encontraron 5 taxones asociados al nogal pecanero, 4 identificados a nivel de especie (Agaricus deserticola, Candolleomyces luteopallidus, Tuber caryophilum y Scleroderma cepa s.l.) y una especie potencialmente nueva (Candolleomyces sp.). De éstos, C. luteopallidus se registra por primera vez en México. Se discute la importancia de la diversidad de los hongos en este sistema agroforestal ya que los hongos saprobios descomponen la materia orgánica y los ectomicorrízicos transfieren macro y micronutrientes esenciales para el crecimiento y productividad del nogal.
Palabras clave: Agroecosistemas; Diversidad de hongos; Candolleomyces; Tuber; Nogal pecanero
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Carya illinoinensis macrofungi in La Comarca Lagunera, Mexico
Abstract
Agroecosystems, such as Carya illinoinensis (pecan tree) orchards, represent an important and constantly growing economic activity in La Comarca Lagunera, Mexico. Most of the macromycetes associated with pecan tree are distributed in the first soil horizons and the plants rhizosphere. To collect the macromycetes associated with pecan trees, we conducted twice a week surveys from May to October 2018 covering 25 hectares of an orchard in La Comarca Lagunera, Mexico. To corroborate their taxonomic identification, we sequenced the DNA ITS region and performed Maximum Likelihood and Bayesian phylogenetic analyses. We found 5 species of macromycetes associated with pecan trees, 4 identified to species level (Agaricus deserticola, Candolleomyces luteopallidus, Tuber caryophylum, and Scleroderma cepa s.l.) and one potential new species (Candolleomyces sp.). Of these, C. luteopallidus is recorded for the first time in Mexico. The importance of fungal diversity in this agroforestry system is discussed because saprobe fungi decompose organic matter and ectomycorrhizal fungi transfer macro and micronutrients essential for pecan trees growth and productivity.
Keywords: Agroecosystems; Fungal diversity; Candolleomyces; Tuber; Pecan tree
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
Carya illinoinensis (Wangenh.) K.Koch (nogal pecanero) es la especie económicamente más importante del género Carya (Casales et al., 2018). Esta especie es nativa del sur de EUA, donde abunda en la cuenca del Mississippi en bosques de latifoliadas, principalmente encinos y también se distribuye en el norte de México, siendo más rara hacia el sur asociada a riveras (Bonito et al., 2011). Se cultiva principalmente por su nuez que es fuente de aceites, proteínas y exquisito sabor, siendo pocos los alimentos que superan su riqueza y digestibilidad (Villasante et al., 2019). La distribución de las especies de Carya al norte de México es el resultado de una buena adaptación ambiental. Al desarrollarse en zonas áridas tienen una menor densidad estomática foliar como un mecanismo adaptativo para reducir las pérdidas de agua por evapotranspiración (Sagaram et al., 2011). En la actualidad, en Coahuila y Durango, en la región conocida como Comarca Lagunera, el valor de la producción de nuez pecana ocupa el primer lugar entre los frutales de la zona, con un valor aproximado de 8,000 millones de pesos. La superficie plantada en 2020 fue de 103,711 ha con una producción de 116,224 toneladas, predominando las variedades Western y Wichita (SIAP, 2021).
El nogal pecanero crece de manera natural en bosques de latifoliadas, principalmente de encinos, con quienes comparte numerosos hongos ectomicorrízicos. En huertas comerciales presenta asociaciones simbióticas con hongos ectomicorrízicos altamente especializados como: Astraeus, Gyrodon, Pisolithus, Russula, Scleroderma, Tuber y Tylopilus (Marx, 1971; Taber, 1984). El único trabajo que ha comparado la diversidad de hongos ectomicorrízicos entre nogal pecanero de huertos y de árboles nativos adyacentes es el de Ge et al. (2017). Ellos encontraron que los árboles nativos están dominados por los linajes: russula-lactarius (12 especies), tomentella-thelephora (7 especies), inocybe (4 especies), /tuber-helvella (3 especies) y amanita (3 especies). Dos terceras partes de estas especies también se encontraron en las raíces del nogal dentro de las huertas. De la diversidad de hongos asociados al nogal pecanero, destacan las especies de Tuber, de las que al menos T. brennemanii y T. floridanum son endémicas de huertas de nogal (Grupe et al., 2018). Estos hongos ectomicorrízicos proporcionan diferentes beneficios en términos de sobrevivencia, nutrición y productividad al nogal.
En huertas de nogal pecanero se han encontrado también hongos saprobios que juegan un papel fundamental en el funcionamiento de este agroecosistema, pues descomponen la materia orgánica y reciclan los nutrientes en el suelo, constituyendo así un recurso forestal integral (Yun y Hall, 2004). Los macromicetos en las huertas de nogal pecanero están distribuidos en los horizontes superficiales del suelo y en la rizosfera de los árboles, manteniendo el funcionamiento, estructura y equilibro en las huertas (Córdoba-Chávez et al., 2014). Por lo tanto, es de suma importancia reconocer el valor ecológico de estos hongos, estudiar su diversidad y distribución (Ramírez, 2013). Este conocimiento se podría aprovechar para promover el crecimiento del nogal y favorecer su adaptación a los diferentes ambientes y condiciones climáticas (Tovar et al., 2004).
Materiales y métodos
Muestreo, caracterización e identificación morfológica
Para la realización de esta investigación se trabajó en un huerto de Carya illinoinensis de La Comarca Lagunera, situada en el municipio de Viesca, Coahuila (25°25’ N, 103°18’ O; 1,200 m snm) (fig. 1). El huerto se maneja de acuerdo al paquete tecnológico de producción de nogal pecanero (INIFAP, 2002), incluyendo el control de malezas por la técnica de “chapoleo” con licuadora mecánica, excluyendo así las prácticas de rastreo y movimiento de suelo. El huerto cuenta con un sistema de plantación marco real 12 × 12 m, un sistema de riego por goteo subsuperficial y un suelo de textura franco —arcillosa, con una capacidad de campo de 0.35 m3/m3 y un punto de marchitez permanente de 0.19 m3/m3. En esta localidad se realizaron recorridos en bloques de 5 km2 en 5 sitios, se realizó la búsqueda y recolección de ejemplares de hongos que crecen espontáneamente, durante 9 meses, comprendidos entre mayo y noviembre de 2018. Los cuerpos fructíferos de los hongos se recolectaron haciendo una revisión de ramas, arbustos, hojarasca y suelo. Cada espécimen se depositó en papel encerado marcado con sus datos de recolecta. Para cada recolecta se realizó registro fotográfico y observaciones como tamaño, color, forma, etc. Los colores de los hongos se registraron usando la guía de colores de Munsell (1954). Para identificar las estructuras microscópicas se analizaron preparaciones temporales con KOH al 5% y reactivo de Melzer en microscopios Velab VE–B3 y ZEISS Scope A1. Se identificaron los ejemplares de acuerdo a los trabajos taxonómicos de Kerrigan (2016), Wächter y Melzer (2020), Guzmán et al. (2013), Healy et al. (2016) y Sánchez-Ledesma et al. (2022). Para el arreglo taxonómico se siguió la clasificación del catálogo de autoridades taxonómicas de los hongos en México (Conabio, 2014) y la clasificación de Index Fungorum (http://www.indexfungorum.org). Se complementaron las identificaciones taxonómicas con análisis filogenéticos para los que se secuenció el ADN de 1 o 2 ejemplares por cada morfoespecie.

Actualmente, el uso de códigos de barras genéticos es una herramienta en la identificación rápida de especies que tiene como propósito, el uso sustentable, la conservación y el conocimiento de la biodiversidad (Bickford et al., 2007; Paz et al., 2011). En este trabajo, además de generar los códigos de barras genéticos, usamos las secuencias de ADN para complementar las identificaciones taxonómicas con análisis filogenéticos para cada uno de los géneros encontrados.
El ADN fue extraído con el método de CTAB y amplificado por PCR en reacciones de 25 µl de acuerdo a Sambrook et al. (1989). Las reacciones consistieron en 2.5 µl de buffer de PCR 10X, 2.0 µl de MgCl 2.5 Mm concentración final, 2.0 µl de dNTPs (CA, USA) 2.0 µl de cada primer de 10 picomoles/µl concentración final, 0.2 µl, (1.5 U) de Taq polimersa (GoTaq®, Flexi ADN Polymerase, Promega, WI), 11.3 µl de agua de grado MiliQ y 3 µl de ADN. La región de los interespaciadores ribosomales (ITS) se amplificó con los oligonucleótidos ITS4 e ITS5 (Vrain et al., 1992). El programa de PCR consistió en un ciclo inicial a 94 °C por 3 min, seguido de 34 ciclos de las temperaturas 94, 51 y 72 °C durante 1 min y una extensión final a 72 °C durante 8 min. La amplificación se llevó a cabo en un termociclador MiniAmp Plus Thermal Cycler.
El ADN, así como los productos de PCR fueron revisados en geles de agarosa al 0.8 y 1.5%, respectivamente. Se utilizó como amortiguador TBE al 0.5% (tris base, ácido bórico, EDTA al 0.5M, pH 8.0). Las muestras fueron teñidas con Gel Red® y se empleó un marcador de peso molecular de 100 pb (Axygen) como referencia. Los geles fueron fotografiados en un fotodocumentador Multidoc-IT (UVP®, Analytik Jena Company, CA).
Tabla 1
Lista de secuencias de ADN ITS utilizadas en los análisis filogenéticos. Las secuencias generadas en este trabajo están en negritas.
| Taxón | GenBank | Referencia |
| Agaricus aridicola | KT951331 | Zhao y Zhou, 2015 |
| Agaricus aridicola | JF797195 | Zhao y Zhou, 2015 |
| Agaricus arrillagarum tipo | NR158301 | |
| Agaricus arvensis | AJ887993 | Didukh y Vilgalys, 2005 |
| Agaricus augustus | JF797193 | Vu y Groenewald, 2017 |
| Agaricus augustus | MH854667 | Vu y Groenewald, 2017 |
| Agaricus augustus | MH859051 | Vu y Groenewald, 2017 |
| Agaricus augustus | KJ847461 | Vu y Groenewald, 2017 |
| Agaricus bellanniae tipo | NR144990 | |
| Agaricus comtulus | KM248904 | Berube y Maaref, 2014 |
| Agaricus deserticola | KM349613 | Kerrigan, 2014 |
| Agaricus deserticola | JF896228 | Kerrigan y Callac, 2011 |
| Agaricus deserticola | HM488747 | Vellinga y Sysouphanthong, 2011 |
| Agaricus deserticola | MZ092920 | Sánchez-Ledesma et al., 2023 |
| Agaricus didymus tipo | NR144990 | |
| Agaricus diminutivus | AY484681 | Geml y Geiser, 2004 |
| Agaricus diminutivus | KM248905 | Berube y Maaref, 2014 |
| Agaricus dulcidulus | KF447894 | Parra, 2013 |
| Agaricus edmondoi | KF447902 | Parra, 2013 |
| Agaricus edmondoi | KT951326 | Zhao y Zhou, 2015 |
| Agaricus evertens | MK550894 | Clements, 2020 |
| Agaricus evertens | MT303143 | Clements, 2020 |
| Agaricus fiardii | HM862464 | Kerrigan, 2010 |
| Agaricus friesianus | KT951316 | Zhao y Zhou, 2015 |
| Agaricus friesianus | KJ877787 | Kerrigan, 2014 |
| Agaricus gemellatus | KJ859081 | Kerrigan, 2014 |
| Agaricus gemellatus | KJ859082 | Kerrigan, 2014 |
| Agaricus jacobi tipo | NR119951 | |
| Agaricus julius tipo | NR144991 | |
| Agaricus kerriganii | KT951333 | Zhao y Zhou, 2015 |
| Agaricus kerriganii | MN992293 | Landry y Berube, 2020 |
| Agaricus kerriganii | MF954612 | Berbee y Bazzicalupo, 2017 |
| Agaricus kerriganii | KT951306 | Zhao y Zhou, 2015 |
| Agaricus lanipes | JF97190 | Didukh y Vilgalys, 2005 |
| Agaricus longistipes tipo | NR151748 | |
| Agaricus marisae | JF797182 | Callac y Parra, 2011 |
| Agaricus matrum | KT951310 | Zhao y Zhou, 2015 |
| Agaricus megalosporus tipo | NR119951 | |
| Agaricus parvibrunneus | MG137001 | He y Hyde, 2017 |
| Tabla 1. Continúa | ||
| Taxón | GenBank | Referencia |
| Agaricus purpulesquameus tipo | NR157484 | Bellchambers y Alvarado, 2018 |
| Agaricus purpurellus | MH620768 | Bellchambers y Alvarado, 2018 |
| Agaricus purpurellus | KU975076 | Callac y Parra, 2011 |
| Agaricus purpurellus | KF447903 | Parra, 2013 |
| Agaricus sandianus tipo | NR144995 | |
| Agaricus sp. | KM349614 | Kerrigan, 2014 |
| Agaricus sp. | KX657047 | He y Chen, 2017 |
| Agaricus sp. | JF691540 | Zhao y Karunarathna, 2011 |
| Agaricus wariatodes | JF495054 | Zhao y Karunarathna, 2011 |
| Candolleomyces badhyzensis tipo | KC992883 | Larsson y Orstadius, 2013 |
| Candolleomyces badiophylus | FN430699 | Nagy, 2011 |
| Candolleomyces bivelatus | MF325962 | Garbelotto y Dovana, 2017 |
| Candolleomyces cacao tipo | NR_148106 | Desjardin y Perry, 2017 |
| Candolleomyces candolleanus tipo | KM030175 | Larsson, 2014 |
| Candolleomyces cladii-marisci | MK080112 | Sicoli y Passalacqua, 2019 |
| Candolleomyces efflorescens | KC992941 | Larsson y Orstadius, 2013 |
| Candolleomyces halophilus | MG825900 | Broussal y Mir, 2018 |
| Candolleomyces hymenocephalus | FJ168609 | Paiva de Carvalho y Mesquita, 2018 |
| Candolleomyces leucotephrus | KC992885 | Larsson y Orstadius, 2013 |
| Candolleomyces littoralis | MG825901 | Broussal y Mir, 2018 |
| Candolleomyces luteopallidus | MG734736 | Yan y Bau, 2018 |
| Candolleomyces luteopallidus | MZ092918 | Sánchez-Ledesma et al., 2023 |
| Candolleomyces luteopallidus tipo | KC992884 | Larsson y Orstadius, 2013 |
| Candolleomyces pseudocandolleanus | KY120973 | Bau y Yan, 2016 |
| Candolleomyces rogueianus | MW412407 | Gordon, 2020 |
| Candolleomyces sp. | MZ092922 | Sánchez-Ledesma et al., 2023 |
| Candolleomyces sp. | MZ092923 | Sánchez-Ledesma et al., 2023 |
| Candolleomyces sp. BAB-4828 | KU571537 | Joshi y Bhatt, 2016 |
| Candolleomyces sp. BAB-4913 | KR155010 | Patel y Bhatt, 2015 |
| Candolleomyces sp. BAB-4964 | KR155054 | Patel y Bhatt, 2015 |
| Candolleomyces subsingeri | MG734725 | Yan y Bau, 2018 |
| Candolleomyces subsingeri tipo | NR_160505 | Yan y Bau, 2018 |
| Candolleomyces sulcatotuberculosus | KJ138422 | Larsson, 2014 |
| Candolleomyces trinitatensis | KC992882 | Larsson y Orstadius, 2013 |
| Candolleomyces tuberculatus | KC992886 | Larsson y Orstadius, 2013 |
| Candolleomyces typhae | DQ389721 | Larsson y Orstadius, 2013 |
| Scleroderma areolatum | EU819438 | Palmer y Lindner, 2008 |
| Scleroderma areolatum | EU819518 | Palmer y Lindner, 2008 |
| Scleroderma areolatum | FM213351 | Phosri y Martin, 2009 |
| Scleroderma areolatum | FM213352 | Phosri y Martin, 2009 |
| Scleroderma areolatum | FM213353 | Phosri y Martin, 2009 |
| Scleroderma bovista | EU819517 | Palmer y Lindner, 2008 |
| Scleroderma bovista | AB211267 | Nara, 2005 |
| Scleroderma bovista | GQ267487 | Walbert y Ramsfield, 2010 |
| Scleroderma cepa | FM213354 | Phosri y Martin, 2009 |
| Scleroderma cepa | MT005950 | Scholler, 2020 |
| Scleroderma cepa s.l. | MZ092921 | Sánchez-Ledesma et al., 2023 |
| Scleroderma citrinum | EU784414 | Brock y Doring, 2009 |
| Scleroderma citrinum | GQ166907 | Avis y Leacock, 2009 |
| Scleroderma dictyosporum | FJ840443 | Sanon y Ba, 2009 |
| Scleroderma dictyosporum | FJ840449 | Sanon y Ba, 2009 |
| Scleroderma laeve | EU718117 | Wilson y Binder, 2008 |
| Scleroderma macalpinei | EU718122 | Wilson y Binder, 2008 |
| Scleroderma meridionale | EU718121 | Wilson y Binder, 2008 |
| Scleroderma michiganese | EU819441 | Palmer y Lindner, 2008 |
| Scleroderma michiganese | FM213347 | Phosri y Martin, 2009 |
| Scleroderma patagonicum | HQ688788 | Nouhra y Hernandez Caffot, 2012 |
| Scleroderma patagonicum | HQ688789 | Nouhra y Hernandez Caffot, 2012 |
| Scleroderma polyrhizum | FM213349 | Martin, 2009 |
| Scleroderma polyrhizum | FM213350 | Phosri y Martin, 2009 |
| Scleroderma septentrionale | FM213336 | Phosri y Martin, 2009 |
| Scleroderma sinnamariense | FM213356 | Phosri y Martin, 2009 |
| Scleroderma sp. | MG211107 | Yuan y Jin, 2017 |
| Scleroderma sp. JZBHM008 | KM048204 | Xu y Li, 2014 |
| Scleroderma sp. UNSCL7 | FM213343 | Phosri y Martin, 2009 |
| Scleroderma suthepense tipo | NR_132871 | Kumla y Suwannarach, 2012 |
| Scleroderma verrucosum | FJ840461 | Sanon y Ba, 2009 |
| Scleroderma verrucosum | AJ629886 | Phosri y Martin, 2009 |
| Tuber lyonii | EU268568 | Bruhn y Pruett, 2007 |
| Tuber lyonii | FJ748911 | Bonito y Vilgalys, 2009 |
| Tuber lyonii | MT156465 | Smith y Healy, 2020 |
| Tuber lyonii | MT156509 | Smith y Healy, 2020 |
| Tuber lyonii | EF202594 | Bruhn y Pruett, 2007 |
| Tuber lyonii | EU268567 | Bruhn y Pruett, 2007 |
| Tuber caryophilum tipo | MZ092919 | Sánchez-Ledesma et al., 2022 |
| Tuber sp. 11 GB-2010 | GQ221453 | Murat y Jeandroz, 2009 |
| Tuber sp. 64 GB-2010 | HM485425 | Bonito y Gryganskyi, 2010 |
| Tuber sp. 65 | JQ925648 | Bonito y Gryganskyi, 2010 |
| Tuber sp. 65 GB-2010 | HM455426 | Bonito y Gryganskyi, 2010 |
| Tuber sp. 66 GB-2010 | HM485427 | Bonito y Gryganskyi, 2010 |
| Tuber sp. 79 | JQ925649 | Bonito y Smith, 2013 |
| Tuber sp. GB-2009e | FJ809887 | Bonito y Trappe, 2009 |
| Tuber sp. scr709 | DQ974798 | Smith y Douhan, 2007 |
| Tuber sp. SOC1404 | JN022530 | Frank y Southworth, 2011 |
| Tuber sp. voucher FLAS: MES-646 | MT156470 | Smith y Healy, 2020 |
| Tuber texense | HM485391 | Bonito y Gryganskyi, 2010 |
| Tuber umbillucatum | FJ797879 | Frank y Southworth, 2011 |
| Muestra ambiental de ectomicorriza | GU907784 | Karpati y Handel, 2011 |
Los productos de PCR se secuenciaron en ambos sentidos en la compañía Macrogen Corp. (Rockville, MD), con los mismos primers de la PCR. Las secuencias nucleotídicas fueron editadas y alineadas en Geneious Prime® versión 2021, con el algoritmo de MUSCLE (Edgar, 2004).
Los alineamientos fueron revisados de forma manual y las regiones ambiguas se excluyeron. Para los análisis filogenéticos se usaron las secuencias de ADN más cercanas obtenidas de la base de datos del Centro Nacional de Información Biotecnológica (NCBI), por medio del algoritmo BLAST (Altschul et al., 1990). También se incluyeron secuencias de referencia incluidas en análisis filogenéticos recientes de los géneros. El análisis de inferencia bayesiana se realizó en Mr.Bayes 3.2.5 (Huelsenbeck y Ronquist, 2001). El análisis de máxima verosimilitud se realizó con RAxML.7.2.6 (Stamatakis, 2006). El modelo de substitución de nucleótidos se eligió por los criterios de información de Akaike en jModelTest (Posada, 2008). En todos los casos, el análisis bayesiano se realizó con un modelo sustitución de nucleótidos JC69, radio de variación gamma, 4 cadenas de Montecarlo, 2 millones de generaciones, muestreando cada 200, descartando el primer 20% y calculando las probabilidades posteriores bayesianas (PPB) para el soporte de las ramas. El análisis de máxima verosimilitud utilizó el modelo de sustitución de nucleótidos JC69 con 1,000 réplicas de bootstrap (MLB). Para Candolleomyces, Agaricus sección arvenses, Tuber sección rufum y Scleroderma, el árbol se enraizó con C. typhae DQ389721, Agaricus sp. KY366488, Tuber sp. JQ925649 y Scleroderma polyrhizum FM213349, respectivamente. En todos los géneros se consideró informativo cuando el valor de bootstrap fue ≥ 70% y la probabilidad posterior fue ≥ 0.9. Las secuencias generadas en este estudio están disponibles en GenBank bajo los números de acceso MZ092918-MZ092923 (tabla 1).
Resultados
Basados tanto en caracteres morfológicos como análisis filogenéticos, se encontraron 5 especies de hongos en la huerta de nogal pecanero, 4 identificadas a nivel de especie (Agaricus deserticola, Candolleomyces luteopallidus, Tuber caryophilum y Scleroderma cepa s.l.) y una especie potencialmente nueva (Candolleomyces sp.) (fig. 2).

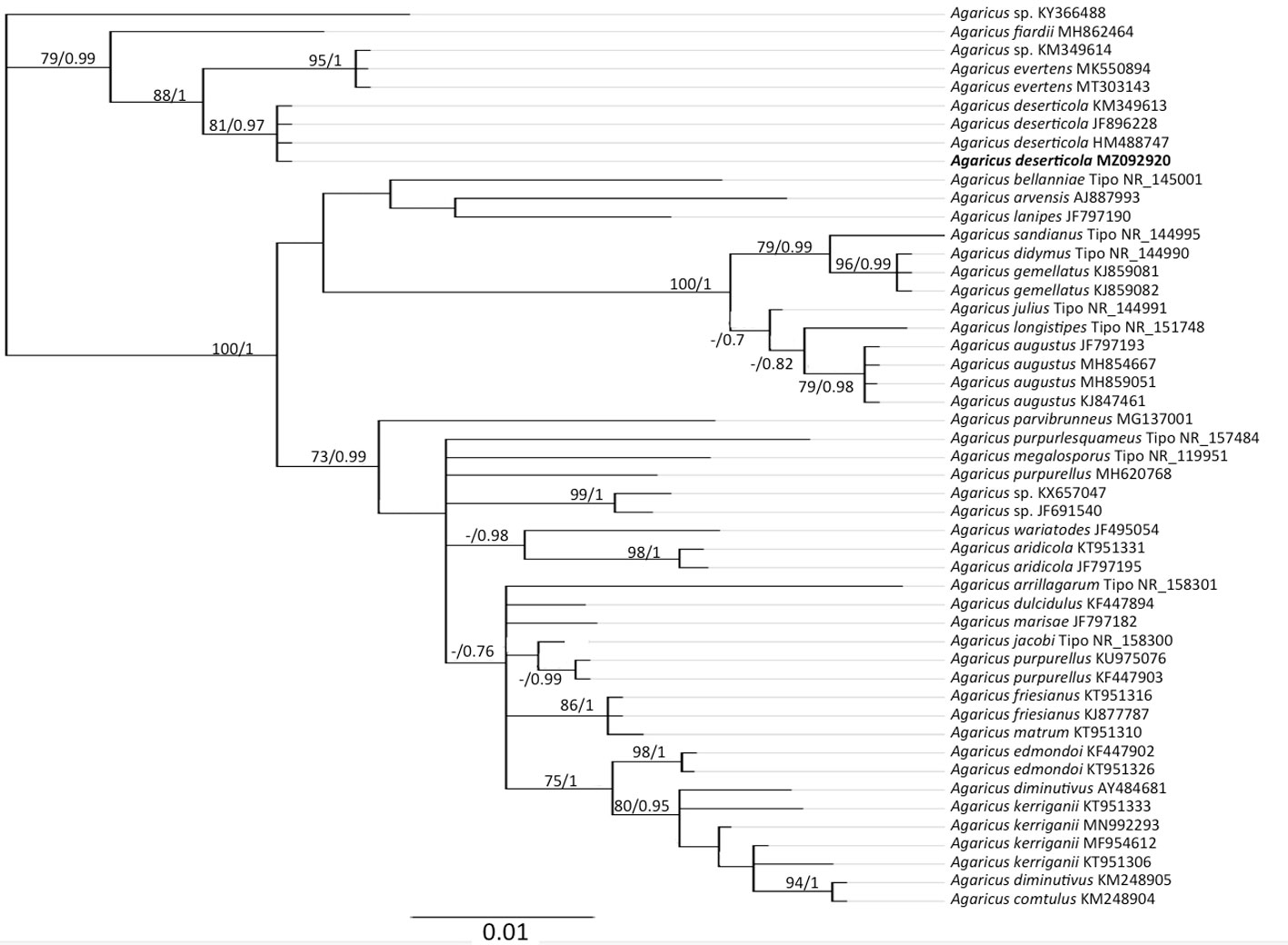

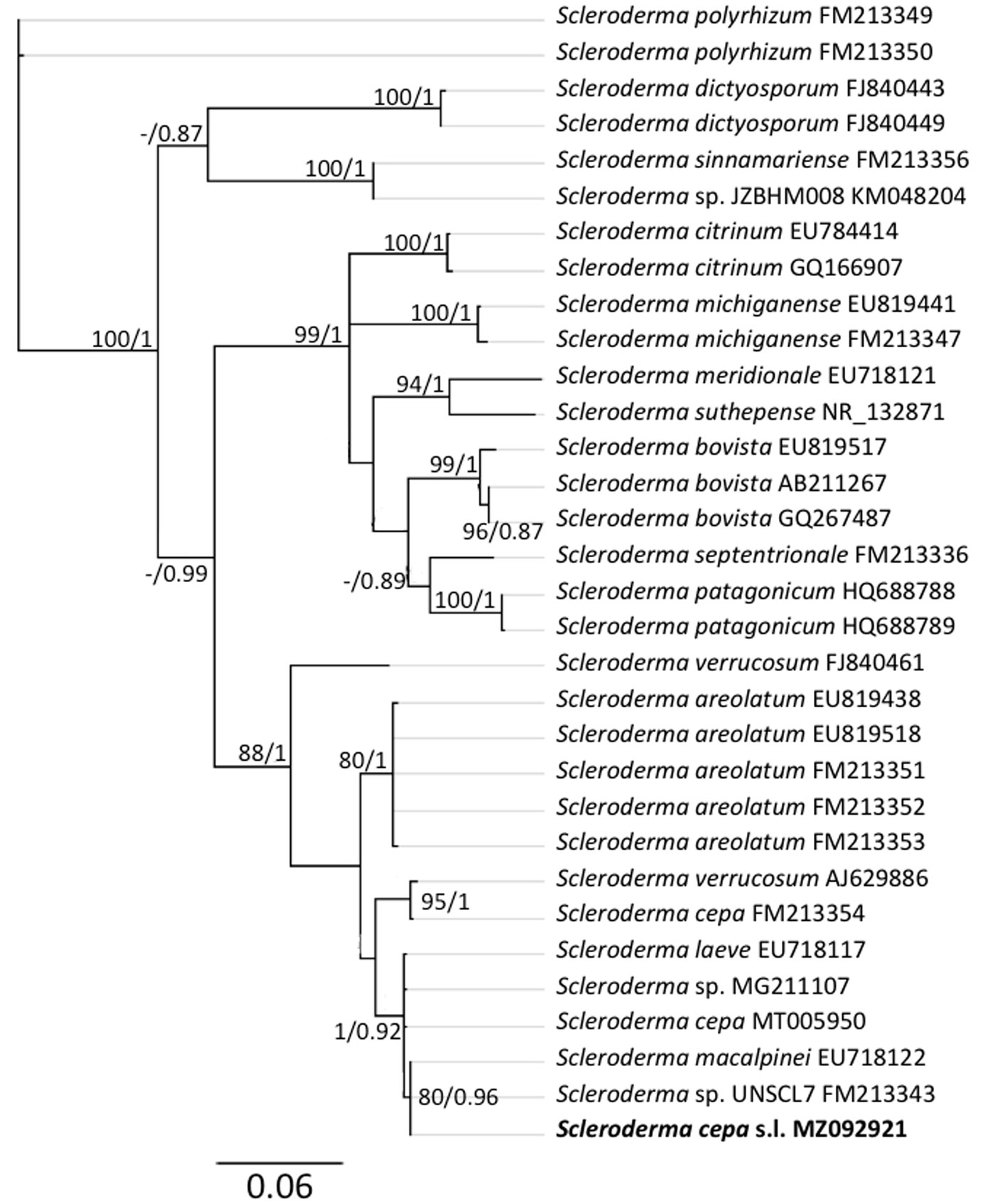
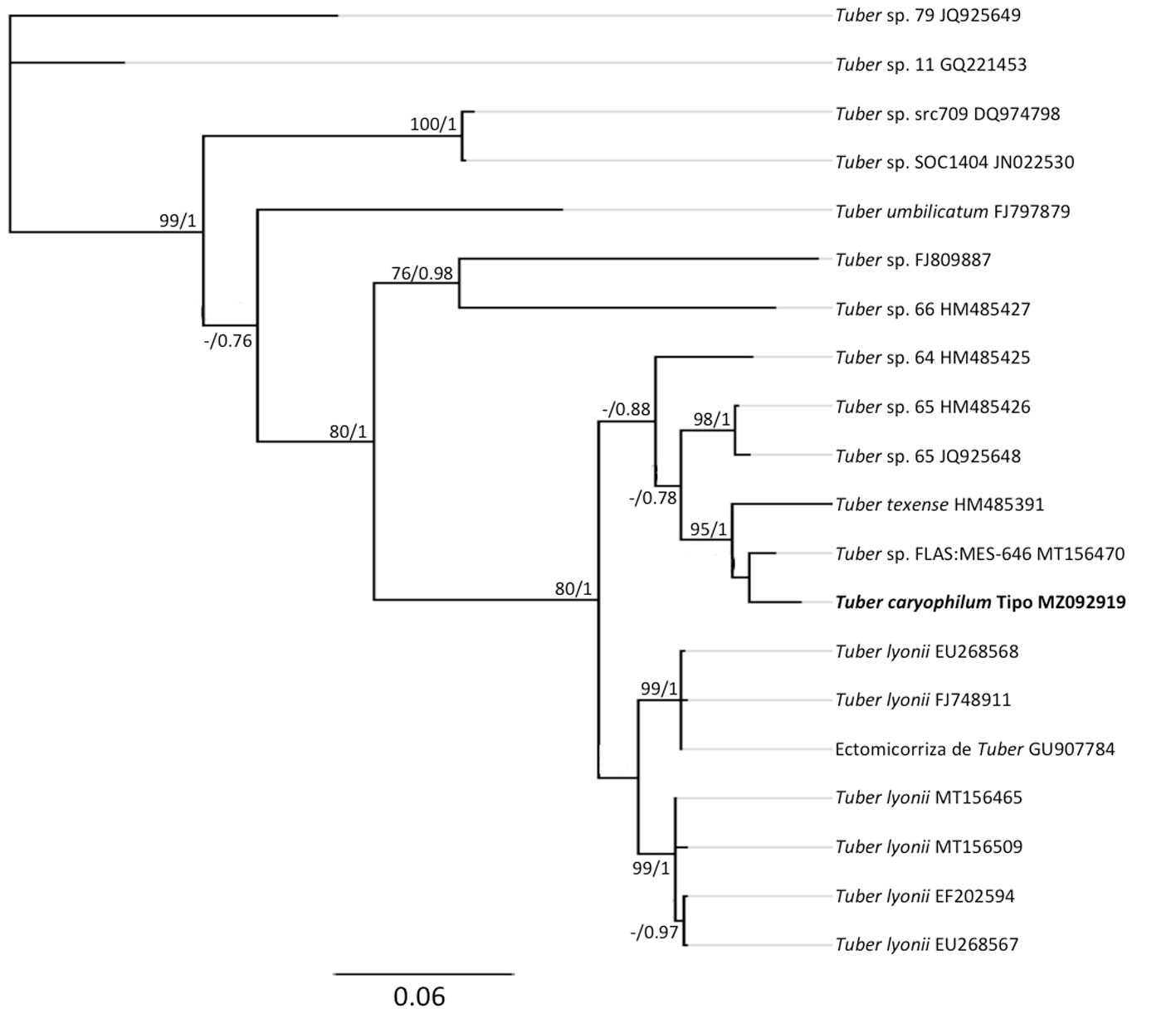
Los análisis filogenéticos demuestran que la muestra st4 se ubicó en Agaricus (fig. 3) dentro del clado de A. deserticola con un buen soporte (PPB = 0.97, MLB = 90). Esta especie es el grupo hermano de A. evertens (PPB = 1, MLB = 95) con un porcentaje de similitud nucleotídica (% SM) de 98.6%. Las muestras st2, st27 y st28 pertenecen a Candolleomyces (fig. 4). La muestra st2 se ubicó dentro del clado de C. luteopallidus con un buen soporte (PPB = 1, MLB = 77). Esta especie es el grupo hermano de C. halophilus y C. littoralis (PPB = 0.99, MLB = 73) (% SM = 96.3%). Por su parte, las muestras st27 y st28 forman un clado independiente, posiblemente relacionado con C. efflorescens (PPB = 0.8, MLB = 40) (% SM = 95.6%) y con un conjunto de muestras identificadas como Candolleomyces sp. (PPB = 0.86, MLB = 42). La muestra st26 se ubicó en Scleroderma (fig. 5) en un clado con numerosas secuencias identificadas como Scleroderma sp. (FM213343, MG212207), S. malcapinei (EU218122), S. laeve (EU718117) o S. cepa (MT005950) con un buen soporte de (PPB = 1, MLB = 92). Estas secuencias, junto con FM213354 y FM213343, pertenecen al complejo de S. cepa s.l. y son el grupo hermano de S. areolatum (PPB = 1, MLB = 78) (% SM = 95.4%). La muestra st3 se ubicó en Tuber (fig. 6) junto con la secuencia del ejemplar FLAS: MES-646 identificada como Tuber sp. en un clado con poco soporte (PPB = 0.69, MLB = 51). Este clado es el grupo hermano de Tuber texense (PPB = 1, MLB = 95) (% SM = 94.4%).
Descripción
Agaricus deserticola
Carpóforo secotioide, píleo subgloboso a ovoide, blanco a pardo a veces escamoso, margen que se fusiona con el estípite durante el desarrollo. No forma láminas, las esporas se encuentran dentro de la gleba. Píleo de 2-6 cm de diámetro, estípite de 5-20 cm de alto y de 10-13 mm de ancho. Contexto blanco que con la desecación y edad adquiere tonos amarillos. Esporas violetas ovales de 7-9 × 4-6 µm. Se encontró en junio-julio, en zonas de la huerta donde hay escasez de agua y riego por goteo subsuperficial, abundante luz y árboles de poco follaje.
Material estudiado: Sánchez-Ledesma st4, huerto de C. illinoinensis Tierra Blanca, Viesca, Coahuila, 4 de julio 2018, MEXU 30228.
Comentarios taxonómicos
Morfológicamente, el ejemplar st4 de A. deserticola corresponde completamente con la descripción de la especie (sensu Moreno et al., 2010) y presentó una similitud nucleotídica entre 99.4 y 100%, con secuencias de ejemplares de esta especie provenientes del suroeste de EUA. Primera vez que se reporta en plantaciones de C. illinoinensis y en Coahuila.
Candolleomyces luteopallidus
Carpóforo con píleo acampanado de 10-20 mm de ancho, marrón que se decolora a blanco, cutícula lisa, estriado. Los cuerpos fructíferos jóvenes tienen colores más intensos que los viejos. Láminas agrietadas, al principio blancas luego púrpuras. Carne frágil, inodora, muy escasa, estípite delicado, de 1-3 cm, aterciopelado, marrón. Esporas (3-6) × (4-5) µm, lisas, rojizas en agua y marrón oscuro en KOH al 5%, elipsoides, cilíndricas ocasionalmente subovoides, con poro germinativo central pequeño. Se encontró abundantemente a inicios de otoño, crecen adheridos a la corteza de los árboles, se considera un hongo saprofito que se alimenta de la corteza.
Material estudiado: Sánchez-Ledesma st2, huerto de C. illinoinensis Tierra Blanca, Viesca, Coahuila, 21 de mayo 2018, MEXU 30229.
Comentarios taxonómicos
Morfológicamente, el ejemplar st2 de C. luteopallidus corresponde bien con la descripción de la especie (sensu Smith, 1972) y presenta una similitud nucleotídica de 99.5% con el holotipo de esta especie (tabla 2). Primer registro de la especie para México y primer reporte en huertas de nogal pecanero.
Tabla 2
Similitud nucleotídica de los hongos asociados a Carya illinoinensis con los registros de la base de datos del GenBank. En todos los casos la probabilidad de error “e-value” fue igual a cero.
| Número de acceso en GenBank | Taxón | Secuencia más cercana en GenBank | Porcentaje de similitud nucleotídica |
| MZ092918 | Candolleomyces luteopallidus st2 | Candolleomyces luteopallidus (holotipo) KC992884 | 99.5 |
| MZ092919 | Tuber caryophilum st31 | Tuber sp. MT156470 | 97 |
| MZ092920 | Agaricus deserticola st4 | Agaricus deserticola HM488747 | 100 |
| MZ092921 | Scleroderma cepa s.l. st26 | Scleroderma FM213343 | 100 |
| MZ092922 | Candolleomyces sp. st27 | Candolleomyces sp. KY563654 | 99.1 |
| MZ092923 | Candolleomyces sp. st28 | Candolleomyces sp. KY563655 | 99.1 |
Candolleomyces sp.
Carpóforo con píleo más o menos acampanado, que oscila entre 4-8 cm de ancho, después se vuelve convexo con el borde enrolado, cutícula lisa, estriado en la parte superior con el centro marrón, higrófano que cambia de gris a gris salmón. Láminas marrón a crema, en ocasiones blancas. Estípite delicado, aterciopelado blanco a crema de 1-8 cm. Contexto frágil, inodoro y muy escaso. Esporas alargadas negruzcas de 6-8 × 4-5 µm, lisas, de color marrón en agua y marrón oscuro en KOH, con poro germinativo central. Saprófito encontrado de mayo a agosto entre la maleza de la huerta.
Material estudiado: Sánchez-Ledesma st27, huerto de C. illinoinensis Tierra blanca, Viesca, Coahuila, 17 de mayo 2018, MEXU 30230; Sánchez-Ledesma st28, 8 de junio 2018, MEXU 30231.
Comentarios taxonómicos
Morfológicamente, Candolleomyces sp. es muy similar a C. candolleanus el cual es un complejo de especies (antes conocido como Psathyrella candolleana) con una gran variación morfológica y amplia distribución geográfica (Al-Habib et al., 2014; Wächter y Melzer, 2020). Los ejemplares identificados como Candolleomyces sp. (st27, st28) presentan una similitud nucleotídica de 99.1% con múltiples muestras identificadas como Psathyrella sp. de la India. Todos estos ejemplares se ubican filogenéticamente en el complejo de C. candolleanus (fig. 4), por lo que podría tratarse de una especie no descrita, aunque hace faltan secuencias de varios holotipos del género.
Scleroderma cepa s.l.
Carpóforos globosos con grietas irregulares, un poco escachados con un exoperidio marcado con escamas poligonales. Cuando inmaduro la gleba es blanca con tonos verdes con una pequeña retícula blanca, después se vuelve gris. Cuando maduro la gleba se vuelve negra violácea y el peridio se rompe por la zona apical y expulsa las esporas pulverulentas. Gleba con olor desagradable. Pseudoestípite radicante con abundante micelio blanquecino-amarillento. En el himenio hay basidios esféricos, hifas emergentes, esporas esféricas con espinas aisladas, sin crestas de conexión de 8-9 × 12-14 μm. Fructifica desde julio a noviembre, siendo más abundante en septiembre y octubre.
Material estudiado: Sánchez-Ledesma st26, huerto de C. illinoinensis Tierra Blanca, Viesca, Coahuila, 3 de septiembre 2018, MEXU 30232.
Comentarios taxonómicos
La secuencia del ejemplar st26 es muy similar (99.7-100%) a decenas de secuencias de ejemplares identificados mayoritariamente como S. cepa, S. laeve o S. malcapinei. Debido a que las especies de este clado son muy plásticas morfológicamente y no existen secuencias de los ejemplares tipo, conservadoramente la identificamos como S. cepa s.l.
Tuber caryophilum
Ascoma hipogeo, globoso a subgloboso, irregular, marrón, sin cambio de color al tacto, venas blancas finas, cerradas y numerosas, con 4-6 verrugas en 1 mm, sólido, superficie seca separable con dermatocistidios y surcos blancos. Peridio de l20 ± 13 µm de grosor, epicutis pseudoparenquimatoso 92 ± 10 µm de ancho, células isodiamétricas 0.5-1 µm de ancho. Subcutis 27 ± 4 µm de grosor delimitado por el epicutis, entretejido por hifas postradas o entrelazadas de 4-5 µm. Esporas de 8-3 × 4-6 µm, hialinas marrón en KOH, paredes 2 µm de grosor. Se encontró en el mes de septiembre, solitario a gregario a 3 cm de profundidad.
Material estudiado: Sánchez-Ledesma st3, huerto de C. illinoinensis Tierra Blanca, Viesca, Coahuila, 3 de agosto 2018, MEXU 30227.
Comentarios taxonómicos
Tuber caryophilum presentó un valor de 97% de similitud nucleotídica con el ejemplar FLAS: MES-646 identificado como Tuber sp. procedente de Texas, EUA. Los ejemplares recolectados en este estudio se usaron recientemente para la descripción de esta especie (Sánchez-Ledesma et al., 2022). Se trata de una especie micorrízica de C. illinoinensis distribuida en el norte de México y sur de EUA.
Discusión
Agaricus cuenta con aproximadamente 500 especies en el mundo (Kirk et al., 2008; Palestina-Villa et al., 2020). En México se han reportado 53 nombres válidos distribuidos en 28 estados (Palestina-Villa et al., 2020). Para A. deserticola se tienen registros de Baja California, Chihuahua y Sonora. Se ha encontrado, principalmente, en hábitats secos o semiáridos, tales como desierto, pradera, matorrales de salvia costera, campos, céspedes y zonas urbanas, así como en bosques de Quercus y bosques de espinas (Harding, 1957; Moreno et al., 2010; Palestina-Villa et al., 2020; Quiñones et al., 1999).
Candolleomyces es un género agarical recientemente segregado de Psathyrella, el cual se subdivide en 13 subclados monofiléticos con numerosas especies no descritas y complejos de especies con alta variación morfológica (Wächter y Melzer, 2020). Candolleomyces luteopallidus es una especie rara, con pocos registros a nivel mundial, aunque se ha reportado en ecosistemas de árboles de interés forestal como una plantación joven de olivos en Europa (Muñoz y Caballero, 2012). Secuencias de ADN de muestras ambientales de esta especie se han encontrado en suelo de matorral xerófilo y raíces de pastos de zonas semidesérticas en el norte de México y sur de EUA. Candolleomyces sp. pertenece al complejo de C. candolleanus, una especie común en América del Norte y América del Sur, que en México se encuentra registrada en el trópico, en bosques mesófilos y bosques de pino en el Cofre de Perote, Veracruz (Guzmán et al., 1988; Smith, 1972).
Las especies de Scleroderma son hongos gasteroides en la familia Sclerodermataceae, tienen una distribución cosmopolita y son simbiontes ectomicorrízicos de distintos hospederos como Castanea, Quercus, Fagus, Nothofagus, Pinus, Prunus y orquídeas, desde zonas tropicales hasta áridas (Cruz et al., 2017; González-Chávez et al., 2018; Phosri et al., 2009). Se ha registrado en Europa y Norteamérica, Chile, Argentina, Brasil, Tailandia, etc. (Cruz et al., 2017; Gurgel et al., 2008; Kumla et al., 2013; Nouhra et al., 2012). En México, se tienen registros de 21 especies de Scleroderma, siendo la más común S. nitidum (Guzmán et al., 2013). Específicamente, se han reportado en la Costa del Pacífico, Puebla, Veracruz y en Chihuahua (González -Chávez et al., 2018; Guzmán et al., 2004; Rivero et al., 1999; Tarango, 2004). Además, en este último estado se tienen registros de ectomicorrizas en huertas de nogal pecanero (Rivero et al., 1999; Tarango, 2004). Los datos corroboran que S. cepa s.l. es un simbionte ectomicorrízico del nogal pecanero en La Comarca Lagunera de manera abundante y natural. Existen registros de S. cepa asociada con Quercus pubescens y Fagus sylvatica (Mrak et al., 2017). Por la amplia y abundante distribución de Scleroderma, se propone muestrearla más ampliamente en las huertas de nogal pecanero en el sur de EUA y el norte de México, con el fin de aumentar el conocimiento sobre la micorrización de C. illinoinensis con Scleroderma spp.
Tuber forma ectomicorrizas con árboles de interés forestal como castaños, robles y nogales (Benucci et al., 2012a; Bonito et al., 2011). El género tiene una distribución geográfica amplia, se han reportado más de 30 especies en Europa, particularmente Portugal, España, Francia e Italia (Benucci et al., 2012b). Para México se tienen registradas 13 especies, distribuidas en el norte, centro y sur, siendo los estados más ricos Nuevo León y Tamaulipas (Bonito et al., 2013; Guevara et al., 2013). Tuber caryophilum pertenece al complejo de T. lyonii (sensu Healy et al., 2016) junto con T. texense. Esto es interesante, ya que estas 3 especies se desarrollan en huertas de nogal pecanero; T. texense y T. lyonii reportadas al sur de EUA y Tuber caryophilum reportada anteriormente (como Tuber sp.) en huertas de C. illinoinensis en México, en particular, en el estado de Chihuahua (González-Chávez et al., 2009; Tarango et al., 2004).
En general, las plantaciones de C. illinoinensis tienen poca diversidad de macromicetos, aunque es importante señalar que durante las recolectas observamos que las especies identificadas fructifican abundantemente en la huerta cubriendo amplias superficies. Si bien existe cierto conocimiento sobre los hongos ectomicorrízicos asociados al nogal, esta es la primera vez que se registran hongos saprobios como C. luteopallidus, Candolleomyces sp. y A. deserticola. Este trabajo contribuye al conocimiento de los macromicetos en plantaciones de C. illinoinensis con nuevos registros para La Comarca Lagunera de Coahuila y México.
Por lo anterior concluimos que, dada a la escasez de trabajos de identificación taxonómica y molecular de hongos asociados al nogal pecanero, la diversidad de macromicetos encontrada es un avance importante para entender su diversidad y función. Asimismo, es importante incrementar los inventarios de hongos en agroecosistemas donde presentan una relevante riqueza micológica.
Agradecimientos
JASL agradece al Conacyt por la beca de posgrado. Parte de los recursos para biología molecular fueron aportados por el proyecto PAPIIT-UNAM (IN212521) otorgado a RGO.
Referencias
Al-Habib, M. N., Holliday, J. y Tura, D. (2014). The pale brittle stem mushroom, Psathyrella candolleana (higher Basidiomycetes): an indigenous medicinal mushroom new to Iraq. International Journal of Medicinal Mushrooms, 16, 617–622. https://doi.org/10.1615/IntJMedMushrooms.v16.i6.110
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. y Lipman, D. J. (1990). Basic local alignment search tool. Journal of Molecular Biology, 215, 403–410.
Benucci, G. M. N., Bonito, G., Falini, L. B. y Bencivenga, M. (2012a). Mycorrhization of Pecan trees (Carya illinoinensis) with commercial truffle species: Tuber aestivum Vittad. and Tuber borchii Vittad. Mycorrhiza, 22, 383–392. https://doi.org/10.1007/s00572-011-0413-z
Benucci, G. M. N., Csorbai, A. G., Falini, L. B., Bencivenga, M., Di Massimo, G. y Donnini, D. (2012b). Mycorrhization of Quercus robur L., Quercus cerris L. and Corylus avellana L. seedlings with Tuber macrosporum Vittad. Mycorrhiza, 22, 639–646. https://doi.org/10.1007/s00572-012-0441-3
Bickford, D., Lohman, D. J., Sodhi, N. S., Meier, R., Winker, K. y Das, I. (2007). Cryptic species as a window on diversity and conservation. Trends in Ecology & Evolution, 22, 148–155. https://doi.org/10.1016/j.tree.2006.11.004
Bonito, G., Brenneman, T. y Vilgalys, R. (2011). Ectomycorrhizal fungal diversity in orchards of cultivated pecan (Carya illinoinensis; Juglandaceae). Mycorrhiza, 21, 601–612. https://doi.org/10.1007/s00572-011-0368-0
Bonito, G., Smith, M. E., Nowak, M., Healy, R. A., Guevara, G., Cázares, E. et al. (2013). Historical biogeography and diversification of truffles in the Tuberaceae and their newly identified southern hemisphere sister lineage. Plos One, 8, e52765. https://doi.org/10.1371/journal.pone.0052765
Casales, F. G., Van der Watt, E. y Coetzer, G. M. (2018). Propagation of pecan (Carya illinoinensis): a review. African Journal of Biotechnology, 17, 586–605. https://doi.org/10.1371/journal.pone.0052765
Conabio. (2014). Catálogos de autoridades taxonómicas (CAT) Biótica 5.0. Ciudad de México: Conabio. Recuperado el 8 noviembre, 2020 de: http://www.conabio.gob.mx/biotica5/documents/CursoEnero2014/SCATBiotica50_enero2014.pdf
Córdova-Chávez, O., Medel, R., Mata, G., Castillo, R. y Vázquez-Ramírez, J. (2014). Evaluación de hongos ectomicorrícicos del grupo de los basidiomicetos en la zona del Cofre de Perote, Veracruz. Madera y Bosques, 20, 97–106.
Cruz, R., Carvajal, L. y Pérez, S. (2017). Identificación de Scleroderma citrinum Pers en una plantación de almendros de la ciudad de Villa Alemana, Chile. Boletín Micológico, 32, 34–38. https://doi.org/10.22370/bolmicol.2017.32.2.1062
Edgar, R. C. (2004). MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics, 5, 1–19.
Ge, Z. W., Brenneman, T., Bonito, G. y Smith, M. E. (2017). Soil pH and mineral nutrients strongly influence truffles and other ectomycorrhizal fungi associated with commercial pecans (Carya illinoinensis). Plant and Soil, 418, 493–505. https://doi.org/10.1007/s11104-017-3312-z
González-Chávez, B. P., Ojeda-Barrios, D. L., Hernández-Rodríguez, O. A., Martínez-Téllez, J. y Núñez-Barrios, A. (2009). Ectomicorrizas en nogal pecanero. Tecnociencia Chihuahua, 3, 138–146.
González-Chávez, M. D., Torres-Cruz, T. J., Sánchez, S. A., Carrillo-González, R., Carrillo-López, L. M. y Porras-Alfaro, A. (2018). Microscopic characterization of orchid mycorrhizal fungi: Scleroderma as a putative novel orchid mycorrhizal fungus of Vanilla in different crop systems. Mycorrhiza, 28, 147–157. https://doi.org/10.1007/s00572-017-0808-6
Grupe, A. C., Sulzbacher, M. A., Grebenc, T., Healy, R., Bonito, G. y Smith, M. E. (2018). Tuber brennemanii and Tuber floridanum: Two new Tuber species are among the most commonly detected ectomycorrhizal taxa within commercial pecan (Carya illinoinensis) orchards. Mycologia, 110, 780–790. https://doi.org/10.1080/00275514.2018.1490121
Guevara, G., Bonito, G. y Cázares, E. (2013). Revisión del género Tuber (Tuberaceae: Pezizales) de México. Revista Mexicana de Biodiversidad, 84, S39–S49. https://doi.org/10.7550/rmb.31981.
Gurgel, F. E., Silva, B. y Baseia, I. G. (2008). New records of Scleroderma from northeastern Brazil. Mycotaxon, 105, 399–405.
Guzmán, G., Cortés-Pérez, A., Guzmán-Dávalos, L., Ramírez-Guillén, F. y Sánchez-Jácome, M. (2013). An emendation of Scleroderma, new records, and review of the known species in Mexico. Revista Mexicana de Biodiversidad, 84, S173–S191. https://doi.org/10.7550/rmb.31979
Guzmán, G., Montoya-Bello, L. y Bandala-Muñoz, V. M. (1988). A new species of Psathyrella (Agaricales, Coprinaceae) from Mexico with discussions on the known species. Brittonia, 40, 229–234.
Guzmán, G., Ramírez-Guillén, F., Miller, O. K. Jr., Lodge, D. J. y Baroni, T. J. (2004). Scleroderma stellatum versus Scleroderma bermudense: the status of Scleroderma echinatum and the first record of Veligaster nitidum from the Virgin Islands. Mycologia, 96, 1370–1379. https://doi.org/10.1080/15572536.2005.11832886
Harding, P. R. Jr. (1957). Notes on Longula texensis var. major. Mycologia, 49, 273–276. https://doi.org/10.1080/00275514.1957.12024640
Healy, R., Bonito, G. M. y Smith, M. E. (2016). A brief overview of the systematics, taxonomy, and ecology of the Tuber rufum clade. En A. Zambonelli, M. Iotti y C. Murat (Eds.), True truffle (Tuber spp.) in the World (pp. 125–136). Suecia: Springer. https://doi.org/10.1007/978-3-319-31436-5_8
Huelsenbeck, J. P. y Ronquist F. (2001) MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics, 17, 754–755.
Index Fungorum (2021). Index Fungorum. Recuperado el 07 octubre, 2020 de: http://www.indexfungorum.org./Names/Names.asp
INIFAP (Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias). (2002). Tecnología de producción en nogal pecanero. Ciudad de México: CELALA-CIRNOC-INIFAP.
Kerrigan, R. W. (2016). Agaricus of North America. Memoirs of the New York Botanical Garden, Volume 114. Nueva York: The New York Botanical Garden Press.
Kirk, P. M., Cannon, P. F., Minter, D. W. y Stalpers, J. A. (2008). Dictionary of the Fungi, 10th edition. Wallingford: CABI.
Kumla, J., Suwannarach, N., Bussaban, B. y Lumyong, S. (2013). Scleroderma suthepense, a new ectomycorrhizal fungus from Thailand. Mycotaxon, 123, 1–7. http://doi.org/10.5248/123.1
Marx, D. H. (1971). Root inhabiting mycorrhizal fungi benefit growth of trees. En Proceedings of the 5th Ann. West. Irri. Pecan Grow. Ass. Conf. (pp. 14–18). New Mexico: New Mexico State University.
Moreno, G., Lizárraga, M., Esqueda, M. y Coronado, M. L. (2010). Contribution to the study of gasteroid and secotioid fungi of Chihuahua, Mexico. Mycotaxon, 112, 291–315.
Mrak, T., Kühdorf, K., Grebenc, T., Štraus, I., Münzenberger, B. y Kraigher, H. (2017). Scleroderma areolatum ectomycorrhiza on Fagus sylvatica L. Mycorrhiza, 27, 283–293. https://doi.org/10.1007/s00572-016-0748-6
Munsell, A. H. (1954) Munsell soil color charts. Baltimore: Munsell Color Company.
Muñoz, G. y Caballero, A. (2012). Contribución al conocimiento del género Psathyrella en la Península Ibérica (I). Boletin Micológico FAMCAL, 7, 37–74.
Nouhra, E. R., Hernández Caffot, M. L., Pastor, N. y Crespo, E. M. (2012). The species of Scleroderma from Argentina, including a new species from the Nothofagus forest. Mycologia, 104, 488–495. https://doi.org/10.3852/11-082
Palestina-Villa, E. N., Parra-Sánchez, L. A., Villegas, M., Garibay-Orijel, R. y Medel-Ortiz, R. (2020). The known species of Agaricus (Agaricales, Agaricaceae) in Mexico, an updated and nomenclatural review. Scientia Fungorum, 50, e1269. https://doi.org/10.33885/sf.2020.50.1269
Paz, A., González, M. y Crawford, A. J. (2011). Códigos de barras de la vida: introducción y perspectiva. Acta Biológica Colombiana, 16, 161–175.
Phosri, C., Martín, M. P., Watling, R., Jeppson, M. y Sihanonth, P. (2009). Molecular phylogeny and re-assessment of some Scleroderma sp. (Gasteromycetes). Anales del Jardín Botánico de Madrid, 66, 83–91. http://dx.doi.org/10.3989/ajbm.2199
Posada. D. (2008). jModelTest: Phylogenetic model averaging. Molecular Biology and Evolution, 25, 1253–1256. https://doi.org/10.1093/molbev/msn083
Quiñónez, M., Garza, F., Mendoza, J. R., García, J., Sáenz, J. y Bolaños, H. (1999). Guía de hongos de la región de bosque Modelo Chihuahua. Chihuahua: Universidad Autónoma de Chihuahua/ Universidad Autónoma de Nuevo León/ Instituto Tecnológico de Ciudad Victoria/ Bosque Modelo Chihuahua, A.C.
Ramírez, L. R. (2013). Los basidiomicetos: una herramienta biotecnológica promisoria con impacto en la agricultura. Fitosanidad, 17, 49–55.
Rivero, S. T., Moorillón, V. N. y Borunda, E. O. (2009). Growth, yield, and nutrient status of pecans fertilized with biosolids and inoculated with rizosphere fungi. Bioresource technology, 100, 1992–1998. https://doi.org/10.1016/j.biortech.2007.12.078
Sagaram, M., Lombardini, L. y Grauke, L. J. (2011). Variation in anatomical characteristics in leaves of pecan seedstocks from Mexico and the United States. Journal of the American Society for Horticultural Science, 136, 103–108. https://doi.org/10.21273/JASHS.1
Sánchez-Ledesma, J. A., Guevara-Guerrero, G., Garibay-Orijel, R., Ángeles-Argáiz, R., Ávila-Rodríguez, V., Arreola-Ávila, J. G. et al. (2022). Tuber caryophilum, a new truffle species growing in Carya illinoinensis orchards. Revista Mexicana de Biodiversidad, 93, e934893. https://doi.org/10.22201/ib.20078706e.2022.93.4893
Sambrook, J., Fritsch, E. y Maniatis, T. (1989). Molecular cloning: a laboratory guide. Nueva York: Cold Spring Harbor Laboratories Press.
SIAP (Servicio de Información Agroalimentaria y Pesquera). (2021). Avances de siembras y cosechas 2021. Recuperado el 26 abril, 2022 de: http://infosiap.siap.gob.mx/gobmx/datosAbiertos.php
Smith, A. H. (1972). The North American species of Psathyrella. Memoirs of the New York Botanical Garden, 24, 1–633.
Stamatakis A. (2006). RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics, 22, 2688–2690. https://doi.org/10.1093/bioinformatics/btl446
Taber, R. A., 1984. Mycorrhyzal fungi associated with pecans. En 18th Western Pecan Conference Proceedings (pp. 135–136). New Mexico: CES-New Mexico State University.
Tarango, R. S. 2004. Micorrizas en nogal pecanero y pistachero. Folleto Técnico No. 16 del INIFAP-Delicias. Chihuahua: Centro de investigaciones regionales norte-centro campo experimental Delicias.
Tovar, A. R., Cásarez, B. X. y Valdés, M. (2004). Ecología molecular de los hongos ectomicorrízicos. Revista Fitotecnia Mexicana, 27, 267–278.
Villasante, J., Pérez-Carrillo, E., Heredia-Olea, E., Metón, I. y Almajano, M. P. (2019). In vitro antioxidant activity optimization of nut shell (Carya illinoinensis) by extrusion using response surface methods. Biomolecules, 9, 883. https://doi.org/10.3390/biom9120883
Vrain, T. C., Wakarchuk, D. A., Levesque, A. C. y Hamilton, R. I. (1992). Intraspecific rDNA restriction fragment length polymorphism in the Xiphinema americanum group. Fundamental and applied Nematology, 15, 563–573.
Wächter, D. y Melzer, A. (2020). Proposal for a subdivision of the family Psathyrellaceae based on a taxon-rich phylogenetic analysis with iterative multigene guide tree. Mycological Progress, 19, 1151–1265. https://doi.org/10.1007/s11557-
020-01606-3
Yun, W. y Hall, I. R. (2004). Edible ectomycorrhizal mushrooms: challenges and achievements. Canadian Journal of Botany, 82, 1063–1073.


