
Ricard Arasa-Gisbert a, *, Víctor Arroyo-Rodríguez a, b, Juan Javier Ortiz-Díaz c y Esteban Martínez d
a Universidad Nacional Autónoma de México, Instituto de Investigaciones en Ecosistemas y Sustentabilidad, Antigua Carretera a Pátzcuaro Núm. 8701, Ex-Hacienda de San José de la Huerta, 58190 Morelia, Michoacán, México
b Universidad Nacional Autónoma de México, Escuela Nacional de Estudios Superiores, Tablaje Catastral Núm. 6998, Carretera Mérida-Tetiz Km. 4.5, 97357 Mérida, Yucatán, México
c Universidad Autónoma de Yucatán, Departamento de Botánica, Campus de Ciencias Biológicas y Agropecuarias, Carretera Mérida-Xmatkuil Km. 15.5, 97000 Mérida, Yucatán, México
d Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Botánica, Herbario Nacional de México, 04510 Ciudad de México, México
*Autor para correspondencia: r_arasa_gisbert@hotmail.com (R. Arasa-Gisbert)
Recibido: 2 de mayo 2020; aceptado: 28 octubre 2020
Resumen
Este estudio presenta la lista florística de árboles, arbustos y palmas en regeneración muestreados en 60 fragmentos de bosque tropical húmedo en 3 regiones distintas (n = 20 por región): Marqués de Comillas (Chiapas), Los Tuxtlas (Veracruz) y región norte de Chiapas. En cada fragmento registramos las plantas leñosas en regeneración presentes en 25 parcelas circulares de 8 m2 (200 m2 por fragmento). En total, registramos 24,612 individuos pertenecientes a 431 especies, 220 géneros y 70 familias, incluyendo 3 registros nuevos, 2 estatales (Piper phytolaccifolium Opiz y Coccoloba acuminata Kunth) y 1 nacional (Piper philodendroides Standl. et Steyerm.). La densidad media de especies fue significativamente más alta en el paisaje mejor conservado (Marqués de Comillas) que en el más degradado (norte de Chiapas); Los Tuxtlas presentó un valor intermedio. De acuerdo con la estrategia de regeneración, 43% de las especies fueron generalistas (especies de sucesión intermedia), 31% tolerantes a la sombra y 13% pioneras (sucesión temprana). Todas las regiones fueron dominadas por pocas especies; la mayoría fueron raras. El porcentaje de especies amenazadas no difirió entre regiones (5.5-6%). Este estudio destaca el potencial regenerativo de los fragmentos de selva y su elevado valor de conservación frente a la creciente deforestación y degradación que amenazan su persistencia.
Palabras clave: Brinzal; Composición florística; Conservación; Diversidad; Regeneración de avanzada; Riqueza taxonómica; Selva alta perennifolia
© 2021 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Regeneration of woody plants in tropical rainforest patches: community structure and new records for Chiapas, Tabasco and Mexico
Abstract
This study presents the floristic list of regenerating trees, shrubs and palms sampled in 60 tropical rainforests fragments in 3 different regions (n = 20 per region): Marqués de Comillas (Chiapas), Los Tuxtlas (Veracruz) and northern region of Chiapas. In each fragment, we recorded saplings in 25 8 m2-circular plots (200 m2 per fragment). In total, we recorded 24,612 individuals belonging to 431 species, 220 genera and 70 families, including 3 new records, 2 at the state level (Piper phytolaccifolium Opiz and Coccoloba acuminata Kunth) and 1 at the national level (Piper philodendroides Standl. et Steyerm.). Mean species density was significantly higher in the best-preserved landscape (Marqués de Comillas) than in the most degraded one (northern Chiapas), and Los Tuxtlas presented an intermediate value. According to the regeneration strategy, 43% of the species were generalists (intermediate succession species), 31% shade tolerant and 13% pioneers (early succession). All regions were dominated by few species; most were rare. The percentage of threatened species did not differ between regions (5.5-6%). This study highlights the regenerative potential of forest fragments and their high conservation value in the face of increasing deforestation and degradation that threaten their persistence.
Keywords: Sapling; Floristic composition; Conservation; Diversity; Advance regeneration; Taxonomic richness; Tropical rainforests
© 2021 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introducción
Los bosques tropicales son el bioma más diverso del planeta y proporcionan servicios ecosistémicos fundamentales para las sociedades humanas (Balvanera et al., 2006; Slik et al., 2015). Sin embargo, el aumento de las poblaciones humanas y de sus demandas de recursos naturales ha provocado un incremento de la deforestación en las regiones tropicales (Hansen et al., 2020; Song et al., 2018). Como resultado, grandes extensiones de bosque tropical continuo han sido convertidas en paisajes fragmentados compuestos, en su mayoría, por pequeños fragmentos de bosque remanente (Taubert et al., 2018). En muchos casos, estos fragmentos se encuentran aislados y rodeados de una matriz no arbolada (e.g., pastizales y campos agrícolas). Esto dificulta el movimiento de muchos animales en el paisaje, limitando la dispersión de semillas y la regeneración natural de los bosques (Arroyo-Rodríguez et al., 2017; Peña-Domene et al., 2018; San José et al., 2020). Sorprendentemente, la mayoría de estudios en paisajes fragmentados se centran en árboles adultos. Muy pocos prestan atención a los individuos en estado inmaduro que han logrado germinar y establecerse (i.e., regeneración de avanzada), a pesar de su papel clave en el ecosistema, ya que determinan la composición y estructura futura de los bosques (Arroyo-Rodríguez et al., 2017; Chazdon y Guariguata, 2016).
En los últimos 30 años, México ha perdido 34% de la cobertura de bosques tropicales, amenazando la supervivencia de un gran número de especies de plantas y animales (Conabio y Semarnat, 2009; Inegi, 2014). Chiapas y Veracruz son el segundo y tercer estado con mayor diversidad de plantas de México pero son también los estados que han sufrido mayor deforestación (Villaseñor, 2016). En la mayor selva tropical húmeda de México, la selva Lacandona en Chiapas, se perdió 31% de la superficie forestal en tan solo 40 años (De Jong et al., 2000). Una situación más alarmante la vive la Reserva de la Biosfera de Los Tuxtlas, en Veracruz, donde hoy queda poco más de 15% de las selvas de tierras bajas (Castillo-Campos y Laborde, 2004; Conanp, 2011).
Aunque las plantas son uno de los grupos taxonómicos más estudiados, son pocos los estudios florísticos y de ecología que analizan la composición y pérdida de la flora nativa (Cayuela et al., 2012; Fazey et al., 2005). Este hecho es especialmente relevante en las regiones tropicales, donde muchas especies no han sido aún descritas y muchas otras carecen de información relativa a su distribución, abundancia e interacciones y dinámicas ecológicas (Collen et al., 2008; Hubbell, 2013; Joppa et al., 2011; Prance et al., 2000). En este sentido, contar con información ecológica como la estrategia de regeneración es clave para predecir la estructura, composición y dinámica futura de los bosques ubicados en paisajes modificados por el hombre (Arroyo-Rodríguez et al., 2017; Bugmann, 1996; Larson y Funk, 2016).
El objetivo de este trabajo es brindar información acerca de la estructura y composición de la comunidad de plantas leñosas en regeneración en fragmentos de bosque tropical húmedo de 3 regiones con diferente patrón e historia de deforestación del sureste de México: Marqués de Comillas (Chiapas), Los Tuxtlas (Veracruz) y región norte de Chiapas. El fin último es mejorar el conocimiento de la flora tropical mexicana, especialmente de la regeneración de avanzada, para resaltar el potencial regenerativo y la enorme diversidad que albergan los fragmentos de bosque. Esta información puede ser útil para promover acciones de investigación, conservación y manejo en paisajes tropicales fragmentados (Arroyo-Rodríguez et al., 2020).
Materiales y métodos
Las 3 regiones de estudio muestran el mismo tipo de vegetación nativa (bosque tropical húmedo) pero difieren en el grado de deforestación e historia de cambio de uso del suelo (fig. 1). La mejor conservada es la región de Marqués de Comillas, que forma parte de la selva Lacandona, Chiapas (91°6’-90°41’ O, 16°19’-16°2’ N). Este bosque tropical húmedo es el más importante de México, tanto por su tamaño (~ 13,000 km2) como por su riqueza florística, ya que alberga más de 50% de las especies propias de este tipo de vegetación del país (Romaní-Cortés, 2011; Villaseñor et al., 2018). Sin embargo, un programa gubernamental de colonización de tierras a finales de los años 70 provocó la pérdida de más de la mitad de los bosques primarios de la región (Carabias et al., 2015). Como consecuencia, el municipio de Marqués de Comillas está hoy dominado por fragmentos de bosque primario de diferente tamaño, los cuales cubren cerca de 45% de la superficie del municipio (Carabias et al., 2015). Sin embargo, la presencia de fragmentos de bosque de gran tamaño (> 1,000 ha) ha permitido la persistencia de especies de animales amenazados como el jaguar, tapir, mono araña, mono aullador, pecarí y guacamaya roja (Arce-Peña et al., 2019; Carrara et al., 2015; Garmendia et al., 2013).
La región de Los Tuxtlas, en Veracruz (95°24’-94°96’ O, 18°68’-18°38’ N) preserva los bosques tropicales húmedos más septentrionales de América y albergan ~ 40% de las especies de plantas vasculares tropicales de México (Dirzo y García, 1992; Villaseñor et al., 2018). Sin embargo, la intensa deforestación provocada por la ganadería extensiva durante la década de los 50’s ha provocado que actualmente solo 20% del territorio esté cubierto por selvas (Castillo-Campos y Laborde, 2004; Conanp, 2011). La mayoría de fragmentos de la región son pequeños (< 8 ha) y se encuentran inmersos en una matriz de potreros y campos de cultivo (Arroyo-Rodríguez et al., 2008; Guevara y Laborde, 2012). Los fragmentos remanentes presentan poblaciones muy reducidas o ausentes de vertebrados medianos y grandes (Dirzo y Miranda, 1991; González-Christen y Coates, 2019), aunque algunos dispersores de semillas (e.g., monos aulladores) aún están presentes en algunos fragmentos (Arroyo-Rodríguez et al., 2008; Galán-Acedo et al., 2018).
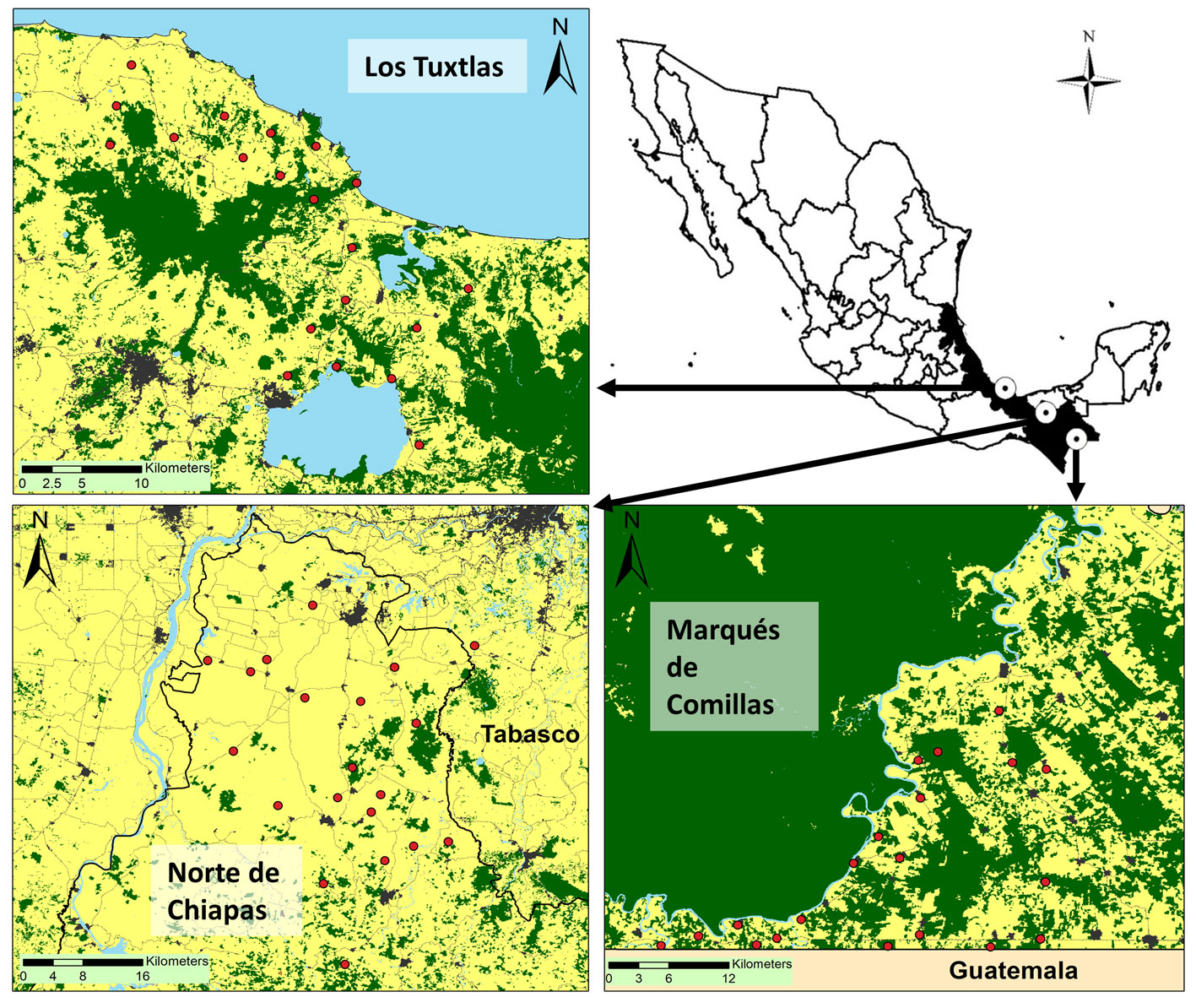
La región de estudio que está más deforestada es la región norte de Chiapas (93°33’-93°00’ O, 17°87’-17°42’ N). Debido a la escasez de fragmentos de bosque primario en la región, uno de los 20 fragmentos de estudio está ubicado fuera del estado de Chiapas, en Tabasco (fig. 1). La introducción del ganado en la década de los 50 y las prospecciones petrolíferas de los 70 provocaron una intensa deforestación en la región (INEGI, 1999). Actualmente, tan solo 5% del territorio es ocupado por bosques primarios y los pocos fragmentos remanentes son extremadamente pequeños (< 5 ha) y se encuentran rodeados por una matriz de pastizales inducidos y carreteras (Anzures-Dadda y Manson, 2007; Renaissance Oil Corp., 2017). Debido a esto, una gran parte de los vertebrados de tamaño mediano y grande (e.g., jaguar, tapir, mono araña) se han extinguido localmente y muchos otros (e.g., jaguarundi, pecarí, venado de cola blanca) están muy amenazados y es posible que se extingan en los próximos años (Renaissance Oil Corp., 2017). No obstante, aún persisten algunas poblaciones de monos aulladores y de otros dispersores de gran importancia, como aves y murciélagos (Galán-Acedo et al., 2018; Renaissance Oil Corp., 2017).
Según la clasificación climática de Köppen, las 3 regiones quedan encuadradas dentro del clima tropical ecuatorial, aunque presentan variaciones pluviométricas: en Los Tuxtlas se superan los 4,000 mm al año, mientras que en el norte de Chiapas oscila entre los 2,500 y 4,000 mm, y en Marqués de Comillas llueve unos 3,000 mm (Breugel et al., 2006; Gobierno de Chiapas, 2013). A pesar de presentar un régimen de lluvias constantes durante todo el año, las 3 regiones presentan una marcada estación seca; ésta se presenta entre marzo y mayo en Los Tuxtlas (Instituto de Biología: http://www.ibiologia.unam.mx/tuxtlas/localizacion/generalidades/frame.htm) y de febrero a abril en las otras 2 regiones (Breugel et al., 2006; Gobierno de Chiapas, 2013).
El muestreo de campo se llevó a cabo de enero a mayo de 2018. En cada región se seleccionaron arbitrariamente 20 fragmentos de bosque primario que cumplieran los siguientes criterios: 1) estar en un rango de tamaño de 1 a 150 ha (a excepción de un fragmento de 640 ha en Los Tuxtlas, y otro de 3,410 ha en Marqués de Comillas); 2) presentar una distancia mínima de 1.5 km con los otros fragmentos de estudio (para poder llevar a cabo, más adelante, estudios a escala de paisaje); 3) sin signos evidentes de perturbación antrópica —árboles talados, senderos humanos, etc.— y 4) estar a una misma elevación (< 600 m snm) y presentar un tipo de suelo similar (fig. 1). En el centro de cada fragmento se establecieron 25 parcelas circulares de 8 m2 cada una (200 m2 por fragmento), formando una cuadrícula de 5 × 5 parcelas, con una separación de 30 m entre sí. En cada parcela se contaron e identificaron todas las plantas leñosas — árboles y arbustos — y palmas en regeneración (i.e., ≥ 30 cm de altura y < 1 cm de diámetro a la altura del pecho). Cabe destacar que la gran mayoría de las especies de palmas muestreadas alcanzan la madurez con tamaños mayores al considerado aquí (Burelo-Ramos et al., 2009; Oyama, 1987; Piñero et al., 1977; Quero y Pérez-Farrera, 2010; Standley y Steyermark, 1952). Aunque algunas especies como Chamaedorea elegans y C. ernesti-augusti pueden presentar individuos adultos con tamaños dentro del rango seleccionado aquí, nosotros excluimos todos los individuos con estructuras reproductivas o con signos de haberlas tenido para enfocarnos en las palmas juveniles. De hecho, tras esta exclusión, solo registramos un individuo de C. elegans y 10 de C. ernesti-augusti. El mismo protocolo fue utilizado con los arbustos de las especies que pudiesen presentar individuos adultos con tamaños dentro del rango considerado aquí.
Los individuos no identificados en campo fueron colectados para su posterior identificación en el Herbario Nacional de México (MEXU, Ciudad de México). Usamos la nomenclatura de Tropicos (Jardín Botánico de Missouri; http://www.tropicos.org). Una vez identificadas, se clasificaron todas las especies con base en la estrategia de regeneración siguiendo la clasificación de Hill y Curran (2003), Arroyo-Rodríguez y Mandujano (2006) y Chazdon (2014): especies pioneras (o especies sucesionales tempranas), especies generalistas (o especies sucesionales intermedias) y especies tolerantes a la sombra. También se incluyeron a las especies introducidas como un grupo independiente. Las plantas pioneras son especies de rápido crecimiento y vida corta, que típicamente producen una gran cantidad de semillas, y germinan y se establecen únicamente en áreas abiertas (i.e., claros dentro del bosque y bordes forestales; Martínez-Ramos, 1994). En contraste, las especies tolerantes a la sombra son de lento crecimiento y vida más larga, que usualmente producen semillas de mayor tamaño que las plantas pioneras, con mayor reserva y que, por tanto, tienen adaptaciones para germinar y establecerse en áreas cerradas, con muy poca luz (Martínez-Ramos, 1994). En un punto intermedio se encuentran las especies generalistas, que pueden germinar en sitios sombríos y soleados, con un crecimiento relativamente rápido, especialmente en sitios soleados, y con una esperanza de vida mayor a la de las plantas pioneras (por esta razón, también son llamadas especies de sucesión intermedia; Chazdon, 2014; Martínez-Ramos, 1994; Poorter et al., 2004). La clasificación se basó en diversas fuentes, incluyendo libros, listas de especies, artículos y
floras (apéndice).
Para conocer la estructura de la comunidad de plantas leñosas en regeneración, realizamos curvas de rango-abundancia en cada región de estudio. Para cuantificar el grado de dominancia en cada región, calculamos el factor de equidad (EF, por sus siglas en inglés) propuesto por Jost (2010). Este índice representa la proporción de especies dominantes en la comunidad. El índice es 1 cuando la comunidad es totalmente equitativa y se aproxima a 0 cuando incrementa la dominancia (i.e., pocas especies dominan, por lo que la mayoría de las especies son raras). Para calcular el índice, consideramos el número efectivo de especies dominantes (2D) y el número total de especies (0D). El factor de equidad se calcula como: EF = 2D /0D (ver fórmulas descritas por Jost, 2006) y para el cálculo de 0D y 2D, utilizamos la paquetería Entropart (Marcon y Herault, 2015) presente en el software de R 3.4.3 (R Core Team, 2013).
Para evaluar cómo varía la composición de especies entre fragmentos y entre regiones se llevó a cabo un escalamiento multidimensional no métrico (NMDS, por sus siglas en inglés). Este análisis multivariado permite ordenar los fragmentos en un espacio de 2 dimensiones en función de su disimilitud composicional (índice de Bray-Curtis). Para evaluar si la composición florística difirió significativamente entre regiones, se hizo un análisis permutacional de la varianza (Permanova) con la paquetería PairwiseAdonis (Arbizu, 2020).
Además, se calculó la densidad de especies (número de especies/200 m2) en cada sitio y el número total de especies (diversidad gamma) para cada región y tipo de regeneración. Para evaluar si la densidad de especies difirió entre regiones y estrategia de regeneración, se aplicó un modelo lineal generalizado (GLMs) con error Poisson, considerando los efectos aislados y el combinado (interacción) de estos 2 factores categóricos. El modelo no presentó sobredispersión. Por último, para la elaboración del mapa de distribución de los nuevos registros, se usaron las localizaciones de los especímenes almacenados en herbarios, cuyas coordenadas están disponibles en la web del Jardín Botánico de Missouri (www.tropicos.org), así como las nuevas localizaciones reportadas en este trabajo.
Resultados
En total, registramos 24,612 individuos pertenecientes a 431 especies, 220 géneros y 70 familias (apéndice). Por regiones, Los Tuxtlas presentó mayor diversidad gamma (265 especies), seguida de Marqués de Comillas (217 especies) y el norte de Chiapas (188 especies). La mayoría de las especies (185 especies; 43%) fueron clasificadas como generalistas, 134 especies (31.2%) como tolerantes a la sombra, 55 (12.8%) como pioneras, 7 (1.6%) como introducidas y 48 especies (11.2%) no pudieron ser clasificadas (12 de ellas por falta de datos de la especie y las 36 restantes porque no se pudo determinar la especie). De las 36 morfoespecies, 17 fueron identificadas a nivel de género y 19 quedaron como morfoespecies.
Las familias más ricas en especies fueron muy similares en todas las regiones, siendo las familias Fabaceae y Rubiaceae las más representativas (tabla 1). Sin embargo, las familias, los géneros y las especies más abundantes difirieron entre las 3 regiones, es decir, cada región estuvo dominada por distintos taxones (tabla 2). De hecho, a nivel de especie, la composición florística difirió significativamente entre regiones (fig. 2, Permanova, F = 8.37, p < 0.001), siendo la región de Los Tuxtlas la que presentó una composición florística más distinta a las otras 2. Los Tuxtlas también fue la región con una mayor variación en la composición florística entre sus fragmentos, seguida del norte de Chiapas y de Marqués de Comillas (fig. 2).
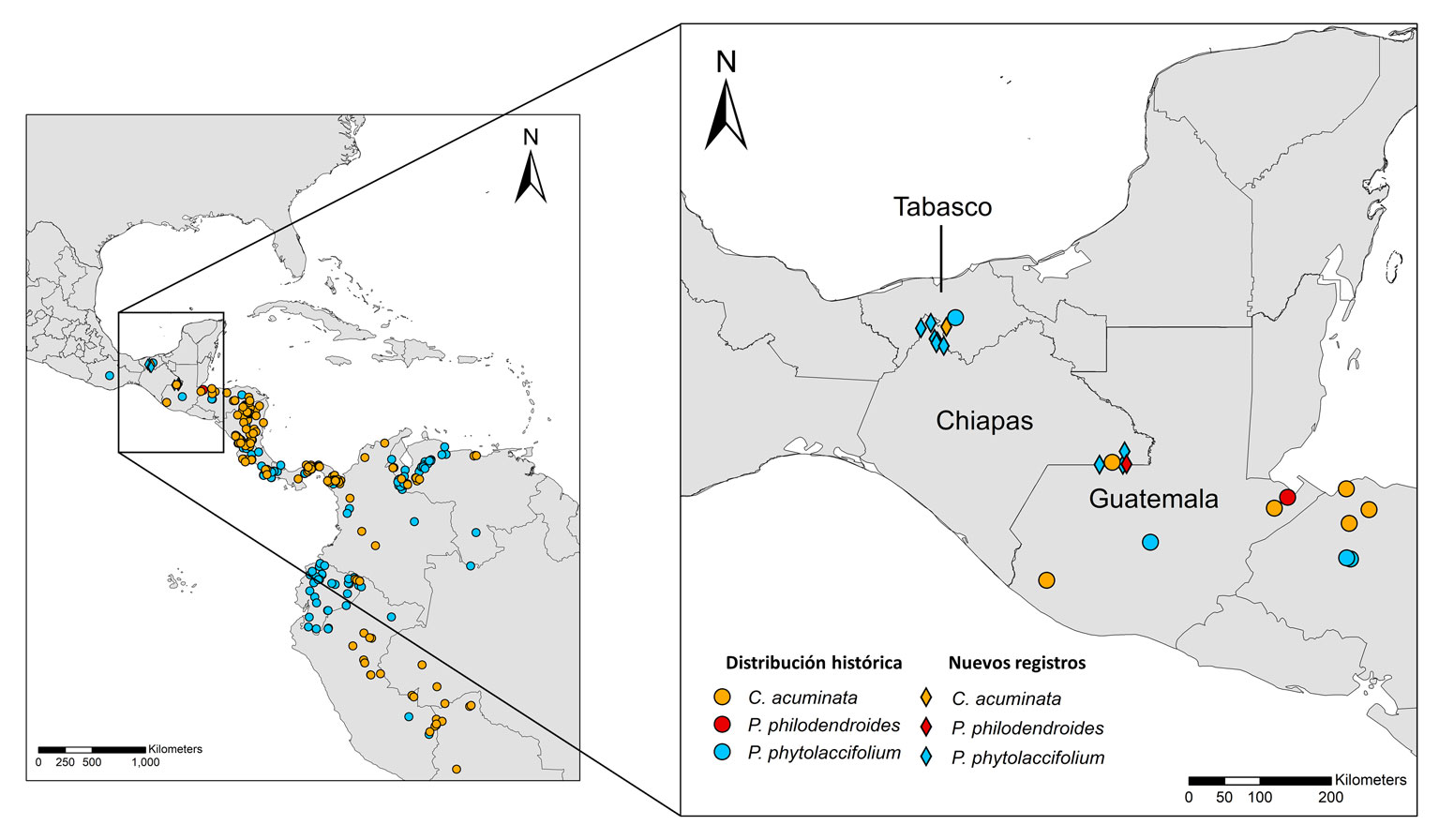
Según la Norma Oficial Mexicana NOM-059-Semarnat-2010, 22 especies encontradas (5% del total, 6% en cada región) cuentan con algún tipo de protección. Cuatro de ellas están sujetas a protección especial, 14 se encuentran como amenazadas y Diospyros conzattii, Ormosia isthmensis, Vatairea lundellii y Mortoniodendron guatemalense se encuentran en peligro (apéndice). Además, reportamos 3 registros nuevos: Piper phytolaccifolium como registro nuevo de Chiapas, Coccoloba acuminata como nuevo de Tabasco y Piper philodendroides como primer registro para México (fig. 3, tabla 3).
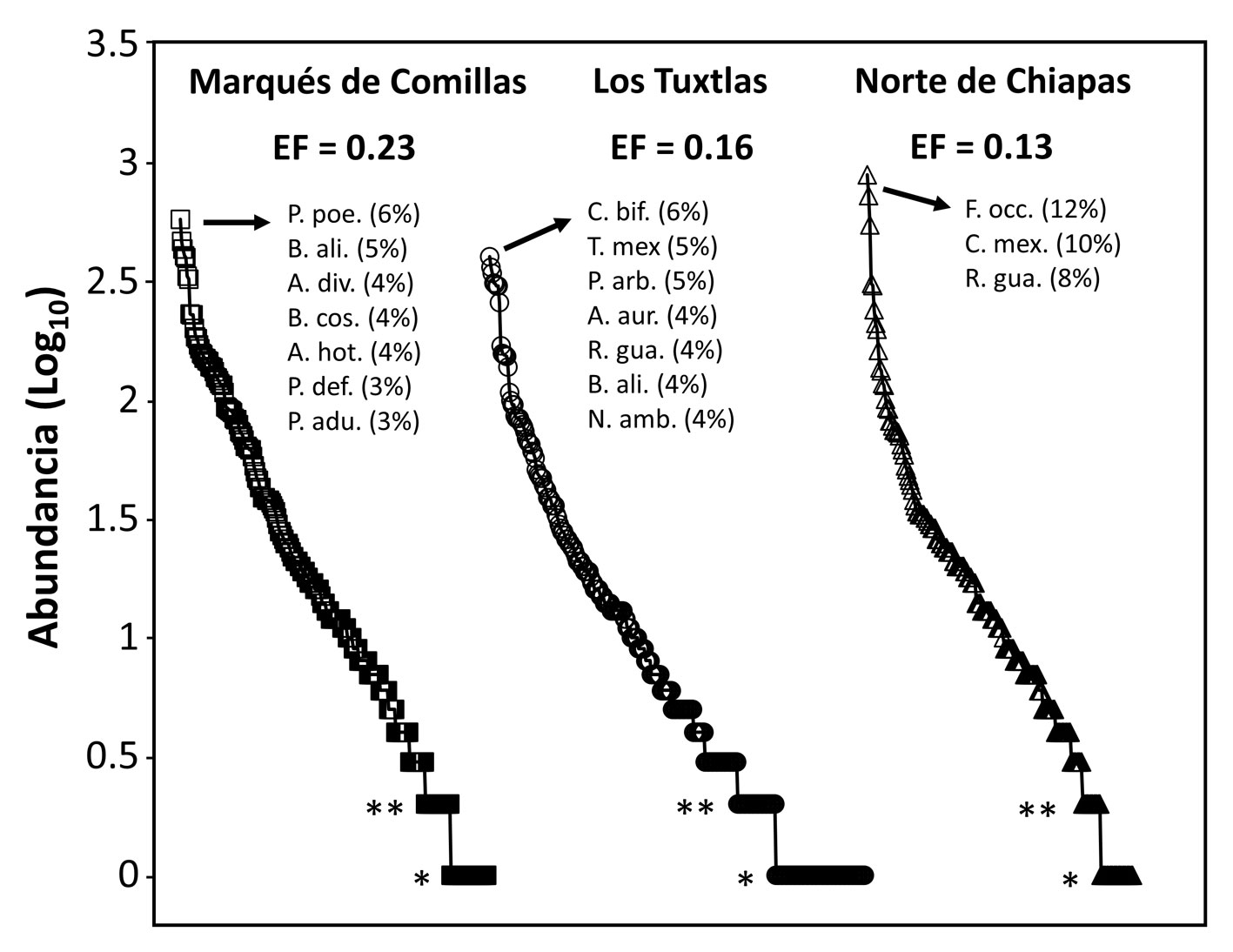
La dominancia fue alta en todas las regiones, ya que 4.8-8.8% de las especies representaron 50% de los individuos en cada región (fig. 4). El norte de Chiapas fue la región con mayor dominancia (EF = 0.13), seguida de Los Tuxtlas (EF = 0.16) y Marqués de Comillas (EF = 0.23). En cuanto a la rareza, Los Tuxtlas fue la región con mayor proporción de especies representadas por un solo individuo (singletons) (23.8% de las especies) o con solo 2 individuos por muestra (doubletons) (10.2%), seguida de Marqués de Comillas (12% singletons y 8.3% doubletons) y el norte de Chiapas (12.2% singletons y 7% doubletons) (fig. 4).
La densidad de especies difirió significativamente entre estrategias de regeneración (χ2 = 9.5, p < 0.001) y entre regiones (χ2 = 53.0, p < 0.001). La interacción entre estos 2 factores fue significativa (χ2 = 27.5, p < 0.001), lo que indica que las diferencias en densidad de especies entre estrategias de regeneración dependen de la región (fig. 5). En particular, la densidad media (± DE) de especies fue 500% mayor en el grupo de generalistas (309 ± 17.4 especies) que en el de pioneras (62 ± 4.6 especies), y 40% mayor que en el de tolerantes a la sombra (221 ± 19.4 especies) (fig. 5). En cuanto a las regiones, Marqués de Comillas presentó la mayor densidad media de especies (71.1 ± 11.9 especies), 62% más alta que en el norte de Chiapas (43.9 ± 8.2 especies) y 25% mayor que en Los Tuxtlas (56.9 ± 13.1 especies). Sin embargo, como indica la interacción significativa entre factores, aunque la densidad de especies generalistas y tolerantes a la sombra difirió entre regiones, la densidad de especies pioneras fue similar en las 3 regiones (fig. 5).

Tabla 1
Las 10 familias con más especies en cada una de las regiones de estudio y en total considerando todas las regiones. Abreviaturas: #S: número de especies por familia; Melastomataceae: Melast.; Moraceae: Mor.; Rutaceae: Rut.; Lauraceae: Lau.; Sapotaceae: Sap.
|
Marqués de Comillas |
Los Tuxtlas |
Norte de Chiapas |
Total |
||||
|
Familia |
#S |
Familia |
#S |
Familia |
#S |
Familia |
#S |
|
Fabaceae |
25 |
Rubiaceae |
25 |
Fabaceae |
22 |
Fabaceae |
44 |
|
Rubiaceae |
24 |
Fabaceae |
24 |
Rubiaceae |
18 |
Rubiaceae |
43 |
|
Melast. |
12 |
Lauraceae |
19 |
Piperaceae |
12 |
Lauraceae |
21 |
|
Arecaceae |
10 |
Euphorbiaceae |
14 |
Malvaceae |
10 |
Piperaceae |
20 |
|
Piperaceae |
9 |
Piperaceae |
13 |
Arecaceae |
8 |
Euphorbiaceae |
18 |
|
Moraceae |
8 |
Sapotaceae |
12 |
Meliaceae |
8 |
Arecaceae |
16 |
|
Malvaceae |
7 |
Arecaceae |
10 |
Moraceae |
8 |
Melast. |
14 |
|
Sapotaceae |
7 |
Myrtaceae |
9 |
Myrtaceae |
7 |
Myrtaceae |
14 |
|
Euphorbiaceae |
6 |
Malvaceae |
8 |
Melast. |
6 |
Sapotaceae |
14 |
|
Myrtaceae |
6 |
Mor. /Rut. |
7 |
Lau. /Sap. |
5 |
Malvaceae |
13 |

Discusión
El presente estudio destaca el potencial regenerativo y gran valor de conservación que tienen los fragmentos de bosque tropical en 3 regiones con diferente grado de perturbación del sureste de México. En tan solo 1.2 ha de muestreo repartidas en 60 fragmentos, encontramos 431 especies de plantas leñosas en regeneración, incluyendo especies amenazadas y 3 nuevos registros, 2 para los estados de Chiapas y Tabasco y uno para México. La dominancia y rareza fue similar en todas las regiones, pero la densidad media de especies difirió significativamente entre regiones, siendo mayor en la región más conservada (Marqués de Comillas) que en la más deforestada (norte de Chiapas). Las regiones de Los Tuxtlas y Marqués de Comillas presentaron una mayor densidad de especies tolerantes a la sombra que la región del norte de Chiapas. Sin embargo, la gran diversidad gamma en las 3 regiones indica que los fragmentos estudiados aún mantienen, en su conjunto, una considerable proporción de la flora de cada región, por lo que pueden actuar como importantes reservorios de biodiversidad.
La regeneración de avanzada en los fragmentos de estudio contiene una porción significativa de la flora vascular de los bosques tropicales húmedos de México. Por ejemplo, las especies muestreadas representan 16-20% de las especies de árboles, arbustos y palmas documentadas para el trópico mexicano (Villaseñor et al., 2003; Villaseñor y Ortiz, 2014). De hecho, la diversidad gamma que encontramos en Los Tuxtlas (265 especies), equivale al ~ 80% de los 333 árboles, arbustos y palmas reportados para la Estación de Biología Tropical de Los Tuxtlas (Ibarra-Manríquez y Sinaca-Colín, 1995), mientras que las 217 especies muestreadas en Marqués de Comillas representan ~ 55% de las 392 especies leñosas reportadas para la región (Martínez et al., 1994). Es importante destacar que la diversidad de estos fragmentos incluye muchas especies amenazadas y nuevos registros a nivel estatal y nacional. Este hallazgo es consistente con inventarios y expediciones florísticas recientes en otros paisajes tropicales fragmentados (León y Martínez-Gordillo, 2008; Solano-Gómez et al., 2011; Vázquez-García, Gómez-Domínguez et al., 2013; Vázquez-García, Pérez-Farrera et al., 2013). Otras investigaciones en las regiones de estudio también encuentran que los fragmentos de bosque pueden ser muy ricos en especies de árboles, incluyendo especies amenazadas y nuevos registros para México (Arroyo-Rodríguez y Mandujano, 2006; Arroyo-Rodríguez et al., 2009; Hernández-Ruedas et al., 2014), lo que destaca su alto valor para salvaguardar la biodiversidad de plantas en paisajes tropicales fragmentados.
A pesar de lo anterior, la deforestación a nivel regional parece limitar el potencial regenerativo de los remanentes de bosque. Aunque otros atributos regionales, como la historia biogeográfica, historia de uso, tipo de suelo, densidad humana (e.g., pisoteo, extracción de leña y árboles, apertura de claros y caminos), presencia de ganado (e.g., ramoneo, pisoteo) o clima, podrían afectar la densidad de especies (Arroyo-Rodríguez et al., 2017), la menor densidad de especies en regiones más deforestadas sugiere que, como ha sido documentado en bosques tropicales de África (Cordeiro y Howe, 2001) y Asia (Stride et al., 2018), la deforestación limita la densidad de especies dentro de los fragmentos. Este patrón fue particularmente evidente al considerar las especies tolerantes a la sombra, sugiriendo que la deforestación tiene un mayor impacto negativo sobre este grupo de especies. De hecho, estudios con árboles (Santos et al., 2008), aves (Carrara et al., 2015; Morante-Filho et al., 2015), mamíferos (Arce-Peña et al., 2019) y anfibios y reptiles (Russildi et al., 2016) han mostrado una tendencia similar: la deforestación tiene un impacto negativo mayor sobre las especies de bosque maduro. No obstante, se requieren estudios adicionales donde se pueda controlar el efecto de otros atributos regionales —e.g., clima, historia de uso— para entender mejor el efecto de la deforestación regional per se sobre el potencial regenerativo de los fragmentos.
Tabla 2
Las 10 familias, 10 géneros y 10 especies más abundantes en cada una de las regiones, y en total, considerando todas las regiones (N = número de individuos).
|
Familias |
N |
Géneros |
N |
Especies |
N |
|
Marqués de Comillas |
|||||
|
Rubiaceae |
2,041 |
Brosimum |
1,026 |
Psychotria poeppigiana |
579 |
|
Moraceae |
1,325 |
Psychotria |
794 |
Brosimum alicastrum* |
468 |
|
Fabaceae |
676 |
Piper |
661 |
Acalypha diversifolia |
434 |
|
Piperaceae |
661 |
Palicourea |
565 |
Brosimum costaricanum* |
407 |
|
Euphorbiaceae |
655 |
Acalypha |
434 |
Ampelocera hottlei* |
400 |
|
Chrysobalanaceae |
451 |
Ampelocera |
400 |
Palicourea deflexa |
334 |
|
Ulmaceae |
400 |
Protium |
304 |
Piper aduncum |
324 |
|
Lauraceae |
367 |
Nectandra |
284 |
Palicourea brachiata |
231 |
|
Burseraceae |
323 |
Licania |
251 |
Calophyllum brasiliense* |
229 |
|
Violaceae |
295 |
Rinorea |
241 |
Nectandra salicifolia* |
429 |
|
Los Tuxtlas |
|||||
|
Moraceae |
1,242 |
Piper |
855 |
Clarisia biflora* |
403 |
|
Piperaceae |
855 |
Nectandra |
551 |
Trophis mexicana |
364 |
|
Acanthaceae |
742 |
Clarisia |
403 |
Piper arboreum |
343 |
|
Lauraceae |
622 |
Trophis |
364 |
Aphelandra aurantiaca |
312 |
|
Rubiaceae |
549 |
Aphelandra |
312 |
Rinorea guatemalensis |
309 |
|
Euphorbiaceae |
455 |
Rinorea |
311 |
Brosimum alicastrum* |
303 |
|
Violaceae |
353 |
Brosimum |
304 |
Nectandra ambigens* |
303 |
|
Arecaceae |
346 |
Psychotria |
258 |
Odontonema callistachyum |
258 |
|
Fabaceae |
222 |
Odontonema |
258 |
Schaueria parviflora |
170 |
|
Myrtaceae |
211 |
Eugenia |
184 |
Omphalea oleifera |
158 |
|
Norte de Chiapas |
|||||
|
Rubiaceae |
1,911 |
Faramea |
890 |
Faramea occidentalis* |
890 |
|
Myristicaceae |
729 |
Compsoneura |
729 |
Compsoneura mexicana |
729 |
|
Violaceae |
555 |
Rinorea |
553 |
Rinorea guatemalensis |
549 |
|
Moraceae |
470 |
Piper |
363 |
Trophis racemosa |
311 |
|
Piperaceae |
363 |
Trophis |
324 |
Posoqueria latifolia |
304 |
|
Fabaceae |
308 |
Posoqueria |
304 |
Piper aduncum |
241 |
|
Euphorbiaceae |
293 |
Psychotria |
293 |
Psychotria flava |
211 |
|
Salicaceae |
292 |
Nectandra |
208 |
Pleuranthodendron lindenii |
199 |
|
Malvaceae |
254 |
Pleuranthodendron |
199 |
Acalypha diversifolia |
162 |
|
Arecaceae |
253 |
Acalypha |
162 |
Mexocarpus tetragonus |
133 |
|
Total |
|||||
|
Rubiaceae |
4,501 |
Piper |
1,879 |
Faramea occidentalis* |
1,138 |
|
Moraceae |
3,037 |
Brosimum |
1,418 |
Rinorea guatemalensis |
983 |
|
Piperaceae |
1,879 |
Psychotria |
1,345 |
Brosimum alicastrum* |
842 |
|
Euphorbiaceae |
1,403 |
Faramea |
1,138 |
Compsoneura mexicana |
729 |
|
Tabla 2. Continúa |
|||||
|
Familias |
N |
Géneros |
N |
Especies |
N |
|
Lauraceae |
1,228 |
Rinorea |
1,105 |
Acalypha diversifolia |
682 |
|
Violaceae |
1,206 |
Nectandra |
1,043 |
Piper aduncum |
661 |
|
Fabaceae |
1,186 |
Trophis |
825 |
Psychotria poeppigiana |
592 |
|
Acanthaceae |
867 |
Palicourea |
749 |
Trophis racemosa |
443 |
|
Myristicaceae |
838 |
Compsoneura |
729 |
Ampelocera hottlei* |
436 |
|
Arecaceae |
766 |
Acalypha |
695 |
Clarisia biflora* |
429 |
* Especies tolerantes a la sombra (el resto son generalistas).
Tabla 3
Localizaciones de los nuevos registros de Coccoloba acuminata Kunth, Piper philodendroides Standl. et Steyerm. y Piper phytolaccifolium Opiz en México. Se incluye el tamaño del fragmento en hectáreas. N = Número de individuos; MC = Marqués de Comillas; NC = norte de Chiapas (incluye el fragmento F1 en el estado de Tabasco).
|
Especie |
Fragmento |
Tamaño |
UTM X |
UTM Y |
N |
Número de folio |
|
Coccoloba acuminata |
F1 (NC) |
3.34 |
0498835 |
1970419 |
3 |
MEXU: 1491354 |
|
0498838 |
1970479 |
1 |
||||
|
0498811 |
1970470 |
2 |
||||
|
0498752 |
1970518 |
1 |
||||
|
0498747 |
1970404 |
1 |
||||
|
Piper philodendroides |
F2 (MC) |
1.72 |
0742598 |
1779553 |
1 |
MEXU: 1491328 |
|
Piper phytolaccifolium |
F3 (NC) |
2.96 |
0464670 |
1968412 |
1 |
|
|
0464591 |
1968433 |
1 |
||||
|
0464623 |
1968441 |
1 |
||||
|
F4 (NC) |
4.75 |
0485566 |
1947747 |
1 |
||
|
F5 (NC) |
4.75 |
0483067 |
1953778 |
1 |
||
|
F6 (NC) |
44.2 |
0486779 |
1950213 |
1 |
||
|
0486708 |
1950146 |
1 |
||||
|
F7 (NC) |
86 |
0495388 |
1943739 |
1 |
||
|
F8 (NC) |
3.38 |
0478033 |
1976058 |
1 |
||
|
F9 (MC) |
18.25 |
0706086 |
1778473 |
3 |
||
|
F10 (NC) |
10.81 |
0739759 |
1797511 |
1 |
||
|
F11 (MC) |
5.96 |
0737788 |
1778641 |
3 |
MEXU: 1491339 |
Las familias más ricas en especies fueron similares en todas las regiones, pero cada región tuvo una composición florística distinta. Siete de las 10 familias con más riqueza de especies (Fabaceae, Rubiaceae, Piperaceae, Arecaceae, Malvaceae, Sapotaceae y Myrtaceae) estuvieron presentes en las 3 regiones de estudio; un resultado consistente con listados florísticos publicados para la selva Lacandona (Durán-Fernández et al., 2016; Levy-Tacher et al., 2006; Meave et al., 2008), Los Tuxtlas (Ibarra-Manríquez y Sinaca-Colín, 1995; Villaseñor et al., 2018) y Tabasco (Cowan, 1983; Vázquez-Negrín et al., 2011). Las diferencias en composición florística entre diferentes regiones tampoco son sorprendentes, ya que es bien sabido que los bosques tropicales tienen un alto recambio de especies —alta diversidad beta— (Arroyo-Rodríguez et al., 2013; Condit et al., 2002; De Cáceres et al., 2012). De hecho, Villaseñor et al. (2018) compararon las floras de la selva Lacandona y Los Tuxtlas y encontraron que tan solo comparten 60% de las especies de plantas vasculares, por lo que es normal que, en nuestro estudio, cada región haya estado dominada por un conjunto distinto de géneros y de especies.

En relación a la estructura de las comunidades, las 3 regiones de estudio muestran un patrón similar de dominancia y rareza. En particular, las comunidades fueron muy inequitativas, con pocas especies dominantes y un gran número de especies raras. Este patrón de distribución de abundancias es común en bosques tropicales de todo el mundo (Slik et al., 2015). La elevada rareza registrada en Los Tuxtlas —34% de especies representadas por 1 o 2 individuos— es similar a la observada por Villaseñor et al. (2018) para esta misma región. De forma interesante, la dominancia fue ligeramente mayor en el norte de Chiapas, lo que puede ser interpretado como una señal de homogenización biótica causada por la extrema deforestación que ha sufrido esta región. En particular, como argumentan Tabarelli et al. (2012), y consistente con evidencias empíricas para el bosque Atlántico de Brasil (Lôbo et al. 2011), las pocas especies —generalmente pioneras y generalistas— capaces de vivir en regiones altamente deforestadas como ésta, pueden acabar dominando los ensambles en los que se encuentran. En contraste, muchas especies, generalmente tolerantes a la sombra, acaban siendo raras, lo que aumenta la inequidad de la comunidad.
En este estudio reportamos a Piper philodendroides Standl. et Steyerm. como registro nuevo para México, un importante hallazgo si tenemos en cuenta que hasta ahora, esta especie tan solo había sido observada en el departamento de Izabal, en Guatemala (Standley y Steyermark, 1952; www.tropicos.org). El ejemplar fue encontrado en un fragmento de bosque maduro de 1.72 ha cercano a la frontera con Guatemala (fig. 6). Debido a la gran distancia que separa las 2 localidades donde se ha encontrado la especie (220 km), es muy probable que también esté presente en otros departamentos de Guatemala como Alta Verapaz, Quiché y Petén, así como en otras partes de la selva Lacandona. Por el contrario, Piper phytolaccifolium Opiz tiene una amplia distribución, extendiéndose desde Ecuador hasta México (fig. 6). Sorprendentemente, la especie fue encontrada tanto en Marqués de Comillas como en el norte de Chiapas (fig. 6) y estuvo presente en un total de 9 fragmentos (tabla 3), indicándonos que la especie podría ser relativamente abundante en ambas regiones. El tercer registro nuevo es el de Coccoloba acuminata Kunth, un arbusto o árbol pequeño de 2 a 8 m de alto que crece a bajas y medianas elevaciones en bosques húmedos y bosques de galería, desde Bolivia hasta México (Davidse et al., 2020). El hallazgo de C. acuminata en Tabasco es importante, ya que representa la distribución más septentrional de la especie, lo que sugiere que la distribución histórica de la especie pudo haber sido mucho más amplia (fig. 6).
En conclusión, el presente estudio destaca el potencial regenerativo y el valor de conservación de los fragmentos de selva. Desafortunadamente, en México gran parte de las expediciones botánicas y estudios sobre composición y diversidad florística son llevados a cabo en reservas y grandes extensiones de bosque (Durán-Fernández et al., 2016; Gómez-Domínguez et al., 2015; Ibarra-Manríquez y Sinaca-Colín, 1995; Meave et al., 2008; Ortiz-Rodríguez et al., 2016; Pérez-Farrera et al., 2012), por lo que el valor florístico de los fragmentos puede ser menospreciado. Nuestro estudio pone de manifiesto la necesidad de estudiar las comunidades de plantas que aún están presentes en los fragmentos de bosque, ya que por unidad de área —i.e., densidad de especies—, pueden albergar una flora similar o incluso más rica que áreas grandes y mejor conservadas (Fahrig, 2020; Watling et al., 2020). Por esta razón, debemos reforzar los esfuerzos de conservación para evitar la desaparición de estos fragmentos relativamente pequeños (Arroyo-Rodríguez et al., 2020). Además de resguardar muchas especies, estos fragmentos incrementan la conectividad del paisaje, constituyen fuentes de propágulos (i.e., semillas) para la futura restauración de la selva en campos o potreros abandonados, proveen una gran cantidad de recursos (agua, alimento, refugio) para un sinnúmero de especies de animales y proveen importantes bienes (madera, leña, carne) y servicios (captura de carbono, protección de cuerpos de agua, polinización, retención de suelo, dispersión de semillas, control de plagas) a las poblaciones humanas (Arroyo-Rodríguez et al., 2020). Por tanto, los incentivos económicos para la conservación de bosques, como el pago por servicios ambientales, no deberían limitarse a los fragmentos más grandes (> 100 ha; Conafor, 2020), ya que la exclusión de los fragmentos pequeños de estos programas de apoyo podría poner en riesgo la viabilidad a largo plazo de estos valiosos remanentes de bosque tropical.
Agradecimientos
La investigación fue financiada por el proyecto SEP-Conacyt (proyecto 2015-253946). RAG recibió una beca de doctorado Conacyt. Este trabajo cumple con parte de los requisitos del programa de doctorado del Posgrado en Ciencias Biológicas de la Universidad Nacional de México (UNAM), al cual RAG agradece todo el apoyo brindado. También agradecemos el apoyo logístico y administrativo dado por el Instituto de Investigaciones en Ecosistemas y Sustentabilidad (IIES-UNAM). RAG agradece a Angélica Ramírez Roa, Alberto Reyes García, Gilda Ortiz Calderón y Clara H. Ramos (Herbario Nacional, MEXU) por su ayuda en la búsqueda e identificación de especies; a Gilberto Jamangapé y a Santiago Sinaca por su inestimable ayuda en el campo y en la identificación in situ de muchos de los ejemplares encontrados; a Miguel Martínez-Ramos por proporcionar información valiosa acerca de la estrategia de regeneración de algunas especies y a Jorge A. Meave por su valioso apoyo en el diseño experimental. Los autores agradecen también a Martín Cervantes-López su gran ayuda en la preparación y elaboración de los análisis NMDS. Finalmente, queremos agradecer a los propietarios de Marqués de Comillas, Los Tuxtlas y el norte de Chiapas por dejarnos acceder a sus propiedades.
Apéndice. Listado de las plantas leñosas en regeneración encontradas en 60 fragmentos de bosque tropical en los 3 paisajes fragmentados estudiados: Marqués de Comillas (MC), Los Tuxtlas (LT) y norte de Chiapas (NC). La región del norte de Chiapas incluye un fragmento en el estado de Tabasco. Seguimos la nomenclatura establecida por la base de datos electrónica del Jardín Botánico de Missouri disponible en http://www.tropicos.org. Indicamos la abundancia de cada especie en cada región y mostramos, entre paréntesis, los fragmentos ocupados por la especie del total de fragmentos de la región (n = 20 fragmentos por región). También se señalan la estrategia de regeneración de cada especie, la referencia bibliográfica que se usó para establecer dicha estrategia, su categoría de riesgo (si se encuentra en alguna) y los nuevos registros. Estrategia de regeneración (Est. Reg.): T = tolerante a la sombra; G = generalista; P = pionera; I = introducida; ND = datos no disponibles. Categoría de riesgo referente a la Norma Oficial Mexicana NOM-059-Semarnat-2010: * = sujeta a protección especial; ** = amenazada; *** = en peligro de extinción. Registros nuevos = †.
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
ACANTHACEAE (5/6) |
||||||
|
Aphelandra aurantiaca Lindl. |
– |
312 (12/20) |
– |
312 |
G |
1, 2 |
|
Bravaisia integerrima (Spreng.) Standl. ** |
– |
– |
13 (2/20) |
13 |
G |
1, 3 |
|
Odontonema albiflorum Leonard |
21 (2/20) |
– |
78(6/20) |
99 |
T |
4 |
|
O. callistachyum (Schltdl. et Cham.) Kuntze |
– |
258 (13/20) |
– |
258 |
G |
5 |
|
Schaueria parviflora (Leonard) T.F. Daniel |
– |
170 (6/20) |
– |
170 |
G |
6 |
|
Stenostephanus silvaticus (Nees) T.F. Daniel |
– |
– |
15 (3/20) |
15 |
G |
4 |
|
ACTINIDIACEAE (1/1) |
||||||
|
Saurauia yasicae Loes. |
– |
1 (1/20) |
– |
1 |
P |
1, 7 |
|
AMARANTHACEAE (1/1) |
||||||
|
Iresine arbuscula Uline et W.L. Bray |
6 (3/20) |
3 (3/20) |
34 (9/20) |
43 |
T |
4, 5 |
|
ANACARDIACEAE (4/5) |
||||||
|
Astronium graveolens Jacq. ** |
12 (6/20) |
– |
– |
12 |
G |
1, 8 |
|
Mangifera indica L. |
– |
– |
2 (1/20) |
2 |
I |
5 |
|
Spondias mombin L. |
6 (6/20) |
– |
20 (7/20) |
26 |
P |
5, 7, 9, 10 |
|
S. radlkoferi Donn. Sm. ** |
1 (1/20) |
5 (4/20) |
– |
6 |
P |
5, 6, 7 |
|
Tapirira mexicana Marchand |
– |
6 (4(20) |
– |
6 |
G |
5, 7 |
|
ANNONACEAE (9/12) |
||||||
|
Annona globiflora Schltdl. |
– |
2 (1/20) |
– |
2 |
G |
4 |
|
A. scleroderma Saff. |
2 (1/20) |
– |
– |
2 |
T |
5 |
|
Cymbopetalum baillonii R.E. Fr. |
– |
57 (14/20) |
– |
57 |
T |
4, 7 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
C. mayanum Lundell |
58 (15/20) |
– |
13 (8/20) |
71 |
T |
4 |
|
Guamia sp. |
– |
24 (10/20) |
– |
24 |
ND |
– |
|
Guatteria anomala R.E. Fr. ** |
7 (4/20) |
– |
1 (1/20) |
8 |
T |
9 |
|
Mosannona depressa (Baill.) Chatrou |
– |
5 (3/20) |
– |
5 |
G |
4, 11 |
|
Rollinia mucosa (Jacq.) Baill. |
– |
3 (2/20) |
2 (2/20) |
5 |
P |
5, 12 |
|
Stenanona cauliflora (J.W. Walker) G.E. Schatz |
– |
– |
4 (2/20) |
4 |
T |
13 |
|
Tridimeris hahniana Baill. |
– |
1 (1/20) |
– |
1 |
T |
14 |
|
T. tuxtlensis G.E. Schatz |
– |
2 (1/20) |
– |
2 |
ND |
– |
|
Xylopia frutescens Aubl. |
74 (8/20) |
– |
7 (3/20) |
81 |
G |
5, 8, 12 |
|
APOCYNACEAE (4/5) |
||||||
|
Aspidosperma megalocarpon Müll. Arg. |
– |
19 (4/20) |
– |
19 |
T |
4, 8, 9 |
|
Stemmadenia donnell-smithii (Rose) Woodson |
– |
13 (8/20) |
– |
13 |
P |
5, 6, 9 |
|
S. litoralis (Kunth) L. Allorge |
30 (14/20) |
28 (9/20) |
20 (7/20) |
78 |
G |
3 |
|
Tabernaemontana alba Mill. |
24 (10/20) |
– |
36 (13/20) |
60 |
G |
3, 5 |
|
Thevetia ahouai (L.) A. DC. |
46 (15/20) |
– |
8 (4/20) |
54 |
P |
1, 3, 5, 8 |
|
AQUIFOLIACEAE (1/2) |
||||||
|
Ilex quercetorum I.M. Johnst. |
– |
1 (1/20) |
– |
1 |
T |
4, 5, 7 |
|
I. valerii Standl. |
– |
1 (1/20) |
– |
1 |
T |
4, 7 |
|
ARALIACEAE (1/1) |
||||||
|
Dendropanax arboreus (L.) Decne. et Planch. |
68 (15/20) |
26 (10/20) |
73 (13/20) |
167 |
G |
7, 8, 9, 15 |
|
ARECACEAE (8/16) |
||||||
|
Astrocaryum mexicanum Liebm. ex Mart. |
– |
84 (11/20) |
33 (6/20) |
117 |
G |
5, 37 |
|
Attalea cohune Mart. |
3 (2/20) |
– |
5 (4/20) |
8 |
ND |
– |
|
A. liebmannii (Becc.) Zona |
– |
1 (1/20) |
– |
1 |
P |
38 |
|
Bactris major Jacq. * |
17 (1/20) |
– |
19(3/20) |
36 |
G |
1 |
|
B. mexicana Mart. |
34 (8/20) |
4 (3/20) |
44 (10/20) |
82 |
T |
1, 4 |
|
Chamaedorea elatior Mart. ** |
– |
1 (1/20) |
– |
1 |
T |
4, 5 |
|
C. elegans Mart. |
1 (1/20) |
– |
– |
1 |
T |
4, 5 |
|
C. ernesti-augusti H. Wendl. ** |
– |
9 (5/20) |
1 (1/20) |
10 |
T |
4, 5 |
|
C. oblongata Mart. ** |
62 (9/20) |
79 (12/20) |
7 (2/20) |
148 |
T |
1, 5 |
|
C. pinnatifrons (Jacq.) Oerst. ** |
28 (12/20) |
74 (14/20) |
7 (4/20) |
109 |
T |
1, 4 |
|
C. tepejilote Liebm. ** |
14 (6/20) |
60 (12/20) |
137 (5/20) |
211 |
T |
1, 5 |
|
C. woodsoniana L.H. Bailey ** |
– |
9 (2/20) |
– |
9 |
T |
1, 4 |
|
Euterpe precatoria Mart. |
4 (3/20) |
– |
– |
4 |
P |
39 |
|
Geonoma interrupta (Ruiz et Pav.) Mart. ** |
3 (2/20) |
– |
– |
3 |
T |
1, 5 |
|
Reinhardtia gracilis (H. Wendl.) Drude ex Dammer * |
– |
25 (6/20) |
– |
25 |
T |
1, 5 |
|
Sabal mauritiiformis (H. Karst.) Griseb. et H. Wendl. |
1 (1/20) |
– |
– |
1 |
P |
5 |
|
ASPARAGACEAE (1/1) |
||||||
|
Dracaena americana Donn. Sm. |
3 (2/20) |
– |
– |
3 |
G |
4 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
ASTERACEAE (3/3) |
||||||
|
Critonia morifolia (Mill.) R.M. King et H. Rob. |
– |
1 (1/20) |
– |
1 |
P |
4 |
|
Koanophyllon pittieri (Klatt) R.M. King et H. Rob. |
– |
44 (12/20) |
– |
44 |
G |
3, 4, 5 |
|
Neurolaena lobata (L.) Cass. |
– |
16 (8/20) |
– |
16 |
P |
1, 3, 7 |
|
BIGNONIACEAE (5/7) |
||||||
|
Amphitecna apiculata A.H. Gentry |
37 (13/20) |
– |
– |
37 |
T |
4 |
|
A. tuxtlensis A.H. Gentry |
– |
10 (5/20) |
– |
10 |
T |
4, 7, 16 |
|
A. steyermarkii (A.H. Gentry) A.H. Gentry |
2 (1/20) |
– |
– |
2 |
T |
4, 17 |
|
Handroanthus guayacan (Seem.) S.O.Grose |
2 (2/20) |
– |
– |
2 |
G |
4, 5, 8 |
|
Rochefortia spinosa (Jacq.) Urb. |
– |
2 (1/20) |
– |
2 |
P |
4, 7 |
|
Parmentiera aculeata (Kunth) Seem. |
– |
– |
13 (1/20) |
13 |
P |
5, 9 |
|
Tabebuia rosea (Bertol.) DC. |
2 (1/20) |
– |
2 (2/20) |
4 |
G |
5, 8, 9, 18 |
|
BURSERACEAE (2/3) |
||||||
|
Bursera simaruba (L.) Sarg. |
19 (8/20) |
3 (3/20) |
11 (7/20) |
33 |
G |
9, 10, 19 |
|
Protium confusum (Rose) Pittier |
165 (9/20) |
– |
– |
165 |
G |
20 |
|
P. copal (Schltdl. et Cham.) Engl. |
139 (17/20) |
7 (2/20) |
20 (5/20) |
166 |
T |
9, 15 |
|
CALOPHYLLACEAE (3/5) |
||||||
|
Calophyllum brasiliense Cambess. ** |
229 (16/20) |
15 (3/20) |
46 (8/20) |
290 |
T |
7, 9 |
|
Calophyllum aff. brasiliense 1 |
– |
1 (1/20) |
– |
1 |
ND |
– |
|
Calophyllum aff. brasiliense 2 |
4 (2/20) |
– |
4 (3/20) |
8 |
ND |
– |
|
Aphananthe monoica (Hemsl.) J.-F. Leroy |
– |
2 (1/20) |
– |
2 |
T |
3, 4 |
|
Trema micrantha (L.) Blume |
1 (1/20) |
– |
– |
1 |
P |
6, 8, 9 |
|
CAPPARACEAE (2/4) |
||||||
|
Capparidastrum mollicellum (Standl.) Cornejo et Iltis |
– |
6 (3/20) |
– |
6 |
G |
4 |
|
C. quiriguense (Standl.) Cornejo et Iltis |
4 (1/20) |
– |
5 (1/20) |
9 |
T |
4 |
|
C. tuxtlense Cornejo et Iltis |
– |
5 (3/20) |
– |
5 |
ND |
– |
|
Crateva tapia L. |
– |
9 (5/20) |
– |
9 |
P |
1, 3, 8 |
|
CARICACEAE (3/3) |
||||||
|
Carica papaya L. |
– |
3 (2/20) |
34 (3/20) |
37 |
P |
6 |
|
Jacaratia dolichaula (Donn. Sm.) Woodson |
12 (7/20) |
1 (1/20) |
– |
13 |
G |
1 |
|
Vasconcellea cauliflora (Jacq.) A. DC. |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
CELASTRACEAE (4/6) |
||||||
|
Crossopetalum filipes (Sprague) Lundell |
– |
2 (2/20) |
– |
2 |
G |
4 |
|
C. parviflorum (Hemsl.) Lundell |
– |
3 (3/20) |
– |
3 |
T |
7, 8 |
|
Elaeodendron xylocarpum (Vent.) DC. |
– |
1 (1/20) |
– |
1 |
G |
4, 21 |
|
Haydenoxylon haberianum (Hammel) M. P. Simmons |
– |
7 (2/20) |
– |
7 |
G |
4 |
|
Maytenus purpusii Lundell |
– |
13 (6/20) |
– |
13 |
T |
4 |
|
M. schippii Lundell |
– |
1 (1/20) |
– |
1 |
T |
4, 7 |
|
CHRYSOBALANACEAE (2/5) |
||||||
|
Hirtella americana L. |
121 (15/20) |
– |
1 (1/20) |
122 |
G |
1, 4, 5 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
H. racemosa Lam. |
79 (17/20) |
– |
65 (11/20) |
144 |
G |
3, 5 |
|
H. triandra Sw. |
– |
10 (1/20) |
– |
10 |
G |
3, 4 |
|
Licania hypoleuca Benth. |
201 (10/20) |
– |
12 (1/20) |
213 |
T |
1, 4, 8 |
|
L. platypus (Hemsl.) Fritsch |
50 (5/20) |
– |
7 (3/20) |
57 |
T |
8, 9, 10 |
|
CLETHRACEAE (1/1) |
||||||
|
Clethra macrophylla M. Martens et Galeotti |
1 (1/20) |
– |
– |
1 |
G |
4, 22 |
|
CLUSIACEAE (2/2) |
||||||
|
Garcinia intermedia (Pittier) Hammel |
13 (4/20) |
100 (13/20) |
– |
113 |
T |
8, 9 |
|
Symphonia globulifera L. f. |
– |
– |
5 (2/20) |
5 |
T |
3, 5, 8 |
|
COMBRETACEAE (1/1) |
||||||
|
Terminalia amazonia (J.F. Gmel.) Exell |
8 (6/20) |
– |
– |
8 |
T |
7, 9, 10 |
|
CORDIACEAE (1/6) |
||||||
|
Cordia alliodora Ruiz et Pav.) Oken |
– |
3 (2/20) |
– |
3 |
G |
8, 9, 10 |
|
C. bicolor A. DC. |
2 (2/20) |
– |
– |
2 |
G |
3 |
|
C. diversifolia Pav. ex A. DC. |
8 (6/20) |
1 (1/20) |
– |
9 |
G |
3 |
|
C. dodecandra DC. |
– |
1 (1/20) |
– |
1 |
G |
3, 7 |
|
C. megalantha S.F. Blake |
– |
3 (2/20) |
– |
3 |
G |
5, 7 |
|
C. stellifera I.M. Johnst. |
– |
7 (2/20) |
– |
7 |
G |
3, 7 |
|
EBENACEAE (1/2) |
||||||
|
Diospyros conzattii Standl. *** |
– |
5 (1/20) |
– |
5 |
T |
20 |
|
D. nigra (J.F. Gmel.) Perr. |
– |
36 (7/20) |
– |
36 |
T |
20 |
|
ELAEOCARPACEAE (1/1) |
||||||
|
Sloanea tuerckheimii Donn. Sm. |
21 (4/20) |
– |
– |
21 |
T |
4, 5 |
|
ERYTHROXYLACEAE (1/1) |
||||||
|
Erythroxylum macrophyllum Cav. |
– |
8 (3/20) |
12 (7/20) |
20 |
G |
4 |
|
EUPHORBIACEAE (13/18) |
||||||
|
Acalypha diversifolia Jacq. |
434 (11/20) |
86 (6/20) |
162 (9/20) |
682 |
G |
1, 8 |
|
A. skutchii I.M. Johnst. |
– |
13 (3/20) |
– |
13 |
G |
6 |
|
Adelia barbinervis Schltdl. et Cham. |
– |
17 (2/20) |
– |
17 |
G |
10 |
|
Alchornea chiapasana Miranda |
– |
3 (1/20) |
– |
3 |
ND |
– |
|
A. latifolia Sw. |
9 (3/20) |
1 (1/20) |
– |
10 |
G |
6, 7, 8 |
|
Cnidoscolus multilobus (Pax) I.M. Johnst. |
– |
4 (3/20) |
– |
4 |
P |
6, 7, 9 |
|
Croton billbergianus Müll. Arg. |
– |
10 (3/20) |
– |
10 |
G |
4 |
|
C. niveus Jacq. |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
C. schiedeanus Schltdl. |
158 (18/20) |
153 (19/20) |
48 (13/20) |
359 |
G |
4 |
|
C. soliman Cham. et Schltdl. |
– |
3 (1/20) |
– |
3 |
P |
4 |
|
Garcia parviflora Lundell |
– |
4 (3/20) |
– |
4 |
T |
4 |
|
Gymnanthes lucida Sw. |
– |
1 (1/20) |
– |
1 |
T |
4 |
|
Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg. |
– |
– |
9 (1/20) |
9 |
I |
1, 8 |
|
Mabea occidentalis Benth. |
47 (4/20) |
– |
74 (4/20) |
121 |
T |
8 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
Omphalea oleifera Hemsl. |
– |
158 (12/20) |
– |
158 |
G |
6 |
|
Pleradenophora tuerckheimiana (Pax et K. Hoffm.) A.L. Melo et Esser |
6 (2/20) |
– |
– |
6 |
T |
4 |
|
Sapium lateriflorum Hemsl. |
1 (1/20) |
– |
– |
1 |
G |
20 |
|
Tetrorchidium rotundatum Standl. ** |
– |
1 (1/20) |
– |
1 |
P |
7 |
|
FABACEAE (25/44) |
||||||
|
Acacia cornigera (L.) Willd. |
17 (4/20) |
– |
– |
17 |
P |
1, 5, 7, 4 |
|
A. mayana Lundell |
18 (8/20) |
– |
3 (1/20) |
21 |
G |
4 |
|
A. polyphylla DC. |
– |
– |
3 (2/20) |
3 |
P |
4, 5 |
|
Albizia leucocalyx (Britton et Rose) L. Rico |
8 (5/20) |
– |
– |
8 |
P |
23 |
|
A. purpusii Britton et Rose |
– |
2 (2/20) |
– |
2 |
P |
4 |
|
A. tomentosa (Micheli) Standl |
– |
14 (8/20) |
– |
14 |
P |
4 |
|
Calliandra tergemina (L.) Benth. |
– |
1 (1/20) |
– |
1 |
G |
4, 5 |
|
Chloroleucon mangense (Jacq.) Britton et Rose |
– |
1 (1/20) |
– |
1 |
P |
4 |
|
Cojoba arborea (L.) Britton et Rose |
4 (2/20) |
1 (1/20) |
– |
5 |
T |
1, 4, 5 |
|
C. graciliflora (S.F. Blake) Britton et Rose |
2 (2/20) |
– |
– |
2 |
G |
4 |
|
Cynometra oaxacana Brandegee |
– |
– |
9 (2/20) |
9 |
T |
20 |
|
C. retusa Britton et Rose |
– |
8 (5/20) |
1 (1/20) |
9 |
T |
4, 7, 20 |
|
Dalbergia glomerata Hemsl. |
6 (3/20) |
2 (2/20) |
8 |
G |
4, 7, 24 |
|
|
Dialium guianense (Aubl.) Sandwith |
89 (17/20) |
1 (1/20) |
14 (8/20) |
104 |
T |
7, 9 |
|
Dussia mexicana (Standl.) Harms |
– |
– |
2 (1/20) |
2 |
T |
7, 9 |
|
Enterolobium cyclocarpum (Jacq.) Griseb. |
– |
– |
18 (2/20) |
18 |
P |
1, 8 |
|
Erythrina folkersii Krukoff et Moldenke |
2 (2/20) |
3 (3/20) |
– |
5 |
G |
6, 9 |
|
Haematoxylum campechianum L. |
– |
– |
4 (1/20) |
4 |
P |
4 |
|
Indigofera suffruticosa Mill. |
15 (6/20) |
– |
– |
15 |
G |
25 |
|
Inga acrocephala Steud. |
16 (5/20) |
– |
16 |
G |
4, 7 |
|
|
I. calcicola M. Sousa |
1 (1/20) |
– |
– |
1 |
T |
4 |
|
I. mexicana (T.D. Penn.) M. Sousa |
– |
21 (8/20) |
– |
21 |
T |
4 |
|
I. paterno Harms |
– |
7 (4/20) |
– |
7 |
G |
5, 7 |
|
I. punctata Willd. |
83 (8/20) |
36 (2/20) |
93 (9/20) |
212 |
G |
1, 26 |
|
I. sapindoides Willd. |
63 (12/20) |
3 (3/20) |
21 (5/20) |
87 |
G |
1, 7 |
|
I. sinacae M. Sousa et Ibarra-Manr. |
– |
6 (3/20) |
– |
6 |
G |
4, 7 |
|
I. thibaudiana DC. |
84 (10/20) |
– |
– |
84 |
G |
1, 4, 8 |
|
I. vera Willd. |
– |
– |
1 (1/20) |
1 |
G |
1, 5, 8, 27 |
|
Leucaena leucocephala (Lam.) de Wit |
– |
– |
3 (2/20) |
3 |
P |
5, 9 |
|
Lonchocarpus cruentus Lundell |
62 (14/20) |
13 (2/20) |
75 (11/20) |
150 |
T |
7, 9 |
|
L. guatemalensis Benth. |
7 (3/20) |
13 (5/20) |
20 (3/20) |
40 |
G |
5, 7 |
|
L. luteomaculatus Pittier |
8 (4/20) |
2 (1/20) |
– |
10 |
G |
4 |
|
L. rugosus Benth. |
– |
1 (1/20) |
– |
1 |
G |
4, 5 |
|
Mariosousa usumacintensis (Lundell) |
7 (5/20) |
– |
6 (4/20) |
13 |
G |
4 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
Ormosia isthmensis Standl. *** |
3 (3/20) |
– |
1 (1/20) |
4 |
G |
20 |
|
O. panamensis Benth. ex Seem. |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
Pithecellobium lanceolatum (Humb. et Bonpl. |
– |
– |
1 (1/20) |
1 |
G |
4 |
|
P. macrandrium Donn. Sm. |
38 (13/20) |
– |
3 (2/20) |
41 |
T |
20 |
|
Platymiscium dimorphandrum Donn. Sm. |
7 (5/20) |
5 (4/20) |
4 (3/20) |
16 |
G |
4 |
|
Pterocarpus rohrii Vahl |
8 (5/20) |
47 (12/20) |
14 (8/20) |
69 |
T |
4, 7, 8 |
|
Senna papillosa (Britton et Rose) |
1 (1/20) |
1 (1/20) |
– |
2 |
P |
1, 7 |
|
Swartzia guatemalensis (Donn. Sm.) Pittier |
17 (4/20) |
17 (6/20) |
8 (3/20) |
42 |
G |
4 |
|
Vatairea lundellii (Standl.) Killip ex Record *** |
116 (16/20) |
– |
2 (2/20) |
118 |
G |
4, 20 |
|
Zygia peckii (B.L. Rob.) Britton et Rose |
10 (4/20) |
– |
– |
10 |
G |
4 |
|
HYPERICACEAE (1/1) |
||||||
|
Vismia camparaguey Sprague et L. Riley |
1 (1/20) |
– |
– |
1 |
P |
3, 5 |
|
ICACINACEAE (1/1) |
||||||
|
Mappia racemosa Jacq. |
– |
3 (2/20) |
– |
3 |
T |
5, 7 |
|
LACISTEMATACEAE (1/1) |
||||||
|
Lacistema aggregatum (P.J. Bergius) Rusby |
147 (19/20) |
2 (2/20) |
26 (9/20) |
175 |
G |
1, 4, 5 |
|
LAMIACEAE (2/4) |
||||||
|
Aegiphila costaricensis Moldenke |
– |
2 (2/20) |
– |
2 |
G |
1, 3, 4, 5 |
|
A. monstrosa Moldenke |
3 (2/20) |
4 (2/20) |
– |
7 |
P |
1, 3, 4, 5 |
|
Cornutia grandifolia (Schltdl. et Cham.) Schauer |
– |
6 (3/20) |
– |
6 |
G |
5 |
|
C. pyramidata L. |
– |
1 (1/20) |
– |
1 |
P |
1, 3, 5 |
|
LAURACEAE (4/21) |
||||||
|
Cinnamomum pachypes Kosterm. |
– |
3 (2/20) |
– |
3 |
ND |
– |
|
Cinnamomum sp. |
– |
1 (1/20) |
– |
1 |
ND |
– |
|
Licaria excelsa Kosterm. |
83 (15/20) |
– |
31 (10/20) |
114 |
G |
4 |
|
L. peckii (I.M. Johnst.) Kosterm. |
– |
2 (2/20) |
– |
2 |
T |
4, 5 |
|
L. velutina van der Werff |
– |
5 (3/20) |
– |
5 |
T |
4, 7 |
|
Nectandra ambigens (S.F. Blake) C.K. Allen |
9 (5/20) |
303 (10/20) |
75 (5/20) |
387 |
T |
4, 7, 9 |
|
N. belizensis (Lundell) C.K. Allen |
10 (4/20) |
– |
14 (1/20) |
24 |
G |
4 |
|
N. coriacea (Sw.) Griseb. |
– |
6 (3/20) |
– |
6 |
G |
4 |
|
N. cuspidata Nees et Mart. |
– |
2 (2/20) |
– |
2 |
G |
4, 5, 8 |
|
N. lundellii C.K. Allen |
– |
48 (8/20) |
– |
48 |
T |
4 |
|
N. reticulata (Ruiz et Pav.) Mez |
36 (11/20) |
9 (5/20) |
118 (16/20) |
163 |
G |
4, 5 |
|
N. salicifolia (Kunth) Nees |
229 (17/20) |
155 (10/20) |
1 (1/20) |
385 |
T |
15 |
|
N. turbacensis (Kunth) Nees |
– |
28 (3/20) |
– |
28 |
G |
4 |
|
Ocotea cernua (Nees) Mez |
– |
3 (1/20) |
– |
3 |
T |
8 |
|
O. dendrodaphne Mez |
– |
6 (4/20 |
– |
6 |
T |
1, 3, 5, 7 |
|
O. helicterifolia (Meisn.) Hemsl. |
– |
19 (2/20) |
– |
19 |
G |
4, 5 |
|
O. rubriflora Mez |
– |
13 (10/20) |
– |
13 |
T |
7, 10 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
O. sinuata (Mez) Rohwer |
– |
1 (1/20) |
– |
1 |
T |
10 |
|
O. subalata Lundell |
– |
9 (1/20) |
– |
9 |
ND |
– |
|
O. uxpanapana T. Wendt et van der Werff |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
Ocotea sp. |
– |
8 (1/20) |
– |
8 |
ND |
– |
|
MAGNOLIACEAE (1/1) |
||||||
|
Magnolia sinacacolinii A. Vázquez |
– |
1 (1/20) |
– |
1 |
ND |
– |
|
MALPIGHIACEAE (2/2) |
||||||
|
Bunchosia lindeniana A. Juss. |
185 (17/20) |
29 (10/20) |
24 (11/20) |
238 |
G |
4, 5 |
|
Malpighia glabra L. |
– |
5 (2/20) |
– |
5 |
G |
4, 6 |
|
MALVACEAE (11/13) |
||||||
|
Hampea nutricia Fryxell |
– |
51 (14/20) |
32 (8/20) |
83 |
G |
4 |
|
H. stipitata S. Watson |
7 (2/20) |
– |
1 (1/20) |
8 |
G |
20 |
|
Heliocarpus appendiculatus Turcz. |
– |
1 (1/20) |
– |
1 |
P |
1, 6, 9 |
|
Lopimia malacophylla (Link et Otto) Mart. |
– |
– |
2 (1/20) |
2 |
G |
4 |
|
Luehea speciosa Willd. |
4 (3/20) |
– |
– |
4 |
P |
8, 9 |
|
Malvaviscus arboreus Cav. |
1 (1/20) |
1 (1/20) |
25 (1/20) |
27 |
G |
4, 5 |
|
Mortoniodendron guatemalense Standl. |
– |
1 (1/20) |
– |
1 |
T |
7, 9 |
|
Pachira aquatica Aubl. |
18 (8/20) |
– |
116 (5/20) |
134 |
T |
5, 8, 20 |
|
Pavonia schiedeana Steud. |
– |
12 (5/20) |
29 (1/20) |
41 |
P |
1, 25 |
|
Quararibea funebris (La Llave) Vischer |
4 (2/20) |
3 (2/20) |
7 (2/20) |
14 |
T |
5, 7, 9 |
|
Q. yunckeri Standl. |
27 (11/20) |
20 (9/20) |
19 (5/20) |
66 |
T |
4, 7 |
|
Sterculia mexicana R. Br. |
– |
6 (3/20) |
17 (4/20) |
23 |
G |
4 |
|
Theobroma cacao L. |
12 (1/20) |
– |
6 (3/20) |
18 |
I |
– |
|
MELASTOMATACEAE (5/14) |
||||||
|
Aciotis rostellata (Naudin) Triana |
10 (2/20) |
– |
– |
10 |
P |
3, 28 |
|
Clidemia petiolaris (Schltdl. et Cham.) |
7 (4/20) |
2 (1/20) |
– |
9 |
G |
20 |
|
C. setosa (Triana) Gleason |
– |
1 (1/20) |
– |
1 |
G |
20 |
|
Conostegia hirtella Cogn. |
35 (12/20) |
– |
3 (2/20) |
38 |
G |
20 |
|
C. xalapensis (Bonpl.) D. Don ex DC. |
2 (2/20) |
2 (2/20) |
– |
4 |
P |
3, 7, 8 |
|
Miconia argentea (Sw.) DC. |
38 (9/20) |
– |
1 (1/20) |
39 |
P |
1, 3, 5, 7, 8 |
|
M. glaberrima (Schltdl.) Naudin |
25 (9/20) |
– |
5 (2/20) |
30 |
G |
3, 7 |
|
M. hondurensis Donn. Sm. |
25 (11/20) |
– |
– |
25 |
G |
4, 5 |
|
M. ibarrae Almeda |
– |
1 (1/20) |
– |
1 |
ND |
– |
|
M. impetiolaris (Sw.) D. Don ex DC. |
1 (1/20) |
– |
1 (1/20) |
2 |
P |
3, 5, 8, 21 |
|
M. trinervia (Sw.) D. Don ex Loudon |
4 (2/20) |
– |
1 (1/20) |
5 |
P |
4, 5, 7 |
|
Miconia sp. |
1 (1/20) |
– |
– |
1 |
ND |
– |
|
Mouriri gleasoniana Standl. |
2 (1/20) |
– |
– |
2 |
T |
4 |
|
M. myrtilloides (Sw.) Poir. |
137 (15/20) |
– |
92 (7/20) |
229 |
T |
8, 29 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
MELIACEAE (3/11) |
||||||
|
Guarea glabra Vahl |
64 (14/20) |
77 (17/20) |
23 (9/20) |
164 |
T |
7, 9 |
|
G. grandifolia DC. |
11 (6/20) |
– |
12 (6/20) |
23 |
T |
7, 8 |
|
Guarea sp. 1 |
– |
14 (9/20) |
– |
14 |
ND |
– |
|
Guarea sp. 2 |
– |
– |
11 (3/20) |
11 |
ND |
– |
|
Swietenia macrophylla King |
6 (2/20) |
– |
1 (1/20) |
7 |
G |
8, 30 |
|
Trichilia breviflora S.F. Blake et Standl. |
– |
39 (10/20) |
30 (5/20) |
69 |
T |
4, 7 |
|
T. erythrocarpa Lundell |
11 (3/20) |
– |
4 (2/20) |
15 |
G |
4 |
|
T. havanensis Jacq. |
3 (2/20) |
– |
4 (3/20) |
7 |
G |
7, 5, 9 |
|
T. martiana C. DC. |
– |
1 (1/20) |
– |
1 |
T |
4 |
|
T. moschata Sw. |
– |
6 (4/20) |
82 (8/20) |
88 |
T |
4, 7 |
|
Trichilia sp |
16 (6/20) |
– |
– |
16 |
ND |
– |
|
MENISPERMACEAE (1/2) |
||||||
|
Hyperbaena mexicana Miers |
2 (2/20) |
5 (3/20) |
– |
7 |
T |
4 |
|
Hyperbaena sp. |
2 (2/20) |
– |
– |
2 |
ND |
– |
|
METTENIUSACEAE (1/2) |
||||||
|
Oecopetalum greenmanii Standl. et Steyerm. |
12 (9/20) |
– |
– |
12 |
G |
20 |
|
O. mexicanum Greenm. et C.H. Thomps. |
– |
– |
2 (1/20) |
2 |
G |
4 |
|
MONIMIACEAE (1/1) |
||||||
|
Mollinedia viridiflora Tul. |
91 (12/20) |
3 (3/20) |
5 (1/20) |
99 |
T |
4, 7 |
|
MORACEAE (6/9) |
||||||
|
Brosimum alicastrum Sw. |
468 (15/20) |
303 (12/20) |
71 (10/20) |
842 |
T |
7, 15 |
|
B. costaricanum Liebm. |
407 (18/20) |
– |
9 (5/20) |
416 |
T |
3, 4, 5 |
|
B. guianense (Aubl.) Huber |
151 (16/20) |
1 (1/20) |
8 (2/20) |
160 |
T |
1, 3, 8 |
|
Castilla elastica Sessé ex Cerv. |
156 (18/20) |
1 (1/20) |
26 (7/20) |
183 |
G |
8, 9 |
|
Clarisia biflora Ruiz et Pav. |
3 (3/20) |
403 (12/20) |
23 (6/20) |
429 |
T |
1, 3, 4, 7, 31 |
|
Poulsenia armata (Miq.) Standl. |
– |
14 (7/20) |
9 (5/20) |
23 |
T |
7, 8, 32 |
|
Pseudolmedia glabrata (Liebm.) C.C. Berg |
3 (3/20) |
156 (19/20) |
– |
159 |
T |
32 |
|
Trophis mexicana (Liebm.) Bureau |
5 (4/20) |
364 (19/20) |
13 (7/20) |
382 |
G |
4 |
|
T. racemosa (L.) Urb. |
132 (16/20) |
– |
311 (19/20) |
443 |
G |
4, 5, 8 |
|
MYRISTICACEAE (2/3) |
||||||
|
Compsoneura mexicana (Hemsl.) Janovec |
– |
– |
729 (13/20) |
729 |
G |
4 |
|
Virola guatemalensis (Hemsl.) Warb. |
– |
36 (9/20) |
– |
36 |
T |
1, 5, 7 |
|
V. koschnyi Warb. |
73 (7/20) |
– |
– |
73 |
T |
1, 5, 8 |
|
MYRTACEAE (6/14) |
||||||
|
Calyptranthes pallens Griseb. |
– |
21 (4/20) |
– |
21 |
G |
4 |
|
Eugenia acapulcensis Steud. |
– |
27 (10/20) |
– |
27 |
G |
4, 7 |
|
E. aeruginea DC. |
– |
22 (4/20) |
– |
22 |
G |
4 |
|
E. breedlovei Barrie |
– |
– |
24 (1/20) |
24 |
T |
4 |
|
E. capuli (Schltdl. et Cham.) Hook. et Arn. |
43 (12/20) |
38 (5/20) |
59 (12/20) |
140 |
G |
4, 7 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
E. colipensis O. Berg |
– |
15 (5/20) |
– |
15 |
T |
4, 7 |
|
E. inirebensis P.E. Sánchez |
– |
13 (4/20) |
– |
13 |
T |
4, 7 |
|
E. mexicana Steud. |
108 (17/20) |
69 (18/20) |
29 (12/20) |
206 |
T |
4 |
|
E. nigrita Lundell |
20 (9/20) |
– |
9 (6/20) |
29 |
G |
20 |
|
E. octopleura Krug et Urb. |
20 (2/20) |
– |
23 (4/20) |
43 |
G |
4, 5 |
|
Myrciaria floribunda (H. West ex Willd.) O. Berg |
– |
1 (1/20) |
– |
1 |
G |
4, 5 |
|
Pimenta dioica (L.) Merr. |
– |
5 (3/20) |
– |
5 |
T |
7, 9, 15 |
|
Psidium friedrichsthalianum (O. Berg) Nied. |
94 (10/20) |
– |
3 (2/20) |
97 |
G |
1, 3, 5 |
|
Syzygium jambos (L.) Alston |
1 (1/20) |
– |
18 (8/20) |
19 |
I |
1, 3, 5 |
|
NYCTAGINACEAE (1/2) |
||||||
|
Neea psychotrioides Donn. Sm. |
– |
2 (2/20) |
– |
2 |
G |
4, 5, 20 |
|
N. stenophylla Standl. |
14 (8/20) |
– |
8 (2/20) |
22 |
G |
4 |
|
OCHNACEAE (1/3) |
||||||
|
Ouratea curvata (A. St.-Hil.) Engl. ex Dwyer |
28 (5/20) |
– |
20 (8/20) |
48 |
G |
3 |
|
O. lucens (Kunth) Engl. |
138 (13/20) |
– |
1 (1/20) |
139 |
G |
3, 4, 5 |
|
O. theophrasta (Planch.) Baill. |
– |
1 (1/20) |
– |
1 |
T |
4, 5 |
|
OLEACEAE (1/1) |
||||||
|
Chionanthus ligustrinus (Sw.) Pers. |
– |
2 (2/20) |
– |
2 |
G |
4 |
|
PASSIFLORACEAE (1/1) |
||||||
|
Erblichia odorata Seem. |
– |
– |
5 (2/20) |
5 |
G |
4 |
|
PICRAMNIACEAE (1/3) |
||||||
|
Picramnia andicola Tul. |
4 (2/20) |
1 (1/20) |
4 (2/20) |
9 |
G |
4 |
|
P. antidesma Sw. |
– |
11 (3/20) |
– |
11 |
G |
4, 5 |
|
P. polyantha (Benth.) Planch. |
8 (1/20) |
– |
7 (1/20) |
15 |
P |
4 |
|
PIPERACEAE (1/20) |
||||||
|
Piper aduncum L. |
324 (20/20) |
96 (14/20) |
241 (13/20) |
661 |
G |
1, 5 |
|
P. aequale Vahl |
150 (12/20) |
138 (17/20) |
11 (2/20) |
299 |
G |
4 |
|
P. amalago L. |
– |
12 (8/20) |
18 (4/20) |
30 |
G |
6 |
|
P. arboreum Aubl. |
21 (9/20) |
343 (12/20) |
6 (4/20) |
370 |
G |
1, 4 |
|
P. fraguanum Trel. |
– |
81 (12/20) |
– |
81 |
T |
4 |
|
P. hispidum Sw. |
64 (6/20) |
67 (13/20) |
– |
131 |
P |
6, 7 |
|
P. ixocubvainense Standl. et Steyerm. |
– |
– |
7 (1/20) |
7 |
ND |
– |
|
P. lapathifolium (Kunth) Steud. |
– |
4 (2/20) |
14 (4/20) |
18 |
G |
6 |
|
P. obliquum Ruiz et Pav. |
– |
96 (2/20) |
– |
96 |
G |
4 |
|
P. oblongum Kunth |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
P. philodendroides Standl. et Steyerm. † |
1 (1/20) |
– |
– |
1 |
ND |
– |
|
P. phytolaccifolium Opiz † |
7 (3/20) |
– |
26 (7/20) |
33 |
G |
4 |
|
P. psilorhachis C. DC. |
2 (1/20) |
– |
– |
2 |
T |
4 |
|
P. sancti-felicis Trel. |
– |
1 (1/20) |
30 (6/20) |
31 |
G |
4 |
|
P. sanctum (Miq.) Schltdl. ex C. DC. |
– |
11 (6/20) |
– |
11 |
P |
6 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
P. umbellatum L. |
– |
1 (1/20) |
– |
1 |
P |
6 |
|
P. variabile C. DC. |
39 (7/20) |
– |
1 (1/20) |
40 |
T |
4 |
|
P. yzabalanum C. DC. |
53 (11/20) |
4 (2/20) |
2 (1/20) |
59 |
T |
5 |
|
Piper sp. 1 |
– |
– |
2 (1/20) |
2 |
ND |
– |
|
Piper sp. 2 |
– |
– |
5 (1/20) |
5 |
ND |
– |
|
POLYGONACEAE (1/6) |
||||||
|
Coccoloba acuminata Kunth † |
– |
– |
8 (1/20) |
8 |
G |
4, 5 |
|
C. barbadensis Jacq. |
– |
3 (2/20) |
– |
3 |
G |
4 |
|
C. belizensis Standl. |
12 (6/20) |
– |
– |
12 |
G |
4, 5 |
|
C. hondurensis Lundell |
– |
30 (4/20) |
3 (3/20) |
33 |
G |
4 |
|
C.montana Standl. |
– |
7 (4/20) |
– |
7 |
T |
4, 5 |
|
Coccoloba sp. |
– |
– |
4 (1/20) |
4 |
ND |
– |
|
PRIMULACEAE (3/5) |
||||||
|
Ardisia compressa Kunth |
– |
8 (6/20) |
– |
8 |
T |
4 |
|
A. tuerckheimii Donn. Sm. |
– |
11 (6/20) |
– |
11 |
G |
4 |
|
Deherainia smaragdina (Planch. ex Linden) Decne. |
– |
65 (2/20) |
– |
65 |
G |
4 |
|
Parathesis conzattii (S.F. Blake) Lundell |
26 (7/20) |
5 (3/20) |
24 (7/20) |
55 |
T |
7 |
|
P. lenticellata Lundell |
19 (8/20) |
2 (2/20) |
11 (3/20) |
32 |
T |
4, 7 |
|
PROTEACEAE (1/1) |
||||||
|
Roupala montana Aubl. |
– |
3 (2/20) |
– |
3 |
P |
3, 5 |
|
RHIZOPHORACEAE (1/1) |
||||||
|
Cassipourea guianensis Aubl. |
11 (6/20) |
– |
– |
11 |
T |
33 |
|
RUBIACEAE (20/43) |
||||||
|
Alibertia edulis (Rich.) A. Rich. |
171 (18/20) |
3 (1/20) |
82 (12/20) |
256 |
G |
5, 8 |
|
Amaioua corymbosa Kunth |
38 (8/20) |
– |
– |
38 |
G |
3, 5, 8 |
|
Appunia guatemalensis Donn. Sm. |
5 (1/20) |
– |
– |
5 |
T |
4 |
|
Arachnothryx tuxtlensis (Lorence et Cast.-Campos) Borhidi |
– |
3 (1/20) |
– |
3 |
ND |
– |
|
Bertiera guianensis Aubl. |
7 (3/20) |
– |
– |
7 |
T |
4 |
|
Blepharidium guatemalense Standl. |
16 (7/20) |
– |
– |
16 |
G |
3, 5 |
|
Coffea arabica L. |
– |
4 (2/20) |
13 (2/20) |
17 |
I |
– |
|
Faramea occidentalis (L.) A. Rich. |
182 (16/20) |
66 (8/20) |
890 (16/20) |
1138 |
T |
4, 7, 8 |
|
Genipa americana L. |
3 (1/20) |
– |
32 (10/20) |
35 |
G |
1, 3, 8, 34 |
|
Hamelia longipes Standl. |
– |
5 (5/20) |
– |
5 |
T |
4, 7 |
|
H. patens Jacq. |
1 (1/20) |
– |
7 (5/20) |
8 |
P |
1, 3, 5, 7, 8 |
|
Hoffmannia altipetens Dwyer ex Torr.-Montúfar |
– |
1 (1/20) |
– |
1 |
T |
4 |
|
H. discolor (Lem.) Hemsl. |
– |
1 (1/20) |
– |
1 |
T |
3 |
|
H. psychotriifolia (Benth.) Griseb. * |
– |
5 (1/20) |
– |
5 |
T |
4 |
|
Mexocarpus tetragonus (Donn. Sm.) Borhidi, |
146 (18/20) |
84 (17/20) |
133 (18/20) |
363 |
G |
1, 7 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
Morinda panamensis Seem. |
1 (1/20) |
– |
1 (1/20) |
2 |
P |
1, 3, 4 |
|
Palicourea brachiata (Sw.) Borhidi |
231 (17/20) |
– |
101 (6/20) |
332 |
G |
4 |
|
P. deflexa (DC.) Borhidi |
334 (15/20) |
– |
12 (3/20) |
346 |
G |
4 |
|
P. faxlucens (Lorence et Dwyer) Lorence |
– |
21 (5/20) |
– |
21 |
T |
4 |
|
P. gardenioides (Scheidw.) Hemsl. |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
P. sousae (Lorence et Dwyer) Lorence |
– |
49 (1/20) |
– |
49 |
ND |
– |
|
Posoqueria latifolia (Rudge) Schult. |
71 (15/20) |
14 (7/20) |
304 (12/20) |
389 |
G |
4, 8 |
|
Psychotria carthagenensis Jacq. |
90 (13/20) |
– |
7 (3/20) |
97 |
G |
1 |
|
P. costivenia Griseb. |
92 (16/20) |
32 (2/20) |
29 (4/20) |
153 |
G |
4 |
|
P. flava Oerst. ex Standl. |
– |
39 (9/20) |
211 (5/20) |
250 |
G |
4 |
|
P. limonensis K. Krause |
16 (6/20) |
108 (14/20) |
25 (5/20) |
149 |
G |
4, 7 |
|
P. marginata Sw. |
12 (5/20) |
– |
– |
12 |
G |
4 |
|
P. micrantha Kunth |
1 (1/20) |
– |
– |
1 |
G |
4 |
|
P. papantlensis (Oerst.) Hemsl. |
– |
4 (2/20) |
– |
4 |
T |
4 |
|
P. pleuropoda Donn. Sm. |
2 (2/20) |
– |
8 (2/20) |
10 |
T |
4 |
|
P. poeppigiana Müll. Arg. |
579 (14/20) |
– |
13 (1/20) |
592 |
G |
1, 4, 8 |
|
P. sarapiquensis Standl. |
– |
7 (1/20) |
– |
7 |
G |
4 |
|
P. simiarum Standl. |
– |
16 (6/20) |
– |
16 |
T |
4 |
|
P. veracruzensis Lorence et Dwyer |
– |
47 (7/20) |
– |
47 |
G |
4 |
|
Psychotria sp. 1 |
– |
5 (1/20) |
– |
5 |
ND |
– |
|
Psychotria sp. 2 |
2 (1/20) |
– |
– |
2 |
ND |
– |
|
Randia armata (Sw.) DC. |
16 (10/20) |
– |
23 (9/20) |
39 |
G |
4, 5 |
|
R. laetevirens Standl. |
– |
1 (1/20) |
– |
1 |
G |
4, 25 |
|
R. loniceroides Dwyer et Lorence |
– |
1 (1/20) |
– |
1 |
T |
4 |
|
R. pterocarpa Lorence et Dwyer |
– |
19 (7/20) |
– |
19 |
T |
3 |
|
Renistipula galeottii (Standl.) Borhidi |
– |
13 (4/20) |
– |
13 |
T |
4 |
|
Ronabea latifolia Aubl. |
2 (1/20) |
– |
– |
2 |
G |
1, 5 |
|
Simira salvadorensis (Standl.) Steyerm. |
23 (3/20) |
– |
20 (7/20) |
43 |
T |
3, 9 |
|
RUTACEAE (3/9) |
||||||
|
Amyris sp. |
– |
10 (1/20) |
– |
10 |
ND |
– |
|
Amyris sylvatica Jacq. |
– |
20 (1/20) |
– |
20 |
G |
4, 5 |
|
Citrus × limetta Risso |
– |
1 (1/20) |
– |
1 |
I |
– |
|
C. × sinensis (L.) Osbeck |
– |
2 (1/20) |
33 (6/20) |
35 |
I |
– |
|
Citrus sp. |
– |
1 (1/20) |
– |
1 |
ND |
– |
|
Zanthoxylum acuminatum (Sw.) Sw. |
4 (2/20) |
– |
7 (3/20) |
11 |
G |
4 |
|
Z. caribaeum Lam. |
1 (1/20) |
– |
– |
1 |
P |
7 |
|
Z. limoncello Planch. et Oerst. |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
Z. rhoifolium Lam. |
– |
2 (2/20) |
– |
2 |
P |
4 |
|
SALICACEAE (5/7) |
||||||
|
Casearia corymbosa Kunth |
12 (7/20) |
5 (1/20) |
38 (7/20) |
55 |
P |
31 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
C. sylvestris Sw. |
9 (6/20) |
2 (2/20) |
53 (14/20) |
64 |
G |
7, 8 |
|
C. tacanensis Lundell |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
Lunania mexicana Brandegee |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
Pleuranthodendron lindenii (Turcz.) Sleumer |
19 (5/20) |
84 (15/20) |
199 (12/20) |
302 |
G |
7, 9 |
|
Xylosma flexuosa (Kunth) Hemsl. |
– |
1 (1/20) |
– |
1 |
P |
1 |
|
Zuelania guidonia (Sw.) Britton et Millsp. |
5 (5/20) |
– |
2 (1/20) |
7 |
G |
1, 8, 9 |
|
SAPINDACEAE (5/9) |
||||||
|
Allophylus camptostachys Radlk. |
– |
2 (2/20) |
– |
2 |
G |
4, 7 |
|
A. cominia (L.) Sw. |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
Cupania belizensis Standl. |
– |
2 (1/20) |
– |
2 |
G |
4 |
|
C. dentata DC. |
93 (9/20) |
– |
62 (14/20) |
155 |
G |
5, 9 |
|
C. glabra Sw. |
8 (5/20) |
43 (11/20) |
9 (4/20) |
60 |
G |
4 |
|
C. macrophylla Mart. |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
Matayba oppositifolia (A. Rich.) Britton |
10 (4/20) |
– |
– |
10 |
G |
4, 5 |
|
Melicoccus oliviformis Kunth |
14 (9/20) |
– |
33 (9/20) |
47 |
G |
4 |
|
Sapindus saponaria L. |
2 (2/20) |
2 (2/20) |
– |
4 |
P |
1, 5, 8, 9 |
|
SAPOTACEAE (4/14) |
||||||
|
Chrysophyllum mexicanum Brandegee ex Standl. |
32 (9/20) |
5 (5/20) |
– |
37 |
T |
7, 9 |
|
C. venezuelanense (Pierre) T.D. Penn. |
– |
1 (1/20) |
– |
1 |
T |
4 |
|
Manilkara chicle (Pittier) Gilly |
– |
2 (1/20) |
– |
2 |
T |
20 |
|
M. zapota (L.) P. Royen |
3 (1/20) |
– |
– |
3 |
T |
9, 15 |
|
Pouteria campechiana (Kunth) Baehni |
4 (2/20) |
3 (2/20) |
1 (1/20) |
8 |
T |
7, 9 |
|
P. durlandii (Standl.) Baehni |
39 (14/20) |
25 (12/20) |
33 (8/20) |
97 |
G |
20 |
|
P. glomerata (Miq.) Radlk. |
13 (3/20) |
3 (1/20) |
1 (1/20) |
17 |
G |
4 |
|
P. reticulata (Engl.) Eyma |
– |
15 (4/20) |
– |
15 |
T |
7, 8, 9 |
|
P. rhynchocarpa T.D. Penn. |
– |
14 (6/20) |
– |
14 |
T |
4 |
|
P. sapota (Jacq.) H.E. Moore et Stearn |
1 (1/20) |
13 (5/20) |
4 (2/20) |
18 |
T |
7, 9 |
|
Pouteria sp. |
118 (15/20) |
– |
42 (5/20) |
160 |
ND |
– |
|
Sideroxylon capiri (A. DC.) Pittier ** |
– |
3 (1/20) |
– |
3 |
T |
1, 3, 5 |
|
S. persimile (Hemsl.) T.D. Penn. |
– |
1 (1/20) |
– |
1 |
T |
1, 3, 7 |
|
S. portoricense Urb. |
– |
18 (8/20) |
– |
18 |
G |
4 |
|
SIMAROUBACEAE (1/1) |
||||||
|
Quassia amara L. |
39 (6/20) |
– |
– |
39 |
T |
1, 5, 8, 34 |
|
SIPARUNACEAE (1/1) |
||||||
|
Siparuna thecaphora (Poepp. et Endl.) A. DC. |
17 (8/20) |
33 (10/20) |
5 (2/20) |
55 |
P |
6 |
|
SOLANACEAE (2/5) |
||||||
|
Cestrum luteovirescens Francey |
– |
1 (1/20) |
– |
1 |
G |
4 |
|
C. oblongifolium Schltdl. |
– |
10 (3/20) |
– |
10 |
G |
4 |
|
C. racemosum Ruiz et Pav. |
5 (4/20) |
1 (1/20) |
9 (7/20) |
15 |
G |
4, 5, 7 |
|
Solanum nudum Dunal |
– |
1 (1/20) |
– |
1 |
G |
1 |
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
S. schlechtendalianum Walp. |
1 (1/20) |
1 (1/20) |
2 (1/20) |
4 |
P |
1, 20 |
|
STAPHYLEACEAE (1/1) |
||||||
|
Turpinia occidentalis (Sw.) G. Don |
– |
2 (2/20) |
– |
2 |
T |
4, 35 |
|
STYRACACEAE (1/1) |
||||||
|
Styrax glabrescens Benth. |
3 (2/20) |
– |
8 (2/20) |
11 |
T |
4, 5 |
|
SURIANACEAE (1/1) |
||||||
|
Recchia simplicifolia T. Wendt et E.J. Lott |
– |
– |
1 (1/20) |
1 |
T |
4, 36 |
|
SYMPLOCACEAE (1/1) |
||||||
|
Symplocos excelsa L.O. Williams * |
– |
16 (3/20) |
– |
16 |
T |
4 |
|
THYMELAEACEAE (1/1) |
||||||
|
Daphnopsis megacarpa Nevling et Barringer |
– |
7 (3/20) |
– |
7 |
T |
4 |
|
ULMACEAE (1/1) |
||||||
|
Ampelocera hottlei (Standl.) Standl. |
400 (13/20) |
19 (2/20) |
17 (2/20) |
436 |
T |
1, 3, 9 |
|
URTICACEAE (4/6) |
||||||
|
Cecropia peltata L. |
– |
– |
1 (1/20) |
1 |
P |
8, 9, 15 |
|
Myriocarpa longipes Liebm. |
– |
61 (11/20) |
– |
61 |
G |
6 |
|
Pourouma bicolor Mart. |
22 (3/20) |
– |
– |
22 |
G |
8 |
|
Urera caracasana (Jacq.) Gaudich. ex Griseb. |
1 (1/20) |
– |
17 (3/20) |
18 |
P |
5, 20 |
|
U. glabriuscula V.W. Steinm. |
– |
24 (7/20) |
– |
24 |
G |
4 |
|
U. simplex Wedd. |
– |
23 (7/20) |
– |
23 |
G |
3, 4 |
|
VIOLACEAE (3/5) |
||||||
|
Hybanthus sylvicola Standl. et Steyerm. |
43 (8/20) |
– |
– |
43 |
T |
4 |
|
Orthion oblanceolatum Lundell |
– |
42 (6/20) |
– |
42 |
T |
4, 5 |
|
O. subsessile (Standl.) Steyerm. et Standl. |
11 (5/20) |
– |
2 (1/20) |
13 |
T |
4, 5 |
|
Rinorea guatemalensis (S. Watson) Bartlett |
125 (8/20) |
309 (9/20) |
549 (16/20) |
983 |
G |
5, 7 |
|
R. hummelii Sprague |
116 (8/20) |
2 (1/20) |
4 (1/20) |
122 |
G |
4, 5 |
|
VOCHYSIACEAE (1/1) |
||||||
|
Vochysia guatemalensis Donn. Sm. |
124 (13/20) |
26 (6/20) |
10 (6/20) |
160 |
G |
4, 20 |
|
NO IDENTIFICADAS (19) |
||||||
|
Especie 1 |
5 (1/20) |
– |
4 (3/20) |
9 |
||
|
Especie 2 |
1 (1/20) |
– |
3 (3/20) |
4 |
||
|
Especie 3 |
– |
– |
1 (1/20) |
1 |
||
|
Especie 4 |
6 (4/20) |
– |
5 (3/20) |
11 |
||
|
Especie 5 |
12 (3/20) |
– |
– |
12 |
||
|
Especie 6 |
25 (8/20) |
– |
– |
25 |
||
|
Especie 7 |
2 (1/20) |
– |
– |
2 |
||
|
Especie 8 |
9 (5/20) |
– |
– |
9 |
||
|
Especie 9 |
1 (1/20) |
– |
– |
1 |
||
|
Especie 10 |
2 (1/20) |
– |
– |
2 |
||
|
Especie 11 |
1 (1/20) |
– |
– |
1 |
||
|
Apéndice. Continúa |
||||||
|
Taxones |
MC |
LT |
NC |
Total |
Est. Reg. |
Ref. |
|
Especie 12 |
– |
– |
1 (1/20) |
1 |
||
|
Especie 13 |
1 (1/20) |
– |
– |
1 |
||
|
Especie 14 |
– |
– |
51 (6/20) |
51 |
||
|
Especie 15 |
– |
– |
2 (1/20) |
2 |
||
|
Especie 16 |
22 (3/20) |
– |
21 (3/20) |
43 |
||
|
Especie 17 |
– |
– |
1 (1/20) |
1 |
||
|
Especie 18 |
1 (1/20) |
– |
– |
1 |
||
|
Especie 19 |
13 (5/20) |
– |
– |
13 |
Referencias bibliográficas: 1. Flora de Nicaragua; 2. Calvo-Irabién e Islas-Luna (1999); 3. Flora Mesoamericana; 4. Revisión de las descripciones de los especímenes en Flora Mesoamericana y en Trópicos; 5. Standley y Steyermark (1952) (Flora of Guatemala); 6. Popma et al. (1988); 7. Arroyo-Rodríguez, V., material no publicado; 8. Condit et al. (2010); 9. Pennington y Sarukhán (2005); 10. Burns et al. (1998); 11. CICY (2020); 12. Lorenzi (1998); 13. Observación personal; 14. Ortiz-Rodríguez et al. (2015); 15. Dickinson et al. (2000); 16. Ortiz-Rodríguez et al. (2016); 17. Pérez-Farrera et al. (2012); 18. Augspurger (1984); 19. Ray y Brown (1995). 20. Miguel Martínez-Ramos, M., material no publicado; 21. Francis (2004); 22. Bárcena (1981); 23. Rico-Arce et al. (2008); 24. Durán-Fernández et al. (2016); 25. Medina-Abreo y Castillo-Campos (1993); 26. Pérez y Condit (2021); 27. Lorenzi (1992); 28. Almeda (1997). 29. Richards y Coley (2007); 30. Snook (1993); 31. Ramírez-Angulo et al. (2006); 32. Strauss-Debenedetti y Bazzaz (1991); 33. Toriola et al. (1998); 34. Barwick y Van Der Schans (2004); 35. Fleming y Estrada (1993); 36. Wendt y Lott (1985); 37. Martínez-Ramos, M. (1985); 38. Galeano y Olivares (2013); 39. Castro-Rodríguez et al. (2015).
Referencias
Almeda, F. (1997). Cytological and nomenclatural notes on the Mesoamerican species of Aciotis (Melastomataceae). Novon, 7, 333–337. https://doi.org/10.2307/3391757
Anzures-Dadda, A. y Manson, R. H. (2007). Patch- and landscape- scale effects on howler monkey distribution and abundance in rainforest fragments. Animal Conservation, 10, 69–76. https://doi.org/10.1111/j.1469-1795.2006.00074.x
Arbizu, P. M. (2020). PairwiseAdonis: Pairwise multilevel comparison using adonis. R package version 0.4.
Arce-Peña, N. P., Arroyo-Rodríguez, V., Dias, P. A. D., Franch-Pardo, I. y Andresen, E. (2019). Linking changes in landscape structure to population changes of an endangered primate. Landscape Ecology, 34, 2687–2701. https://doi.org/10.1007/s10980-019-00914-8
Arroyo-Rodríguez, V. y Mandujano, S. (2006). The Importance of tropical rain forest fragments to the conservation of plant species diversity in Los Tuxtlas, Mexico. Biodiversity and Conservation, 15, 4159–4179. https://doi.org/10.1007/s105
31-005-3374-8
Arroyo-Rodríguez, V., Mandujano, S. y Benítez-Malvido, J. (2008). Landscape attributes affecting patch occupancy by howler monkeys (Alouatta palliata mexicana) at Los Tuxtlas, Mexico. American Journal of Primatology, 70, 69–77. https://doi.org/10.1002/ajp.20458
Arroyo-Rodríguez, V., Pineda, E., Escobar, F. y Benítez-Malvido, J. (2009). Value of small patches in the conservation of plant-species diversity in highly fragmented rainforest. Conservation Biology, 23, 729–739. https://doi.org/10.1111/j.1523-1739.2008.01120.x
Arroyo-Rodríguez, V., Rös, M., Escobar, F., Melo, F. P. L., Santos, B. A., Tabarelli, M. et al. (2013). Plant β-diversity in fragmented rain forests: testing floristic homogenization and differentiation hypotheses. Journal of Ecology, 101, 1449–1458. https://doi.org/10.1111/1365-2745.12153
Arroyo-Rodríguez, V., Melo, F. P. L., Martínez-Ramos, M., Bongers, F., Chazdon, R., Meave, J. A. et al. (2017). Multiple successional pathways in human-modified tropical landscapes: New insights from forest succession, forest fragmentation and landscape ecology research. Biological Reviews, 92, 326–340. https://doi.org/10.1111/brv.12231
Arroyo-Rodríguez V., Fahrig, L., Tabarelli, M., Watling, J.I., Tischendorf, L., Benchimol, M. et al. (2020). Designing optimal human-modified landscapes for forest biodiversity conservation. Ecology Letters, 23, 1404–1420. https://doi.org/10.1111/ele.13535
Augspurger, C. K. (1984). Seedling survival of tropical tree Species: interactions of dispersal distance, light-gaps and pathogens. Ecology, 65, 1705–1712. https://doi.org/
10.2307/1937766
Balvanera, P., Pfisterer, A. B., Buchmann, N., He, J-S., Nakashizuka, T., Raffaelli, D. et al. (2006). Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters, 9, 1146–1156. https://doi.org/10.1111/j.1461-0248.2006.00963.x
Bárcena, A. (1981). Clethraceae. En A. Gómez-Pompa, V. Sosa, L. I. Nevling Jr, M, Nee, N. P. Moreno y B. Ludlow-Wiechers (Eds.), Flora de Veracruz (pp. 1–18). Xalapa, Veracruz: Instituto Nacional de Investigaciones sobre Recursos Bióticos.
Barwick, M. y Van Der Schans, A. (2004). Tropical and subtropical trees: a worldwide encyclopaedic guide. London: Thames et Hudson.
Breugel, M. van, Martínez-Ramos, M. y Bongers, F. (2006). Community dynamics during early secondary succession in Mexican tropical rain forests. Journal of Tropical Ecology, 22, 663–674. https://doi.org/10.1017/S0266467406003452
Bugmann, H. (1996). Functional types of trees in temperate and boreal forests: Classification and testing. Journal of Vegetation Science, 7, 359–70. https://doi.org/10.2307/3236279
Burelo-Ramos, C. M., Ortiz-Gil, G. y Almeida-Cerino, C. M. (2009). Notas sobre el género Bactris (Arecaceae) en el estado de Tabasco, México. Kuxulkab’, 29, 83–87. https://doi.org/10.19136/kuxulkab.a16n29.433
Burns, R. M., Mosquera, M. S., Whitmore, J. L. (1998). Useful trees of the tropical region of North America. Washington: North American Forestry Commission publication.
Calvo-Irabién, L. M. e Islas-Luna, A. (1999). Predispersal predation of an understory rainforest herb Aphelandra aurantiaca (Acanthaceae) in gaps and mature forest. Americal Journal of Botany, 86, 1108–1113. https://doi.org/10.2307/2656973
Carabias, J., De la Maza, J. y Cadena, R. (2015). Conservación y desarrollo sustentable en la Selva Lacandona: 25 años de actividades y experiencias. Ciudad de México: Natura y Ecosistemas Mexicanos, A.C.
Carrara, E., Arroyo-Rodríguez, V., Vega-Rivera, J. H., Schondube, J. E., Freitas, S. M. y Fahrig, L. (2015). Impact of landscape composition and configuration on forest specialist and generalist bird species in the fragmented Lacandona rainforest, Mexico. Biological Conservation, 184, 117–126. https://doi.org/10.1016/j.biocon.2015.01.014
Castillo-Campos, G. y Laborde, J. (2004). La vegetación. En S. Guevara, J. Laborde y G. Sánchez-Ríos (Eds.), Los Tuxtlas. El paisaje de la sierra (pp. 231–265). Xalapa: Instituto de Ecología y Unión Europea.
Castro-Rodríguez, S. Y., Barrera-García, J. A., Carrillo-Bautista, M. P. y Hernández-Gómez, M. S. (2015). Asaí (Euterpe precatoria): cadena de valor en el sur de la región amazónica. Bogotá: Instituto Amazónico de Investigaciones Científicas- Sinchi.
Cayuela L., Gálvez-Bravo, L., de Albuquerque, F. S., Golicher, D. J., González-Espinosa M., Ramírez-Marcialet, N. et al. (2012). La Red Internacional de Inventarios Forestales (BIOTREE-NET) en Mesoamérica: avances, retos y perspectivas futuras. Ecosistemas, 21, 126–135.
CICY (Centro de Investigación Científica de Yucatán). (2020). Flora de la península de Yucatán. Recuperado el 13 de junio, 2021 de: https://www.cicy.mx/sitios/flora%20digital/index.php
Chazdon, R. L. (2014). Second growth: the promise of Tropical Forest regeneration in an age of deforestation. Chicago, Illinois: The University of Chicago Press.
Chazdon, R. L. y Guariguata, M. R. (2016). Natural regeneration as a tool for large-scale forest restoration in the tropics: prospects and challenges. Biotropica, 48, 716–730. https://doi.org/10.1111/btp.12381
Collen, B., Ram, M., Zamin, T. y McRae, L. (2008). The tropical biodiversity data gap: addressing disparity in global monitoring. Tropical Conservation Science, 1, 75–88. https://doi.org/10.1177/194008290800100202
Conabio y Semarnat (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad y Secretaría de Medio Ambiente y Recursos Naturales. (2009). Cuarto Informe Nacional de México al Convenio sobre Diversidad Biológica (CDB). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad y Secretaría de Medio Ambiente y Recursos Naturales.
Conafor (Comisión Nacional Forestal). (2020). Apoyos Conafor. Recuperado el 13 de junio, 2021 de: https://www.gob.mx/conafor/acciones-y-programas/apoyos-conafor
Conanp (Comisión Nacional de Áreas Naturales Protegidas). (2011). Actualización de la tasa de cambio del uso del suelo en la Reserva de la biosfera de Los Tuxtlas, Informe Final. Proyecto Sierra de Santa Marta, A. C. Recuperado el 13 de junio, 2021 de: https://simec.conanp.gob.mx/pdf_evaluacion/tuxtlas.pdf.
Condit, R., Pitman, N., Leigh, E. G., Chave, J., Terborgh, J., Foster, R. B. et al. (2002). Beta-diversity in tropical forest trees. Science, 295, 666–669. https://doi.org/10.1126/science.1066854
Condit, R., Pérez, R. y Daguerre, N. (2010). Trees of Panama and Costa Rica. Princeton y Oxford: Princeton University Press.
Cordeiro, N. J. y Howe, H. F. (2001). Low Recruitment of trees dispersed by animals in African forest fragments. Conservation Biology, 15, 1733–1741. https://doi.org/10.1046/j.1523-1739.
2001.99579.x
Cowan, C. P. (1983). Flora de Tabasco. Ciudad de México: Instituto de Biología, UNAM.
Davidse, G., Sousa Sánchez, M., Knapp, S. y Chiang Cabrera, F. (2020). Cycadaceae a Connaraceae. En G. Davidse, M. Sousa Sánchez, S. Knapp y F. Chiang Cabrera (Eds.), Flora Mesoamericana. St. Louis: Missouri Botanical Garden.
De Cáceres, M., Legendre, P., Valencia, R., Cao, M., Chang, L-W., Chuyong, G. et al. (2012). The variation of tree beta diversity across a global network of forest plots. Global Ecology and Biogeography, 21, 1191–1202. https://doi.org/10.1111/j.1466-8238.2012.00770.x
De Jong, B. H. J., Ochoa-Gaona S., Castillo-Santiago, M. A., Ramírez-Marcial, N. y Cairns, M. A. (2000). Carbon flux and patterns of land-use/land-cover change in the Selva Lacandona, Mexico. Ambio, 29, 504–511. https://doi.org/
10.1579/0044-7447-29.8.504
Dickinson, M. B., Whigham, D. F. y Hermann, S. M. (2000). Tree regeneration in felling and natural treefall disturbances in a semideciduous tropical forest in Mexico. Forest Ecology and Management, 134, 137–151. https://doi.org/10.1016/S0378-1127(99)00252-2
Dirzo, R. y Miranda, A. (1991). Altered patterns of herbivory and diversity in the forest understory: a case of study of the possible consequences of contemporary defaunation. En P. W. Price, T. M. Lewinsohn, G. W. Fernandes y W. W. Benson (Eds.), Plant-animal interactions: evolutionary ecology in tropical and temperate regions (pp. 273–287). Nueva York: John Wiley et Sons.
Dirzo, R. y García, M. C. (1992). Rates of deforestation in Los Tuxtlas, a Neotropical area in Veracruz, Mexico. Conservation Biology, 6, 84–90. https://doi.org/10.1046/
j.1523-1739.1992.610084.x
Durán-Fernández, A., Aguirre-Rivera, J. R., García-Pérez, J., Levy-Tacher, S. y Nova-Vázquez, J. A. (2016). Inventario florístico de la comunidad lacandona de Nahá, Chiapas, México. Botanical Sciences, 94, 157–184. https://doi.org/10.17129/botsci.248
Fahrig, L. (2020). Why do several small patches hold more species than few large patches? Global Ecology and Biogeography, 29, 615–628. https://doi.org/10.1111/geb.13059
Fazey, I., Fischer, J. y Lindenmayer, D. B. (2005). What do conservation biologists publish? Biological Conservation, 124, 63–73. https://doi.org/10.1016/j.biocon.2005.01.013
Fleming, T. H. y Estrada, A. (1993). Frugivory and seed dispersal: ecological and evolutionary aspects. Dordrecht: Kluwer Academic Publishers.
Francis, J. K. (2004). Wildland shrubs of the United States and its territories: Thamnic descriptions. Volume 1. San Juan, Puerto Rico: Gen. Tech. Rep. IITF-GTR-26.
Galán-Acedo, C., Arroyo-Rodríguez, V., Estrada, A. y Ramos-Fernández, G. (2018). Drivers of the spatial scale that best predict primate responses to landscape structure. Ecography, 41, 2027–2037. https://doi.org/10.1111/ecog.03632
Galeano, G. y Olivares, I. (2013). Palma de vino (Attalea butyracea). En R. Bernal y G. Galeano (Eds.), Cosechar sin destruir. Aprovechamiento sostenible de palmas colombianas (pp. 165–174). Bogotá: Universidad Nacional de Colombia.
Garmendia, A., Arroyo-Rodríguez, V., Estrada, A., Naranjo E. J. y Stoner, K. E. (2013). Landscape and patch attributes impacting medium- and large-sized terrestrial mammals in a fragmented rain forest. Journal of Tropical Ecology, 29, 331–344. https://doi.org/10.1017/S0266467413000370
Gobierno de Chiapas (2013). Informe de Gobierno, Región VIII-Norte. Recuperado el 13 de junio, 2021 de: http://www.ceieg.chiapas.gob.mx/productos/files/MAPASTEMREG/REGION_VIII_NORTE_Post.pdf
Gómez-Domínguez, H., Pérez-Farrera, M. A., Espinosa-Jiménez, J. A. y Márquez-Reynoso, M. I. (2015). Listado florístico del Parque Nacional Palenque, Chiapas, México. Botanical Sciences, 93, 559–578. https://doi.org/10.17129/botsci.151
González-Christen, A. y Coates, R. (2019). Los mamíferos no voladores de la región de Los Tuxtlas, Veracruz, México. Revista Mexicana de Biodiversidad, 90, e902580. http://dx.doi.org/10.22201/ib.20078706e.2019.90.2580
Guevara, S. y Laborde, J. (2012). The Mesoamerican rain forest environmental history. Livestock and landscape biodiversity at Los Tuxtlas, Mexico. Pastos, 42, 219–248.
Hansen, M. C., Wang, L., Song, X-P., Tyukavina, A., Turubanova, S., Potapov, P. V. et al. (2020). The fate of tropical forest fragments. Science Advances, 6, eaax8574. https://doi.org/10.1126/sciadv.aax8574
Hernández-Ruedas, M. A., Arroyo-Rodríguez, V., Meave, J. A., Martínez-Ramos, M., Ibarra-Manríquez, G., Martínez, E. et al. (2014). Conserving tropical tree diversity and forest structure: The value of small rainforest patches in moderately-managed landscapes. Plos One, 9, e98931. https://doi.org/10.1371/journal.pone.0098931
Hill, J. L. y Curran, P. J. (2003). Area, shape and isolation of tropical forest fragments: effects on tree species diversity and implications for conservation. Journal of Biogeography, 30, 1391–1403. https://doi.org/10.1046/j.1365-2699.2003.
00930.x
Hubbell, S. P. (2013). Tropical rain forest conservation and the twin challenges of diversity and rarity. Ecology and Evolution, 3, 3263–3274. https://doi.org/10.1002/ece3.705
Ibarra-Manríquez, G. y Sinaca-Colín, S. (1995). Lista florística comentada de la Estación de Biología Tropical “Los Tuxtlas”, Veracruz, México. Revista de Biología Tropical, 43, 75-115.
INEGI (Instituto Nacional de Estadística, Geografía e Informática). (1999). Juárez, estado de Chiapas. Cuaderno estadístico municipal. Mexico D.F.: Instituto Nacional de Estadística, Geografía e Informática.
INEGI (Instituto Nacional de Estadística, Geografía e Informática). (2014). Información de uso del suelo y vegetación. Escala 1:250000. Recuperado el 13 de junio, 2021 de: https://www.inegi.org.mx/temas/bosques/default.html#Tabulados
Joppa, L. N., Roberts, D. L. y Pimm, S. L. (2011). How many species of flowering plants are there? Proceedings of the Royal Society B, 278, 554–559. https://doi.org/10.1098/rspb.
2010.1004
Jost, L. (2006). Entropy and diversity. Oikos, 113, 363–375. https://doi.org/10.1111/j.2006.0030-1299.14714.x
Jost, L. (2010). The relation between evenness and diversity. Diversity, 2, 207–232. https://doi.org/10.3390/d2020207
Larson, J. E. y Funk, J. L. (2016). Regeneration: an overlooked aspect of trait-based plant community assembly models. Journal of Ecology, 104, 1284–1298. https://doi.org/10.1111
/1365-2745.12613
León, B. y Martínez-Gordillo, M. (2008). Especie nueva de Croton sección Eluteria (Euphorbiaceae) de Chiapas, México. Novon, 18, 189–191. https://doi.org/10.3417/2006097
Levy-Tacher, S., Aguirre-Rivera, J. R., García-Perez, J. D. y Martínez-Romero, M. M. (2006). Aspectos florísticos de Lacanhá Chansayab, Selva Lacandona, Chiapas. Acta Botanica Mexicana, 77, 69–98. https://doi.org/10.21829/abm77.2006.1026
Lôbo, D., Leão, T., Melo, F. P. L., Santos, A. M. M. y Tabarelli, M. (2011). Forest fragmentation drives Atlantic forest of northeastern Brazil to biotic homogenization. Diversity and Distributions, 17, 287–296. https://doi.org/10.1111/j.1472-4642.2010.00739.x
Lorenzi, H. (1992). Brazilian trees, Volume 1. A guide to the identification and cultivation of Brazilian native trees. Nova Odessa: Instituto Plantarum de Estudos da Flora.
Lorenzi, H. (1998). Brazilian trees, Volume 2. A guide to the identification and cultivation of Brazilian native trees. Nova Odessa: Instituto Plantarum de Estudos da Flora.
Marcon, E. y Herault, B. (2015). entropart: An R package to measure and partition diversity. Journal of Statistical Software, 67, 1–26. https://doi.org/10.18637/jss.v067.i08
Martínez, E., Ramos, C. H. A. y Chiang, F. (1994). Lista florística de la Lacandona, Chiapas. Boletín de la Sociedad Botánica de México, 54, 99–177. https://doi.org/10.17129/botsci.1430
Martínez-Ramos, M. (1985). Claros, ciclos de vida de los árboles tropicales y la regeneración natural de las selvas altas perennifolias. En A. Gómez-Pompa y S. del Amo (Eds.), Investigaciones sobre la regeneración de selvas altas en Veracruz II (pp. 191–239). México: Alhambra.
Martínez-Ramos, M. (1994). Regeneración natural y diversidad de especies arbóreas en selvas húmedas. Boletín de la Sociedad Botánica de México, 54, 179–224. https://doi.org/10.17129/botsci.1431
Meave, J. A., Romero-Romero, M. A., Valle-Doménech, A., Rincón-Gutiérrez, A., Martínez, E. y Ramos, C. H. (2008). Plant diversity assessment in the Yaxchilán Natural Monument, Chiapas, Mexico. Boletín de la Sociedad Botánica de México, 83, 53–76. https://doi.org/10.17129/botsci.1788
Medina-Abreo, M. E. y Castillo-Campos, G. (1993). Vegetation and checklist of the Barranca de Acazónica, Veracruz, Mexico. Botanical Sciences, 53, 73–111. https://doi.org/10.
17129/botsci.1417
Morante-Filho, J. C., Faria, D., Mariano-Neto, E. y Rhodes, J. (2015). Birds in anthropogenic landscapes: The responses of ecological groups to forest loss in the Brazilian Atlantic Forest. Plos One, 10, e0128923. https://doi.org/10.1371/journal.pone.0128923
Ortiz-Rodríguez, A. E., Sosa, V. y Ruiz-Sánchez, E. (2015). Flora del Bajío y de regiones adyacentes. Annonaceae. Pátzcuaro, Michoacán: Instituto de Ecología A.C. https://doi.org/10.21829/fb.54.2015.191
Ortiz-Rodríguez, A. E., Burelo Ramos, C. M. y Gomez-Dominguez, H. (2016). A new species of Amphitecna (Bignoniaceae) endemic to Chiapas, Mexico. Phytokeys, 65, 15–23. https://doi.org/10.3897/phytokeys.65.8454
Oyama, K. (1987). Demografía y dinámica poblacional de Chamaedorea tepejilote Liebm. en la selva de Los Tuxtlas, Veracruz (México) (Tesis de maestría). Facultad de Ciencias, UNAM. Ciudad de México.
Peña-Domene, M., Martínez-Garza, C., Ayestarán-Hernández, L. M. y Howe, H. F. (2018). Plant attributes that drive dispersal and establishment limitation in tropical agricultural landscapes. Forests, 9, 620. https://doi.org/10.3390/f9100620
Pennington, T. D. y Sarukhán, J. (2005). Árboles tropicales de México. Manual para la identificación de las principales especies. Ciudad de México: Ediciones Científicas Universitarias.
Pérez, R. y Condit, R. (2021). Tree Atlas of Panama. Recuperado el 13 de junio, 2021 de: http://ctfs.si.edu/PanamaAtlas/maintreeatlas.php
Pérez-Farrera, M. A., Martínez-Camilo, R., Martínez-Meléndez, N., Farrera-Sarmiento, O. y Maza-Villalobos, S. (2012). Listado florístico del Cerro Quetzal (Polígono III) de la Reserva de la Biosfera El Triunfo, Chiapas, México. Botanical Sciences, 90, 113–142.
Piñero, D., Sarukhán, J. y González, E. (1977). Estudios demográficos en plantas. Astrocaryum mexicanum Liebm. 1. Estructura de las poblaciones. Botanical Sciences, 37, 69–118. https://doi.org/10.17129/botsci.1164
Poorter, L., Bongers, F., Kouamý, F. N. y Hawthorne, W. D. (2004). Biodiversity of West African forests: an ecological atlas of woody plant species. Oxon y Cambridge: Cabi Publishing.
Popma, J., Bongers, F., Martínez-Ramos, M. y Veneklaas, E. (1988). Pioneer species distribution in treefall gaps in Neotropical rain forest; a gap definition and its consequences. Journal of Tropical Ecology, 4, 77–88. https://doi.org/
10.1017/S0266467400002534
Prance, G. T., Beentje, H., Dransfield, J. y Johns, R. (2000). The tropical flora remains undercollected. Annals of the Missouri Botanical Garden, 87, 67–71. https://doi.org/
10.2307/2666209
Quero, H. J. y Pérez-Farrera, M. A. (2010). El género Geonoma (Arecaceae: Arecoideae) en México. Revista Mexicana de Biodiversidad, 81, 231–237. https://doi.org/10.22201/ib.20078706e.2010.002.250
R Core Team. (2013). R: A language and environment for statistical computing, versión 3.4.3. R Foundation for Statistical Computing, Vienna, Austria.
Ray, G. J. y Brown, B. J. (1995). Restoring Caribbean dry forests: Evaluation of tree propagation techniques. Restoration Ecology, 3, 86–94. https://doi.org/10.1111/j.1526-100X.19
95.tb00081.x
Renaissance Oil Corp. (2017). Manifiesto de Impacto Ambiental Regional (MIA-R) para las áreas contractuales 11 (Campo Malva), 15 (Campo Mundo Nuevo) y 25 (Campo Topén). Recuperado el 13 de junio, 2021 de: http://104.209.210.233/gobmx/2018/2do_T/A73/FI_201806/d/R-09-DLA0605-10-17-DGGEERC.pdf
Richards, L. A. y Coley, P. D. (2007). Seasonal and habitat differences affect the impact of food and predation on herbivores: a comparison between gaps and understory of tropical forest. Oikos, 116, 31–40. https://doi.org/10.1111/
j.2006.0030-1299.15043.x
Rico-Arce, M. L., Gale, S. L. y Maxted, N. (2008). A taxonomic study of Albizia (Leguminosae: Mimosoideae: Ingeae) in Mexico and Central America. Anales del Jardín Botánico de Madrid, 65, 255–305. https://doi.org/10.3989/ajbm.2008.v65.i2.294
Romaní-Cortés, J. (2011). De necedades a necesidades. Anatomía de un conflicto agrario-ambiental en la zona norte de la Lacandona. México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Russildi, G., Arroyo-Rodríguez, V., Hernández-Ordóñez, O., Pineda, E. y Reynoso, V. H. (2016). Species- and community-level responses to habitat spatial changes in fragmented rainforests: assessing compensatory dynamics in amphibians and reptiles. Biodiversity and Conservation, 25, 375–392. https://doi.org/10.1007/s10531-016-1056-3
San José, M., Arroyo-Rodríguez, V. y Meave, J. A. (2020). Regional context and dispersal mode drive the impact of landscape structure on seed dispersal. Ecological Applications, 30, e02033. https://doi.org/10.1002/eap.2033
Santos, B. A., Peres, C. A., Oliveira, M., Grillo, A., Alves-Costa, C. P. y Tabarelli, M. (2008). Drastic erosion in functional attributes of tree assemblages in Atlantic forest fragments of northeastern Brazil. Biological Conservation, 141, 249–260. https://doi.org/10.1016/j.biocon.2007.09.018
Slik, J. W. F., Arroyo-Rodríguez, V., Aiba, S-I., Alvarez-Loayza, P., Alves, L. F., Ashton, P. et al. (2015). An estimate of the number of tropical tree species. PNAS, 112, 7472–7477. https://doi.org/10.1073/pnas.1423147112
Snook, L. K. (1993). Stand dynamics of mahogany (Swietenia macrophylla King) and associated species after fire and hurricane in the tropical forests of the Yucatán Peninsula, Mexico (Tesis doctoral). Yale University, Yale.
Solano-Gómez, R., Jiménez Machorro, R. y Damon, A. A. (2011). Two new records and one rediscovery for Orchidaceae of Mexico. Acta Botanica Mexicana, 96, 59–72. https://doi.org/10.21829/abm96.2011.259
Song, X. P., Hansen, M. C., Stehman, S. V., Potapov, P. V., Tyukavina, A., Vermote, E. F. et al. (2018). Global land change from 1982 to 2016. Nature, 560, 639–643. https://doi.org/10.1038/s41586-018-0411-9
Standley, P. C. y Steyermark, J. A. (1952). Flora of Guatemala. Chicago: Chicago Museum of Natural History.
Strauss-Debenedetti, S. y Bazzaz, F. A. (1991). Plasticity and acclimation to light in tropical Moraceae of different sucessional positions. Oecologia, 87, 377–387. https://doi.org/10.1007/BF00634595
Stride, G., Thomas, C. D., Benedick, S., Hodgson, J. A., Jelling, A., Senior, M. J. M. et al. (2018). Contrasting patterns of local richness of seedlings, saplings, and trees may have implications for regeneration in rainforest remnants. Biotropica, 50, 889–897. https://doi.org/10.1111/btp.12605
Tabarelli, M., Peres, C. A. y Melo, F. P. L. (2012). The ‘few winners and many losers’ paradigm revisited: Emerging prospects for tropical forest biodiversity. Biological Conservation, 155, 136–40. https://doi.org/10.1016/j.biocon.
2012.06.020
Taubert, F., Fischer, R., Groeneveld, J., Lehmann, S., Müller, M.S., Rödig, E. et al. (2018). Global patterns of tropical forest fragmentation. Nature, 554, 519–522. https://doi.org/10.1038/nature25508
Toriola, D., Chareyre, P. y Buttler, A. (1998). Distribution of primary forest plant species in a 19-year old secondary forest in French Guiana. Journal of Tropical Ecology, 14, 323–340. https://doi.org/10.1017/S026646749800025X
Tropicos. (2020). Flora Mesoamericana. Jardín Botánico de Missouri. Recuperado el 13 de junio, 2021 de: http://legacy.tropicos.org/Project/FM
Tropicos. (2020). Flora de Nicaragua. Jardín Botánico de Missouri. Recuperado el 13 de junio, 2021 de: http://legacy.tropicos.org/Project/FN
Vázquez-García, J. A., Gómez-Domínguez, H., López-Cruz, A., Espinosa-Jiménez, J. A., Sahagún-Godínez, E. y Muñiz-Castro, M. A. (2013). Magnolia perezfarrerae, a new species and a key to Mexican species of Magnolia (section Talauma, subsection Talauma, Magnoliaceae). Botanical Sciences, 91, 417–425. https://doi.org/10.17129/botsci.38
Vázquez-García, J. A., Pérez-Farrera, M. A., Martínez-Camilo, R., Muñiz-Castro, M. A. y Martínez-Meléndez, N. (2013). Magnolia lacandonica (subsection Talauma, Magnoliaceae), a new rainforest species from Chiapas, Mexico. Phytotaxa, 79, 30–36. https://doi.org/10.11646/phytotaxa.79.1.2
Vázquez-Negrín, I., Castillo-Acosta, O., Valdez-Hernández, J. I., Zavala-Cruz, J. y Martínez-Sánchez, J. L. (2011). Estructura y composición florística de la selva alta perennifolia en el ejido Niños Héroes Tenosique, Tabasco, México. Polibotánica, 32, 41–61.
Villaseñor, J. L., Meave, J. A., Ortiz, E. y Ibarra-Manríquez, G. (2003). Biogeografía y conservación de los bosques tropicales húmedos de México. En J. J. Morrone y J. Llorente (Ed.). Una perspectiva latinoamericana de la biogeografía (pp. 209–216). México, D.F.: Universidad Nacional Autónoma de México.
Villaseñor, J. L. y Ortiz, E. (2014). Biodiversidad de las plantas con flores (División Magnoliophyta) en México. Revista Mexicana de Biodiversidad, Supl. 85, S134–S142. https://doi.org/10.7550/rmb.31987
Villaseñor, J. L. (2016). Checklist of the native vascular plants of Mexico. Revista Mexicana de Biodiversidad, 87, 559–902. https://doi.org/10.1016/j.rmb.2016.06.017
Villaseñor, J. L., Ortiz, E. y Campos-Villanueva, A. (2018). High richness of vascular plants in the tropical Los Tuxtlas Region, Mexico. Tropical Conservation Science, 11, 1–12. https://doi.org/10.1177/1940082918764259
Watling, J. I., Arroyo-Rodríguez, V., Pfeifer, M., Baeten, L., Banks-Leite, C., Cisneros, L. M. et al. (2020). Support for the habitat amount hypothesis from a global synthesis of species density studies. Ecology Letters, 23, 674–681. https://doi.org/10.1111/ele.13471
Wendt, T. y Lott, E. J. (1985). A new simple-leaved species of Recchia (Simaroubaceae) from southeastern Mexico. Brittonia, 37, 219–225. https://doi.org/10.2307/2806115


