Importancia de las interacciones semilla-mamífero para Heteroflorum (Leguminosae), un género monoespecífico endémico de México
Lina Adonay Urrea-Galeano, Ellen Andresen y Guillermo Ibarra-Manríquez *
Instituto de Investigaciones en Ecosistemas y Sustentabilidad, Universidad Nacional Autónoma de México, Antigua Carretera a Pátzcuaro No. 8701, Ex-Hacienda de San José de La Huerta, C.P. 58190, Morelia, Michoacán, México
* Autor para correspondencia: gibarra@cieco.unam.mx (G. Ibarra-Manríquez)
Resumen
Se identificaron algunas interacciones entre los frutos/semillas de Heteroflorum sclerocarpum (Leguminosae) y los mamíferos terrestres de un bosque tropical caducifolio de México. Usando cámaras trampa se detectaron los mamíferos que consumen los frutos y/o semillas de esta especie, y mediante observaciones en cautiverio y campo se determinó cómo los mamíferos manipulan estas estructuras y podrían influir en su dispersión. Se hicieron pruebas de germinación en campo y casa sombra para determinar si las semillas podían germinar estando fuera de los frutos, en el interior de frutos intactos o luego de haber sido manipulados por mamíferos. En el laboratorio se colocaron a germinar semillas escarificadas mecánica o químicamente por algunos consumidores. En 3 localidades se determinó la distribución espacial de los individuos de H. sclerocarpum, así como la proporción de individuos dispersados y no dispersados. Registramos interacciones entre los frutos y semillas de H. sclerocarpum y algunos mamíferos terrestres (coatíes, cuiniques, ratones, vacas y venados) durante la época de fructificación, favoreciéndose la liberación de las semillas de los frutos, y permitiendo y/o acelerando la germinación. Un conocimiento más adecuado de las interacciones planta-animal contribuye al entendimiento de la estructura y funcionamiento de los bosques tropicales y permite el desarrollo de estrategias adecuadas para su conservación.
Palabras clave:
Bosque tropical seco; Destino de semillas; Dispersión de semillas; Distribución espacial; Escarificación de semillas; Germinación
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Importance of the seed-mammal interaction for Heteroflorum (Leguminosae), a monospecific genus endemic to Mexico
Abstract
We identified some interactions between fruits/seeds of Heteroflorum sclerocarpum (Leguminosae) and terrestrial mammals of a Mexican tropical dry forest. The mammals that consume the fruits and seeds of this species were detected using camera traps, and by observations in captivity and field, it was determined how mammals manipulate these structures and could influence their dispersion. Germination tests were carried out in the field and shade house to determine if seeds could germinate being outside the fruits, or inside intact fruits, or after having been manipulated by mammals. In the laboratory, we placed to germinate seeds mechanical or chemically scarified by some consumers. In 3 locations the spatial distribution of H. sclerocarpum individuals was determined, as well as the proportion of dispersed and undispersed individuals. We recorded interactions between the fruits and seeds of this species and some terrestrial mammals (cows, coatis, deer, ground squirrels and mice) during the fruiting season, which favored the release of seeds from the fruits, allowing and/or accelerating germination. A good knowledge of plant-animal interactions contributes to the understanding of the structure and functioning of tropical forests, and allows the development of adequate strategies for their conservation.
Keywords:
Tropical dry forest; Seed fate; Seed dispersal; Spatial distribution; Scarification; Germination
Introducción
Es ampliamente conocido que en los bosques tropicales hay un alto número de especies vegetales que desarrollan frutos adaptados para el consumo animal (Jordano, 2014), así como un gran número de especies animales (e.g., aves y mamíferos) que incluyen frutos dentro de su dieta (Fleming y Kress, 2013). Esta disponibilidad y consumo de alimento permite el establecimiento de interacciones planta-animal, las cuales son importantes para el funcionamiento y mantenimiento de los bosques tropicales. Como resultado de estas interacciones, las especies implicadas pueden verse no afectadas, perjudicadas o beneficiadas, reflejando, en este último caso, los efectos positivos en la supervivencia, crecimiento y/o reproducción de los individuos (del Val y Boege, 2012).
Un tipo de interacción cuyo resultado puede tener diversas consecuencias positivas para las plantas es la establecida entre semillas y mamíferos, la cual puede influir de las siguientes maneras: i) mediante la dispersión efectiva de semillas se permite el escape de una alta mortalidad cerca de los árboles madre, la llegada determinística de las semillas a micrositios que favorecen la superviviencia y establecimiento, y/o la colonización aleatoria de nuevos hábitats, favoreciendo el crecimiento demográfico de la planta (Howe y Smallwood, 1982); ii) durante la manipulación de los frutos en la boca o tracto digestivo de los animales, se puede reducir o eliminar la dormancia de las semillas, lo que favorece su germinación (Traveset et al., 2007); iii) de acuerdo al comportamiento de los animales, las semillas son depositadas en patrones espaciales que afectan la distribución espacial de los individuos reclutados (Fragoso et al., 2003; Russo et al., 2006).
En la medida que se cuente con un buen entendimiento de las interacciones entre plantas y animales y sus consecuencias, se pueden desarrollar estrategias de manejo y conservación de los bosques tropicales. Esto puede ser de particular importancia para especies que se encuentren en alguna categoría de vulnerabilidad, como puede llegar a ser el caso de las especies endémicas. Los bosques tropicales secos de México cuentan con una alta diversidad de animales y plantas, así como un alto grado de endemismo (Ceballos y Valenzuela, 2010; Linares-Palomino et al., 2011). Se calcula que aproximadamente el 5% de los géneros y 45-60% de las especies del bosque tropical seco son endémicos del país (Lott y Atkinson, 2010; Trejo, 1998; Villaseñor y Ortiz, 2014). Una especie arbórea que representa este endemismo es Heteroflorum sclerocarpum M. Sousa (Leguminosae), un género monoespecífico cuya distribución se restringe a la cuenca baja del río Balsas y a la planicie costera del Pacífico, desde el nivel del mar y hasta 600 m de altitud (Salas-Morales et al., 2007; Sousa, 2005).
No obstante, diversos aspectos de la biología básica de Heteroflorum sclerocarpum (de aquí en adelante, la especie se nombrará únicamente mencionando el género), son poco conocidos y en particular, aquellos relacionados con su regeneración natural. El poco conocimiento que se tiene es información anecdótica proporcionada por los campesinos, quienes describen que los frutos de Heteroflorum permanecen durante largos periodos en la copa de los árboles (Sr. Rojas-López, com. pers.). Sin embargo, se desconoce qué animales consumen sus frutos, cómo se dispersa la especie o qué factores favorecen la germinación de las semillas en el campo, dada la naturaleza indehiscente de las vainas. Los ejidatarios consideraban que posiblemente las vacas, que pastorean libremente en el bosque, podrían estar dispersando las semillas, debido a que observaban plántulas en excretas de vacas, lejos de árboles con frutos. Sin embargo, se debe considerar que la fauna silvestre también está presente y podría estar desempeñando algún papel importante en las primeras fases de la regeneración de la especie.
Por lo tanto, el objetivo general de este estudio fue documentar aspectos de la historia natural de Heteroflorum, identificando posibles interacciones durante la época de fructificación con mamíferos terrestres que la consumen. Se consideraron los siguientes objetivos particulares: i) determinar las especies de mamíferos terrestres que consumen los frutos y/o semillas de esta especie y describir cómo manipulan estas estructuras, ii) evaluar el destino de los frutos que, por gravedad, caen debajo de las copas de los árboles madre, iii) determinar si las semillas pueden germinar estando fuera de los frutos, en el interior de frutos intactos o con huellas de manipulación por mamíferos, iv) cuantificar el posible efecto de animales sobre la dispersión y germinación de las semillas, y v) explorar si la distribución espacial de los individuos no reproductivos de Heteroflorum es o no agregada. Si los dispersores son granívoros que almacenan alimento, se espera una distribución espacial agregada; por otro lado, si los dispersores son mamíferos frugívoros terrestres, que se mueven largas distancias, se espera una distribución espacial no agregada (Vander Wall y Beck, 2012).
Materiales y métodos
El estudio se realizó en las localidades de El Guaricho (18º42’22” N, -101º40’19.7” O), La Joya de la Niña (18º42’1.0” N, -101º39’47.6” O) y El Zipimo (18º42’23” N, -101º39’06” O), con altitudes de 370, 466 y 550 m, respectivamente, en el municipio de Churumuco, Michoacán, México. Este municipio pertenece a la provincia fisiográfica de la Depresión del Balsas. El área de estudio presenta un clima caliente y seco, con temperatura promedio anual de 29.4 ºC, precipitación promedio anual de 564 mm, y una época seca entre octubre y junio. El tipo de vegetación dominante es el bosque tropical caducifolio. Más información sobre la flora del sitio de estudio se puede encontrar en Cortés-Flores et al. (2017) y Luna-Nieves et al. (2017) y sobre la fauna en Urrea-Galeano et al. (2016).
Es importante mencionar que las localidades estudiadas son Áreas de Conservación Comunitaria, localizadas en la reserva de la Biosfera Zicuirán-Infiernillo, en las que los pobladores del ejido Llano Ojo de Agua tienen un acceso restringido, llevando a cabo actividades de monitoreo de la biota y de los cuerpos de agua, lo que permite que el bosque tenga un buen estado de conservación y que sin dificultad se registre la presencia de especies de fauna nativa (Urrea-Galeano et al., 2016). El único indicio claro de actividades productivas antropogénicas es el libre pastoreo de ganado vacuno, el cual no presenta densidades importantes. En las localidades estudiadas no se talan árboles de Heteroflorum.
Heteroflorum es una especie dioica, caducifolia y presenta alturas de hasta15 m. La floración y fructificación ocurren exclusivamente en la época seca (Luna-Nieves et al., 2017). Sus frutos son vainas leñosas, indehiscentes y escleróticas, de 14-18 cm de largo y 2.3‑2.7 cm de ancho (fig. 1A); cada fruto tiene 6-12 semillas, de 8.3-11 mm de largo y 6-8 mm de ancho (Sousa, 2005; fig. 1B). Las vainas maduras permanecen por algunos meses en la copa de los árboles y a partir de la mitad de la época seca caen por gravedad, observándose una gran cantidad de éstos alrededor del tronco. Debido a su peso (5.58-18.36 g) y a la morfología del fruto, que carece de apéndices alados, éstos no pueden ser dispersados por viento. Las vainas y semillas de Heteroflorum poseen altos contenidos de hidratos de carbono (47%) y fibra cruda (36%), así como bajos contenidos de proteína (11%) y grasa (2%), considerándose potencial forraje para los rumiantes (Campos, 2006).

Para identificar los mamíferos que consumen los frutos/semillas caídos bajo la copa de Heteroflorum, en la localidad de El Zipimo se colocaron cámaras trampa en árboles en fructificación. En 6 árboles focales (distancia ≥ 30 m entre árboles) se instaló una cámara que estuvo activa durante 13 meses (octubre 2013‑octubre 2014). Posteriormente (marzo 2014), se colocaron 6 cámaras adicionales en árboles vecinos a los focales, las cuales estuvieron activas durante 5 meses. Las cámaras trampa fueron programadas para tomar fotografías durante las 24h, al momento de detectar el paso o movimiento de algún animal; el lapso entre fotografías fue de 5 segundos. Se consideró una fotocaptura al conjunto de fotografías tomadas a los individuos de una misma especie animal durante un periodo de 24 h (Medellín et al., 2006). Sólo aquellas especies de mamíferos que se registraron interactuando con los frutos/semillas de Heteroflorum fueron consideradas como consumidoras.
Para obtener información más detallada de cómo los mamíferos consumidores registrados en las fotografías manipulan los frutos/semillas de Heteroflorum, se hicieron observaciones en cautiverio (mamíferos grandes) y campo (roedores). En cautiverio se ofrecieron, como único alimento, frutos de Heteroflorum a 11 venados (Odocoileus virginianus), 10 pecarís (Pecari tajacu) y 3 coatís (Nasua narica) en el Parque Zoológico Benito Juárez (Morelia, Michoacán), así como a 9 vacas (Bos taurus) en una localidad cercana a la ciudad de Morelia. Con base en una revisión previa de 161 frutos de Heteroflorum, se obtuvo un promedio de 5 semillas por fruto. A cada especie animal se le ofrecieron 31 frutos (i.e., un estimado de 155 semillas). Después de la manipulación y/o consumo por los animales, se recolectaron las semillas que quedaron sobre el suelo y se observaron bajo el microscopio estereoscópico para registrar marcas de dentición. Durante 3 días se revisaron las heces para extraer las semillas defecadas. Para cada especie animal se obtuvieron los siguientes valores: número de semillas sobre el suelo (con o sin signo de impresión dental) que cayeron durante la masticación del fruto, sin haber sido tragadas, número de semillas defecadas, número de semillas tragadas (restando el número de semillas sobre el suelo del número de semillas ofrecidas) y el número de semillas digeridas (restando el número de semillas defecadas enteras del número de semillas tragadas).
Las observaciones de campo se hicieron para cuiniques (Notocitellus adocetus) y roedores nocturnos. La actividad de los primeros fue registrada por 2 personas en 3 árboles focales, durante 3 días entre las 7:00 y las 13:00 h, en La Joya de la Niña. Para roedores nocturnos, se instaló en El Zipimo una cámara trampa (registro continuo por 1 mes) a 1 m de la entrada de una pequeña cueva rocosa, que a su vez estaba a 2 m de un árbol de Heteroflorum. En ese lugar se habían observado previamente vainas abiertas, lo que sugirió el forrajeo por ratones.
Para determinar el destino de frutos/semillas que caen bajo las copas de los árboles, se trabajó con los mismos 6 árboles focales antes mencionados. A 2 m del tronco de cada árbol se colocaron 19 frutos de Heteroflorum sobre el suelo, en un área de 1 m2. Estos frutos fueron recolectados de la copa de los 6 árboles focales y se les hizo una marca externa para distinguirlos de los frutos que habían caído naturalmente, los cuales no fueron removidos para no alterar, en lo posible, el comportamiento de los animales. Los frutos focales se revisaron diariamente por una semana y posteriormente una vez al mes, hasta completar 5 meses (marzo-julio del 2014). Una vez finalizado el periodo de seguimiento, los frutos se categorizaron dentro de alguno de los siguientes destinos: a) frutos intactos no dispersados, cuando no hubo evidencia de manipulación por animales, b) frutos intactos dispersados, cuando los frutos fueron desplazados de su posición inicial, c) frutos abiertos no dispersados, cuando la vaina fue abierta dejando expuestas sus semillas (en la vaina o en el suelo) y d) frutos depredados, cuando sólo quedaron fragmentos de vaina y/o semillas sobre el suelo.
Para determinar si es posible que las semillas germinen al encontrarse fuera del fruto o estando en el interior de las vainas intactas o parcialmente dañadas por la manipulación por mamíferos, se realizó un experimento en condiciones naturales (El Zipimo) y otro en una casa sombra ubicada en el poblado del ejido. A partir de 10 árboles en fructificación, se recolectaron 24 frutos maduros de la copa de cada uno. Una parte del total de frutos (n = 40) fueron usados para la obtención de semillas sin daño físico externo, que fueron sometidas previamente a una prueba de flotación para remover las que fueran vanas. Con los frutos y semillas se formaron 20 lotes, cada uno integrado por 5 semillas y 10 frutos. La mitad de éstos fueron frutos intactos, que no presentaban daños por insectos o roedores, mientras que el resto incluyeron frutos abiertos manualmente para simular el consumo parcial por roedores observado previamente en campo.
La mitad de los lotes fueron colocados en el campo, en bolsas plásticas de vivero, sobre un sustrato obtenido del área. Las bolsas se colocaron dentro de una parcela de 2.25 m2, localizada bajo la copa de un árbol en fructificación de Heteroflorum. Esta parcela estuvo protegida con una malla metálica (apertura de malla 0.5 × 0.5 cm) para evitar la remoción de los frutos. Los otros 10 lotes se colocaron en la casa sombra (80% de sombreado). Las semillas fueron colocadas en bolsas plásticas, mientras que los frutos de cada categoría (intactos y con manipulación simulada), se ubicaron en recipientes plásticos independientes. Todos los frutos y semillas se colocaron sobre una mezcla de peat-moss, arena y arcilla (2:1:1). No se suministró riego a ninguno de los lotes, con el propósito de que estuvieran expuestas a las condiciones ambientales naturales. El registro de la germinación se hizo durante 13 meses (noviembre 2013-2014), con revisiones mensuales. El criterio para considerar un evento de germinación fue registrar la presencia de la radícula.
Por otro lado, para evaluar el efecto de los venados y vacas sobre la germinación, se colocaron las semillas en una cámara de germinación durante un mes, con los siguientes tratamientos: i) semillas no tragadas, pero con marcas de dentición de venados; ii) semillas tragadas y defecadas por vacas y iii) semillas control (extraídas directamente de los frutos). Se calcularon los porcentajes de germinación y se realizó una prueba de independencia usando la prueba exacta de Fisher para el cálculo de la probabilidad binomial. Por otro lado, se evaluó si las semillas (n = 42) de Heteroflorum presentaban dormancia física siguiendo las recomendaciones de Baskin y Baskin (1998).
Para determinar la densidad y distribución espacial de Heteroflorum, así como la proporción de individuos que son producto de un evento de dispersión, se mapearon todos los individuos de esta especie en 3 parcelas de 1,000 m2 (una en cada localidad). Los individuos se clasificaron en 2 grupos: a) reproductivos, aquellos árboles con un diámetro a la altura del pecho (DAP) ≥ 10 cm, y b) no reproductivos, incluyendo desde plántulas hasta árboles con DAP < 10 cm. Los individuos no reproductivos luego se categorizaron como ‘no dispersados’, si estaban debajo del área proyectada de la copa de un individuo reproductivo femenino o a una distancia de hasta 3 m a partir del borde de la copa (también se consideraron individuos reproductivos que estuvieran fuera de los límites de la parcela como posibles árboles madre, siempre y cuando el borde de su copa estuviera a ≤ 3 m del límite). Todos los demás individuos no reproductivos se consideraron como ‘dispersados’. La distancia de 3 m que se usó para establecer si un individuo es o no dispersado, si bien es arbitraria, se basa en Webb y Peart (2001). Además, este valor se considera conservador si se toma en cuenta que: i) las localidades estudiadas presentan una topografía relativamente plana, lo cual hace improbable que los frutos caídos se alejen más de esta distancia de la planta progenitora, ii) el tamaño y peso de los frutos no permite que éstos sean dispersados por viento, iii) si bien los frutos pueden permanecer durante la época de lluvias en el suelo, es altamente improbable que se presenten fuertes corrientes de agua en el sotobosque, ya que durante esa época se presenta una densa cobertura de hierbas, que forma un obstáculo que limita un posible desplazamiento de los frutos, y iv) aunque en la época de secas se pueden presentar fuertes pero inusuales eventos de lluvia, es muy probable que el agua se filtre rápidamente por el sustrato predominantemente rocoso, lo cual podría limitar que se formen corrientes de agua que pudieran desplazar los frutos. Se realizó una prueba de bondad de ajuste para evaluar las diferencias en la proporción de individuos dispersados vs. no dispersados en El Guaricho y La Joya de la Niña; dado el bajo número de individuos registrados en El Zipimo, para este sitio se calculó la probabilidad exacta binomial con la prueba de Fisher. Finalmente, se calculó el índice de dispersión de Morisita (1962) para identificar el patrón de distribución espacial de los individuos no reproductivos en las 3 localidades.
Tabla 1
Lista de las especies de mamíferos terrestres consumidores de frutos/semillas de Heteroflorum, fotocapturas (una fotocaptura representa un conjunto de fotografías tomadas a los individuos de una misma especie animal durante un periodo de 24 h) y periodo de actividad de cada especie (1 = madrugada, 2 = mañana, 3 = tarde y 4 = noche; en negritas el periodo con el mayor número de fotocapturas).
|
Orden/Familia |
Especie |
Nombre común |
Fotocapturas (%) |
Periodo de actividad |
|
Artiodactyla |
||||
|
Bovidae |
Bos taurus |
Vaca |
5 (1.27) |
2, 3 |
|
Cervidae |
Odocoileus virginianus |
Venado cola blanca |
77 (19.49) |
1, 2, 3, 4 |
|
Tayassuidae |
Pecari tajacu |
Pecarí de collar |
102 (25.82) |
1, 2, 3, 4 |
|
Carnivora |
||||
|
Procyonidae |
Nasua narica |
Coatí |
168 (42.53) |
1, 2, 3, 4 |
|
Rodentia |
||||
|
— |
— |
Ratones |
38 (9.62) |
1, 2, 4 |
|
Sciuridae |
Notocitellus adocetus |
Cuinique |
5 (1.27) |
2, 3 |
Resultados
Con un esfuerzo de muestreo total de 3,282 días-trampa, se obtuvo un total de 466 fotocapturas de mamíferos terrestres. Se identificaron 15 taxones, 14 a nivel de especie y 1 a nivel de orden; 6 especies se categorizaron como consumidoras (tabla 1). Los mamíferos que manipularon los frutos fueron animales herbívoros/frugívoros (vacas, venados cola blanca, pecarís de collar y coatís) o granívoros (ratones y cuiniques). En el primer caso, los individuos fueron fotografiados con el fruto en la boca (e.g., venado cola blanca; fig. 1C) o entre sus patas delanteras (e.g., coatí), mientras que en el segundo, los animales se registraron transportando los frutos en la boca o abriéndolos (e.g., N. adocetus). Los coatís, pecarís y venados fueron las especies de mamíferos consumidoras registradas con mayor frecuencia (168, 102 y 77 fotocapturas, respectivamente).
A partir de las observaciones en cautiverio, para los coatís, de un total de 31 frutos ofrecidos, 26 fueron manipulados. Ninguna semilla fue consumida, 90 (58%) quedaron sobre el suelo, sin signos de impresión dental, y las demás quedaron en el interior de frutos enteros o parcialmente abiertos. Los pecaríes consumieron todos los frutos ofrecidos, ingiriendo el 98% de las semillas y dejando caer al suelo 3 semillas, con impresión dental. No hubo semillas en sus excretas, lo que sugiere que todas las semillas tragadas fueron digeridas. De los frutos ofrecidos a venados, 2 fueron removidos por roedores; los 29 frutos restantes fueron manipulados por los venados. Del total de semillas estimadas dentro de estos frutos, 36 (25%) fueron tragadas, pero ninguna fue recuperada de las heces, por lo que se asume su digestión. Del suelo se recuperaron 109 (75%) semillas, 57 de ellas con impresión dental (fig. 1D). Las vacas manipularon 28 de los 31 frutos ofrecidos. Un total de 123 semillas (79%) fueron tragadas, 17 (11%) cayeron al suelo y 15 (10%) quedaron en los frutos no manipulados. De las semillas ingeridas, 13 (11%) fueron defecadas enteras (fig. 1E) y de las recuperadas del suelo, 9 (53%) tuvieron impresión dental.
Con respecto a las observaciones del comportamiento de roedores en campo, se registró que los cuiniques forrajean en búsqueda de frutos de Heteroflorum, tanto en la copa de los árboles (fig. 1F), como en el suelo, cerca de individuos reproductivos (fig. 2C). En ambos casos, los cuiniques abren los frutos y guardan algunas semillas en sus abazones; las demás semillas quedan sobre el suelo, dentro de la porción cerrada o parcialmente abierta de la vaina (fig. 2A, C). Las madrigueras de los cuiniques se encontraron debajo del suelo, algunas entradas fueron localizadas entre y debajo de rocas grandes, cerca o lejos de la base de un árbol. La extensión de algunas madrigueras fue de aproximadamente 2-3 m, a partir de una entrada hasta una salida ubicada a otro lado de la madriguera. Cerca de algunas entradas de las madrigueras, se encontraron restos de semillas depredadas de Heteroflorum, pero también semillas intactas (fig. 2C). En el caso de los roedores nocturnos, se registraron 4 fotocapturas en las que se observó a los ratones manipulando 5 vainas de Heteroflorum en la entrada de su madriguera. Revisiones posteriores revelaron que todas las semillas habían sido extraídas de los frutos y no se encontraron restos de ellas.

De un total de 114 frutos usados para determinar los posibles destinos de los frutos que caen bajo la copa, el 75% fueron frutos abiertos no dispersados (i.e., con semillas intactas en el suelo o en la vaina abierta; fig. 2A), el 24% frutos depredados (i.e., fragmentos de frutos y/o semillas) y el 1% frutos intactos dispersados. El fruto dispersado se encontró intacto a 3 m de la posición original.
En condiciones de campo (bajo árbol madre) o en la casa sombra, ninguna semilla germinó, en ninguno de los tratamientos (semillas fuera de frutos, semillas en el interior de las vainas intactas o parcialmente abiertas). Durante la época de secas, todos los frutos y semillas permanecieron en la misma condición y posición en la que habían sido colocados. En la época de lluvias, algunos frutos y las semillas contenidas en éstos se infestaron superficialmente de hongos y no se observó su desintegración (fig. 2B); estos hongos no se registraron en las semillas que se encontraban fuera de los frutos.
En el caso de los experimentos de germinación realizados para evaluar el efecto de los consumidores, se encontró que ninguna semilla control (n = 70) germinó. De las semillas con signos de impresión dental de los venados (n = 57), germinó el 14%. De las excretadas por las vacas (n = 13) germinó el 23%. Hubo un efecto positivo significativo sobre la germinación causado por la impresión dental de los venados (probabilidad exacta binomial, p = 0.006) y el paso por el tracto digestivo de las vacas (p = 0.002). Ninguna semilla utilizada para evaluar la dormancia física mostró aumento en su peso, lo que indica que la testa de la semilla es impermeable.
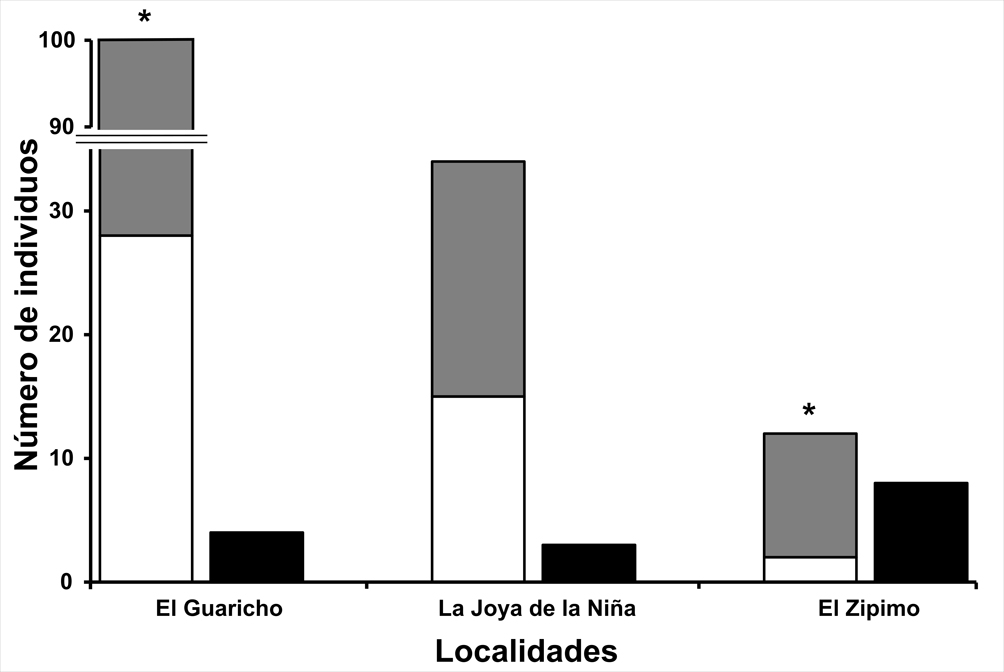
En las 3 localidades se registró un total de 161 individuos, 15 reproductivos y 146 no reproductivos. Tomando en cuenta que el área de muestreo en cada sitio fue de 0.1 ha, la mayor densidad poblacional se registró en El Guaricho, con 104 individuos (4 reproductivos y 100 no reproductivos), seguido de La Joya de la Niña y El Zipimo, con 37 (3 reproductivos y 34 no reproductivos) y 20 individuos (8 reproductivos y 12 no reproductivos), respectivamente. Del total de individuos registrados, el 91% se categorizó como no reproductivo (n = 142 individuos). Para éstos, en El Guaricho y El Zipimo, fueron más los que se categorizaron como individuos dispersados (El Guaricho: χ2 = 18.49, gl = 1, p < 0.0001; El Zipimo: probabilidad exacta binomial, p = 0.019; fig. 3). En La Joya de la Niña la proporción de dispersados fue igual a lo esperado por azar (χ2 = 0.26; gl = 1; p = 0.47). En las 3 localidades los individuos no reproductivos presentaron una distribución significativamente agrupada (El Guaricho, Id = 1.47, p < 0.05; La Joya de la Niña, Id = 1.74, p < 0.05; El Zipimo, Id = 2.19, p < 0.05).
Discusión
Los resultados del presente estudio indican que los pecaríes son únicamente depredadores de las semillas de Heteroflorum, pero los cuiniques, ratones, venados, vacas, y posiblemente coatíes, son potencialmente dispersores de algunas semillas de Heteroflorum y contribuyen positivamente con la liberación de las semillas de la vaina (fig. 2C). La apertura de la vaina para la liberación de las semillas parece ser una acción necesaria para que ocurra la primera fase de regeneración natural de esta especie de árbol.
Algunas especies de Leguminosae se caracterizan por tener semillas con dormancia física (e.g., Argel y Paton, 1999; Ray y Brown, 1994), por lo que requieren algún mecanismo de escarificación para germinar. Dada la impermeabilidad de la testa de las semillas de Heteroflorum registrada en este estudio, se considera que sus semillas también presentan este tipo de dormancia. Además, los resultados de los experimentos de germinación en campo, casa sombra y laboratorio, sugieren que sin la acción de algún mamífero que interrumpa esta dormancia, las semillas no germinan. Además, las semillas que no germinan podrían perder su viabilidad debido al ataque de patógenos (e.g., hongos; Traveset et al., 2007), en especial en aquellas que permanecen en el interior de las vainas. Sin embargo, estudios futuros tendrán que evaluar la viabilidad de las semillas de Heteroflorum a largo plazo, así como el efecto demográfico que pudiera tener un retraso en la germinación. También será necesario determinar si factores climáticos, como la fluctuación en la temperatura, podrían favorecer la germinación de las semillas, al estar expuestas sobre el suelo durante un tiempo superior al evaluado.
La manipulación de frutos y semillas por venados y vacas en cautiverio sugiere que ambas especies promueven la germinación mediante la escarificación de las semillas. La escarificación mecánica por venados (e.g., durante la masticación) y química por vacas (e.g., mediante el paso por el tracto digestivo), permitió una germinación del 14% y 23% de las semillas caídas y defecadas, respectivamente. Estos mecanismos de escarificación son importantes para la germinación de algunas leguminosas (Malo y Suárez, 1995; Peinetti et al., 1993; Zhang et al., 2013) y en este caso, se sugiere que también lo son para Heteroflorum. El tipo de escarificación realizada por los venados en este estudio y su efecto sobre la germinación, no han sido documentados con anterioridad para estos animales.
En 2 de las 3 localidades estudiadas se encontró una mayor proporción de individuos no reproductivos que habían sido producto de un evento de dispersión de semillas. Lo anterior sugiere que los mamíferos terrestres, mediante su comportamiento de forrajeo, podrían estar facilitando la regeneración de Heteroflorum al desplazar las semillas fuera del área de influencia de un individuo reproductivo. Sin embargo, dado el comportamiento de forrajeo y manipulación de las semillas observados, las distancias de dispersión podrían ser, en su mayoría, relativamente cortas, generando la distribución espacial agregada observada para la especie. Los cuiniques, roedores y venados podrían ser los vectores responsables de la dispersión a cortas distancias, mientras que las vacas posiblemente serían responsables de eventos a distancias mayores. Efectos de la dispersión de semillas por mamíferos terrestres sobre la distribución espacial agregada de las plantas, han sido documentados para otras especies de árboles tropicales (e.g., Fragoso et al., 2003).
Durante el consumo de frutos por venados, algunos estudios han mostrado que las semillas de algunas especies son tragadas y luego digeridas en el rumen, mientras que otras son escupidas y por lo tanto dispersadas (Bodmer, 1991); ambas situaciones ocurrieron en este estudio. Otros estudios también han documentado el papel de los venados en la dispersión de semillas de algunas especies de plantas (Jara-Guerrero et al., 2018; Mandujano et al., 1994; Vellend et al., 2003). Con respecto a las vacas, dado el largo tiempo de retención de las semillas en el tracto digestivo (entre 48 y 72 h) y las distancias que recorren, se podría pensar que desempeñan un papel importante en eventos de dispersión de larga distancia (Reynolds, 1954). La afirmación anterior se podría reforzar con las observaciones en el campo, donde se encontraron semillas y plántulas de Heteroflorum en las excretas de vacas (fig. 2D, E) localizadas lejos de individuos adultos. Estudios previos han documentado la relevancia de las vacas en la dispersión de semillas de otras leguminosas (Ramos et al., 2005; Zhang et al., 2013). Se postula que, junto con los caballos, las vacas pueden estar jugando un papel crucial en la regeneración de especies de árboles, muchas de ellas leguminosas, que hasta el Pleistoceno eran dispersados por especies de megafauna, hoy extintas (Janzen y Martin, 1982). Debido a que en este estudio no se realizó un seguimiento detallado a las semillas liberadas por los animales consumidores, no hay evidencia contundente del papel de éstos en la dispersión. Por lo tanto, lo planteado anteriormente son sólo hipótesis que requieren de estudios futuros que las comprueben, particularmente a través de cuantificar la proporción de semillas que son dispersadas o depredadas, así como el destino final de las semillas dispersadas, y el impacto de estos consumidores en la demografía de la planta, su distribución espacial y estructura
genética.
Es importante mencionar que la función que los mamíferos desempeñan al liberar las semillas de los frutos durante su forrajeo puede ser importante para las plantas. Para muchas especies de semillas, la separación del fruto es necesaria para que ocurra la germinación (Traveset et al., 2007), aunque en el caso de Heteroflorum esta separación no parece ser suficiente para estimular la germinación. Otros efectos positivos derivados de la apertura de la vaina y exposición de las semillas, que posiblemente sí estén ocurriendo en Heteroflorum, incluyen un menor ataque por patógenos, y un aumento en la probabilidad de dispersión secundaria por otros animales. En el caso de Heteroflorum, los vectores más probables de dispersión secundaria serían los roedores. La remoción por roedores con comportamiento de almacenaje de semillas puede tener efectos positivos o negativos para la planta. Los negativos ocurren cuando las semillas son depredadas (Howe, 1989) o depositadas en micrositios que impidan la germinación o emergencia de la plántula (Forget, 1993). Sin embargo, en muchos casos las semillas sobreviven al ser olvidadas en reservorios lejos de la planta madre, lo cual favorece el establecimiento y subsecuente desempeño de las plántulas (Vander Wall y Beck, 2012). Estos 2 posibles escenarios se registraron para Heteroflorum, cerca de las madrigueras de los cuiniques, ya que algunas semillas fueron depredadas, pero otras se encontraron sin ningún daño aparente (fig. 2C). Será necesario realizar un seguimiento detallado del destino de las semillas para evaluar la relevancia de estos destinos en la estructura demográfica de la especie.
Es evidente que existen interacciones entre los frutos y semillas de Heteroflorum y algunos mamíferos terrestres en este sistema de estudio, las cuales permiten el escape de las semillas del fruto, y favorecen y/o aceleran el proceso de la germinación. Realizar investigaciones con este enfoque ecológico contribuye en el entendimiento de la estructura y funcionamiento de las comunidades bióticas de los bosques tropicales. Un enfoque en las interacciones bióticas es imprescindible para el desarrollo de estrategias que favorezcan el aprovechamiento, mantenimiento y conser-
vación de los ecosistemas (Valiente-Banuet et al., 2015).
Agradecimientos
Al Posgrado en Ciencias Biológicas, de la Universidad Nacional Autónoma de México, por los estudios de maestría de la primera autora, así como al Consejo Nacional de Ciencia y Tecnología por la beca otorgada. A IDEA WILD por el aporte financiero para la compra de cámaras-trampa. A las autoridades del Parque Zoológico Benito Juárez de Morelia, M.V.Z. Martín Zavala Mondragón, M.V.Z. Alberto Arrés Rangel y M.V.Z. Roberto López, por facilitar el trabajo con los animales en cautiverio. A los miembros del ejido Llano Ojo de Agua por permitir el acceso a las áreas de conservación, y al señor Misael Rojas López por su invaluable ayuda en campo. Agradecemos a la M. en C. Ma. Guadalupe Cornejo Tenorio por su apoyo en la elaboración de las figuras 1 y 2.
Referencias
Argel, P. J. y Paton, C. J. (1999). Overcoming legume hardseedness. En D. S. Loch y J. E. Ferguson (Eds.), Forage seed production: tropical and sub-tropical species (pp. 247–265). Wallingford, United Kingdom: CABI.
Baskin, J. M. y Baskin, C. C. (1998). Seeds: ecology, biogeography, and evolution of dormancy and germination. San Diego: Academic Press.
Bodmer, R. E. (1991). Strategies of seed dispersal and seed predation in Amazonian ungulates. Biotropica, 23, 255–261.
Campos, R. V. (2006). Evaluación nutrimental y toxicológica de algunas plantas cultivables y silvestres mexicanas (Tesis). Facultad de Química, Universidad Nacional Autónoma de México. Ciudad de México, México.
Ceballos, G. y Valenzuela, D. (2010). Diversidad, ecología y conservación de los vertebrados de Latinoamérica. En G. Ceballos, L. Martínez, A. García, E. Espinoza, J. Bezaury y R. Dirzo (Eds.), Diversidad, amenazas y áreas prioritarias para la conservación de las selvas secas del Pacífico de México (pp. 93–118). Ciudad de México: Fondo de Cultura Económica.
Cortés-Flores, J., Hernández-Esquivel, K. B., González-Rodríguez, A. e Ibarra-Manríquez, G. (2017). Flowering phenology, growth forms and pollination syndromes in a tropical dry forest species: influence of phylogeny and abiotic factors. American Journal of Botany, 104, 39–49.
del Val, E. y Boege, K. (2012). ¿Por qué estudiar las interacciones bióticas? En E. del Val y K. Boege (Eds.), Ecología y evolución de las interacciones bióticas (pp. 11–13). Ciudad de México: Universidad Nacional Autónoma de México/ Fondo de Cultura Económica.
Fleming, T. H., Breitwhisch, R. y Whitesides, G. H. (1987). Patterns of tropical vertebrate frugivore diversity. Annual Review in Ecology and Systematics, 18, 91–109.
Fleming, T. H. y Kress, W. J. (2013). The ornaments of life. Chicago: The University of Chicago Press.
Forget, P. M. (1993). Postdispersal predation and scatterhoarding of Dipteryx panamensis (Papilionaceae) seeds by rodents in Panama. Oecologia, 94, 255–261.
Fragoso, J. M. V., Silvius, K. M. y Correa, J. A. (2003). Long-distance seed dispersal by tapirs increases seed survival and aggregates tropical trees. Ecology, 84, 1998–2006.
Howe, H. F. (1989). Scatter and clump dispersal and seedling demography: hypothesis and implications. Oecologia, 79, 417–426.
Howe, H. F. y Smallwood, J. (1982). Ecology of seed dispersal. Annual Review of Ecology and Systematics, 13, 201–228.
Janzen, D. H. y Martin, P. S. (1982). Neotropical anachronisms: the fruits the gomphotheres ate. Science, 215, 19–27.
Jara-Guerrero, A., Escribano-Ávila, G., Espinosa, C. I., De la Cruz, M. y Méndez, M. (2018). White-tailed deer as the last megafauna dispersing seeds in Neotropical dry forest: the role of fruit and seed traits. Biotropica, 50, 169–177.
Jordano, P. (2014). Fruits and frugivory. En R. S. Gallagher (Eds.), Seeds: the ecology of regeneration in plant communities (pp.18–61). Wallingford, United Kingdom: CABI.
Linares-Palomino, R., Oliveira-Filho, A. T. y Pennington R. T. (2011). Neotropical seasonally dry forest: diversity, endemism, and biogeopgraphy of woody plants. En R. Dirzo, H. S. Young, H. A. Mooney y G. Ceballos (Eds.), Seasonally dry tropical forest ecology and conservation (pp. 3–22). Washington D.C.: Island Press.
Lott, E. J. y Atkinson, T. H. (2010). Diversidad florística. En G. Ceballos, L. Martínez, A. García, E. Espinoza, J. Bezaury y R. Dirzo (Eds.), Diversidad, amenazas y áreas prioritarias para la conservación de las selvas secas del Pacífico de México (pp. 63–76). Ciudad de México: Fondo de Cultura Económica.
Luna-Nieves, A. L., Meave, J. A., Cerdeira, L. P. e Ibarra-Manríquez, G. (2017). Reproductive phenology of useful seasonally dry tropical forest trees: guiding patterns for seed collection and plant propagation in nurseries. Forest Ecology and Management, 393, 52–62.
Malo, J. E. y Suárez, F. (1995). Cattle dung and the fate of Biserrula pelecinus L. (Leguminosae) in a Mediterranean pasture: seed dispersal, germination and recruitment. Botanical Journal of the Linnean Society, 118, 139–148.
Mandujano, S., Gallina, S. y Bullock, S. H. (1994). Frugivory and dispersal of Spondias purpurea (Anacardiaceae) in a tropical dry forest of México. Revista de Biología Tropical, 42, 105–112.
Medellín, R. A., Azuara, D., Maffei, L., Zarza, H., Bárcenas, H., Cruz, E. et al. (2006). Censos y monitoreos. En G. Ceballos y C. Chávez (Eds.), Memorias del primer Simposio: El jaguar mexicano en el siglo XXI: situación actual y manejo (pp. 25–35), Ciudad de México: Conabio/ Alianza WWF-Telcel/ Universidad Nacional Autónoma de México.
Morisita, M. (1962). Morisita index, a measure of dispersion of individuals. Researches on Population Ecology, 4, 1–7.
Peinetti, R., Pereyra, M., Kin, A. y Sosa, A. (1993). Effects of cattle ingestion on viability and germination rate of calden (Prosopis caldenia) seeds. Journal of Range Management, 46, 483–486.
Ramos, M. E., Robles, A. B., Cardoso, J. A., Ruiz-Mirazo, J. y González-Rebollar, J. L. (2005). Dispersión endozoócora por ganado ovino de cuatro leguminosas de interés forrajero. En B. de la Roza, A. Argamentería, A. Martínez y K. Osoro (Eds.), Producciones agroganaderas: gestión eficiente y conservación del medio natural (pp. 92–929). Asturias, España: Serida.
Ray, G. J. y Brown, B. J. (1994). Seed ecology of woody species in a Caribbean dry forest. Restoration Ecology, 2, 156-–163.
Reynolds, H. G. (1954). Some interrelations of the Merrian Kangaroo rat to Velvet Mesquite. Journal of Range Management, 7, 176–180.
Russo, S. E., Portnoy, S. y Augspurger, C. K. (2006). Incorporating animal behavior into seed dispersal models: implications for seed shadows. Ecology, 87, 3160–3174.
Salas-Morales, S. H., Schibli, L., Nava-Zafra, A. y Saynes-Vásquez, A. (2007). Flora de la costa de Oaxaca, México (2): lista florística comentada del Parque Nacional Huatulco. Boletín de la Sociedad Botánica de México, 81, 101–130.
Smythe, N. (1986). Competition and resource partitioning in the guild of neotropical terrestrial frugivorous mammals. Annual Review of Ecology and Systematics, 17, 169–188.
Sousa, M. (2005). Heteroflorum: un nuevo género del grupo Peltophorum (Leguminosae: Caesalpinioideae: Caesalpinieae), endémico para México. Novon, 15, 213–218.
Traveset, A., Robertson, A. W. y Rodríguez-Pérez, J. (2007). A review on the role of endozoochory in seed germination. En A. J. Dennis, E. W. Schupp, R. J. Green y D. A. Westcott (Eds.), Seed dispersal. Theory and its application in a changing world (pp. 78–103). Wallingford, United Kingdom: CABI.
Trejo, I. (1998). Distribución y diversidad de selvas bajas en México: relaciones con el clima y el suelo (Tesis doctoral). Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad de México, México.
Urrea-Galeano, L. A., Rojas-López, M., Sánchez-Sánchez, L. e Ibarra-Manríquez, G. (2016). Registro de Puma yagouaroundi en la Reserva de la Biosfera Zicuirán-Infiernillo, Michoacán. Revista Mexicana de Biodiversidad, 87, 548–551.
Valiente-Banuet, A., Aizen, M. A., Alcántara, J. M., Arroyo, J., Cocucci, A., Galetti, M. et al. (2015). Beyond species loss: the extinction of ecological interactions in a changing world. Functional Ecology, 29, 299–307.
Vander Wall, S. B. y Beck, M. J. (2012). A comparison of frugivory and scatter-hoarding seed-dispersal syndromes. Botanical Review, 78, 10–31.
Vellend, M., Myers, J. A., Gardescu, S. y Marks, P. L. (2003). Dispersal of Trillium seeds by deer: implications for long-distance migration of forest herbs. Ecology, 54, 1067–1072.
Villaseñor, J. L. y Ortiz, E. (2014). Biodiversidad de las plantas con flores (División Magnoliophyta) en México. Revista Mexicana de Biodiversidad, 85 (Suplem.), S134–S142.
Webb, C. O. y Peart, D. R. (2001). High seed dispersal rates in faunally intact tropical rain forest: theoretical and conservation implication. Ecology Letters, 4, 491–499.
Zhang, X., Yang, Y., Li, H. y Tian, X. (2013). Potential role of endozoochory by cattle and sheep as an important dispersal mechanism of Zoysia japonica seeds. African Journal of Agricultural Research, 8, 3967–3972.


