Phenotypic and molecular analysis of Mexican Metarhizium anisopliae strains
Eder Barbier a, b, *, Hannah Nunes c, d, Patrício Adriano da Rocha c, d, Fabiana Lopes Rocha c, e, Pedro Cordeiro-Estrela c
a Departamento de Zoologia, Centro de Biociências, Universidade Federal de Pernambuco, Av. Prof. Moraes Rego, 1235, Cidade Universitária – Recife PE 50670-901, Brazil
b Programa de Pós-graduação em Biologia Animal, Centro de Biociências, Universidade Federal de Pernambuco, Av. Prof. Moraes Rego, 1235, Cidade Universitária – Recife PE 50670-901, Brazil
c Laboratório de Mamíferos, Departamento de Sistemática e Ecologia, Centro de Ciências Exatas e da Natureza, Universidade Federal da Paraíba – João Pessoa PB 58051-900, Brazil
d Programa de Pós-graduação em Ciências Biológicas, Centro de Ciências Exatas e da Natureza, Universidade Federal da Paraíba, Cidade Jardim Universitário, s/n, Castelo Branco – João Pessoa PB 58051-900, Brazil
e Programa de Pós-Graduação em Ecologia e Monitoramento Ambiental, Centro de Ciências Aplicadas e Educação, Universidade Federal da Paraíba, Campus IV – Rio Tinto PB 58297-000, Brazil
*Corresponding author: barbier.eder@gmail.com (E. Barbier)
Abstract
Metarhizium anisopliae is an anamorphic fungus widely studied due to its use as a biological control agent. In Mexico, it is mainly used as an integrated management strategy to control populations of spittlebugs (Hemiptera: Cercopidae) that affect sugarcane fields and cattle pastures. In this study, 13 strains isolated from Aeneolamia spp. (Hemiptera: Cercopidae) from the main areas of sugarcane production of Mexico and 11 reference strains of Metarhizium from different insect hosts and geographic origins were evaluated based on their conidial dimensions, thermotolerance at 15, 25, 30, and 35 ºC, random amplification of polymorphic DNA (RAPD) patterns using a combined analysis with 14 different oligonucleotides and diversity estimators, and a phylogenetic analysis with the ribosomal RNA internal transcribed spacer (ITS) region. All strains used in this study showed typical morphological characteristics corresponding to M. anisopliae and were able to grow at 25 and 30 ºC with restricted growth at low (15 ºC) and high temperatures (35 ºC). Polymorphism analysis clustered all strains from Aeneolamia sp. in one well-defined group with low variability among them. Phylogenetic relationships based on nuclear ribosomal ITS region sequences recovered a subclade within M. anisopliae formed by 10 of the Mexican strains.
Keywords:
Metarhizium; Sugarcane; Cattle pastures; ITS region; RAPD
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Análisis fenotípico y molecular de cepas mexicanas de Metarhizium anisopliae
Resumen
Metarhizium anisopliae es un hongo anamórfico ampliamente estudiado debido a su uso como agente de control biológico. En México se utiliza principalmente como una estrategia de manejo integrado para controlar poblaciones de mosca pinta (Hemiptera: Cercopidae) que afectan los campos de caña de azúcar y pastizales de ganado. Estudios previos muestran evidencia de variabilidad fisiológica y filogenética entre las cepas de Metarhizium anisopliae. En este estudio, 13 cepas aisladas de Aeneolamia spp. (Hemiptera: Cercopidae) que provienen de las principales áreas de producción de caña de azúcar de México y 11 cepas de referencia de Metarhizium de diferentes insectos hospederos y orígenes geográficos, fueron evaluadas en función de sus dimensiones conidiales, su termotolerancia a 15, 25, 30 y 35 ºC, amplificación aleatoria de patrones de ADN polimórfico (RAPD) usando un análisis combinado con 14 oligonucleótidos diferentes, análisis de estimadores de diversidad y un análisis filogenético con la región de los espaciadores internos transcritos (ITS) del gen del ARN ribosómico. Todas las cepas utilizadas en este estudio mostraron características morfológicas típicas correspondientes a M. anisopliae, crecieron a 25 y 30 ºC con restricción a bajas (15 ºC) y altas (35 ºC) temperaturas. El análisis del polimorfismo agrupó todas las cepas de Aeneolamia sp. en un grupo bien definido con baja variabilidad entre ellos. El análisis filogenético evidenció un subclado dentro de M. anisopliae formado por 10 de las cepas mexicanas.
© 2019 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Palabras clave:
Metarhizium; Caña de azúcar; Pastizales de ganado; Región ITS; RAPD
Introduction
Metarhizium anisopliae is an entomopathogenic fungus widely used as a biological control agent throughout the world to regulate insect populations that affect agricultural areas. In Mexico, M. anisopliae is the main agent used against spittlebugs (Hemiptera: Cercopidae) that affect sugarcane (one of Mexico’s most profitable crops) and cattle pastures (Senasica, 2017). Brunner-Mendoza et al. (2017) and Rezende et al. (2015) have demonstrated the variability of the genus Metarhizium. The internal transcribed spacer (ITS) region of nuclear ribosomal DNA was adopted as the primary fungal barcode marker by the Consortium for the Barcode of Life (Schoch et al., 2012), due to its easy amplification and high yield, providing positive results in most phyla. Although ITS sequences have been surpassed at species level resolution, they are still being used to investigate fungal diversity and interactions with each other and with the environment, now with a metagenomic approach (Cuadros-Orellana et al., 2013). On the other hand, the evaluation of cold and heat activity of isolates has been used as a rapid tool to identify Metarhizium species (Fernandes et al., 2009), as well as to demonstrate the relation between fungal genotypes with habitats (Bidochka et al., 2001). In addition, molecular tools have allowed the development of various methods for detecting genetic polymorphisms at the DNA sequence level and aided the understanding of genetic diversity in fungi (Fungaro et al., 1996; Paelowska & Taylor, 2004). Other authors have previously used analysis of sequence data from the ITS region, the 28S rDNA D3 region, and Random Amplification of Polymorphic DNA (RAPD) patterns to reassess the relationships within the Metarhizium genus (Curran et al., 1994; Driver et al., 2000). The RAPD technique has shown advantages when used as an initial screening tool (Kumari & Thakur, 2014), allowing the differentiation of strains even at the intra-species level, and therefore, is more suitable for the rapid discrimination of a high number of newly isolated microorganisms. The aim of this study was to evaluate thermotolerance and conidial size, and present molecular analyses of Mexican Metarhizium anisopliae strains using RAPD and ITS markers. These data will serve as a proposal for their use as routine identification tools, as well as for the characterization of indigenous Metarhizium biodiversity in Mexico.
Table 1
Strains of M. anisopliae from Aeneolamia sp. in sugar cane fields, Mexico.
|
Strains |
Origin (Mexican states) |
GenBank accesion number |
|
EH-467 |
Veracruz |
MH828214 |
|
EH-468 |
Veracruz |
MH828215 |
|
EH-469 |
Veracruz |
MH828216 |
|
EH-470 |
Veracruz |
MH828217 |
|
EH-471 |
San Luis Potosí |
MH828218 |
|
EH-472 |
San Luis Potosí |
MH828219 |
|
EH-473 |
San Luis Potosí |
MH828220 |
|
EH-474 |
San Luis Potosí |
MH828221 |
|
EH-476 |
San Luis Potosí |
MH828222 |
|
EH-477 |
Oaxaca |
MH828223 |
|
EH-478 |
Oaxaca |
MH828224 |
|
EH-479 |
Veracruz |
MH828225 |
|
EH-480 |
Veracruz |
MH828226 |
Materials and methods
The 13 strains used in this study were isolated from infected Aeneolamia spp. coming from the main areas of sugarcane production in the country, obtained from the Colección de Hongos Entomopatógenos of the Centro Nacional de Referencia de Control Biológico (CNRCB), Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (Senasica), Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (Sagarpa), Colima, Mexico (Table 1) and 11 reference strains from the United States of America, Department of Agriculture, Agricultural Research Service (USDA, ARS) and the Commonwealth Scientific and Industrial Research Organization (CSIRO) collections (Table 2). All strains were identified by macro and micro morphology as M. anisopliae. The strains were maintained in sterile water, mineral oil, and liquid nitrogen cryopreservation at the fungal collection of the “Laboratorio de Micología Básica, Departamento de Microbiología y Parasitología, Facultad de Medicina, Universidad Nacional Autónoma de México” (UNAM), registered at the World Federal Culture Collection (WFCC) as BMFM-UNAM 834. To guarantee their purity, single spore cultures were prepared with each isolate from Mexico using the method described by Goettel and Inglis (1997).
Conidial size. Each isolate was grown on PDA (potato dextrose agar, Bioxon, México City) at 28 ºC for 9 days. Thirty randomly selected conidia per isolate were measured (length × width). A one-way Anova (p = 0.05) was calculated to detect size differences.
Thermotolerance. Strain growth was evaluated at different temperatures (15, 25, 30, 35 ºC) on SDYA [1% peptone (Bioxon, México City), 4% dextrose (Droguería Cosmopolita, México City), 1% yeast extract (Yestal, México City), and 1% agar (Bioxon, México City)] plates. Five plates per isolate and per temperature were inoculated with 2 μL of conidial suspension (approximately 105 conidia/mL), incubated for 14 days. These experiments were carried out in triplicate. Fungal growth was calculated as the mean of the perpendicular diameters of colonies at 14 days. Data were subjected to analysis of variance (Anova, p = 0.05) to detect differences between isolates and temperatures. In case of significant differences, the Tukey multiple comparison test was applied.
DNA extraction. The mycelial-phase of each original strain was cultured in a 100-mL YPG broth (1% yeast extract, 1% soy peptone, 4% glucose), incubated at 28 °C at 100 rpm for 5 days. Mycelia were harvested by filtration, washed 3 times with sterile Milli-Q water (Millipore, USA), and dried between 2 sterile paper filters. For the extraction process, the DNeasy Plant Mini Kit was used (Qiagen, USA) as described by the manufacturer. For quantification of DNA, a spectrophotometer (DeNovix Inc., USA) was used, using 2 μL of genomic DNA.
Table 2
Metarhizium reference strains.
|
Name |
ARSEF* |
Host |
Country |
|
CSIRO** |
|||
|
M. album |
1941* |
Nephotettix virescens (Hemiptera) |
Philippines |
|
M. acridum |
FI-985** |
Austracis guttulosa (Orthoptera) |
Australia |
|
M. acridum |
FI-987** |
Ornithacris cavroisi (Orthoptera) |
Nigeria |
|
M. anisopliae |
7450* |
Heteronyx piceus (Coleoptera) |
Australia |
|
M. anisopliae |
FI-1029** |
Schistocerca gregaria (Orthoptera) |
Ethiopia |
|
M. flavoviride |
1184* |
Otiorhynchus sulcatus (Coleoptera) |
France |
|
M. novozealandicum^ |
FI-698** |
Lepidoptera |
New Zealand |
|
M. pemphigi^ |
FI-72** |
Pemphigus treherni (Hemiptera) |
England |
|
M. brasiliense |
2948* |
Hemiptera |
Brazil |
|
M. lepidiotae^ |
FI-147** |
Lepidiota consobrina (Coleoptera) |
Australia |
|
M. majus |
1914* |
Oryctes rhinoceros (Coleoptera) |
Philippines |
|
M. minus^ |
2037* |
Niliparvata lugens (Hemiptera) |
Philippines |
|
M. pingshaense |
7929* |
Cryptotermes brevis (Isoptera) |
Australia |
|
M. robertsii |
2575* |
Curculio caryae (Coleoptera) |
Australia |
*ARSEF = Agricultural Research Service of Entomopathogenic fungi, USA.
** CSIRO = Commonwealth Scientific and Industrial Research Organization, Australia.
Metarhizium names according to USDA-ARS Collection of Entomopathogenic Fungal Cultures ARSEF, Catalog of Strains, compiled 16 January 2014, except for strain FI-72 and ^ Names according to Kepler et al. (2014).
The purity of the genomic DNA was also determined fluorometrically and checked against standard λ-phage (Gibco BRL, USA) concentrations through 0.8% agarose gel electrophoresis with the SYBR®Safe DNA gel stain (ThermoFisher Scientific, USA).
RAPD-PCR. Amplifications were done in 25 μL reactions containing 1X PCR buffer, 2.5 mΜ MgCl2, 0.2 mM of each oligonucleotide, 200 μM dNTPs (Applied Biosystems, USA), 10-50 ng DNA, and 1 U Taq polymerase (Rocher, France). Primers (Table 3) and PCR programs were those used by Cobb and Clarkson (1994) and Driver et al. (2000). A positive control (PCR mix with reference strain CSIRO FI-1029) and negative control (PCR mix without DNA) were used. Reactions were placed in a T100™ thermocycler (BioRad, USA). The amplification products were analyzed through gel electrophoresis with 1.2% agarose with the SYBR®Safe in 0.5 X TBE buffer (45 mM Tris-Base, 45 mM boric acid, 1 mM EDTA). Band profiles were analyzed using a presence/absence matrix. The degree of similarity among isolates was evaluated according to the Jaccard coefficient (Real & Vargas, 1996) and the dendrogram was developed with the “Unweighted Pair Group Method With Arithmetic Mean” (UPGMA) using the combined data of the 14 primers (5,000 replicates). The cophenetic correlation coefficient (CCCr) (Sneath & Sokal, 1973) was calculated to determine the degree of relationship between the dendrogram and original matrices. For the relationship among isolates a multidimensional diagram was constructed by main components. The ordering of the isolates was multi-dimensionally established by analysis of minimum tree lines, using Numerical Taxonomic and Multivariate Analysis System (NTSYS-PC) software (Rholf, 1988). The genetic variability of the RAPD markers was estimated with the Shannon index (S), considering that each phenotypic marker represents a distinct locus (Allnut et al., 1999).
PCR amplification and sequencing of ITS rDNA region. For the amplification of the ITS rDNA region of the 13 isolates studied, TW81 (5´-GTTTCCGTAGGTGAACCTGC-3´) and AB28 (5´-ATATGCTTAAGTTCAGCGGGT-3´) primers were used (Sigma-Aldrich, USA). These oligonucleotides amplify a 601 bp fragment and were previously tested by Curran et al. (1994) and Driver et al. (2000). The PCR reactions were performed in 25 μL reactions containing 1X PCR buffer, 2.0 mM MgCl2, 200 mM dNTPs, 100 pmol/μL of each oligonucleotide, 10-20 ng of genomic DNA, and 1 U Taq polymerase. Amplifications were performed in a BioRad T100™ thermocycler with the program reported by Curran et al. (1994). The amplification products were analyzed through 1.2% agarose gel electrophoresis with SYBR®Safe in 0.5 X TBE buffer (45 mM Tris-Base, 45 mM boric acid, 1 mM EDTA). PCR products were sequenced using the same primers described above with Sanger chemistry in the High Throughput Genomic Center, Seattle, Washington.
Analysis of DNA sequences. For the phylogenetic analysis, 10 sequences were retrieved from GenBank (AF135210, AF137061, AF137062, AF137065, AF137067, AF138267, AF138271, AF139850, AF139851, AF139854) corresponding to the ITS sequences from Metarhizium reference strains, described by Driver et al. (2000). The ITS sequences used for the analysis were edited with Geneious R8.1.5. Alignments were made using MAFFT version 7.017 with default settings. Phylogenetic analysis was developed by Bayesian Inference (BI) with MrBayes version 3.2.6 (Huelsenbeck & Ronquist, 2001) to determine posterior probabilities. The analysis was run 4 times with each run including 4 MCMC chains, 1,100,000 generations, and sampling every 200 generations.
Table 3
RAPD-PCR primers.
|
Primer |
Sequence |
|
H01* |
5’-GGTCGGAGAA-3’ |
|
H02* |
5’-TCGGACGTGA-3’ |
|
F06* |
5’-GGGAATTCGG-3’ |
|
F07* |
5’-CCGATATCCC-3’ |
|
F08* |
5’-GGGATATCGG-3’ |
|
F10* |
5’-GGAAGCTTGG-3’ |
|
OPA01** |
5’-CAGGCCCTTC-3’ |
|
OPA04** |
5’-AATCGGGCTG-3’ |
|
OPA05** |
5’-AGGGGTCTTG-3’ |
|
OPA08** |
5’-GTGACGTAGG-3’ |
|
OPA09** |
5’-GGGTAACGCC-3’ |
|
OPA010** |
5’-GTGATCGCAG-3 |
|
OPA016** |
5’-AGCCAGCGAA-3’ |
|
OPA019** |
5’-CAAACGTCGG-3’ |
* Driver et al. (2000). ** Cobb & Clarkson (1994).
Results
Conidial size. Microscopic examination revealed hyaline hyphae, smooth, septate, predominantly cylindrical, uninuclear conidia with rounded ends forming basipetal chains characteristic of M. anisopliae. The conidial size of isolates from Mexico were 7.35 to 7.94 × 2.56 μm (p < 0.01) (Table 4).
Thermotolerance. All isolates tested in this study were able to grow at 25 and 30 ºC in SDYA medium. At 15 ºC most of the isolates showed a limited growth (< 3 cm diameter), where EH-472, 1941, 2037, 2948, and FI-987 showed the lowest growth values (< 1 cm diameter). On the other hand, at 35 ºC most isolates assayed showed a restricted and slow growth compared to 28 ºC, except for strains 985 (M. acridum) and FI-1029 (M. anisopliae) both isolated from Orthoptera. Their growth was > 2 cm of diameter (Table 5).
Cluster analysis of the RAPD data. The band profiles generated by the amplification using the 14 primers (Table 3) generated 339 informative characters. The profiles among duplicates of reactions were consistent. The cluster analysis of the RAPD data showed the integration of 2 well-defined groups (Fig. 1). Group I include only strains from Aeneolamia sp. collected from different agricultural regions of Mexico, showing 24% similarity among them. Average polymorphism was 41.59% and the number of effective alleles was 1.13 (SD = 0.250). The value for genetic diversity estimated from the Shanon index (a value not sensitive to the number of strains analyzed) was 0.1461 (SD = 0.214). On the other hand, group II clustered all reference strains of different species, hosts, and origins, from ARSEF and CSIRO. The CCCr (0.965, p < 0.0002) indicates that the dendrogram is a good representation of genetic relationships among strains and also that the groups generated preserve the pairwise distance among the original data.
Analysis of ITS sequences. The amplification of the ITS region resulted in a single product for all strains. The size of the product was approximately 600 bp for all strains. The ITS sequence data set consisted of 599-620 aligned positions. From the results of the BI analysis, all Mexican strains from Aeneolamia spp. formed a well-supported clade (Mani 2) as reported by Brunner-Mendoza et al. (2017) (pp = 1); besides the apparent host specificity of this clade, the morphology of the colony, and the dimensions of conidia do not present differences with respect to species of the M. anisopliae complex (Fig. 2). These findings match and support the group integrated by native strains from Mexico generated by RAPD data.
Table 4. Conidia dimensions of Metarhizium strains from Mexico and reference strains.
|
Isolate |
Conidia dimensions (μm) |
|
EH-467* |
7.353 × 2.564 |
|
EH-468* |
7.371 × 2.564 |
|
EH-469* |
7.692 × 2.564 |
|
EH-470* |
7.947 × 2.564 |
|
EH-471* |
7.692 × 2.564 |
|
EH-472* |
7.691 × 2.564 |
|
EH-473* |
7.435 × 2.564 |
|
EH-474* |
7.692 × 2.564 |
|
EH-476* |
7.692 × 2.564 |
|
EH-477* |
7.692 × 2.564 |
|
EH-478* |
7.400 × 2.564 |
|
EH-479* |
7.570 × 2.564 |
|
EH-480* |
7.692 × 2.564 |
|
M. album^ (FI-MaF) |
4 – 6 × 1.5 – 2.5 |
|
M. acridum^ (FI-987) |
4.5 × 2.6 |
|
M. anisopliae^ (FI-1029) |
5 -7 × 2 – 3.5 |
|
M. flavoviride^ (FI-405) |
8.0 – 12.0 × 3.5 – 6.0 |
|
M. novozealandicum^ (FI-698) |
6.0 × 2.4 |
|
M. pemphigi^ (FI-72) |
5.4 × 2.4 |
|
M. lepidiotae^ (FI-147) |
7.3-10.6 × 3-4.1 |
|
M. majus^ (FI-388) |
8.5-14.5 × 2.5-5.0 |
|
M. minus^ ( FI- 403) |
ND |
* = Strains from Mexico; ^ = reference strains; ND = not determined.
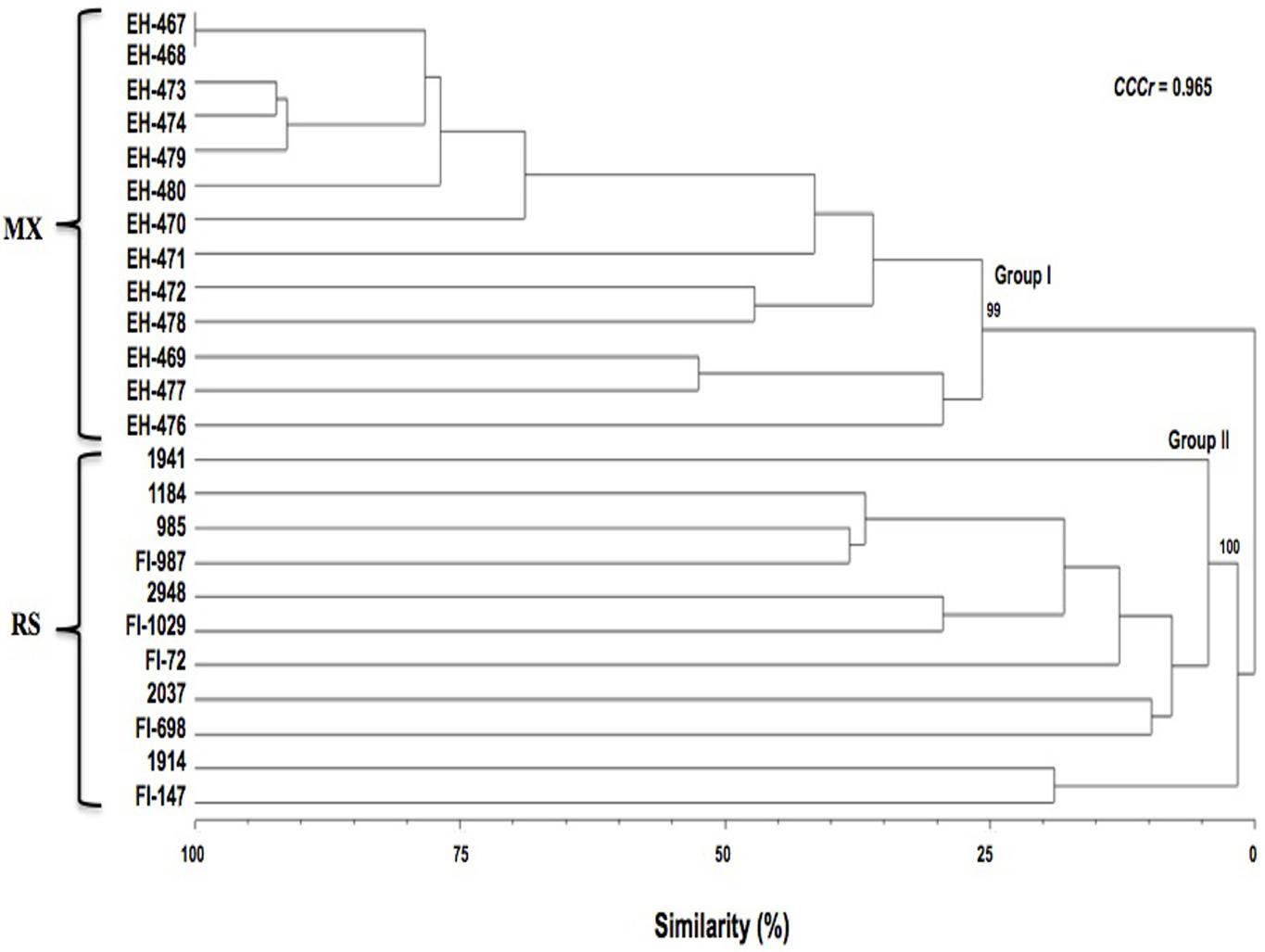
Table 5
Colony diameter (cm) of strains from Mexico and reference strains on SDYA medium.
|
Isolates |
15 °C |
25 °C |
30 ºC |
35 ºC |
|
EH-467* |
2.45 ± 0.086a |
6.75 ± 0.050bcd |
7.62 + 0.109a |
0.83+ 0.044b |
|
EH-468* |
2.53 ± 0.120a |
7.14 + 0.82ab |
7.70 + 0a |
0.70 + 0 0cd |
|
EH-469* |
2.00 ± 0bc |
6.96 ± 1.4abc |
7.20 + 0c |
0.75 + 0.093bc |
|
EH-470* |
1.84 ± 0.054bc |
7.14 ± 0.65ab |
7.40 + 0b |
0.60 + 0d |
|
EH-471* |
2.02 ±0.044bc |
5.27 ± 0.58f |
5.64 + 0.089e |
1.05 + 0a |
|
EH-472* |
0.60 ± 0h |
4.30 ± 0.272g |
5.18 + 0.057f |
0.600 + 0d |
|
EH-473* |
1.70 ± 0cde |
5.99 ± 0.151e |
6.99 + 0.022d |
0.82 + 0.109b |
|
EH-474* |
1.77 ± 0.043bcd |
6.52 ± 0.171d |
7.16 + 0.041c |
0.60 + 0d |
|
EH-476* |
1.34 ± 0.041ef |
6.05 ± 0.086e |
6.95 + 0.11d |
0.60 + 0d |
|
EH-477* |
1.40 ± 0def |
5.44± 0.041f |
6.97 + 0.044d |
0.62 + 0.044d |
|
EH-478* |
1.96 ± 0.310bc |
6.62 ± 0.135cd |
7.18 + 0.044c |
0.60 + 0d |
|
EH-479* |
2.20 ± 0.044ab |
6.56 ± 0.191cd |
7.20 + 0c |
0.79 + 1.29bc |
|
EH-480* |
1.06 ± 0.044fg |
6.87 ± 0.143abc |
4.79 + 0.022h |
0.60 + 0d |
|
1941** |
0.70 ± 0g |
1.30 ± 0g |
1.04 ± 0.065h |
0 ± 0c |
|
1914** |
2.00 ± 0b |
4.67 ± 0.057c |
5.14 ± 0.089b |
0.66 ± 0.019b |
|
1184** |
2.00 ± 0b |
4.58 ± 0.057c |
2.72 ± 0.109e |
0 + 0c |
|
2037** |
0.79 ± 0.022ef |
2.80 ± 0e |
2.70 ± 0.106e |
0 ± 0c |
|
2948** |
0.900 + 0e |
2.80 ±0.339e |
1.57 ± 0.057f |
0 + 0c |
|
985** |
1.28± 0.027d |
4.23 ± 0.044d |
4.18 ± 0.109c |
2.04 + 1.341a |
|
FI-147** |
1.92 ± 0.125bc |
5.32 ± 0.083b |
3.82 ± 0.044d |
0 + 0 c |
|
FI-1029** |
2.80 ± 0.136a |
7.62 ± 0.130a |
6.02 ± 0.044a |
2.180 + 0.658a |
|
FI-698** |
1.88 ± 0.057bc |
1.90 ± 0f |
0.60 ± 0i |
0 + 0 c |
|
FI-987** |
0.900 ± 0e |
4.70 ± 0c |
3.97 ± 0.044d |
0.720 + 0.178b |
|
FI-72** |
1.86 ± 00.740c |
4.50 ± 0.061c |
1.39 ± 0.022g |
0 + 0 c |
*Strains from Mexico (F = 82.088; d.f. = 12; p < 0.001), same letters with no significant differences. **Reference strains (F = 1126.871; d.f. = 10; p < 0.001), same letters with no significant differences. Strains in SDAY at day 14.
Discussion
Conidial morphology has been considered the only potentially useful morphological character to distinguish several Metarhizium species (Driver et al., 2000). Although it is possible to differentiate many species in this genus using morphological features, the molecular approach has shown the existence of cryptic species that are morphologically indistinguishable (Bischoff et al., 2009). Our results showed a range of conidial dimensions in the strains from Mexico corresponding to M. anisopliae (Driver et al., 2000) and to cryptic species reported later by Bischoff et al. (2009), M. anisopliae, M. brunneum, M. lepidiotae, M. pingshaense, and M. robertsii. Although conidial morphology is a useful tool, it is not possible to distinguish among very closely related species within the M. anisopliae complex.
Most of the entomopathogenic fungi are considered mesophilic (optimal growth temperature between 25 and 35 ºC) (Cooney & Emerson, 1964), although there are cold and heat active strains of M. anisopliae reported by Bidochka et al. (2001). But until now, there is no evidence of a strong correlation between conidial heat resistance and latitude of origin (Rangel et al., 2010). It is necessary to evaluate the behavior of native strains to elucidate their situation and provide a better and more integrated field management. Our thermotolerance results showed a limited growth of most of the strains used in this study at 35 ºC, compared to 28 ºC, except for the strains isolated from Orthoptera.
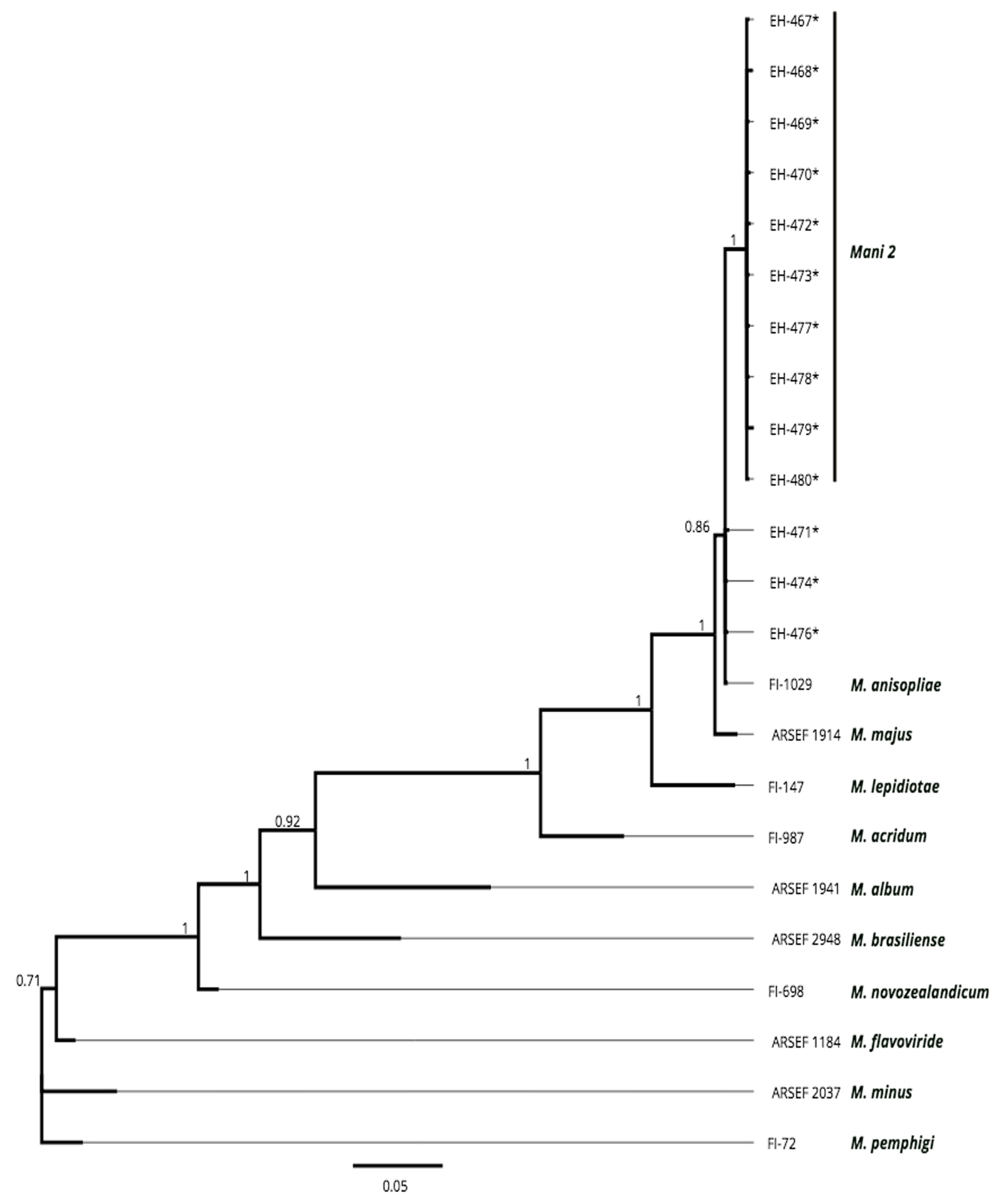
RAPD markers have been widely used for the study of genetic diversity / polymorphism in fungi (Nirmaladevi et al., 2016; Skoneczny et al., 2015). In Metarhizium, this technique was also useful to conclude that M. anisopliae contained a number of cryptic species (Bidochka et al., 1993). Other authors, like Fegan et al. (1993) and Fungaro et al. (1996), demonstrated a possible correlation between the insect host and persistence of particular fungal genotypes in specific locations. Our results showed 2 major clusters: Group I that included all strains from Mexico isolated from Hemiptera in sugar cane fields, and Group II with all reference strains obtained from different hosts and geographic origins. Strains from Mexico formed a cluster with high support values from both RAPD data and ITS sequences analysis, suggesting that the existing populations of M. anisopliae are in a process of divergence and the resulting linages are undergoing a process of speciation, as shown in previous studies with the same strains identified as a subclade (Mani 2) within M. anisopliae (Brunner-Mendoza et al. 2017) revealing a greater number of phylogenetic species. Besides, Fungaro et al. (1996) demonstrated that M. anisopliae strains are extremely diverse but insect strains present a moderate degree of variation, suggesting the development of host specificity. Many factors could influence genetic diversity, such as environmental and anthropic influences (Amos & Harwood, 1998), and climate conditions, thereby enhancing gene exchange and leading to high genetic diversity (Joshi et al., 2000); however, further investigations are required.
The phylogenetic analysis using ITS sequences clustered all strains from Mexico with the sequences of FI-1029 (M. anisopliae) with a high value. This region is still being considered the official DNA barcode marker for fungi (Hawksworth, 1991), despite that in some genera, such as Metarhizium, the lack of informative sites results in poor resolution for some clades (Driver et al., 2000). Nevertheless, it has been useful to identify a broad group of sampled fungi and is one of the most commonly used markers with one of the largest databases (Raja et al., 2017).
These findings show the importance of evaluating strains from different Mexican locations with molecular tools, when these strains have the potential of being used as microbial agents for biological control, because most investigations demonstrate differences related to their behavior as insect pathogens and in physiological parameters. In this study we showed molecular differences between strains from Mexico and other strains of the genus Metarhizum.
Acknowledgments
To the financial support by PAPIIT-DGAPA, UNAM, project IT202012 and Consejo Nacional de Ciencia y Tecnología, project PDCPN 2015 1247. The authors thank the Colección de Hongos Entomopatógenos from the CNRCB, Sagarpa, Senasica for the native Metarhizium anisopliae strains from Mexico and Dr. Richard Humber (ARSEF) for providing the Metarhizium reference strains.
References
Allnut, T. R., Newton, A. C., Lara, A., Premoli, A., Armesto, J. J., Vergara, R. et al. (1999). Genetic variation in Fitzroya cupressoides (alerce), a threatened South American conifer. Molecular Ecology, 8, 975–987.
Amos, W., & Harwood, J. (1998). Factors affecting levels of genetic diversity in natural populations. Philosophical Transactions of the Royal Society of London. Series B, 1366, 177–186.
Bidochka, M. J., Kamp, M. A., Lavendar, M. T., Dekoning, J., & Amritha De Croos, J. N. (2001). Habitat association in two genetic groups of the insect-pathogenic fungus Metarhizium anisopliae: Uncovering cryptic species? Applied Environmental Microbiology, 67, 1335–1342.
Bidochka, M. J., McDonald, M. A., St. Leger, R. A., & Roberts, D. W. (1993). Differentiation of species and strains of entomopathogenic fungi by Random Amplified Polymorphic DNA (RAPD). Current Genetics, 21, 107–113.
Bischoff, J. F., Rehner, S. A., & Humber, R. A. (2009). A multilocus phylogeny of the Metarhizium anisopliae lineage. Mycologia, 101, 512–530.
Brunner-Mendoza, C., Moonjely, S., Reyes-Montes, M. R., Toriello, C., & Bidochka, M. J. (2017). Physiological and phylogenetic variability of Mexican Metarhizium strains. BioControl, 62, 779–791.
Cobb, B. D., & Clarkson, J. M. (1994). A simple procedure for optimizing the Polymerase Chain Reaction (PCR) using modified Taguchi methods. Nucleic Acids Research, 18, 3801–3805.
Cooney, D. G., & Emerson, R. (1964). Thermophilic fungi: an account of their biology, activities and classification. San Francisco and London: Freeman, W.H. and Co.
Cuadros-Orellana, S., Rabelo-Leite, L., Smith, A., Medeiros, J. D., Badotti, F., Fonseca, P. L. C. et al. (2013). Assessment of fungal diversity in the environment using metagenomics: a decade in review. Fungal Genomics & Biology, 3, 100.
Curran, J., Driver, F., Ballard, J. W. O., & Milner, R. J. (1994). Phylogeny of Metarhizium: Analysis of ribosomal DNA sequence data. Mycological Research, 98, 547–552.
Driver, F., Milner, R. J., & Trueman, J. H. (2000). A taxonomic revision of Metarhizium based on a phylogenetic analysis of rDNA sequence data. Mycological Research, 104, 134–150.
Fegan, M., Manners, J. M., Maclean, D. J., Irwin, J. A. G., Samuels, K. D. Z., Holdom, D. G. et al. (1993). Random amplified polymorphic DNA markers reveal a high degree of genetic diversity in the entomopathogenic fungus Metarhizium anisopliae var. anisopliae. Journal of General Microbiology, 13, 2075–2081.
Fernandes, E. K. K., Keyser, C. A., Chong, J. P., Rangel, D. E. N., Miller, M. P., & Roberts, D. W. (2009). Characterization of Metarhizium species and varieties based on molecular analysis, heat tolerance and cold activity. Journal of Applied Microbiology, 108, 115–128.
Fungaro, F. H. P., Vieira, M. L. C., Pizzirani-Kleiner, A. A., & de Azevedo, J. L. (1996). Diversity among soil and insect strains of Metarhizium anisopliae var. anisopliae detected by RAPD. Letters in Applied Microbiology, 2, 389–392.
Goettel, M. S., & Inglis, D. (1997). Fungi: Hyphomycetes. In L. A. Lacey (Ed.), Manual of techniques in insect pathology (pp. 213–248). San Diego: Academic Press.
Hawksworth, D. L. (1991). The fungal dimension of biodiversity: magnitude, significance, and conservation. Mycological Research, 95, 641–655.
Huelsenbeck, J. P., & Ronquist, F. (2001). MrBayes: Bayesian inference of phylogenetic trees. Bioinformatics (Oxford, England), 17, 754–755.
Joshi, S. P., Gupta, V. S., Agarwal, R. K., Ranjekar, P. K., & Brar, D. S. (2000). Genetic diversity and phylogenetic relationship as revealed by inter simple sequence repeat (ISSR) polymorphism in the genus Oryza. Theoretical and Applied Genetics, 100, 1311–1320.
Kepler, R. M., Humber, R. A., Bischoff, J. F., & Rehner, S. A. (2014). Clarification of generic and species boundaries for Metarhizium and related fungi through multigene phylogenetics. Mycologia, 106, 811–829.
Kumari, N., & Thakur, K. S. (2014). Randomly amplified polymorphic DNA-A brief review. American Journal of Animal and Veterinary Sciences, 9, 6–13.
Nirmaladevi, D., Venkataramana, M., Srivastava, R. K., Uppalapati, S. R., Gupta, V. K., Yli-Mattila, T. et al. (2016). Molecular phylogeny, pathogenicity and toxigenicity of Fusarium oxysporum F. Sp. Lycopersici. Scientific Reports, 6, 21367.
Paelowska, T. E., & Taylor, J. W. (2004). Organization of genetic variation in individuals of arbuscular mycorrhizal fungi. Nature, 427, 733–737.
Raja, H. A., Miller, A. N., Pearce, C. J., & Oberlies, N. H. (2017). Fungal identification using molecular tools: A primer for the natural products research community. Journal of Natural Products, 80, 756–770.
Rangel, D. E. N., Fernandes, K. K., Dettenmaier, S. J., & Roberts, D. W. (2010). Thermotolerance of germlings and mycelium of the insect-pathogenic fungus Metarhizium spp. and mycelial recovery after heat stress. Journal of Basic Microbiology, 50, 344–350.
Real, R., & Vargas, J. M. (1996). The probabilistic basis of Jaccard’s index of similarity. Systematic Biology, 45, 380–385.
Rezende, J. M., Riguetti-Zanardo, A. B., Lopes, M. S., Delalibera, I., & Rehner, S. A. (2015). Phylogenetic diversity of Brazilian Metarhizium associated with sugarcane agriculture. BioControl, 60, 495–505.
Rholf, F. J. (1988). NTSYS-pc. Numerical taxonomy and multivariate analysis system version 2.02e. Exeter Sofware. New York.
Schoch, C. L., Seifert, K. A., Huhndorf, S., Robert, V., Spouge, J. L., Levesque, C. A. et al. (2012). Nuclear ribosomal internal transcribed spacer (ITS) region as universal DNA barcode marker for fungi. Proceedings of the National Academy Sciences USA, 109, 6241–6246.
Senasica (Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria). (2017). Programas de trabajo. Retrieved on May 16th, from https://www.gob.mx/senasica/documentos/programas-de-trabajo-sanidad-vegetal
Skoneczny, D., Oskiera, M., Szczech, M., & Bartoszewski, G. (2015). Genetic diversity of Trichoderma atroviride strains collected in Poland and identification of loci useful in detection of within-species diversity. Folia Microbiologica, 60, 297–307.
Sneath, P. H. A., & Sokal, R. R. (1973). Taxonomic structure. In W. H. Freeman (Ed.), Numerical taxonomy (pp.188–305). San Francisco: Freeman, W.H. and Co.


