
Beder D. Madrid-Peralta * y Neis J. Martínez-Hernández
Universidad del Atlántico, Semillero de investigación de Artrópodos NEOPTERA, Atlántico, Km 7 antigua vía Puerto Colombia, Atlántico, Colombia
*Autor para correspondencia: bederdmp@gmail.com (B.D. Madrid-Peralta)
Recibido: 14 octubre 2022; aceptado: 15 mayo 2023
Resumen
Los estudios poblacionales de mariposas en el Caribe colombiano son escasos, razón por la cual se analizó la estructura poblacional y el desplazamiento de Hamadryas Hübner (1806) en diferentes usos del suelo en un paisaje fragmentado en los Montes de María, San Jacinto, Bolívar. Se realizaron 6 muestreos entre marzo y junio de 2018. En el área de estudio se ubicaron 7 estaciones donde se colocaron 5 trampas Van Someren-Rydon cebadas con mezcla de fruta fermentada. Mediante el método de captura-marcaje-recaptura, se registraron 541 individuos del género Hamadryas (312 machos – 229 hembras). La especie más abundante fue H. feronia (Linnaeus, 1758) con 291 individuos registrados y la menos frecuente H. amphinome (Linnaeus, 1767) con 33 registros. Las proporciones sexuales variaron, aunque el número de machos siempre fue mayor y las hembras experimentaron una mayor distancia de desplazamiento. La estructura de edades y las recapturas mostraron un pico de emergencia de imagos entre abril-junio. Por último, la variación de la abundancia de Hamadryas se asocia, principalmente, con la cobertura del dosel, indicando que este factor juega un papel determinante en la dinámica poblacional de este grupo de mariposas.
Palabras clave: Abundancia; Paisaje; Estructura
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Population aspects and movement of thunderring butterflies Hamadryas (Nymphalidae) in different land uses in Montes de María, Colombia
Abstract
Population studies about butterflies in the Colombian Caribbean are scarce, which is why we analyzed the variation in the population structure and displacement of Hamadryas in different land uses in a fragmented landscape in Montes de María, San Jacinto, Bolívar. Six samplings were conducted between March and June, 2018. Seven sampling stations were located in the study area where 5 Van Someren-Rydon traps baited with fermented fruit mix were placed in each station. Using the capture-mark-recapture method, 541 individuals of the genus Hamadryas were recorded (including 312 males and 229 females). The most abundant species was H. feronia with 291 individuals recorded and the least frequent was H. amphinome with 33 individuals records. Sex ratios varied, although the number of males was always greater and the females experienced greater travel distance. The age structure and recapture showed a peak of imagos emergence for these butterflies between April-June. Finally, the variation in the abundance of Hamadryas is mainly associated with variables such as canopy cover, indicating that this factor plays a determining role in the population dynamics of this group of butterflies.
Keywords: Abundance; Landscape; Structure
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
En los Montes de María (Caribe colombiano), la destrucción del bosque seco tropical (BST) avanza de forma acelerada, con la expansión de las actividades antrópicas como la ganadería, siembra de cultivos, plantaciones forestales y extracción de madera (Castellanos-Barliza et al., 2018; González et al., 2014; Pizano y García, 2014; Rangel y Martínez, 2017). Lo anterior influye en la disponibilidad de hábitats para taxones de insectos como las mariposas (Mercado et al., 2018; Vargas-Zapata et al., 2012). Paralelo a la modificación y reducción de los hábitats, se producen cambios en los atributos de las especies como la reducción en sus poblaciones, cambios en su estructura y alteraciones en sus patrones de vuelo y desplazamiento (Bell et al., 1991; Márquez y Martínez, 2020; Townsend et al., 2001). Adicionalmente, la composición y usos del suelo en un paisaje juegan un papel importante en el grado de disponibilidad y distribución de los recursos nutricionales, sitios de apareamiento y la dispersión de las mariposas, ya que estos factores influyen en la estructura y desplazamiento de estos insectos (Andrade et al., 2007; Brown et al., 1991; Constantino et al., 2007; Rueda, 2016).
Por otro lado, los cambios en la estructura vegetal de los fragmentos y la pérdida de cobertura en el BST inciden en la dinámica espacio-temporal de la abundancia y en el desplazamiento de algunos grupos de mariposas (Vargas-Zapata et al., 2015). Con el fin de aportar a esta discusión, es necesario utilizar taxones focales dentro de estos insectos como las mariposas del género Hamadryas Hübner (1806), debido a la relación estrecha que mantienen con los cambios de diversidad y abundancias de las especies vegetales y variables ambientales (Brown y Freitas, 2002; Kremen et al., 1993). Así mismo, dichos cambios pueden afectar a las poblaciones adultas dentro del género, ya que éstos se alimentan de la savia de los árboles y de las frutas fermentadas; además, suelen utilizar estos sustratos para posarse, exhibirse, realizar búsqueda de pareja y defender su territorio y sitios de alimentación (Marini-Filho, 1996). De este modo, la pérdida de hábitat es una amenaza para las Hamadryas en regiones como los Montes de María (MM) en el departamento de Bolívar, donde la reducción de los relictos de BST es constante y avanza de forma acelerada (Ahumada et al., 2019; Pizano y García, 2014).
El género Hamadryas comprende un grupo de 20 especies de mariposas neotropicales (Lamas et al., 2004; Reverdin, 1914), su tamaño varía entre los 40 mm en H. cloe hasta los 80 mm de envergadura en H. guatemalena (Monge-Nájera, 1992; Monge-Nájera y Hernández, 1991). Para el Caribe colombiano se han reportado 4 especies de Hamadryas (Andrade et al., 2007), la mayoría de estos registros proviene de inventarios sobre la composición y estructura de lepidópteros en distintos fragmentos de BST en el departamento del Atlántico (Boom-Urreta et al., 2013; Montero et al., 2009; Prince-Chacón et al., 2011; Vargas-Zapata et al., 2015), Córdoba (Campos-Salazar et al., 2011), Magdalena (Vargas-Zapata et al., 2011) y Sucre (Mercado et al., 2018). Sin embargo, el seguimiento de este grupo de insectos desde un enfoque poblacional y su desplazamiento entre áreas con vegetación boscosa y las matrices adyacentes (áreas de pastizales, cultivos anuales y permanentes y bordes) se desconoce; razón por la que se espera que las especies presenten variaciones en su abundancia y desplazamiento en los distintos usos de suelo dentro del paisaje.
Con la finalidad de aportar información acerca de la dinámica poblacional de este grupo de mariposas, se analizaron la abundancia, tamaño poblacional, proporciones sexuales, estructura de edades y el desplazamiento de las especies del género Hamadryas (Biblidinae) en diferentes usos del suelo en un paisaje fragmentado en los Montes de María, San Jacinto, Bolívar y su relación con las variables ambientales y la cobertura vegetal.
Materiales y métodos
El paisaje de BST se encuentra en la vereda La Flecha (VLF), en el sistema montañoso de los Montes de María, municipio de San Jacinto, Bolívar (09°51’12.4” N, 75°10’41.4” O) a una altitud que oscila entre 324 y 500 m (fig. 1). El área presenta un clima tropical determinado por los vientos alisios del norte y noreste, los sistemas montañosos y el mar (Aguilera, 2013), con temperaturas oscilantes entre 27 y 32 °C. Su precipitación se clasifica como bimodal tetraestacional, con un periodo de extensa sequía y lluvias regulares entre abril, julio, septiembre y noviembre, con un nivel de precipitación entre 1,000 y 1,900 mm al año. Además, presenta una humedad relativa promedio de 75% (Hernández, 2008), presentando transición entre periodos secos y húmedos.

La vegetación predominante en los fragmentos de BST es la higrotropofítica (Holdridge, 1978), con paisaje montañoso (USAID, 2016). El arbolado varía entre los 22 y 25 m de altura, con estratos vegetales de tipo arbustivo, sotobosque y dosel. Entre las especies vegetales que predominan se pueden encontrar Bursera simaruba (L.) (Sarg.) (indio encuero), Anacardium excelsum (Bertero ex Kunth) Skeels (caracolí), Aspidosperma polyneuron (Müll. Arg.) (carreto), Bocageopsis sp. (yaya), Caesalpinia coriaria (Jacq.) Wild. (dividivi), Pseudobombax septenatum (Jacq.) Dugand. (majagua), Crescentia cujete L. (totumo), Gustavia superba (Kunth) O. Berg. (membrillo), Bulnesia arborea (Jacq.) Engl. (guayacán), Hura crepitans L. (ceiba lechosa), Spondias mombin L. (jobo), Guazuma ulmifolia Lam. (guácimo), Chrysophyllum cainito L. (caimito), Quadrella adoratissima (Jacq.) Hutch. (olivo), Sterculia apetala (Jacq.) H. Karst. (camajón), Gliricidia sepium (Jacq.) Walp. (matarraton), Ficus sp. (caucho) entre otras, y a su vez se encuentran palmas como Bactris guineensis (L.) H.E. Moore, Attalea butyracea (Mutis ex L.F.) Wess. Boer (palma de vino) y Sabal mauritiiformis (H. Karst) Griseb. et H. Wendl. (palma amarga) entremezcladas en la zona de bosque. Alrededor del fragmento de bosque, se encuentran pastizales (gramíneas introducidas), pastizales arbolados (gramíneas, arboles maderables y palmas amargas), cercas vivas y la presencia de policultivos como Mangifera indica L. (mango), Dioscorea esculenta (Lour) Burkill (ñame), Zea mays L. (maíz), Capsicum annuum L. (ají), Persea americana Mill. (aguacate), Manihot esculenta Crantz. (yuca), Musa x paradisiaca L. (plátano), Ananas comosus L. Merr. (piña), entre otros.
En el área de estudio se delimitó una ventana de 2 x 2 km para un total de 4 km2, donde se establecieron 7 estaciones (E1 – E7) de muestreo con un mínimo de 180 m de distancia y un máximo de 275 m con el fin de cubrir diversos usos de suelo. Entre los usos, se destacan los cultivos mixtos permanentes temporales (CMPT) o E1, compuesta por cultivos mixtos temporales (D. esculenta, Z. mays, C. annuum, M. esculenta, C. papaya) y permanentes de P. americana (aguacate), A. butyracea (palma de vino), M. indica (mango), C. cainito (caimito) y B. guineensis con algunos árboles típicos del bosque como A. excelsum (Bertero ex Kunth) Skeels (caracolí), H. crepitans (ceiba blanca), S. mombin (jobo) y A. polyneuron (carreto) con remanentes de vegetación adyacentes a quebradas temporales. Borde de bosque (BB) o E2, con presencia de cercas vivas constituida por especies predominantes como H. crepitans (ceiba lechosa), S. mombim (jobo), Quadrella adoratissima (olivo) y S. apetala (camajón) y de manera adyacente G. sepium (matarratón) y enredaderas. Las estaciones E3, E4 y E5 corresponden a zona de bosque (Bos); mientras que, E6 corresponde a una zona de vegetación secundaria (VS) con 12 – 15 años de abandono, con presencia de especies vegetales como H. crepitans (ceiba blanca), Bursera simaruba (indio encuero), Q. adoratissima (olivo); con alturas entre 3 y 15 m y DAP entre 3 (mín) y 15.2 cm2 (máx), así como enredaderas y arbustos como G. sepium (matarratón). La E7 corresponde a un área de cultivos mixtos permanentes (CMP), donde predominan árboles frutales como P. americana (aguacate), M. indica (mango) con alturas de 3 – 10 m, Musa sp. (guineo cuatro filos), Annona muricata L. (guanábana), S. purpurea L. (ciruela), palmas como A. butyracea (palma de vino) entremezcladas con zonas de cultivos de M. x paradisiaca (plátano) y A. comosus (piña).
Se realizaron 6 muestreos separados por un promedio de 15 días, entre marzo y junio de 2018, de los cuales 2 se desarrollaron durante época seca (M1- marzo 17 y M2 – abril 3) y los restantes durante las lluvias (M3 – abril hasta M6 – junio 6). La captura de las mariposas se realizó con la ayuda de trampas Van Someren-Rydon (VSR), las cuales fueron cebadas con una mezcla de frutas (M. x paradisiaca, C. papaya, A. comosus, M. indica) y panela fermentada durante 48 h (Devries y Walla, 2001; Ribeiro y Freitas, 2011). Las trampas fueron cebadas a las 7:00 am y revisadas en horas de la tarde del primer día y a las 24, 48 y 96 hrs, periodo en el cual se realizaba el reemplazo del atrayente, iniciando consecutivamente desde la E1 hasta la E7.
En cada una de las 7 estaciones se colocaron 5 trampas Van Someren-Rydon (VSR) (Devries, 1988) distribuidas en formas de “zig-zag” y separadas por un promedio de 61.8 ± 9 m entre sí (mínimo 50 m, máximo 70 m), a una altura entre 1 y 1.2 m del suelo (Ribeiro y Freitas, 2011; Ribeiro et al., 2008, 2009, 2010, 2012; Sambhu et al., 2018). Las trampas fueron georreferenciadas con un GPS (GARMIN – Monterra) con el fin de determinar la distancia de cada mariposa una vez que fuera recapturada, donde los datos de recapturas eran tomados en cuenta si los individuos eran determinados registrados nuevamente entre muestreos. La captura de los ejemplares se realizó de forma pasiva, utilizando la técnica de captura-marcaje-recaptura (CMR). Cada ejemplar capturado fue registrado con los datos de captura de cada punto con su respectivo sexo. Adicionalmente, se sacrificaron un ejemplar macho y uno hembra, representante de cada especie por la técnica de sujeción torácica propuesta por Neild (1996); para la corroboración taxonómica se siguieron las claves propuestas por Chacón y Montero (2007) y la página ilustrada Butterflies of America (Warren et al., 2013). Las mariposas capturadas fueron marcadas con un marcador permanente “Sharpie” en la celda discal del ala anterior con una combinación alfa-numérica, correspondiente al muestreo y un número consecutivo del marcaje de individuos (ej. Individuo 10A). Posteriormente estas fueron liberadas a su ambiente natural, con el objetivo de no contar nuevamente los individuos (Constantino, 2007; Ribeiro y Freitas, 2011; Ribeiro et al., 2008, 2010, 2012) y así registrar los individuos recapturados con los datos de cada punto y de esta manera determinar su desplazamiento; el cual fue determinado como la distancia recorrida por los individuos desde los puntos donde era capturados hasta los puntos donde eran recapturados (Prieto et al., 2005).
Adicional a lo anterior, en cada punto (hora de revisión por trampa en cada muestreo) se midieron la temperatura ambiente (°C) y la humedad relativa (%), con la ayuda de un termohigrómetro marca Extech-IR, la intensidad lumínica con un luxómetro marca Extech-DT-1308, y la cobertura del dosel con un densitómetro Marca Spherical Densiometer Model-A. Para esto último, se siguió la metodología propuesta por Mostacedo y Fredericksen (2000), quien describe que el espejo del equipo debe colocarse de manera horizontal y contar el número de cuadros donde, por lo menos, la mitad de su área se encuentre cubierto por ramas, hojas o troncos, y a su vez, estos se multiplicaron por un factor de corrección de 1.04 para obtener la cobertura del dosel. De esta manera, en cada punto, la medición se tomó haciendo 4 lecturas, orientando el densitómetro hacia los puntos norte, oeste, sur y este de 5 m de distancia de la respectiva trampa en cada sitio.
Con los registros de los individuos capturados (C), marcados (M) y recapturados (R) o CMR por trampa, estación y muestreo, se determinó el número total de individuos capturados o abundancia total (Ntotal) y el número neto de individuos o abundancia neta (Nnet = Ntotal – número de individuos recapturados). Posteriormente, con los datos de CMR se realizó una matriz de Trellis sin tener en cuenta el sexo respectivo para cada especie (Krebs, 1989; Lemos et al., 2005). De esta manera, esta matriz se empleó para calcular el tamaño poblacional de las especies H. feronia y H. februa en la VLF, mediante el estimador de Jolly-Seber (J.S.) (1964), descrito por Krebs (1989), Henderson (2003) y Lemos et al. (2005), empleando el programa de Ecological Methodology versión 5.1 (Krebs, 1998). Los cálculos fueron realizados para estas 2 especies, ya que fueron las que registraron el mayor porcentaje de recaptura. La proporción de sexos se calculó de acuerdo a la siguiente formula: número de machos/número de machos + número de hembras; el valor resultante de la proporción sexual es 0.5 si el número de machos y hembras es igual (Wilson y Hardy, 2002). Este método es el más recomendado para estimar las proporciones sexuales en la naturaleza, debido a que considera a los individuos como unidades discretas y por lo tanto refleja la abundancia relativa de cada sexo en una población (Ancona et al., 2017).
Los individuos adultos marcados se agruparon en 3 categorías de edad, las cuales se basaron en el estado de las alas (Freitas y Ramos, 2001). Las categorías involucradas son: nuevos (individuos recién emergidos), intermedios (adultos con alas en estado medio) y viejos (incluye individuos viejos o alas muy desgastadas). De esta manera, la estructura de edades se estimó como la proporción de individuos de cada categoría en relación con el total de individuos marcados. Mediante los datos de CMR de cada especie (Freitas y Ramos, 2001), se estimaron los desplazamientos totales para cada mariposa por especie y sexo (Freitas y Ramos, 2001). A su vez, se registró la media o el valor de desplazamiento medio, la mediana y la distancia mínima o valor de desplazamiento más bajo y el tiempo promedio de desplazamiento, teniendo en cuenta el tiempo de residencia o tiempo en el que los individuos de una especie permanecieron en los distintos usos de suelo dentro del paisaje (Silva y Hernández, 2015). De esta forma, se calculó la tasa media de desplazamiento (Tmm) como la media de desplazamiento en relación con el desplazamiento promedio de cada especie. Las distancias recorridas por individuo en línea recta entre áreas se calcularon utilizando el software ArGIS® Versión 10.5.
Por otro lado, se aplicaron los modelos lineales generalizados (MLG) para determinar el efecto de los usos y épocas (muestreos) como factores y la cobertura del dosel (CD) sobre la abundancia de las Hamadryas registradas. Los datos de abundancia (por especie) se modelaron siguiendo una distribución de Poisson (O’Hara y Kotze, 2010). En el modelo se incluyeron los efectos fijos, como el número de trampas, los factores como uso del suelo (estaciones) y épocas (muestreos: seca y lluvias), y la CD. La temperatura, la humedad ambiental y la intensidad lumínica se eliminaron del modelo debido a la alta correlación con la cobertura del dosel. El mejor modelo fue seleccionado siguiendo el criterio de información Akaike (AIC) más bajo, corregido para un tamaño de muestra pequeño (AICc; según complejidad, el número de parámetros y la aptitud de cada modelo) manteniéndose aquellos modelos con una disminución de 2 unidades AIC (20%) con respecto al modelo nulo. Este análisis se realizó utilizando la función glmer en el paquete lme4 (Bates et al., 2015) con el programa R Studio versión 4.1.2 de 64 bit (Balzarini et al., 2008).
Resultados
Se capturaron y liberaron 541 individuos del género Hamadryas, de los cuales 312 fueron machos y 229 hembras. La especie más abundante fue H. feronia con 291 individuos marcados y liberados (164 machos: 127 hembras). La segunda especie más abundante fue H. februa con 137 individuos (79 machos: 58 hembras), mientras que H. amphichloe presentó un total de 80 individuos (47 machos: 33 hembras) y H. amphinome con 33 individuos (22 machos: 11 hembras) fue la especie menos abundante dentro del paisaje (tabla 1). El mayor número de capturas se registró en marzo (M2) con 150 individuos (85 machos; 65 hembras). De manera general, el mayor número de machos registrados se presentó específicamente en la estación 2 (E2) durante el M2, al igual que el mayor número de hembras con 36 y 29 capturas, respectivamente (tabla 1). El menor número de capturas se presentó durante inicios de junio (M6) con 17 individuos, el cual coincidió con la etapa de post lluvia, respectivamente (tabla 1).
Se registraron un total de 71 recapturas (16.1%), de los cuales 41 fueron machos y 30 hembras. Del total de recapturas, 62 individuos (38 machos; 24 hembras) fueron recapturados una sola vez, 5 (3 machos; 2 hembras) se recapturaron 2 veces, mientras que 2 individuos (2 hembras) fueron recapturados 3 veces. La especie H. feronia presentó el mayor porcentaje de recapturas (20.96%) para un total de 11.68% de machos y 9.28% de hembras, presentándose el mayor valor (12 individuos) en la E7 (cultivos mixtos) para el M3 y M5 (8 machos – 4 hembras), respectivamente. Para H. februa, el registro de recapturas fue de 12.4% con un total de 7.3% machos y 5.1% para las hembras, y el mayor registro de individuos recapturados (4 individuos) se presentó en el borde (E2) durante el M2. De las 4 especies de Hamadryas capturadas en el área de estudio, H. amphichloe presentó el porcentaje más bajo de recapturas con 6.25% (3.75% machos; 2.5% hembras), y el valor máximo de recapturas (2 individuos) se presentó durante el M4 (1 macho – 1 hembra), en borde (E2) y cultivos mixtos (E7), mientras que para la especie H. amphinome todos los individuos recapturados fueron machos con 12.12% de la población y el menor valor de recapturas se presentó durante el M3 en las E5 (bosque) y E7.
El tamaño estimado de la población varió durante los eventos de captura y recaptura para cada una de las especies en los diferentes usos del suelo en la zona. A través del registro directo de individuos observados se determinó un menor tamaño poblacional para H. feronia en el M6 (inicio de junio), con 9 individuos y un mayor valor en el M2 (abril) con 93 individuos (tabla 1, fig. 5). Para esta especie, mediante el estimador de Jolly-Seber (JS), el valor máximo estimado fue de 329.9 individuos en el M3 (mediados de abril) y el menor en el M5 (inicios de mayo) con 36 individuos (ICI = 18.218 y ICS = 134.535). Las probabilidades de supervivencia más altas se observaron en M1 (marzo) con un valor de 0.681 (tabla 2). En el caso de la especie H. februa, el menor valor observado fue de 9 individuos en el M6 (junio) y el mayor valor en M1 (marzo) con 60 individuos. Con el estimador de Jolly-Seber (JS), el mayor tamaño poblacional para esta especie fue de 133 individuos en M2 (abril) y el menor en M5 (mayo) con 4 individuos (ICI = 2.479 e ICS = 47.902). Las probabilidades de supervivencia más alta se observaron en el M1 (marzo) con un valor de 0.77 y el más bajo se presentó en el muestreo 2 con 0.26 (tabla 2).
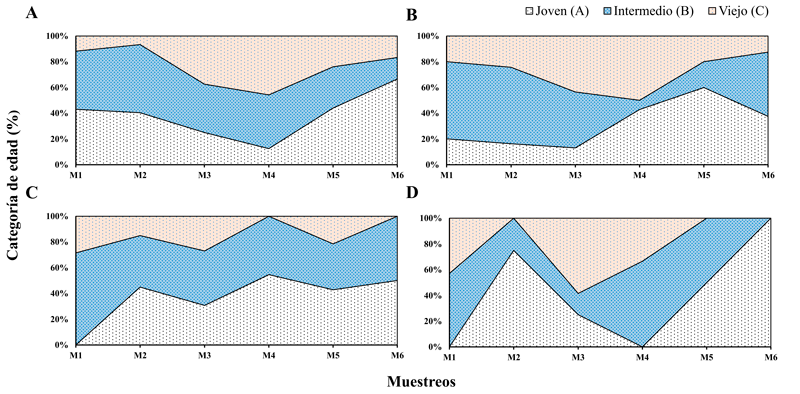
La estructura de edades varió para cada una de las 4 especies durante los 6 eventos de muestreos. La clase joven (A) dominó en cada una de las especies de Hamadryas involucradas en este estudio durante los últimos muestreos, mientras las clases intermedias (B) y vieja (C) presentaron una mayor frecuencia durante los primeros muestreos para cada una de las especies. De esta manera, la estructura de edades no fue constante en ninguna de las especies de estudio, presentando fluctuaciones durante los eventos de captura dentro del paisaje en la VLF (fig. 2).
De acuerdo con los valores de ASR de las 4 especies estudiadas, se determina una predominancia de los machos para cada una de las especies de Hamadryas durante los eventos de muestreos. El solapamiento de los intervalos de confianza (IC de 95%) indica que no hubo diferencias significativas en los valores de ASR observados (fig. 3). Para H. amphichloe, el rango de variación de los IC del ASR fue mayor durante la época de lluvias (M3, M4, M5, M6) con una clara dominancia de los machos. En cuanto a la especie H. amphinome, se determinó una alta dominancia de los machos desde el M2 (mediados de marzo), mientras que la especie H. februa presentó una tendencia hacía los machos en la época seca (M1, M2) y con respecto a la época de las primeras lluvias, se determinó una tendencia hacia las hembras (M3). En los últimos periodos (lluvias fuertes) se observó una inclinación hacia las hembras en los valores de ASR observados. Por otro lado, para la especie H. feronia se determinó una dominancia de los machos en la temporada seca y primeras precipitaciones, mientras que durante los últimos periodos de muestreo (lluvias fuertes) se determinó una alta frecuencia de las hembras influenciadas por las lluvias (M5, M6) (fig. 3).
Dentro de las distancias recorridas promedio estimadas, H. feronia fue la especie con el menor valor (127.825 m; IC = 43.15), sin embargo, presentó la tasa media de movimiento más alta (0.163 m/días). El individuo con mayor desplazamiento fue el registrado como 434D con 476 m entre la E6 hasta la E7 (tabla 3). La especie H. februa presentó un valor promedio de desplazamiento de 208.846 m (IC = 99.63); el individuo con mayor distancia recorrida fue el 38A con 820 m, desde E5 (bosque) hasta E7 (cultivos mixtos), mientras que la tasa media de desplazamiento estimada para la especie fue de 0.095 m/días.
Para H. amphichloe se determinó un valor promedio desplazado de 285.6 m (IC = ± 98.42) y el individuo con la mayor distancia recorrida dentro de todo el paisaje fue un macho (marca 18A), con aproximadamente 564 m desde la E4 (bosque) hasta la E2 (borde). La tasa media de desplazamiento estimada para la especie fue de 0.070 m/días, mientras que para H. amphinome, el promedio de desplazamiento presentado fue de 278.5 m (IC = ± 66.56), y el individuo con mayor registro de vuelo fue el 230 B con 607 m recorridos desde E5 (bosque) hasta E6 (vegetación secundaria). Esta especie presentó la tasa media de desplazamiento más baja con 0.067 m/días (tabla 3). Se destaca que no se registraron desplazamientos entre las E1 (cultivos mixtos con remanentes de vegetación) con el resto de estaciones para ninguna de las especies.
Tabla 1
Variación de la abundancia de Hamadryas durante los muestreos y estaciones en un paisaje en la vereda la Flecha (VLF), San Jacinto, Bolívar. Abreviaturas: muestreo (M), estación (E), números en paréntesis (macho-hembra). Los muestreos M1 y M2 corresponden a la época seca y los restantes (M3 hasta M6) a la época de lluvias. E1 corresponde a zona de cultivo mixto permanente temporales (CMPT), E2 a zona de borde de bosque (BB), E3, E4 y E5 son zonas de bosque (Bos), E6 es vegetación secundaria (VS) y E7 es zona de cultivo mixto permanente (CMP).
| Especie | E | M1
(17 mar) |
M2
(3 abr) |
M3
(14 abr) |
M4
(28 abr) |
M5
(9 may) |
M6
(2 jun) |
Total |
| H. feronia | E1 | 3 (2-1) | 1 (1-0) | 5 (2-3) | 2 (2-0) | 5 (4-1) | 0 (0-0) | 16 (11-5) |
| E2 | 7 (4-3) | 29 (14-15) | 26 (15-11) | 15 (11-4) | 8 (2-6) | 3 (0-3) | 88 (46-42) | |
| E3 | 4 (3-1) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 4 (3-1) | |
| E4 | 4 (2-2) | 3 (1-2) | 0 (0-0) | 2 (1-1) | 0 (0-0) | 0 (0-0) | 9 (4-5) | |
| E5 | 6 (3-3) | 3 (2-1) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 9 (5-4) | |
| E6 | 1 (0-1) | 17 (9-8) | 9 (8-1) | 3 (2-1) | 1 (0-1) | 0 (0-0) | 31 (19-12) | |
| E7 | 26 (12-14) | 37 (21-16) | 32 (19-13) | 26 (18-8) | 11 (6-5) | 3 (2-1) | 135 (78-57) | |
| Total | 51 (26-25) | 90 (48-42 | 63 (44-28) | 48 (34-14) | 25 (12-13) | 6 (2-4) | 292 (166-126) | |
| H. februa | E1 | 3 (2-1) | 0 (0-0) | 0 (0-0) | 1 (0-1) | 0 (0-0) | 0 (0-0) | 4 (2-2) |
| E2 | 15 (10-5) | 22 (14-8) | 19 (9-10) | 10 (6-4) | 3 (2-1) | 6 (5-1) | 75 (46-29) | |
| E3 | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | |
| E4 | 19 (10-9) | 3 (2-1) | 1 (0-1) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 23 (12-11) | |
| E5 | 11 (5-6) | 2 (2-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 13 (7-6) | |
| E6 | 5 (2-3) | 5 (4-1) | 0 (0-0) | 1 (0-1) | 1 (0-1) | 0 (0-0) | 12 (6-6) | |
| E7 | 7 (4-3) | 5 (2-3) | 3 (1-2) | 2 (2-0) | 1 (1-0) | 2 (2-0) | 20 (12-8) | |
| Total | 60 (33-27) | 37 (24-13) | 23 (10-13) | 14 (8-6) | 5 (3-2) | 8 (7-1) | 147 (85-62) | |
| H. amphichloe | E1 | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 2 (1-1) | 0 (0-0) | 2 (1-1) |
| E2 | 0 (0-0) | 13 (8-5) | 14 (9-5) | 5 (2-3) | 7 (5-2) | 1 (1-0) | 40 (25-15) | |
| E3 | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | |
| E4 | 5 (3-2) | 2 (1-1) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 7 (4-3) | |
| E5 | 0 (0-0) | 0 (0-0) | 1 (1-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 1 (1-0) | |
| E6 | 0 (0-0) | 5 (2-3) | 6 (3-3) | 3 (1-2) | 1 (1-0) | 0 (0-0) | 15 (7-8) | |
| E7 | 2 (1-1) | 0 (0-0) | 5 (1-4) | 3 (2-1) | 4 (4-0) | 1 (1-0) | 15 (9-6) | |
| Total | 7 (4-3) | 20 (11-9) | 26 (14-12) | 11 (5-6) | 14 (11-3) | 2 (2-0) | 80 (47-33) | |
| H. amphinome | E1 | 0 (0-0) | 0 (0-0) | 1 (1-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 1 (1-0) |
| E2 | 0 (0-0) | 0 (0-0) | 3 (2-1) | 0 (0-0) | 1 (1-0) | 0 (0-0) | 4 (3-1) | |
| E3 | 1 (0-1) | 0 (0-0) | 1 (0-1) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 2 (0-2) | |
| E4 | 2 (1-1) | 1 (1-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 3 (2-1) | |
| E5 | 4 (0-4) | 1 (1-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 0 (0-0) | 5 (1-4) | |
| E6 | 0 (0-0) | 0 (0-0) | 1 (1-0) | 1 (1-0) | 1 (1-0) | 1 (1-0) | 4 (4-0) | |
| E7 | 0 (0-0) | 1 (1-0) | 6 (5-1) | 2 (1-1) | 4 (4-0) | 0 (0-0) | 13 (11-2) | |
| Total | 7 (1-6) | 3 (3-0) | 12 (9-3) | 3 (2-1) | 5 (5-0) | 1 (1-0) | 31 (21-10) |
Relación entre la abundancia de Hamadryas con las variables ambientales y la cobertura dosel. Con los MLG se determinó que el uso del suelo y la época, así como su interacción con la CD, presentaron un efecto estadísticamente significativo (p < 0.05) sobre la abundancia de Hamadryas. Así mismo, se determinó que las variaciones de la CD, teniendo en cuenta los usos de suelo, influyen en la variación de la abundancia de la especie H. feronia. Con respecto a la especie H. amphichloe, tanto la época como los diferentes usos de suelo fueron significativos en la explicación de la variación de su abundancia, así como los cambios en la cobertura del dosel por la interacción de los muestreos (épocas) y los usos del suelo en la zona. Por otro lado, para H. februa y H. amphinome los usos del suelo influyen en la variación de la abundancia de estas especies (tabla 4).
Tabla 2
Tamaño poblacional estimado por el método de Jolly-Seber en el paisaje en la VLF para las especies H. feronia y H. februa. Øt: Probabilidad de supervivencia; Bt: número de ganancias; EEØt: error estándar supervivencia; EEBt: error estándar número de las ganancias.
| H. feronia | H. februa | |||||
| Muestreos | Proporción marcada | Øt (EEØt) | Bt (EEBt) | Proporción marcada | Øt (EEØt) | Bt (EEBt) |
| M1 (mar) | 0 | 0.681 (0.22) | 0 (0) | 0 | 0.77 (0.482) | 0 (0) |
| M2 (abr) | 0.111 | 0.67 (0.23) | 120.7 (103.9) | 0.289 | 0.26 (0.325) | 33.5 (46.6) |
| M3 (abr) | 0.233 | 0.29 (0.101) | 30.1 (33.8) | 0.25 | 0.243 (0.383) | 47.2 (86.6) |
| M4 (may) | 0.306 | 0.382 (0.178) | (-)12 (12.3) | 0.133 | 0.093 (0.082) | (-)1.9 (7.5) |
| M5 (may) | 0.769 | 0 (0) | 0 (0) | 0.5 | 0 (0) | 0 (0) |
| M6 (jun) | 0.571 | 0 (0) | 0 (0) | 0.222 | 0 (0) | 0 (0) |
Discusión
El presente trabajo representa el primer estudio ecológico sobre la dinámica poblacional y el desplazamiento de las mariposas del género Hamadryas en diferentes usos del suelo en los Montes de María (San Jacinto, Bolívar). Además, se demuestra el papel de los fragmentos de BST y las matrices adyacentes de cultivos y vegetación secundaria en la dinámica poblacional de las poblaciones de mariposas en el Caribe colombiano (Pizano y García, 2014).
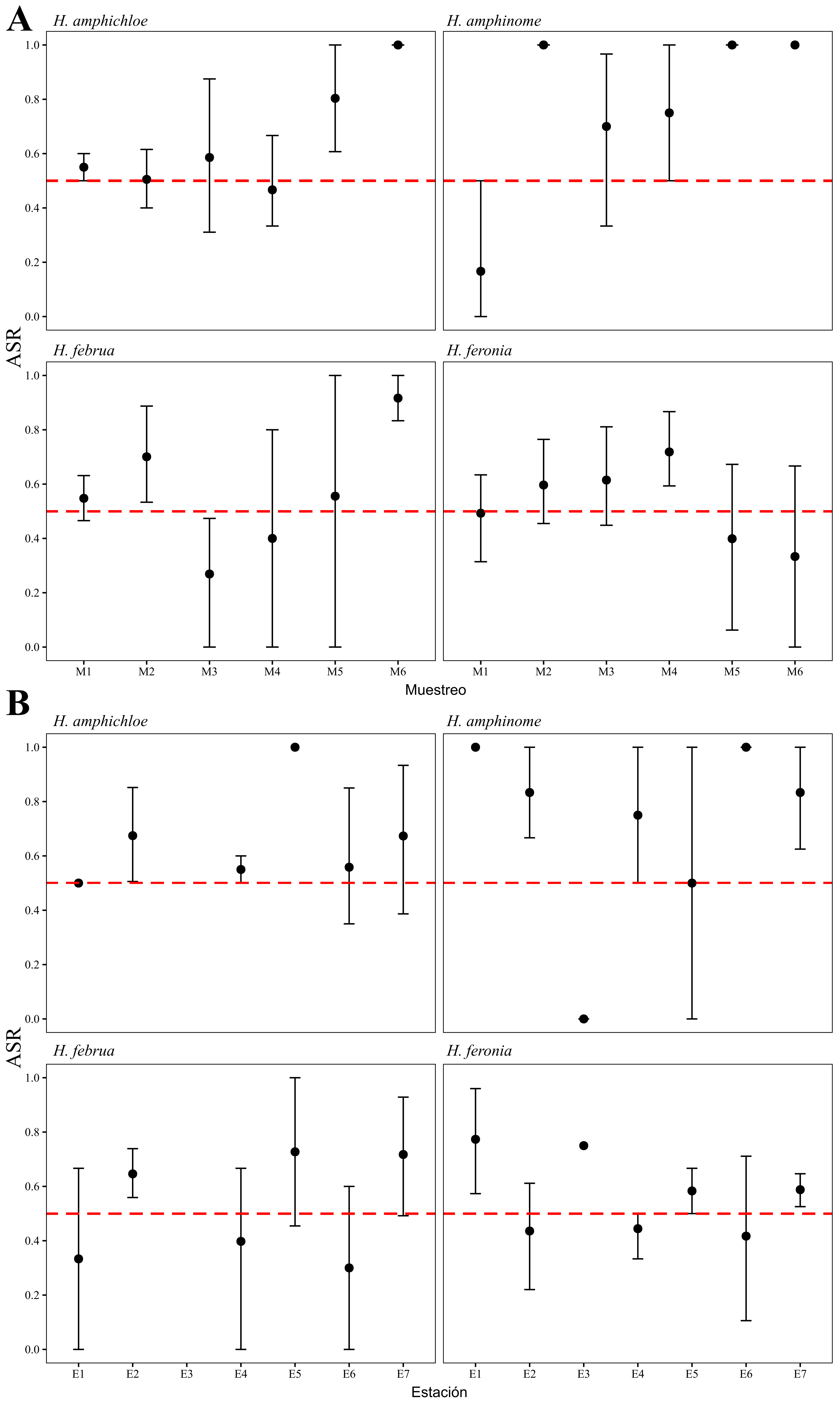
Tabla 3
Desplazamiento de Hamadryas en el paisaje en la VLF. Se indican el número de machos y hembras, las categorías de edad —A (joven), B (intermedio), C (viejo)—, la tasa media de desplazamiento (Tmm; m/días), los individuos recapturados en la misma trampa (S), los valores de movimiento y el tiempo promedio de desplazamiento de las recapturas, así como el número de capturas (C) y recapturas (R) por especie y su porcentaje (%), el número de machos (♂) y hembras (♀) recapturados.
| Especie | Individuos | Sexo | Clase de edad | Movimiento (m/días) | ||||||||||
| C | R | % | ♂ | ♀ | A | B | C | Tmm | S | Media | Mediana | Max | T (días) | |
| H. feronia | 291 | 61 | 20.96 | 34 | 27 | 6 | 21 | 34 | 0.162 | 17 | 20.8 | 18 | 43 | 20.8 |
| H. februa | 137 | 17 | 12.4 | 10 | 7 | 0 | 10 | 7 | 0.095 | 4 | 19.538 | 18 | 28 | 19.538 |
| H. amphichloe | 80 | 5 | 6.25 | 3 | 2 | 0 | 1 | 4 | 0.07 | 0 | 20 | 18 | 27 | 20 |
| H. amphinome | 33 | 4 | 12.2 | 4 | 0 | 1 | 1 | 2 | 0.067 | 0 | 18.75 | 13.5 | 37 | 18.75 |
Tabla 4
Resumen de los resultados de los MLG para la variación de la abundancia del género y cada especie de Hamadryas por uso de suelo, épocas y la CD en el área de estudio. Se presentan las estimaciones factoriales y significancia estadística para la abundancia de las especies y las respectivas interacciones de los factores y la variación de la cobertura vegetal.
| Especies | Variables | LR Chisq | Df | Pr (> Chisq) |
| Hamadryas | Épocas | 114.752 | 3 | < 2.2e-16 *** |
| Uso | 4.009 | 1 | 0.04526 * | |
| CD | 16.564 | 1 | 4.704e-05 *** | |
| Usos x CD | 110.023 | 3 | < 2.2e-16 *** | |
| H. feronia | Épocas | 5.164 | 1 | 0.0230580 * |
| Usos | 153.018 | 3 | < 2.2e-16 *** | |
| CD | 6.628 | 1 | 0.0100410 * | |
| Usos x CD | 24.082 | 3 | 2.401e-05 *** | |
| H. februa | Usos | 49.323 | 3 | 1.113e-10 *** |
| H. amphichloe | Épocas | 5.798 | 1 | 0.0160428 * |
| Usos | 15.608 | 3 | 0.0013641 ** | |
| Épocas x CD | 10.7704 | 1 | 0.0010314 ** | |
| Usos x CD | 12.3598 | 3 | 0.0062470 ** | |
| H. amphinome | Usos | 14.3148 | 3 | 0.002507 ** |
La mayor abundancia de H. februa y H. feronia en usos de suelo como borde de bosque y cultivos mixtos permanentes puede estar relacionada con la disponibilidad de refugio o recursos como frutos en descomposición (fermentación). Por ejemplo, en la zona de cultivos mixtos permanentes (E7), hay presencia de árboles como M. indica, A. muricata, P. americana y palmas como A. butyracea que tienen cortezas grisáceas, los cuales les permiten hacer cripsis con este tipo de sustrato; así como líquenes adheridos que son utilizadas por este grupo de mariposas para camuflarse e interactuar con individuos de la misma especie (Jenkis, 1983; Mercado et al., 2018; Monge-Nájera et al., 1998; Prince et al., 2011; Sargent, 1985; Vargas-Zapata et al., 2015). Adicionalmente, estas 2 especies prefieren zonas de bordes y cultivos con claros expuestos a la luz (observaciones en campo), donde aplican estrategias de comportamiento asociadas con la territorialidad (posar con las alas abiertas contra la superficie y con la cabeza apuntando hacia abajo), y la presencia de suelos desnudos con bastantes arcillas para obtener sales y nutrientes (Devries, 1988; Monge-Nájera et al., 1998). La alta abundancia de estas 2 especies coincide con lo descrito por Montero et al. (2009), Vargas-Zapata et al. (2011, 2015), Cerpa y Flórez (2016), Mercado et al., (2018) y Ahumada et al., (2019), quienes coinciden en que fueron las especies más comunes dentro de este género para zonas de bosque seco en el Caribe colombiano.
La poca abundancia de H. amphichloe y H. amphinome registrada dentro del paisaje puede estar relacionada con la estructura y composición vegetal de las estaciones de muestreos dentro del paisaje. Es posible que esta baja abundancia indique una rareza de hábitat para estas especies de mariposas, ya que pueden presentar interés por perchar y aprovechar otros sitios con algunas condiciones particulares, como troncos de colores oscuros y la presencia de barro con contenido de sales (González et al., 2014; Jenkis, 1983; Monge-Nájera et al., 1998). De este modo, H. amphinome suele posar más frecuentemente en troncos con cortezas oscuras con DAP grande, localizados en el interior de zonas boscosas o adyacentes a márgenes de quebradas. Adicionalmente, esta especie presenta un patrón de coloración distintivo con franjas de color rojizo, las cuales las hacen más vistosas al depredador; razón por la que requiere de hábitats con vegetación más conservada y con áreas boscosas con varios estratos y bastante dosel (Moreno y Acuña, 2015). Lo anterior explica la abundancia alta de esta especie en las estaciones ubicadas en zonas boscosas (E4 y E5) y en E6, esta última localizada en una zona de relictos secundarios de bosque seco con cierto tipo de cobertura vegetal.
La mayor abundancia de H. februa a mediados de marzo, puede atribuirse a que esta especie tenga sincronizado su ciclo reproductivo con las primeras precipitaciones, las cuales favorecen la productividad primaria del bosque, brindando así beneficios a este grupo de mariposas debido al aumento en la disponibilidad de recursos. La baja abundancia de Hamadryas durante las lluvias fuertes (muestreos 4 y 5, valores de precipitación entre 79.5 mm y 113 mm) de finales de abril y mediados de mayo se debe a que en la zona se registraron altos niveles de nubosidad (poca luminosidad), los cuales pueden producir disminución de la capacidad de vuelo y sus periodos de actividad (Kingsolver, 1983). Adicionalmente, durante este periodo, debido a la influencia de las lluvias se deteriora el estado alar de las mariposas con envergadura grande como las Hamadryas (Jenkis, 1983; Monge-Nájera et al., 1998), factor que incide negativamente en las rutas de vuelo, desplazamiento y en la disminución de sus poblaciones durante este periodo. Conjuntamente, con la influencia de las lluvias fuertes se disminuye el efecto del atrayente y de los frutos fermentados naturales del bosque, lo cual disminuye la probabilidad de captura de estas mariposas durante este periodo.
Los resultados observados indican que las poblaciones se comportan como poblaciones abiertas en el área y que éstas pueden estar reguladas por factores intrínsecos como son la tasa de natalidad, fecundidad, supervivencia y mortalidad (Graziano et al., 2002). De este modo, algunas zonas son utilizadas como fuentes de recursos (interior del bosque, zona de policultivos), mientras que los nuevos individuos dentro de las estaciones pueden deberse a un pico de emergencia de los imagos o a la entrada de nuevos individuos provenientes de áreas adyacentes. Así mismo, los diferentes usos de suelo representan atributos significativos para estas poblaciones incidiendo así en sus rutas de vuelo y en el flujo de entrada y salida de individuos, por lo que los adultos suelen asociarse con lugares que brinden un refugio con recursos óptimos para subsistir y completar su ciclo.
La variación en el porcentaje de recapturas entre cada una de las especies de Hamadryas puede estar relacionado con las tasas de dispersión de estas mariposas. Estas especies pueden experimentar distintas rutas de vuelo dentro de la matriz debido a la distribución aleatoria o al patrón de disposición de los recursos (Rueda y Andrade, 2016; Silva y Hernández, 2015). Además, según Jenkis (1983), Monge-Nájera et al. (1998) y Murillo y Córdoba (2013), estas mariposas suelen desplazarse a distintos sitios de acuerdo con la disponibilidad de recursos alimenticios, zonas para la ovoposición y asociadas al camuflaje. Por ejemplo, la mayor frecuencia en las zonas con cultivos permanentes se puede atribuir a que estos lugares le brindan una alta cantidad de recursos que podrían ser explotados por especies como H. feronia y H. februa, con árboles aprovechables para su desarrollo y distintas actividades como el apareamiento. Así mismo, la zona de borde presentó una alta representatividad de individuos recapturados para cada una de las especies monitoreadas y esto estaría asociado con la distribución de especies vegetales típicas de las zonas de bosque y de áreas abiertas, factor que permite a los adultos de las Hamadryas desplazarse hacia el interior del bosque y utilizar esta zona como área de corredor biológico.
La residencia estimada en cada una de las especies presentó variaciones, lo cual puede atribuirse a que este grupo de especies puede tener varias generaciones durante el año (Cerpa y Flórez, 2016; Moreno y Acuña, 2015). Estas especies se caracterizan por presentar ciclos de vida relativamente cortos, donde la mayor duración (alrededor de 15 días) la experimentan durante su fase larval (Brown, 1991; Jenkis, 1983; Kremen, 1992; Pearson, 1994); de esta manera, los tiempos de residencia muy cortos de los adultos en esta zona puede ser un indicativo del carácter multivoltino de estas especies. El menor tiempo de residencia de H. amphinome se atribuye a la fidelidad que tiene esta especie por permanecer en puntos fijos en el interior del bosque, donde se encuentren troncos que presentan tonalidades más oscuras y por estratos altos con bastante dosel (Monge-Nájera, 1998).
Con respecto a las proporciones sexuales para cada una de las especies, la mayor proporción de machos coincide con lo descrito en lepidópteros, donde es más común encontrar una mayor proporción de machos que de hembras en los bosques tropicales (Ramos y Freitas, 1999). El mayor número de machos estaría asociado a patrones en el comportamiento entre los sexos, lo que influye en su probabilidad de recaptura (Ehrlich y Gilbert, 1973; Marini-Filho y Benson, 2010; Young, 1982). Los machos de estas especies se observaron emitiendo los sonidos típicos “clicks” en las diferentes estaciones de muestreo, lo que se asocia a funciones de defensa, territorialidad y cortejo (González et al., 2014; Yack et al., 2000). Este comportamiento hace que muchos machos permanezcan cerca de sus territorios, lo que aumenta su probabilidad de ser recapturados en comparación a las hembras.
Por otro lado, el hecho de que los machos de cada una de las especies se hayan desplazados menos entre estaciones que las hembras, se asocia a que los primeros suelen patrullar y defender las zonas (territorialidad) que les brinden mayores recursos y frecuentar los sitios que les permitan ser más visibles para las hembras (García et al., 2002; Marini-Filho y Benson, 2010; Yack et al., 2000). Además, algunas hembras del grupo se observaron en estratos medios y altos, asociadas a la búsqueda de sitios óptimos con plantas hospederas para la ovoposición, lo que puede aumentar las distancias de desplazamiento. Por otro lado, las hembras permanecen un menor tiempo en las zonas con respecto a los machos, debido a que muchas estarían en la búsqueda de recursos fundamentales para el proceso de maduración de huevos como sales, nutrientes, ácido úrico y azucares (Baker, 1972; Gilbert, 1975; Gilbert y Singer, 1975; Jones, 1884; Nieves et al., 2015; Sterry, 1997).
La variación temporal de la abundancia de Hamadryas en la VLF (apéndice), indica que hay una tendencia a aumentar con las primeras lluvias. Conforme llegan las lluvias, la fenología del paisaje cambia (mayor cobertura vegetal), incidiendo de manera indirecta en la dinámica poblacional de estas mariposas, especialmente en los muestreos posteriores (Vargas-Zapata et al., 2011). A su vez, en la transición de época seca a lluvias se observaron cambios en la fenología y cobertura de la vegetación como arbustos, plantaciones y enredaderas (observaciones en campo), lo que permite que el paisaje sea más homogéneo y el desplazamiento de estas mariposas se incremente. Esto concuerda con Ramírez et al. (2014) y Vargas-Zapata et al. (2015), quienes reportan mayor abundancia de mariposas Hamadryas para BST en la época de las primeras lluvias. La mayor frecuencia de H. feronia y H. februa durante la época seca se debe a que estas especies pueden aprovechar las zonas abiertas con altos valores de intensidad lumínica, debido a sus condiciones heliotérmicas. A su vez, ésto concuerda con Ribeiro et al. (2012), quienes indican que la distribución y la abundancia de mariposas no son aleatorias, sino que siguen un patrón de distribución de recursos que está relacionado con aquellos hábitats con condiciones óptimas para llevar a cabo sus funciones biológicas.
En conclusión, las poblaciones de Hamadryas presentaron variación espacial (usos) y temporal (muestreos) en la vereda La Flecha; lo que demuestra que la heterogeneidad del paisaje y la estacionalidad juegan un papel importante en la dinámica poblacional de estas especies. Las poblaciones de Hamadryas se desplazan entre áreas de bordes, plantaciones y vegetación secundaria dentro del paisaje, sugiriendo que el movimiento de los individuos está determinado por los cambios de cobertura entre las distintas áreas. Sumado a lo anterior, el desplazamiento de las hembras está asociado a los patrones de disposición de las plantas hospederas y distribución de los recursos óptimos para la ovoposición.
Agradecimientos
El primer autor agradece a Johana Márquez, José D. Sarmiento y Kevin Caballero por su colaboración durante la fase de campo. BMP y NJMH agradecen a Dylan Padilla Pérez, beneficiario de la Convocatoria 775 de 2017 “Jóvenes investigadores e innovadores por la paz 2017” del Departamento Administrativo de Ciencia, Tecnología e Innovación, COLCIENCIAS, Colombia y la Universidad del Atlántico, enmarcadas dentro del macroproyecto “Diversidad, distribución y algunos tópicos comportamentales de grupos selectos de artrópodos en zonas de posconflicto en el Caribe colombiano: mariposas fruteras de la familia Nymphalidae”, por la financiación del proyecto. De igual modo, se agradece a la primera convocatoria interna para el fortalecimiento de la Red Institucional de Semilleros de Investigación -2019; proyecto “Abundancia y desplazamiento de Hamadryas Hübner. 1806 (Lepidoptera: Nymphalidae: Biblidinae), en un paisaje de bosque seco tropical en San Jacinto, Bolívar” CB152-CIS2019. Finalmente, agradecemos Oscar García y a Adalberto García, quienes nos permitieron el ingreso, nos brindaron hospedaje dentro de la reserva y siempre estuvieron dispuestos a ayudar en la realización del proyecto.
Apéndice. Variables ambientales registradas en la VLF por estaciones y muestreos. Se indican las fechas de muestreo (M), los usos de suelo y estaciones (Usos-E), valores de temperatura ambiente (T.A), humedad relativa (H.R), intensidad lumínica (IL), cobertura del dosel (C.D.) y precipitaciones (Prec). CMPT = Cultivo mixto permanente temporales), BB = borde de bosque, Bos = bosque, VS = vegetación secundaria, CMP = cultivo mixto permanente.
| M | Usos-E | T.A (°C) | H.R.% | I.L. (KLUX) | C.D. (%) | Prec (mm) |
| mar-17 | CMPT-E1 | 30.03 ±0.99 (28.7 – 31.1) | 68.14 ±4.82 (61.1-73.06) | 0.42±0.21 (0.15-0.66) | 80.62±1.73 (18-22) | 0 |
| BB-E2 | 34.69 ±2.56 (30.7-37.06) | 49.76 ±6.63 (42.9-58.8) | 9.55±4.87 (5.8-17.4) | 35.94±5.47 (3.7-18) | ||
| Bos-E3 | 32.93 ±0.81 (32.1-34.1) | 53.1 ±1.60 (50.5-54.7) | 0.34±0.17 (0.15-0.62) | 94.18±0.89 (21.5-23.5) | ||
| Bos-E4 | 35.95 ±1.10 (34.3-37.2) | 42.85 ±1.47 (40.9-44.7) | 0.82±0.46 (0.37-1.5) | 59.90±3.02 (11-18.5) | ||
| Bos-E5 | 35.47 ±0.66 (34.8-36.4) | 42.22 ±2.65 (39.8-47.1) | 0.32±0.11 (0.18-0.49) | 84.03±5.20 (11-23.5) | ||
| VS-E6 | 31.49 ±1.58 (29.4-33.4) | 55.29 ±4.68 (49.4-59.5) | 0.32±0.18 (0.17-0.59) | 82.11±0.81 (19-20.7) | ||
| CMP-E7 | 29.6 ±0.66 (28.9-30.6) | 59.15 ±0.83 (58.5-60.5) | 0.40±0.19 (0.25-0.63) | 89.93±0.95 (20.7-22.7) | ||
| abr-03 | CMPT-E1 | 27.97 ±1.65 (26.3-29.7) | 79 ±4.3 (74.4-85.08) | 2.29±0.56 (1.9-3.09) | 92.85±1.21 (21.2-24) | 6 |
| BB-E2 | 32.58 ±2.65 (28.6-35.1) | 60.95 ±7.48 (52.6-69.5) | 0.92±0.36 (0.53-1.44) | 78.29±2.21 (15.2-21.2) | ||
| Bos-E3 | 31.65 ±1.00 (30.7-33.2) | 66.19 ±4.50 (61-72.9) | 1.02±0.57 (0.3-1.7) | 97.76±0.27 (23.2-23.7) | ||
| Bos-E4 | 31.46 ±1.03 (30.2-33.06) | 67.9 ±7.09 (56.9-73.6) | 6.65±9.22 (0.88-23) | 85.52±2.03 (17.2-22.2) | ||
| Bos-E5 | 32.05 ±1.99 (28.5-33.3) | 58.16 ±1.91 (55.4-60.2) | 2.06±1.50 (0.74-4.6) | 96.42±0.94 (21.7-24) | ||
| VS-E6 | 30.9 ±0.27 (30.6-31.2) | 72.76 ±2.08 (70.3-74.7) | 4.12±2.48 (0.97-7.9) | 89.52±1.7 (19.5-23.7) | ||
| CMP-E7 | 28.64 ±0.76 (28.01-29.9) | 73.51 ±3.59 (70.7-79.7) | 1.11±0.38 (0.73-1.6) | 97.2±0.59 (22.7-249 | ||
| abr-14 | CMPT-E1 | 25.38 ±0.38 (24.8-25.8) | 83.88 ±1.36 (81.6-85.1) | 7.36±5.11 (1.03-14.6) | 92.85±0.90 (21.2-23.2) | 60 |
| BB-E2 | 29.97 ±0.70 (28.9-30.7) | 68.59±3.33 (64.9-73.08) | 7.26±5.34 (1.8-15.9) | 94.59±1.07 (21-23.7) | ||
| Bos-E3 | 29.46 ±0.34 (29-29.7) | 66.7 ±2.17 (63.1-68.5) | 0.71±0.41 (0.16-1.3) | 94.51±0.63 (21.7-23.2) | ||
| Bos-E4 | 29.04 ±1.22 (28.01-30.6) | 70.69±3.75 (65.1-74.4) | 0.6±0.16 (0.43-0.78) | 93.26±1.03 (20.7-23.2) | ||
| Bos-E5 | 30.88 ±0.35 (30.3-31.2) | 62.58±4.04 (58.3-68.03) | 0,5±0.16 (0.26-0.7) | 96.17±0.40 (22.7-23.5) | ||
| VS-E6 | 32.31 ±0.23 (31.9-32.5) | 59.83±1.49 (58.06-61.8) | 0.52±0.31 (0.25-0.95) | 95.59±0.55 (22.2-23.5) | ||
| CMP-E7 | 32.11 ±0.48 (31.3-32.4) | 60.2±1.97 (58.9-63.6) | 0.82±0.40 (0.18-1.2) | 95.09±0.97 (21.5-23.7) | ||
| abr-28 | CMPT-E1 | 28.19 ±0.53 (27.4-28.7) | 79.58±2.70 (75.4-82.9) | 1.6±0.86 (0.76-3) | 94.09±1.07 (21.2-24) | 79.5 |
| BB-E2 | 27.78 ±0.38 (27.5-28.4) | 77.60±3.19 (72.2-80.6) | 1.88±0.98 (1.01-3.5) | 95.43±1.31 (21-24) | ||
| Bos-E3 | 27.53 ±0.37 (27.1-28.1) | 82.48±2.39 (79.4-86.01) | 1.17±0.75 (0.4-2.3) | 98.09±0.60 (22.7-24) | ||
| Bos-E4 | 26.85 ±0.64 (26.1-27.8) | 83.88±3.05 (80-88.4) | 0.84±0.38 (0.55-1.5) | 98.25±0.37 (23-24) | ||
| Bos-E5 | 27.59 ±0.37 (27.1-28.1) | 80.05±1.38 (78.08-81.4) | 0.78±0.16 (0.61-1.05) | 98.50±0.34 (23.2-24) | ||
| VS-E6 | 27.17 ±0.67 (26.5-28.2) | 85.22±3.04 (80.4-88.8) | 1.21±0.64 (0.76-2.3) | 95.76±0.40 (22.5-23.5) | ||
| CMP-E7 | 25.66 ±1.16 (23.9-26.9) | 85.83 ±1.42 (84.4-88) | 0.57±0.42 (0.23-1.1) | 98.50±0.34 (23.2-24) | ||
| may-09 | CMPT-E1 | 28.63 ±4.56 (25.4-36.6) | 82.27 ±2.72 (80.1-86.6) | 1.36±0.44 (0.71-1.9) | 97.67±0.32 (23.2-24) | 113 |
| BB-E2 | 22.22 ±2.82 (20.7-24.6) | 82.24±3.13 (89.4-96.8) | 1.36±0.51 (0.15-1.4) | 97.67±0.65 (22.5-24) | ||
| Bos-E3 | 24.22 ±0.80 (23.6-25.6) | 93.09 ±2.24 (91.2-96.7) | 0.84±0.47 (0.41-1.6) | 98.50±0.43 (23.2-24) | ||
| Bos-E4 | 24.70 ±0.40 (24.2-25.2) | 87.91 ±4.08 (82.1-92.7) | 0.73 ±0.27 (0.38-1.1) | 98.42±0.28 (23.2-24) | ||
| Bos-E5 | 25.37 ±0.52 (24.7-26.03) | 87.23 ±2.51 (83.5-90.2) | 0.77±0.19 (0.53-1) | 99.09±0.16 (23.7-24) | ||
| VS-E6 | 26.01±0.44 (25.4-26.5) | 90.81 ±0.94 (89.3-91.5) | 1.17±0.57 (0.51-1.8) | 97.26±0.79 (22-24) | ||
| CMP-E7 | 27.27 ±0.54 (26.4-27.7) | 79.56 2.49 (75.7-81.5) | 0.69±0.35 (0.39-1.3) | 23.75±0.32 (23.2-24) | ||
| jun-02 | CMPT-E1 | 27.83 ±0.68 (26.8-28.6) | 82.76 ±2.76 (81.1-87.6) | 0.77±0.22 (0.46-1.1) | 98.59±0.44 (23-24) | 5 |
| BB-E2 | 29.93 ±0.65 (29-30.8) | 80.96 ±2.82 (77.7-84.1) | 0.83±0.83 (0.24-2.2) | 97.92±0.77 (22.2-24) | ||
| Apéndice. Continúa | ||||||
| M | Usos-E | T.A (°C) | H.R.% | I.L. (KLUX) | C.D. (%) | Prec (mm) |
| Bos-E3 | 30.49 ±0.57 (29.5-31.01) | 78.89 ±3.66 (74.1-83.4) | 0.46±0.37 (0.14-1.1) | 98.34±0.49 (23-24) | ||
| Bos-E4 | 29.5 ±0.05 (29.4-29.5) | 83.55 ±2.55 (80.2-86.1) | 0.34±0.09 (0.21-0.48) | 98.34±0.49 (23-24) | ||
| Bos-E5 | 29.76 ±0.18 (29.5-30.01) | 83.74 ±3.36 (80.2-88.5) | 0.29±0.07 (0.19-0.38) | 97.84±0.43 (23.2-24) | ||
| VS-E6 | 30.55 ±0.54 (30-31.3) | 82.09 ±1.47 (79.5-83.1) | 0.6±0.45 (0.31-1.4) | 97.42±0.58 (22.5-24) | ||
| CMP-E7 | 30.53 ±0.29 (30.3-31.01) | 78.52 ±2.29 (74.6-80.05) | 0.97±0.17 (0.71-1.2) | 99.59±0.13 (23.7-24) |
Referencias
Aguilera, M. (2013). Montes de María: una subregión de economía campesina y empresarial. Cartagena, Colombia: Banco de la República, Centro de Estudios Económicos Regionales (CEER). https://doi.org/10.32468/dtseru.195
Ahumada, C. D., Segovia, P. A. y Navas, S. G. (2019). Butterflies (Lepidoptera: Papilionoidea) of Montes de María sub-region: preliminary list of species from El Carmen de Bolívar (Bolívar, Colombia). Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 43, 521–530. https://doi.org/10.18257/raccefyn.808
Andrade, C. M., Campos-Salazar, L., González-Montaña, L. y Pulido, B. H. (2007). Santa María mariposas alas y color: guía de campo. Bogotá D.C.: Universidad Nacional de Colombia. Facultad de Ciencias. Instituto de Ciencias Naturales.
Baker, R. R. (1972). Territorial behaviour of the Nymphalid butterflies, Aglais urticae (L.) and Inachis io (L.). Journal of Animal Ecology, 41, 453–469. https://doi.org/10.2307/3480
Balzarini, M. G., González, L., Tablada, M., Casanoves, F., Di Rienzo, J. A. y Robledo, C. W. (2008). Manual del usuario. Córdoba, Argentina: Editorial Brujas.
Barliza, J. C., León, P. J. y Campo, J. (2018). Recovery of biogeochemical processes in restored tropical dry forest on a coal mine spoil in La Guajira, Colombia. Land Degradation & Development, 29, 3174–3183. https://doi.org/10.1002/ldr.3069
Bell, S. S., Mccoy, E. D. y Mushinsky, H. R. (2012). Habitat structure: the physical arrangement of objects in space. Florida: Springer-Science-Business Media, B.V.
Boom-Urreta, C., Seña-Ramos, L., Vargas-Zapata, M. y Martínez-Hernández, N. (2013). Mariposas Hesperioidea y Papilionoidea (Insecta: Lepidoptera) en un fragmento de bosque seco tropical, Atlántico, Colombia. Boletín Científico Centro de Museos. Museo De Historia Natural, 17, 149–167.
Brown, K. S. (1997). Diversity, disturbance, and sustainable use of Neotropical forests: insects as indicators for conservation monitoring. Journal of Insect Conservation, 1, 25–42.
Brown, K. S. y Freitas, A. V. L. (2002). Butterfly communities of urban forest in Campinas, São Paulo, Brazil: Structure, instability, environmental correlates, and conservation. Journal of Insect Conservation, 6, 217–231. https://doi.org/10.1023/A:1024462523826
Campos, L. R., Gómez, J. y Andrade, C. M. G. (2011). Mariposas (Lepidoptera: Hesperioidea-Papilionoidea) de las áreas circundantes a las ciénagas del departamento de Córdoba, Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 35, 45–60.
Chacón, I. A. y Montero, J. J. (2007). Mariposas de Costa Rica / Butterflies and moths of Costa Rica. Santo Domingo. Revista de Biología Tropical, 56, 366.
Cerpa, J. M. P. y Flórez, G. R. (2016). Mariposas diurnas de tres fragmentos de bosque seco tropical del alto valle del Magdalena. Tolima-Colombia. Revista de la Asociación Colombiana de Ciencias Biológicas, 1, 57–66.
Devries, P. J. (1988). The butterflies of Costa Rica and their natural history. Papilionidae. Vol. I: Papilionidae, Pieridae, Nymphalidae. The Journal of Research on the Lepidoptera, 24, 290–333.
Devries, P. J. y Walla, T. R. (2001). Species diversity and community structure in neotropical fruit- feeding butterflies. Biological Journal of the Linnean Society, 74, 1–15. https://doi.org/10.1111/j.1095-8312.2001.tb01372.x
Ehrlich, P. R. y Gilbert, L. E., (1973). Population structure and dynamics of the Tropical butterfly Heliconius ethilla. Biotropica, 5, 69–82. https://doi.org/10.2307/2989656
Freitas, A. V. L. y Ramos, R. R. (2001). Population biology of Parides anchises nephalion (Papilionidae) in a coastal site in Southeast Brazil. Brazilian Journal of Biology, 61, 623–630. https://doi.org/10.1590/S1519-69842001000400011
García-Robledo, C. A., Constantino, L. M., Heredia, M. D. y Kattan, G. (2002). Guía de campo: mariposas comunes de la cordillera central de Colombia. Valle del Cauca: Wildlife Conservation Society Colombia.
Gilbert, L. E. (1975). Ecological consequences of a coevolved mutualism between butterflies and plants. In Coevolution of Animals and Plants: Symposium V, First International Congress of Systematic and Evolutionary Biology, 1973, 210–240. https://doi.org/10.7560/710313-011
Gilbert, L. E. y Singer, M. C. (1975). Butterfly ecology. Annual Review of Ecology and Systematics, 6, 365–397. https://doi.org/10.1146/annurev.es.06.110175.002053
González, M. R., Isaacs, P., García, H. y Pizano, C. (2014). Memoria técnica para la verificación en campo del mapa de bosque seco tropical en Colombia. Escala 1:100.000. Bogotá, Colombia: Instituto de Investigaciones de Recursos Biológicos “Alexander von Humboldt” – Ministerio de Ambiente y Desarrollo Sostenible.
Graziano, J. V., Pinto, V. M., Collado, J. L. y Robles, R. R. (1997). Ecología de poblaciones de insectos. México D.F.: Colegio de Postgraduados, Institución de Enseñanzas e Investigación en Ciencias Agrícola.
Hammer, Ø., Harper, D. A. T. y Ryan, P. (2001). Past: Paleontological statistics software package for education and data analysis. Paleantologica Electronica, 4, 9.
Hammer, Ø., Harper, D. A. T. y Ryan, P. (2013). PAST: Paleontological satistics. Version 3.0. Reference manual. Oslo: Natural History Museum University of Oslo.
Henderson, P. A. (2003). Practical methods in ecology. Lymington: Blackwell Publishing.
Hernández, C. V. (2008). Aproximación a los avances de cinco entidades en el proceso de implementación del documento. Propuesta técnica para la formulación de un Plan de Acción Nacional en Biodiversidad. Colombia biodiversidad siglo XXI. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Jenkins, D. W. (1983). Neotropical Nymphalidae. I. Revision of Hamadryas. Sarasota: Bulletin of the Allyn Museum.
Jolly, G. M. (1965). Explicit estimates from capture-recapture data with both death and immigration-stochastic model. Biometrika, 52, 225–247. https://doi.org/10.2307/2333826
Jones, E. D. (1884). Metemorphoses of Lepidoptera from San Paulo, Brazil, in the free public museum, Liverpool (with nomenclature and description of new forms by F. Moore). Second series. Liverpool: Museum Report.
Kingsolver, J. G. (1983). Thermoregulation and flight in Colias butterflies: elevational patterns and mechanistic limitations. Ecology, 64, 534–545. https://doi.org/10.2307/1939973
Krebs, C. J. (1989). Ecological methodology. New York: Happer Collins PubF lishers.
Krebs, C. J. (1998). Ecological methodology. New Jersey: Pearson Education, Publisher.
Kremen, C. (1992). Assessing the indicator properties of species assemblages for areas monitoring. Ecological Applications, 2, 203–217. https://doi.org/10.2307/1941776
Kremen, C., Colwell, R. K., Erwin, T. L., Murphy, D. D., Noss, R. A. y Sanjayan, M. A. (1993). Terrestrial arthropod assemblages: their use in conservation planning. Conservation Biology, 7, 796–808. https://doi.org/10.1046/j.1523-1739.1993.740796.x
Lamas, G., Viloria, A. L. y Pyrcz, T. W. (2004). Subtribe Pronophilina. En G. Lamas (Eds.), Atlas of Neotropical Lepidoptera, Checklist: Part 4A, Hesperoidea – Papilionoidea (pp. 206–215). Gainesville: Association for Tropical Lepidoptera.
Lemos, E. J. A., Rojas, G. R. I. y Zúñiga, V. J. J. (2005). Técnicas para el estudio de poblaciones de fauna silvestre. México D.F.: Conabio.
Marini-Filho, O. J. (1996). Defesa de recursos alimentares e interações aéreas entre borboletas x|simpátricas do gênero Hamadryas (Tesis doctoral). Universidade Estadual de Campinas, Instituto de Biologia: Campinas, Brasil.
Marini-Filho, O. J. y BensOn, W. W. (2010). Use of sound and aerial chases in sexual recognition in Neotropical Hamadryas butterflies (Nymphalidae). Journal of Research on the Lepidoptera, 42, 5–12. https://doi.org/10.5962/p.266510
Márquez, J. y Martínez, N. (2020). Estructura poblacional de Morpho helenor peleides Kollar, 1850 (Lepidoptera: Nymphalidae) en un paisaje de bosque seco tropical, departamento de Bolívar, Colombia. Boletín Científico Centro de Museos Museo de Historia Natural, 24, 169–189. https://doi.org/10.17151/bccm.2020.24.2.12
Mercado-Gómez, Y. L., Mercado-Gómez, J. D. y Giraldo-Sánchez, C. E. (2018). Mariposas en un fragmento de bosque seco tropical en Montes de María (Colombia). Ciencia en Desarrollo, 9, 35–45.
Monge-Nájera, J. (1992). Clicking butterflies, Hamadryas, of Panama: their biology and classification (Lepidoptera, Nymphalidae). En D. Quintero y A. Aiello (Eds.), Insects of Panama and Mesoamerica: selected studies (pp. 567–572). Oxford: Oxford University.
Monge-Nájera, J. y Hernández, F. (1991). A morphological search for the sound mechanism of Hamadryas butterflies (Lepidoptera: Nymphalidae). Journal of Research on the Lepidoptera, 30, 196–208. https://doi.org/10.5962/p.266644
Monge-Nájera, J., Hernández, F., González, M. I., Soley, J., Araya, J. y Zolla, S. (1998). Spatial distribution, territoriality and sound production by tropical cryptic butterflies (Hamadryas, Lepidoptera: Nymphalidae): implications for the “industrial melanism” debate. Revista de Biología Tropical, 46, 297–330. https://doi.org/10.15517/rbt.v46i2.19532
Montero, F., Moreno, M. y Gutiérrez, L. C. (2009). Mariposas (Lepidoptera: Hesperioidea y Papilionoidea) asociadas a fragmentos de bosque seco tropical en el departamento del Atlántico, Colombia. Boletín Científico. Centro de Museos. Museo de Historia Natural, 13, 157–173.
Moreno, M. G. y Acuña-Vargas, J. (2015). Caracterización de lepidópteros diurnos en dos sectores del Santuario de Flora y Fauna Los Flamencos (San Lorenzo de Camarones, La Guajira). Boletín Científico Centro de Museos de Historia Natural, 19, 221–234.
Mostacedo, B. y Fredericksen, T. (2000). Manual de métodos básicos de muestreo y análisis en ecología vegetal, Vol. 87. Santa Cruz, Bolivia: Proyecto de Manejo Forestal Sostenible (BOLFOR).
Murillo-Hiller, L. y Córdoba-Alfaro, J. (2013). Taxonomía e historia natural de las mariposas tronadoras Hamadryas spp. (Lepidoptera, Nymphalidae), Revista de Ciencias, 17, 61–66.
Neild, A. F. E. (1996). The butterflies of Venezuela. Part 1: Nymphalidae 1 (Limenitidinae, Apaturinae, Charaxinae). A comprehensive guide to the identification of adult Nymphalidae, Papilionidae and Pieridae. Annals of the Entomological Society of America, 90, 701-702. https://doi.org/10.1093/aesa/90.5.701
Nieves-Uribe, S., Flores-Gallardo, A., Hernández-Mejía, B. y Llorente-Bousquets, J. (2015). Exploración morfológica del corion en Biblidinae (Lepidoptera: Nymphalidae): aspectos filogenéticos y clasificatorios. Southwestern Entomologist, 40, 589–648. https://doi.org/10.3958/059.040.0318
Pearson, D. L. (1994). Selecting indicator taxa for the quantitative assessment of biodiversity. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 345, 75–79. https://doi.org/10.1098/rstb.1994.0088
Pizano, C. y García, H. (2014). Bosque seco tropical en Colombia; generalidades y contexto. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Prieto, C., Takegami, C. y Rivera, J. (2005). Estructura poblacional de Morpho sulkowskyi Kollar, 1850 (Lepidoptera: Nymphalidae) en un sector de la cordillera occidental, departamento del Cauca (Colombia). Entomotropica, 20, 15–22.
Prince-Chacón, S., Vargas-Zapata, M. A., Salazar, J. y Martínez, N. J. (2011). Mariposas Papilionoidea y Hesperioidea (Insecta: Lepidoptera) en dos fragmentos de bosque seco tropical en Corrales de San Luis, Atlántico, Colombia. Boletín de la Sociedad Entomológica Aragonesa, 48, 243–252.
Ramírez, R. L., Chacón, D.U. y Constantino, L. (2007). Diversidad de mariposas diurnas (Lepidoptera: Papilionoidea y Hesperioidea) en Santiago de Cali, Valle del Cauca, Colombia. Revista Colombiana de Entomología, 33, 54–63.
Ramírez, C., Gallusser, S., Lachaume, G. y Blandin, P. (2014). The ecology and life cycle of the Amazonian Morpho cisseis phanodemus Hewitson, 1869, whit a comparative review of early stages in the genus Morpho (Lepidoptera: Nymphalidae: Morphinae). Tropical Lepidoptera Research, 24, 67–80.
Ramos, R. R. y Freitas, A.V. (1999). Population Biology and wing varation in Heliconius erato Phyllis (Nymphalidae). Journal of the Lepidopterists Society, 53, 11–21.
Rangel-Acosta, J. L. y Martínez-Hernández, N. J. (2017). Comparación de los ensamblajes de escarabajos copronecrófagos (Scarabaeidae: Scarabaeinae) entre fragmentos de bosque seco tropical y la matriz adyacente en el departamento del Atlántico-Colombia. Revista Mexicana de Biodiversidad, 88, 389–401. https://doi.org/10.1016/j.rmb.2017.03.012
Reverdin, J. (1914). A hitherto unknown organ in the ancillary appendages of the Lepidoptera (Ageronia sps.). The Entomologist’s Record and Journal of Variation, 27, 97–98.
Ribeiro, D. B., Batista, R., Prado, P. I., Brown, K. S. y Freitas, A. V. (2012). The importance of small scales to the fruit-feeding butterfly assemblages in a fragmented landscape. Biodiversity and Conservation, 21, 811–827. https://doi.org/10.1007/s10531-011-0222-x
Ribeiro, D. B. y Freitas, A. V. (2011). Large-sized insects show stronger seasonality than small-sized ones: a case study of fruit-feeding butterflies. Biological Journal of the Linnean Society, 104, 820–827. https://doi.org/10.1111/j.1095-8312.2011.01771.x
Ribeiro, D. B., Prado, P. I., Brown Jr, K. S. y Freitas, A. V. (2008). Additive partitioning of butterfly diversity in a fragmented landscape: importance of scale and implications for conservation. Diversity and Distributions, 14, 961–968. https://doi.org/10.1111/j.1472-4642.2008.00505.x
Ribeiro, D. B., Prado, P. I., Brown Jr., K. S. y Freitas, A. V. (2010). Temporal diversity patterns and phenology in fruit-feeding butterflies in the Atlantic Forest. Biotropica, 42, 710–716. https://doi.org/10.1111/j.1744-7429.2010.00648.x
Ribeiro, M. C., Metzger, J. P., Martensen, A. C., Ponzoni, F. J. y Hirota, M. M. (2009). The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biological Conservation, 142, 1141–1153. https://doi.org/10.1016/j.biocon.2009.02.021
Rueda, R. N. M. (2016). Parámetros poblacionales y aportes para la conservación de las especies del género Heliconius Kluk, 1780 en hábitats de diferente grado de conservación de la amazonia colombiana (Tesis doctoral). Universidad Nacional de Colombia. Bogotá, Colombia.
Sambhu, H., Nankishore, A., Turton, S. M. y Northfield, T. D. (2018). Trade-offs for butterfly alpha and beta diversity in human-modified landscapes and tropical rainforests. Ecology and Evolution, 8, 12918–12928. https://doi.org/10.1002/ece3.4732
Sargent, T. D. (1985). Melanism in Phigalia titea (Cramer) (Lepidoptera: Geometridae) in southern New England: A response to forest disturbance? Journal of the New York Entomological Society, 93, 1113–1120.
Silva, P. G. D. y Hernández, M. I. M. (2015). Spatial patterns of movement of dung beetle species in a tropical forest suggest a new trap spacing for dung beetle biodiversity studies. Plos One 10, e0126112. https://doi.org/10.1371/journal.pone.0126112
Sterry, P. (1997). Insects a portrait of the animal world. New York: New Line Books.
Townsend, D. I., Masters, R. E., Lochmiller, R. L., Leslie, D. M., Demaso, S. J. y Peoples, A. D. (2001). Characteristics of nest sities of northern bobwhites in western Oklahoma. Rangeland Ecology & Management/Journal of Range Management Archives, 54, 260–264. https://doi.org/10.2307/4003244
Vargas-Zapata, M. A., Boom-Urueta, C. J., Seña-Ramos, L. I., Echeverry-Iglesias, A. L. y Martínez-Hernández, N. J. (2015). Composición vegetal, preferencias alimenticias y abundancia de Biblidinae (Lepidoptera: Nymphalidae) en un fragmento de bosque seco tropical en el departamento del Atlántico, Colombia. Acta Biológica Colombiana, 20, 79–92. https://doi.org/10.15446/Abc.V20n3.42545
Vargas-Zapata, M. A., Martínez-Hernández, N. J., Gutiérrez-Moreno, L. C., Prince-Chacón, S., Herrera-Colón, V. y Torres-Periñan, L. F. (2011). Riqueza y abundancia de Hesperioidea y Papilionoidea (Lepidoptera) en la Reserva Natural Las Delicias, Santa Marta, Magdalena, Colombia. Acta Biológica Colombiana, 16, 43–60.
Vargas-Zapata, M. A., Prince-Chacón, S. y Martínez-Hernández, N. J. (2012). Estructura poblacional de Heliconius erato hydara Hewitson, 1867 (Lepidoptera: Nymphalidae) en La Reserva Campesina La Montaña (RCM), departamento del Atlántico, Colombia. Boletín de la Sociedad Entomológica Aragonesa (S.E.A.), 51, 273–281.
Warren, A. D., Davis, K. J., Stangeland, E. M., Pelham, J. P. y Grishin, N. V. (2013). Illustrated lists of American butterflies. Recuperado el 01 de agosto, 2023 de: http://www.butterfliesofamerica.com/
Wilson, K. y Hardy, I. C. (2002). Statistical analysis of sex ratios: an introduction. En I. Hardy (Eds.), Sex ratios: concepts and research methods (pp. 48–92). Cambridge, Cambridge: University Press.
Yack, J. E., Otero, L. D., Dawson, J. W., Surlykke, A. y Fullards, J. H. (2000). Sound production and hearing in the blue cracker butterfly Hamadryas feronia (Lepidoptera, Nymphalidae) from Venezuela. Journal of Experimental Biology, 203, 3689–3702. https://doi.org/10.1242/jeb.203.24.3689
Young, A. (1982). Population biology of tropical insects. New York, Boston: Plenum Pub Corp.


