
Miguel Ángel Suastes-Jiménez a, Ludivina Barrientos-Lozano b, Jorge Gutiérrez-Rodríguez a, c, Jovana M. Jasso-Martínez a, d, Alejandro Zaldívar-Riverón a, *
a Universidad Nacional Autónoma de México, Instituto de Biología, Colección Nacional de Insectos, 3er Circuito Exterior s/n, Cd. Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
b Tecnológico Nacional de México- Instituto Tecnológico de Cd. Victoria, Blvd. Emilio Portes Gil No. 1301, 87010 Ciudad Victoria, Tamaulipas, Mexico
c Estación Biológica de Doñana CSIC, Departmento de Ecología y Evolución, C/Américo Vespucio, 26, 41092, Isla de la Cartuja, Sevilla, Spain
d Smithsonian Institution, National Museum of Natural History, Department of Entomology, 10th St. & Constitution Ave. NW, Washington, D.C. 20560, USA
*Corresponding author: azaldivar@ib.unam.mx (A. Zaldívar-Riverón)
Received: 30 November 2022; accepted: 28 March 2023
http://zoobank.org/urn:lsid:zoobank.org:pub:31072EA4-A6E1-4A2D-98FC-6B8B4240A2BE
Abstract
We carried out a taxonomic revision of the Stenopelmatus talpa species group (Stenopelmatidae: Stenopelmatini), a poorly studied group of orthopterans whose species are distributed along the Trans-Mexican Volcanic Belt and adjacent areas of the Sierra Madre Oriental and Sierra Madre del Sur in Central Mexico. We examined both external morphological and male internal genitalia features, and also carried out a molecular species delimitation based on DNA barcoding. An integrative approach based on congruence of morphological and molecular evidence helped to delimit 9 species for the S. talpa species group, 7 of which are new to science whereas the remaining 2 are already described, S. talpa and S. typhlops. We described 4 out of the above 7 new species: Stenopelmatus chilango sp. nov., S. purepecha sp. nov., S. sierragordaensis sp. nov. and S. tlaxcalli sp. nov. A key to the described species of the S. talpa species group is provided. Our study highlights the relevance of the male internal genitalia as a valuable character system to delimit species in Stenopelmatus.
Keywords: Male internal genitalia; Diagnosis; Description; Ensifera; Mexico
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Revisión taxonómica del grupo de especies Stenopelmatus talpa (Orthoptera: Stenopelmatidae: Stenopelmatini)
Resumen
Realizamos una revisión taxonómica del grupo de especies Stenopelmatus talpa (Stenopelmatidae: Stenopelmatini), un grupo de ortópteros poco estudiado cuyas especies se distribuyen a lo largo de la Faja Volcánica Transmexicana y áreas adyacentes de la Sierra Madre Oriental y Sierra Madre del Sur en el centro de México. Examinamos características tanto de la morfología externa como de genitales internos masculinos y también llevamos a cabo una delimitación de especies basada en códigos de barras de ADN. Un enfoque integrador basado en la congruencia de evidencia morfológica y molecular ayudó a delimitar 9 especies para el grupo de especies S. talpa, 7 de las cuales son nuevas para la ciencia, mientras que las 2 restantes ya están descritas, S. talpa y S. typhlops. Describimos 4 de las 7 especies nuevas: Stenopelmatus chilango sp. nov., S. purepecha sp. nov., S. sierragordaensis sp. nov. y S. tlaxcalli sp. nov. Se proporciona una clave para las especies descritas del grupo de especies S. talpa. Nuestro estudio destaca la relevancia de los genitales internos masculinos como un sistema de caracteres valioso para delimitar especies en Stenopelmatus.
Palabras clave: Genitales internos masculinos; Diagnosis; Descripción; Ensifera; México
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
The tribe Stenopelmatini comprises a group of cricket-like ensiferous orthopterans that belong to the family Stenopelmatidae (Gorochov, 2021). Members of this tribe are commonly known as Jerusalem crickets in the U.S. (Weissman et al., 2008), and as “cara de niño” or “niños de la tierra” in Mexico (Weissman, 2005; Sánchez-Xolalpa et al., 2017). The supraspecific classification of this family was recently updated by Gorochov (2021), who changed Stenopelmatini from subfamily to tribe level. Currently, Stenopelmatini contains 2 valid genera, Ammopelmatus Tinkham, 1965 and Stenopelmatus Burmeister, 1938. The first one is distributed from southwestern Canada across the western part of the U.S. to Baja California in northern Mexico, and the second mainly occurs in high altitude regions along Mesoamerica from northern Mexico to Panama and in Ecuador in northern South America (Weissman et al., 2021).
Weissman et al. (2021) recently revised the systematics of Stenopelmatus employing nuclear genome-wide (anchor hybrid enrichment) and mitochondrial (mt) DNA sequence data, external morphological, ethological and cytological information. Based on the phylogenetic relationships recovered with the molecular data and on the observed variation of the remaining examined character systems, the authors proposed that Viscainopelmatus Tinkham, 1970 and Stenopelmatopterus Gorochov, 1988 were junior synonyms of Ammopelmatus and Stenopelmatus, respectively. They also described 14 new species of Stenopelmatus, classified 13 species of this genus as nomina dubia, and transferred 16 species to Ammopelmatus. As a result of this revision, Stenopelmatus was considered to include 19 described species, although its actual species diversity is considerably higher (Gutiérrez-Rodríguez et al., 2022; Weissman et al., 2021). More recently, Gutiérrez-Rodríguez et al. (2022) conducted a species delimitation and phylogenetic study for Stenopelmatus using nuclear-genome wide (3RAD) and mtDNA sequence data. These authors proposed the existence of 4 species groups within the genus, the S. faulkneri, S. talpa, Central America and S. piceiventris groups.
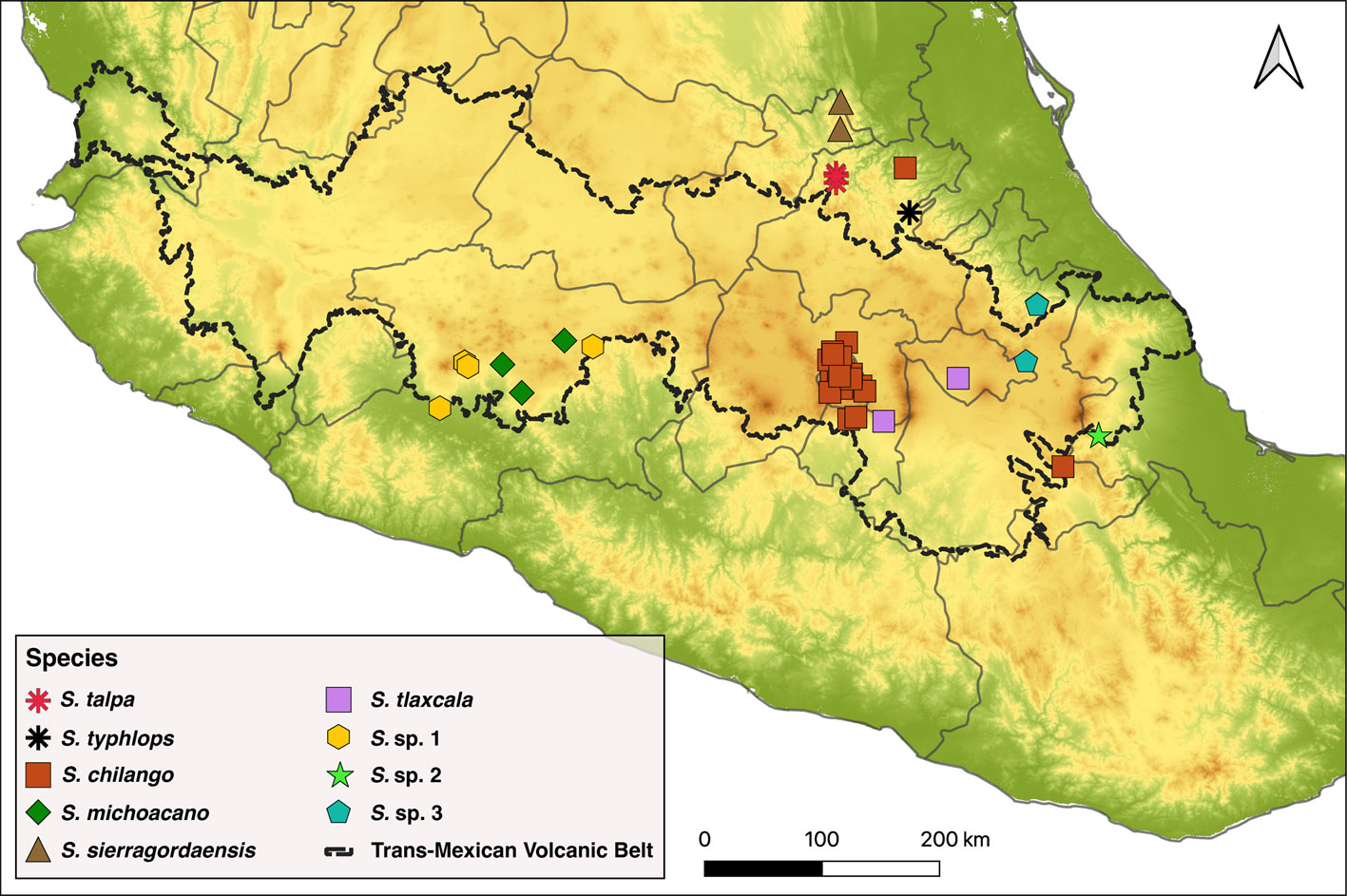
The S. talpa species group is represented by species with a relatively large size and usually with a reddish-orange and black color pattern except by S. typhlops Rehn, 1903, which is completely black (Gutiérrez-Rodríguez et al., 2022; Ruiz-Mendoza et al., 2023). This group receives its name after S. talpa Burmeister, 1838, a species originally assumed to have a wide geographical distribution throughout central México, though it appears to be restricted to pine and pine-oak forests in its type locality and its surroundings in the municipalities of Jacala and Zimapán, in north-northwest Hidalgo (Gutiérrez-Rodríguez et al., 2022; Ruiz-Mendoza et al., 2023; Weissman et al., 2021). Besides the 2 aforementioned species, the S. talpa species group also appeared to contain at least 5 undescribed species that were delimited based on nuclear genome-wide and mitogenome DNA sequence data (Gutiérrez-Rodríguez et al., 2022; Ruiz-Mendoza et al., 2023). Members of this species group occur in central Mexico across the Trans-Mexican Volcanic Belt province and adjacent areas of the Sierra Madre Oriental and Sierra Madre del Sur provinces (Gutiérrez-Rodríguez et al., 2022; Ruiz-Mendoza et al., 2023).
Species of Stenopelmatus are characterized by their highly conserved external morphology (Weissman, 2001; Weissman et al., 2008, 2021). For this reason, Weissman et al. (2021) proposed that the newly described species of the genus need to be diagnosed based on a combination of alternative features, including karyotype chromosome number, geographic distribution, call drum pattern, and mt and/or nuclear DNA profiles. However, a character system that requires more attention is the male genitalia, which has never been thoroughly assessed within the entire family Stenopelmatidae (Gorochov, 2021).
Here, we carried out a taxonomic revision for the S. talpa species group based on examination of external morphological and male genitalia features of specimens collected along central Mexico. We also molecularly assessed the species diversity within the group based on the DNA barcoding locus (Hebert et al., 2003). Based on the gathered information, we redescribed the 2 previously described species, S. talpa and S. typhlops, and described 4 new species mainly based on male genitalia features, a character system that was found to be informative for species delimitation within the genus.
Materials and methods
We examined a total of 112 specimens assigned to the S. talpa species group that were collected from a number of localities situated along central Mexico (Fig. 1). All specimens are deposited in the Colección Nacional de Insectos (CNIN), Instituto de Biología, Universidad Nacional Autónoma de México (IB-UNAM). We also examined the external morphological features of the neotype of S. talpa, which is deposited in the California Academy of Sciences, San Francisco, California, USA (CAS).
Observation and description of external morphological features were made using a stereo microscope ZEISS Discovery V8 with a lens ZEISS Achromat S 1.5x FWD 28 mm lens. We examined the following characters: shape of frontal fastigium, shape of pronotum (dorsal and lateral view), shape of cerci, shape of subgenital and supra-anal plates, and shape of internal genitalia in males and of ovipositor in females. Measurements were taken for the body total length (from vertex to tip of abdomen), and for the following morphological features: length and width of pronotum (dorsal view), femora (anterior, middle and posterior), length of cerci, length and width of supra-anal and subgenital plates, and length of ovipositor. Measurements of morphological characters greater than 0.5 cm were taken with a millimetric ruler and a Vernier. For smaller features, we used the ZEN 3.1 (blue edition) program to measure structures from photographs taken by a ZEISS AxioCam ERc5s fixed camera and a 60N-C 2/3” (0.63x) adapter.
The internal male genitalia of 15 adult specimens belonging to most of the DNA barcode species that were delimited were obtained following the extraction protocol described by Rocha-Sánchez et al. (2018). We followed the terminology of the internal male genitalia structures proposed by Gorochov (2021) for the families Stenopelmatidae and Anostostomatidae. Males were confirmed to reach adult stage by the presence of a pair of black, evident hooks near the cerci. The terminalia of each male specimen was placed in a petri dish with hot water for 45 minutes and it was subsequently removed with entomological needles and forceps. The terminalia was then placed in a petri dish with 10% potassium hydroxide until the muscle tissue was dissolved. The remaining genitalia was subsequently washed and examined with a ZEISS DiscoveryV8 stereoscopic microscope and a ZEISS Achromat S 1.5x FWD 28 mm lens. Digital photographs of both external morphological and male genitalia features were taken with a ZEISS AxioCam ERc 5s camera, and edited using the ZEN 3.1 (blue edition) imaging system. The male genitalia were preserved in Eppendorf tubes with glycerin and deposited in the CNIN IBUNAM.
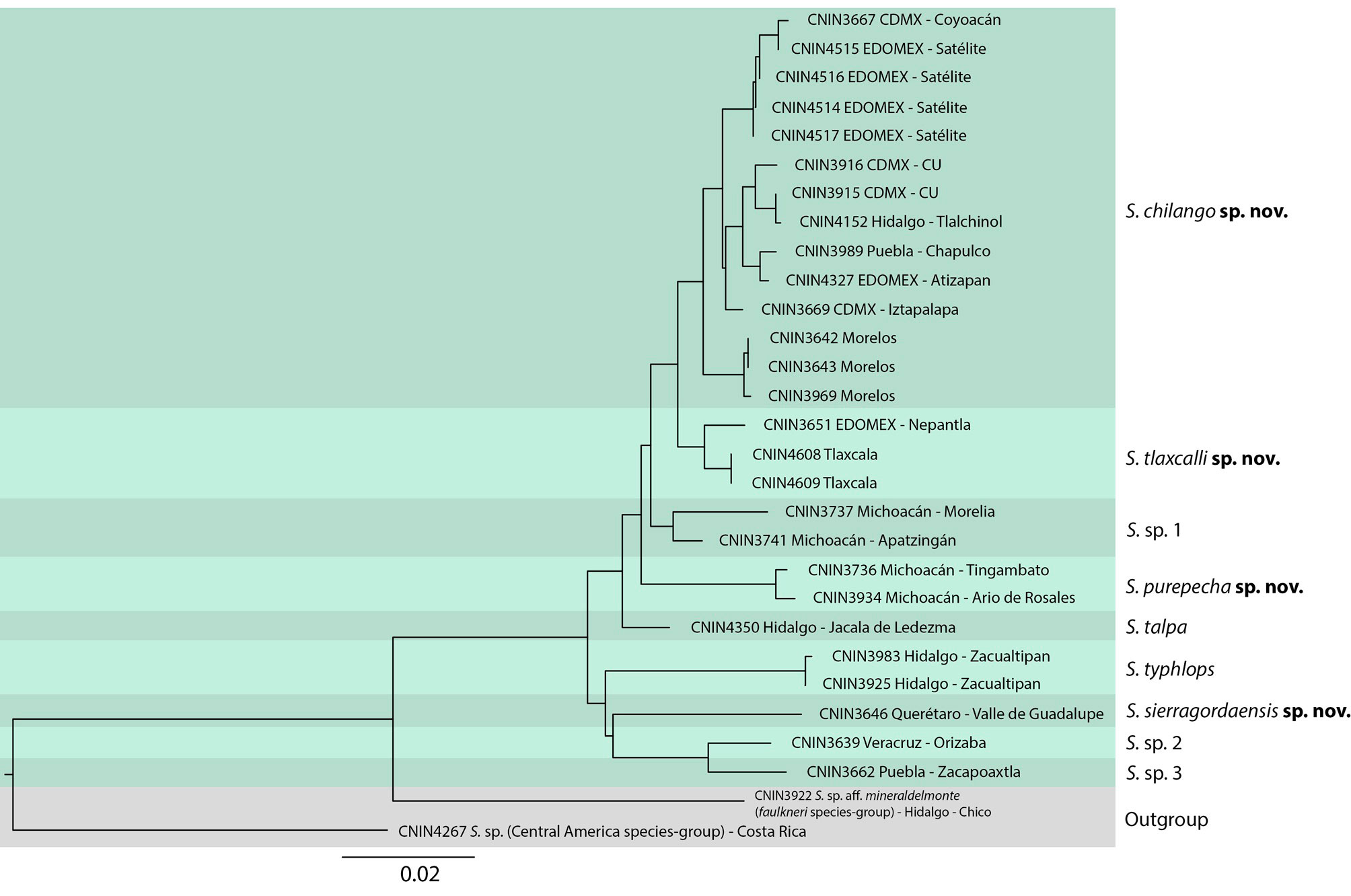
We assessed the species limits within the S. talpa species group with the DNA barcoding locus including representative specimens of all the localities where we collected fresh tissue samples. We sequenced a ~ 658 bp fragment of the cytochrome oxidase I (COI) mtDNA gene for 10 specimens assigned to the S. talpa species group. We also generated COI sequences of 2 species belonging to 2 other Stenopelmatus species groups as outgroups, S. sp. CNIN3922 (GenBank accession no. OP778773; S. faulkneri species group) and Stenopelmatus sp. CNIN4267 (GenBank accession no. OP778776; Central America species group), using the second species to root the tree. Moreover, we included in our analysis 19 previously published COI sequences of specimens belonging to the S. talpa species group (Gutiérrez-Rodríguez et al., 2022; Ruiz-Mendoza et al., 2023), of which 14 specimens were obtained from assembled mitogenomes. For the sequenced specimens, we first removed muscle tissue from a hind femur of each specimen, and genomic DNA was then extracted with the EZ-10 Spin Kit minipreps DNA Genomic Column Kit (BIOBasic, Toronto, Ontario, Canada) following the manufacturer’s instructions. Amplification was performed using the following primers especially designed for species of Stenopelmatus: COX1-F: 5 CTCRACRAATCATAAAGATATYGG and COX1-R2: 5´GTRTCWACATCTATWCCTACAG. We also included in our analysis 9 previously published COI sequences of specimens of the above species group (Gutiérrez-Rodríguez et al., 2022).
Table 1
Uncorrected COI distances among the species of the Stenopelmatus talpa species group. * = Interspecific pairwise distance < 2%. Numbers in bold = intraspecific variation.
| S. chilango sp. nov. | S. tlaxcalli sp. nov. | S. purepecha sp. nov. | S. sp. 1 | S. talpa | S. typhlops | S. sierragordaensis | S. sp. 2 | S. sp. 3 | |
| S. chilango sp. nov. | 0-0.019 | – | – | – | – | – | – | – | – |
| S. tlaxcalli sp. nov. | 0.013*-0.027 | 0-0.01 | – | – | – | – | – | – | – |
| S. purepecha sp. nov. | 0.019*-0.039 | 0.019*-0.029 | 0.019 | – | – | – | – | – | – |
| S. sp. 1 | 0.033-0.05 | 0.036-0.04 | 0.025-0.044 | 0.005 | – | – | – | – | – |
| S. talpa | 0.024-0.034 | 0.023-0.023 | 0.020-0.028 | 0.0310-0.033 | – | – | – | – | – |
| S. typhlops | 0.05 -0.068 | 0.057-0.059 | 0.056-0.07 | 0.065-0.068 | 0.041-0.045 | 0 | – | – | – |
| S. sierragordaensis sp. nov. | 0.052-0.063 | 0.055-0.058 | 0.055-0.061 | 0.064-0.066 | 0.043 | 0.058-0.058 | – | – | – |
| S. sp. 2 | 0.048-0.062 | 0.046-0.053 | 0.047-0.058 | 0.056-0.061 | 0.043 | 0.058-0.06 | 0.053 | – | – |
| S. sp. 3 | 0.052-0.065 | 0.049-0.053 | 0.048-0.06 | 0.056-0.061 | 0.041 | 0.058-0.054 | 0.053 | 0.021 | – |
The examined COI sequences were manually aligned and verified based on their reading frame with the program Geneious version 10.2.6 (Kearse et al., 2012). We used the 2% COI divergence criterion for molecular species delimitation (Hebert et al., 2003), calculating uncorrected COI genetic distances with the program PAUP version 4.0a (Swofford, 2002) and further visualizing them reconstructing a Neighbor Joining (NJ) distance tree with the program PAUP version 4.0 (Swofford, 2004). The DNA voucher and GenBank accession numbers of the sequenced specimens is included in the taxonomic part.
The species names were selected during a ludic auction that took place on the 25th of August 2023 at the Pabellón de la Biodiversidad, Instituto de Biología, Universidad Nacional Autónoma de México, in Mexico City, Mexico.
Results
The 2% COI divergence criterion yielded a total of 9 barcoding species for the specimens assigned to the S. talpa species group, of which 2 belong to specimens assigned to S. talpa and S. typhlops (Table 1; Fig. 2). We only found 3 cases of values < 2% between the barcoding species, of which 2 of them were marginally lower and involved the species from Morelia described below as S. purepecha sp. nov. with those of S. chilango sp. nov. (1.9–3.9%) and S. tlaxcalli sp. nov. (1.9-2.9%). The third case of a value < 2% was between a specimen of S. tlaxcalli sp. nov. and 1 of S. chilango sp. nov. (1.3%-2.7%). Based on an integrative taxonomic approach by congruence (Padial et al., 2010) using the above molecular evidence and a detailed morphological revision of the available specimens, here we describe 4 species for which we had adult males, and redescribe the 2 previously described species of the group, S. talpa and S. typhlops. All these species are mainly distinguished by features of male internal genitalia.
Descriptions
Stenopelmatus talpa species group
Diagnosis. Members of this group can be distinguished from those of the remaining 3 species groups (Central America, S. piceiventris and S. faulkneri species groups) by having the following morphological features: apterous (generally brachypterous to macropterous in species of the Central America and piceiventris species groups; apterous in species of the faulkneri species group); and head, legs and thorax with a variety of red to orange coloration, abdominal tergites with a dark color, except in S. typhlops which is mostly black (mostly black in species of the faulkneri species group; variable but mainly reddish to orange in species of the remaining 2 species groups). Species of the S. talpa species group appear to be restricted to high elevated areas along the Trans-Mexican Volcanic Belt and adjacent areas of the Sierra Madre Oriental and Sierra Madre del Sur provinces in central Mexico.

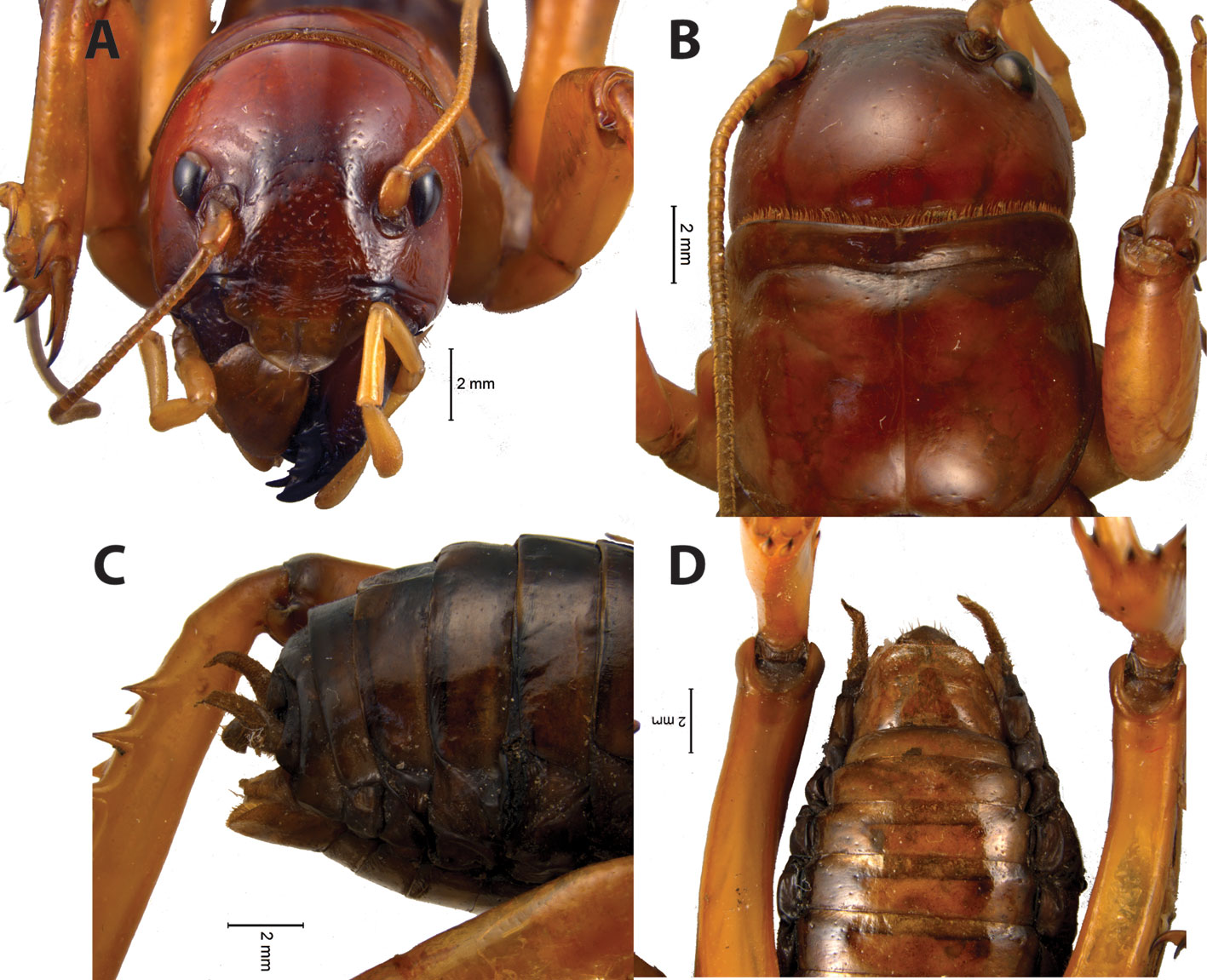
Description. Adults. Usually with large size (females 20.0-50.0 mm; males 19.0-44.0 mm). Body color: head, legs and thorax with different varieties of red to orange color, abdominal tergites black to dark brown, except in S. typhlops, whose main body color is black. Head: hypognate, relatively large compared to body size, with an oval shape that narrows near the jaw area. Antennae longer than body, thin, easily brittle and filiform; scape, pedicel and first flagellomere longer than the rest of the antennal segments; antennal segments with similar shape and size. Vertex strongly convex, fastigium of vertex absent. Eyes small, oval or rhomboid in shape, situated close to and almost at the same level as antennae insertions. Forehead slightly bulging. Labrum with the ability to extend and retract, with ability to hide the jaws in front view. Clypeus rectangular. Thorax: pronotum convex in lateral view, anterior margin slightly concave in dorsal view, with short and thin setae. Lateral lobe with a bisinuate lower edge, longer than wide in lateral view; anterior angle almost right in angle and with a rounded corner. Pronotum with lateral margins smooth in dorsal view, with a deep furrow parallel to the anterior border, with a distinct, shallow furrow running along midline of pronotum. Prosternum with a hexagon-like shape, with a sinking and thick groove that appears to divide it into 2 isosceles trapezoids of different size. Mesosternum shaped in form of a laterally elongated hexagon or trapezoidal. Metasternum shaped in form of a laterally elongated hexagon. Abdomen: oval shaped, slightly elongated. First tergite with modified anterior part, 10th tergite considerably modified, with cerci, supra-anal plate, epiproct, and, in adult males, with black hooks near cerci. Subgenital plate with an “isosceles triangle” shape in ventral view in adult females, and with a “hexagonal” or “trapezoidal” shape in adult males. Males with the 8 sternite having a third, or less the length of subgenital plate, and half or less the length of the seventh sternite. Legs: anterior femora dorsally thickened, with inner side straight and the outer curved; anterior tibiae with 3 spurs and 4 or 5 calcars; middle femora simple; medium tibiae with 8 calcars and 2 spurs; posterior femora thicker than previous ones, with a visible depression along its entire length; posterior tibiae with 5 inner and 3 or 4 outer spines, 6 calcars and 2 spurs. Posterior tibiae with most calcars and internal spines distinctly larger than external ones. Inner portion of posterior and middle calcars flat or slightly concave.

Stenopelmatus talpa Burmeister, 1838
(Figs. 3A, 4A-D, 5A, 6A, B)
Stenopelmatus talpa Burmeister, H. 1838. Handbuch der Entomologie 2 2(I-VIII): 721
Stenopelmatus talpa Graber. 1876[1875]. Denkschr. Akad. Wiss. Wien 36(2): 32
Stenopelmatus talpa Brunner von Wattenwyl. 1888. Verh. der Zoologisch-Botanischen Gesellsch. Wien 38: 260
Stenopelmatus talpa Saussure & Pictet. 1897. Biologia Centrali-Americana 1: 288
Stenopelmatus talpa Rehn, J.A.G. 1901[1900]. Trans. Amer. Entomol. Soc. 27: 222
Stenopelmatus talpa Kirby, W.F. 1906. A Synonymic Catalogue of Orthoptera (Orthoptera Saltatoria, Locustidae vel Acridiidae) 2: 111
Stenopelmatus talpa Karny. 1927. Zeitschr. gesam. Naturwiss. 88: 9
Stenopelmatus talpa Karny. 1928. Stett. Entomol. Z. 89: 249
Stenopelmatus talpa Karny. 1929. Ann. Naturhist. Mus. Wien 43: 40
Stenopelmatus talpa Hebard. 1932. Trans. Amer. Entomol. Soc. 58(3): 342
Stenopelmatus talpa Karny. 1937. Genera Insectorum 206: 45
Stenopelmatus talpa ICZN. 1943. Bull. Zool. Nomencl.
Stenopelmatus talpa ICZN. 1954. Bull. Zool. Nomencl.
Stenopelmatus talpa Weissman. 2001. In Field, L.H. [Ed.]. The Biology of Wetas, King Crickets and their Allies 58
Stenopelmatus talpa Gorochov & Cadena-Castañeda. 2016. Zoosystematica Rossica 25(1): 142
Stenopelmatus talpa Sánchez-Xolalpa, Álvarez, De la Torre-Anzúres & Jiménez-García. 2017. SW Entomol. 42(3): 745-752
Stenopelmatus talpa Weissman, Vandergast, H. Song, Shin, McKenna & Ueshima. 2021. Zootaxa 4917(1): 97
Stenopelmatus talpa Gutiérrez-Rodríguez, Zaldívar-Rive-
rón, Weismann & Vandergast. 2022. Inv. Syst. 36(1): 18.

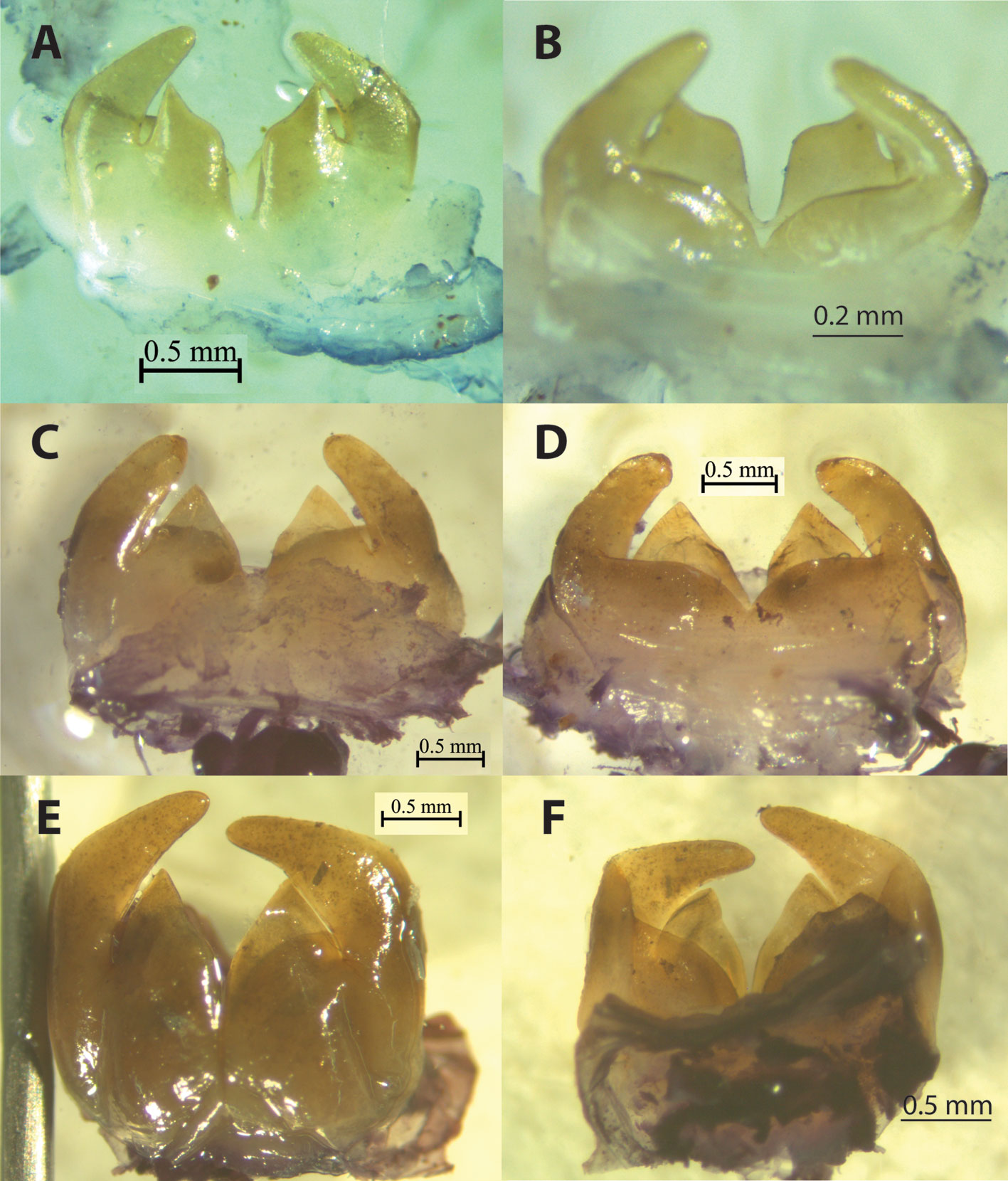
Diagnosis. Adult males and females of S. talpa can be distinguished from the remaining members of the S. talpa species group by having the head, pronotum and legs with a bright reddish-orange color when alive (Fig. 3A) (dark reddish orange to black in the remaining species; Fig. 3B-D). Moreover, adult males of this species can be distinguished from those of the remaining species of the group by having the following combination of features: 1) sternites 8 and 9 (subgenital plate) not overlapping in lateral view (Fig. 4D) (overlapping in the remaining species; Figs. 8C, 10C, 11C, 12C, and 13C, respectively); 2) subgenital plate with a “square-like” shape, and a distal truncated edge in lateral view (Fig. 4D) (similar in S. typhlops, Fig. 8C; “rectangle-like” with a truncated edge in S. sierragordaensis, Fig. 12C; irregular trapezoid-like in S. chilango, S. purepecha and S. tlaxcalli; Figs. 10C, 11C, and 13C, respectively); 3) ventral lobes of male internal genitalia in form of an irregular triangle, considerably concave proximally with a sharp apex in ventral view (Fig. 6A) (equilateral triangle with a sharp apex in S. typhlops and S. sierragordaensis, Fig. 6C, E; isosceles triangle with a sharp apex in S. purepecha, Fig. 7A; irregular triangle, slightly concave proximally with sharp apex in S. chilango, Fig. 7C; sub-rectangular and folded in S. tlaxcalli, Fig. 7E).
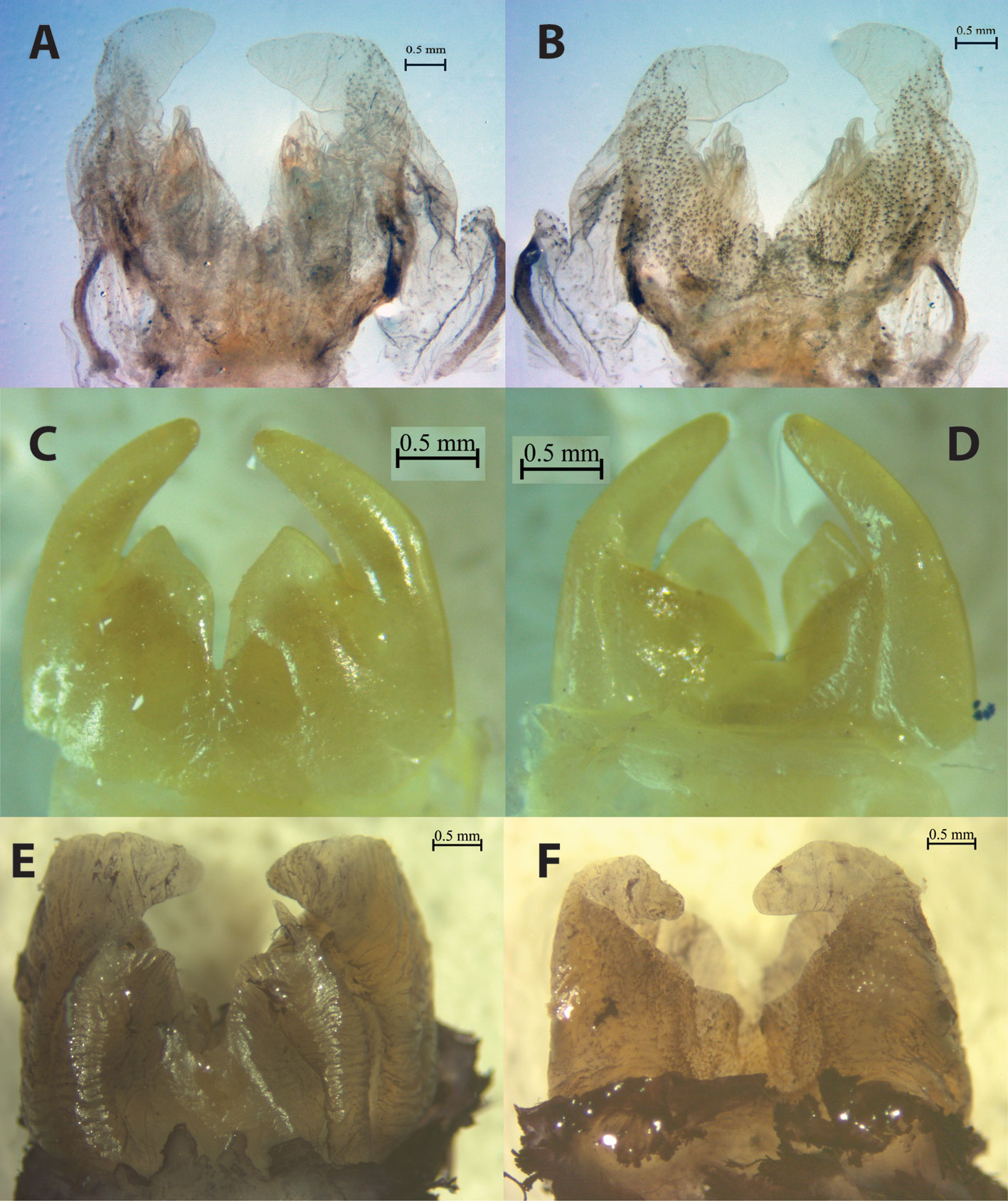
Redescription. Males. Measurements: body length from anterior margin of vertex to end of abdomen 25-31 mm. Pronotum length along midline 5.2-6.0 mm. Anterior, mid and posterior femora length 6.6-7.5, 6.1-6.7 and 11-11.9 mm, respectively. Body color: head, legs and thorax dark reddish orange; tergites dark reddish-orange on basal half, dark brown to black on apical half; abdominal sternites light brown to brown usually with apical, thin, black stripes. The neotype only has abdominal sternites with reddish-brown tones. Head: head hypognate, relatively large compared to body size (0.5 times length of remaining part of body), with an oval shape that narrows near the jaw area. Antennae longer than body, thin, easily brittle, composed of 70 or more segments. Vertex strongly convex. Fastigium of vertex absent. Fastigium frontalis with an “omega-like” pattern (Ω), slightly flattened and with elongated opening lines curved downwards and with the presence of a furrow (Fig. 4A). Eyes small, oval or rhomboid in shape, situated close to and almost at the same level as antennae insertions (Fig. 4A). Forehead slightly bulging; genae with no apparent divisions. Thorax: pronotum convex in lateral view, anterior margin slightly concave in dorsal view, with short and thin setae (Fig. 4B). Lateral lobe with a bisinuate lower edge, greater in length on the horizontal axis, anterior angle almost rectangular and with a rounded corner in lateral view. Pronotum with lateral margins smooth in dorsal view, with a deep furrow parallel to the anterior border, a distinct, shallow furrow running along midline of pronotum, without overlapping with other furrows and becoming diffuse near margin of anterior furrow. Anterior-medial area of the pronotum with a pair of shallow and slightly distinct grooves on each side of the median line and apparently symmetrical; grooves slightly paler and at an approximate angle of 45º. Marked and shallow furrow, parallel to the posterior margin of pronotum, bisinuate rear margin (Fig. 4B). Legs: anterior femora dorsally thickened, with inner side straight and the outer curved; anterior tibiae with 3 spurs and 4 calcars; middle femora flat; medium tibiae with 8 calcars and 2 spurs; posterior femora thicker than remaining ones, laterally with a visible depression along its entire length (Fig. 4D); posterior tibiae with 5 inner and 3 outer spines, 6 calcars and 2 spurs (Fig. 5A). Abdomen (terminalia): cerci thicker basally, tapering towards the apex, ending in a fine point (Fig. 4D), with 2 dark yellow hooks near the cerci. Lateral lobes of internal genitalia visible in ventral and lateral view, near subgenital plate. Supra-anal plate with a “half-moon-like” shape, with its tips pointing towards proximal area. Subgenital plate with “hexagonal-like” shape and slightly elongated laterally in ventral view, “square-like” shape with a distal truncated edge in lateral view (Fig. 3C, D). Male internal genitalia: wider than long (Fig. 6A, B). Ventral lobes in form of an irregular triangle, concave proximally and pointed apically (Fig. 6A); lateral lobes bent inwards almost at a right angle in dorsal view, distally subconical and with rounded apices, proximally broader.
Females. Similar to males. Body length from anterior margin of vertex to end of abdomen 33 mm. Pronotum length along midline 6.4-6.6 mm. Anterior, mid, and posterior femora length 6.7-7.7, 6.5-6.6, and 12.5-12.6 mm, respectively. Distance from apex of ovipositor to apex of subgenital plate 4.0-4.1 mm, in lateral view. Supra-anal plate with “triangle-like” shape in ventral view, proximally concave (Fig. 9D). Subgenital plate with an “isosceles triangle” shape in ventral view, and a “right triangle” shape in lateral view (Fig. 4E, F). Ovipositor strongly curved at a 90 º angle, claw-shaped (Fig. 4F).
Taxonomic summary
Neotype (designated by D. B. Weissman et al., 2021): CAS. ♂ México, Hidalgo, Hwy 85 at km sign post 163.5 toward Zimapán, 03/06/2008, 20°55’42” N, 99°12’29.2” W, S08-39, R08-13 & R08-42. DBW and DC Lightfoot, CAS CASENT type #20086.
Examined material. One adult male, 2 adult females. ♂ México, Hidalgo, Jacala de Ledezma, 01/09/2019, 20°56’54.445” N, 99°12’41.2878” W, K. López-Estrada, M. Suastes-Jiménez & J. Gutiérrez-Rodríguez leg., DNA voucher number CNIN 4571. ♀ México, Hidalgo, Jacala de Ledezma, 01/09/2019, 20°56’55.1796” N, 99°12’42.2346” W, K. López-Estrada, M. Suastes-Jiménez & J. Gutiérrez-Rodríguez leg., DNA voucher number CNIN 4350 (GenBank accession no. MZ313375). ♀ México, Hidalgo, Zimapán, Encarnación, frente a un criadero de truchas, 09/2020, 20°52’26.4828” N, -99°12’37.3608” W, A. Zaldívar-Riverón, I. Garzón, J. Jasso-Martínez leg., DNA voucher number CNIN 4602.
Other examined material. Six nymphs. Nymph, México, Hidalgo, Jacala de Ledezma, 01/09/2019, 20°56’55.1796” N, 99°12’42.2346” W, K. López-Estrada, M. Suastes-Jiménez, J. Gutiérrez-Rodríguez leg., DNA voucher number CNIN 4349. Four nymphs, México, Hidalgo, Jacala de Ledezma, 01/09/2019, 20°56’55.1796” N, 99°12’42.2346” W, K. López-Estrada, M. Suastes-Jiménez, J. Gutiérrez-Rodríguez leg., DNA voucher number CNIN 4523-4526. Nymph, México, Hidalgo, Jacala de Ledezma, 09/2020, 19°56’55.1796” N, 99°12’42.2346” W, A. Zaldívar-Riverón, I. Garzón, J. Jasso-Martínez leg., DNA voucher number CNIN 4603.
Geographic distribution. This species is only known from its type locality and surrounding areas within Los Mármoles National Park in the municipalities of Jacala de Ledezma and Zimapán in north-northwestern Hidalgo, central Mexico.

Remarks
Specimens of this species have been mainly collected walking on the ground or under stones and logs. Specimens at different stages of their development have been found under the same rocks and logs. Weissman et al. (2021) mentioned that this species characterizes by its hopping, medium to large-sized adults, and a drum with soft, short, irregular series (R08-13 and R08-42) of 2 drums over 0.48-0.56s, up to 6 drums, over 1.6s (3.8 d/s), and super series of 11 variable series over 52.7s at 20.5°C.
Stenopelmatus typhlops Rehn, 1903
(Figs. 5E, 6C, D, 8A-D)
Stenopelmatus typhlops Rehn, J.A.G. 1903. Trans. Amer. Entomol. Soc. 29(1): 15
Stenopelmatus typhlops Kirby, W.F. 1906. A Synonymic Catalogue of Orthoptera (Orthoptera Saltatoria, Locustidae vel Acridiidae)
Stenopelmatus nieti Hebard. 1932. Trans. Amer. Entomol. Soc. 58(3): 342
Stenopelmatus typhlops Karny. 1937. Genera Insectorum 206: 1-317
Stenopelmatus typhlops Weissman. 2001. In Field, L.H. [Ed.]. The Biology of Wetas, King Crickets and their Allies 58, 105
Stenopelmatus typhlops Gutiérrez-Rodríguez, Zaldívar-Riverón, Weissman & Vandergast. 2022. Invertebrate Systematics 36(1): 18
Diagnosis. Males. Adult males and females of S. typhlops can be distinguished from those of the remaining species of the group by having a mostly black body color. Adult males of this species can also be distinguished from the remaining species of the group by having the following combination of morphological features: 1) subgenital plate with a “square-like” shape and a distal truncated edge in lateral view (Fig. 8C) (similar in S. talpa, Fig. 4D; “rectangle-like” with a distal truncated edge in S. sierragordaensis, Fig. 12C; irregular “trapezoidal-like” in S. chilango, S. purepecha, and S. tlaxcalli, Figs. 10C, 11C, and 13C, respectively); 2) ventral lobes of male internal genitalia in form of an equilateral triangle with a sharp apex (Fig. 6C, D) (similar in S. sierragordaensis, Fig. 6E, F; isosceles triangle with sharp apex in S. purepecha, Fig. 7A, B; irregular triangle, concave proximally with sharp apically in S. talpa, Fig. 6A, B; irregular triangle, slightly concave proximally with sharp apex in S. chilango, Fig. 7C, D; sub-rectangular and folded in S. tlaxcalli, Fig. 7E, F). Moreover, adult females of this species appear to have a unique feature for the group, the supra-anal plate considerably narrow (Fig. 9B) (moderately narrow in the remaining species; Fig. 9A, C, D).

Redescription. Males. Measurements body length from anterior margin of vertex to end of the abdomen 27 mm. Pronotum length, along midline: 6 mm. Anterior, mid, and posterior femora length: 4.5, 5.0, 9.5 mm, respectively. Body color: head, thorax, abdominal tergites and anterior and mid legs black, abdominal sternites and femora dark red with apical, thin, black stripes, with fine yellow lines at the junction of abdominal tergites 1-7. Head: head hypognate, relatively large (0.4 times length of remaining part of body), with an oval shape that becomes narrower near the jaws’area (Fig. 8A). Antennae long, thin, easily brittle, composed of 80 or more segments. Vertex strongly convex. Fastigium of vertex absent. Fastigium frontalis almost indistinct, with an “omega-like” pattern (Ω), and a wider constriction compared to other species. Eyes relatively small, oval or rhomboid in shape, situated close to and almost at the same level as the antennae insertions (Fig. 8A). Forehead slightly bulging; genae with no apparent divisions. Thorax: pronotum convex or slightly convex in lateral view, anterior margin slightly concave in dorsal view, with short and thin setae. Lateral lobe with a bisinuate lower edge and larger in the horizontal axis; anterior angle almost in a right angle and with a rounded corner in lateral view. Lateral margins of pronotum smooth in dorsal view, with a deep furrow parallel to the anterior border and a marked and shallow furrow running along midline of pronotum, without overlapping with other furrows (Fig. 8B). Anterior-medial part of pronotum with a pair of shallow grooves on each side of the median line and apparently symmetrical; grooves at an approximate 45º angle. Furrow distinct and shallow, parallel to posterior margin of pronotum, with a straight or slightly bisinuate rear margin (Fig. 8B). Legs: anterior femora dorsally thickened, with inner side straight and the outer curved; anterior tibiae with 3 spurs and 4 calcars; middle femora flat; medium tibiae with 8 calcars and 2 spurs; posterior femora thicker than remaining ones, laterally with a visible depression along its entire length; rear tibia with 5 inner and 3 outer spines, 6 calcars and 2 spurs (Fig. 5E). Abdomen (terminalia): cerci proximally thick, tapering towards the apex, ending in a sharp point (Fig. 8C). Two hooks, 1 on each side near the cerci. Supra-anal plate with a “triangle-like” shape, visibly biconcave proximally. Subgenital plate with an irregular “hexagonal-like” shape in ventral view (Fig. 8D), and a “square-like” shape with a distal truncated edge in lateral view (Fig. 8C). Male internal genitalia: wider than long. Ventral lobes in form of an equilateral triangle with a sharp apex; emargination between ventral lobes funnel-shaped, narrower than in S. talpa (Fig. 6C). Lateral lobes bent inwards almost at a right angle, subconical distally, proximally broad and with rounded apices in dorsal view. Distance between the middle area of the lateral lobes comparatively greater than any of the other species, except S. talpa (Fig. 6C, D).
Females. Similar to males. Body length (from anterior margin of vertex to end of abdomen) 28.5 mm. Pronotum length along midline: 6.0 mm. Anterior, mid, and posterior femora length: 4.5, 5.0, 10 mm, respectively. Distance from apex of ovipositor to apex of the subgenital plate in lateral view: 4.0 mm. Supra-anal plate considerably narrow in dorsal view (Fig. 9B). Subgenital plate with an “isosceles triangle” shape in ventral view, with a “right triangle” shape in lateral view (Fig. 8E, F). Ovipositor strongly curved at a 90 º angle, claw-shaped (Fig. 8F).
Taxonomic summary
Examined material. One adult male, 1 adult female. ♂ México, Hidalgo, Zacualtipán, 09/03/2018, 20°38’15.4464” N, 99°10’58.0296” W, C. Pedraza-Lara, J. Gutiérrez-Rodríguez leg., DNA voucher number CNIN 3925 (GenBank accession no. OQ525962). ♀ México, Hidalgo, Zacualtipán, 09/03/2018, 20°38’09.6” N, 98°37’41.16” W, C. Pedraza-Lara, J. Gutiérrez-Rodríguez leg., DNA voucher number CNIN 3983 (GenBank accession no. MZ313424).
Geographic distribution. This species is only known from its type locality and nearby areas within Zacualtipán, in the eastern part of the state of Hidalgo. In the original description this locality was incorrectly written.
Stenopelmatus chilango Suastes-Jiménez, Barrientos-Lozano & Zaldívar-Riverón, sp. nov.
(Figs. 5B, 7C, D, 10A-D)
http://zoobank.org/urn:lsid:zoobank.org:act:B0E9AB8E-
FE1A-40C8-9837-88303FC47BBF
Diagnosis. Adult males of S. chilango can be distinguished from those of the remaining species of the talpa species group by having the following combination of morphological features: 1) subgenital plate with an irregular “trapezoid-like” shape in lateral view (Fig. 10C) (similar in S. purepecha and S. tlaxcalli, Figs. 11C and 13C, respectively; “square-like” with a distal truncated edge in S. talpa and S. typhlops, Figs. 4D and 8C, respectively; “rectangle-like” with a distal truncated edge in S. sierragordaensis, Fig. 12C); and 2) ventral lobes of male internal genitalia in form of an irregular triangle, slightly concave proximally with sharp apex (Fig. 7C, D) (sub-rectangular and folded in S. tlaxcalli (Fig. 7E, F); irregular triangle, considerably concave proximally with sharp apex in S. talpa; Fig. 6A, B; equilateral triangle with a sharp apex in S. typhlops and S. sierragordaensis (Fig. 6C, D and 6E, F, respectively); isosceles triangle with sharp apex in S. purepecha (Fig. 7A, B).
Description. Males. Measurements: body length (from anterior margin of vertex to end of the abdomen): 21-38 mm. Pronotum length, along midline: 4.5-6.4 mm. Anterior, mid, and posterior femora length: 5.3-8.2, 5.0-7.8, 8.2-13 mm, respectively. Body color: head, legs and thorax dark red to dark reddish orange, abdominal tergites black, sternites brown to light brown with a pattern of apical thin, dark brown stripes. Head: head hypognate, relatively large (0.45 times length of remaining part of body), with an oval shape that becomes narrower near the jaws’ area (Fig. 10A). Antennae longer than body, thin, easily brittle, composed of 80 or more segments. Vertex strongly convex. Fastigium of vertex absent. Fastigium frontalis with an “omega-like” pattern (Ω), slightly flattened and with elongated opening lines curved downwards and with the presence of a furrow. Eyes relatively small, oval or rhomboid in shape, situated close to and almost at the same level as the antennae insertions (Fig. 10A). Forehead slightly bulging; genae with no apparent divisions. Thorax: pronotum convex, anterior margin slightly concave in dorsal view, with short and thin setae. Lateral lobe with a bisinuate lower edge and larger in the horizontal axis; the anterior angle almost at right angle and a rounded corner in lateral view. Lateral margins of pronotum smooth in dorsal view, with a deep furrow parallel to the anterior border and marked and shallow furrow running along midline of pronotum, without overlapping with other furrows (Fig. 10B). Anterior-medial part of pronotum with a pair of shallow and poorly distinct grooves, 1 on each side of the median line and apparently symmetrical; grooves at an approximate 45º angle and a small discoloration in the furrows. Furrow distinct and shallow, parallel to posterior margin of pronotum, straight or slightly bisinuate rear margin (Fig. 10B). Legs: anterior femora dorsally thickened, with inner side straight and the outer curved; anterior tibiae with 3 spurs and 5 calcars; middle femora flat; medium tibiae with 8 calcars and 2 spurs; posterior femora thicker than remaining ones, with a lateral depression along its entire length; posterior tibia with 5 inner and 3 outer spines, 6 calcars and 2 spurs. Most calcars and internal spines are significantly larger than calcars and external spines. Abdomen (terminalia): cerci proximally thick, tapering towards the apex, ending in a sharp point (Fig. 10C). Two hooks near the cerci. Supra-anal plate with “triangle-like” shape, proximally concave. Subgenital plate with a very marked, wide keel, with an irregular “hexagon-like” shape in ventral view (Fig. 10D), and an irregular “trapezoid-like” shape in lateral view (Fig. 10C). Male internal genitalia: almost as long as wide. Ventral lobes in form of an irregular triangle, slightly concave proximally with sharp apex (Fig. 7C); emargination between ventral lobes Y-shaped. Lateral lobes bent inward almost at an obtuse angle in ventral view, anterior part subconical, broad at the base, apices rounded (Fig. 7C, D).
Females. Similar to males. Body length (from anterior margin of vertex to end of abdomen): 25-35 mm. Pronotum length, along midline: 5.0-6.8 mm. Anterior, mid and posterior femora length: 6.3-8.4, 5.3-7.8, 9.3-13.4 mm, respectively. Distance from apex of ovipositor to apex of subgenital plate, in lateral view: 1.5-4.0 mm. Color: head, legs and thorax a dark red to dark reddish orange, abdominal tergites black, sternites brown to light brown with a pattern of apical, thin, dark, brown stripes. Supra-anal plate with a “half-moon-like” or “triangle-like” shape (Fig. 9A), proximally concave. Subgenital plate with an “isosceles triangle” shape in ventral view, and a “rectangle triangle” shape in lateral view (Fig. 10E, F). Ovipositor strongly curved at a 90º angle, claw-shaped (Fig. 10F).
Taxonomic summary
Holotype. Adult male. México, CDMX, Copilco Universidad, 19/08/2016, 19°20’15.4464” N, 99°10’58.0296” W, A. Zaldívar-Riverón leg., DNA voucher number CNIN 3918.
Paratypes. Twenty-one adult males and 17 adult females. ♂ México, CDMX, Coyoacán, Villa Panamericana, 28/08/2012, 19°18’23.5008” N, 99°10’29.4996” W, N. Percino leg., DNA voucher number CNIN 3667, GenBank accession no. MZ313403. ♂ México, CDMX, Calle San Apolinar, 15/11/2017, 19°18’24.7680” N, 99°08’57.8940” W, J. Jasso-Martínez leg., DNA voucher number CNIN 4557. ♂ México, CDMX, Coyoacán, Copilco, Av. Universidad 2016, 10/12/2018, 19°20’15.9360” N, 99°10’58.3608” W, A. Zaldívar-Riverón leg., DNA voucher number CNIN 4556. ♂ México, CDMX, Copilco. Avenida Universidad 2016, 20/06/2014, 19°20’15.4464” N, 99°10’58.0296” W, A. Zaldívar-Riverón leg., DNA voucher number CNIN 4541. ♂ México, CDMX, Sede de la Licenciatura en Ciencias Forenses CU, 20/08/2018, 19°19’37.5168” N, 99°10’28.8984” W, C. Pedraza-Lara leg., DNA voucher number CNIN 4547. ♂ México, CDMX, Col. del Valle, 2014, 19°22’41.844” N, 99°10’48.8964” W, DNA voucher number CNIN 4328. ♂ México, CDMX, Deportivo Del. Benito Juárez, 11/10/2016, 19°22’17.0868” N, 99°09’34.3548” W, K. López-Estrada leg., DNA voucher number CNIN 4527. Four ♂ México, CDMX, Sede de la Licenciatura en Ciencias Forenses CU, verano 2017, 19°19’37.5168” N, 99°10’28.8984” W, DNA voucher number CNIN 4550-4553. ♂ México, CDMX, Carretera México-Cuernavaca km. 23.5, 10/1979, 19°14’30.9912” N, 99°10’01.8804” W, F. Figueroa leg., DNA voucher number CNIN 4597. ♂ México, CDMX, Ciudad Universitaria, 29/07/2012, 19°19’09.8364” N, 99°11’43.8036” W, M. García-París & N. Percino leg., CNIN 3915 (GenBank accession no. OQ525957). ♂ México, CDMX, Delegación Coyoacán, Cineteca Nacional, 20/09/2020, 19°21’37.1736” N, 99°09’50.7888” W, A. Zaldívar-Riverón leg., CNIN 4599. ♂ México, CDMX, Alcaldía Benito Juárez, entre calles Palenque y Torres, 15/11/2021, 19°23’19.32” N, 99°09’08.28” W, J. Jasso-Martínez leg., CNIN 4604. ♂ México, Estado de México, municipio Naucalpan de Juárez, colonia Echegaray, calle Fresnos no. 3, 10/10/2017, 19°27’57.1464” N, 99°16’11.0028” W, A. Hernando leg., DNA voucher number CNIN 4534. ♂ México, Estado de México, Ciudad Satélite, 20/10/2020, Jardín de casa. E. Pérez-Silva leg., DNA voucher number CNIN 4514 (GenBank accession no. OQ525955). Two ♂ México, Estado de México, Sierra de Guadalupe, 08-12/2017, 19°36’18.9036” N, 99°07’39.45” W, DNA voucher number CNIN 4591-4592. ♂ México, Estado de México, Calzada, Colonia Calacoaya, Atizapan de Zaragoza, col. Calacoaya, 17/08/2018, 19°31’52.0032” N, 99°14’21.1596” W, A. Almas-Altamirano leg., CNIN 4327 (GenBank accession no. OQ525958). ♂ México, Puebla, Chapulco, 5/11/2016, 18°37’10.9992” N, 97°24’38.0016” W, CNIN 3989 (GenBank accession no. OP778775). ♀ México, CDMX, Coyoacán Av. Universidad 2016 Colonia Copilco, 09/2019, 19°20’15.9360” N, 99°10’58.3608” W, A. Zaldívar-Riverón leg., DNA voucher number CNIN 4531. ♀ México, CDMX, Ajusco, 06/10/2012, 19°12’26.9892” N, 99°15’28.998” W, Z. N. Mora-Ramírez leg., DNA voucher number CNIN 4539. ♀ México, CDMX, Tláhuac. San Antonio Tecomilt, 27/05/2019, 19°13’12.36” N, 98°58’56.28” W, V. de Jesús Bonilla leg., DNA voucher number CNIN 4554. ♀ México, CDMX, Coyoacán Viveros, 19°21’12.5208” N, 99°10’17.2092” W, J. Gutiérrez-Rodríguez leg., DNA voucher number CNIN 4570. ♀ México, CDMX, Iztapalapa, Cerro de la Estrella, 05/11/2017, 19°20’36.6792” N, 99°05’21.0732” W, K. Guevara leg., DNA voucher number CNIN 3669 (GenBank accession no. OP778771). ♀ México, CDMX, Tulyehualco, 12/11/2012, 19°15’31.59” N, 99°00’50.8392” W, DNA voucher number CNIN 3666. ♀ México, CDMX, Coyoacán, 28/10/2018, 19°21’02.1312” N, 99°08’24.198” W, R. MacGregor leg., CNIN 4587. ♀ México, CDMX, Delegación Iztapalapa, Colonia el Molino, 14/11/2005, 19°21’17.1432” N, 99°05’19.0716” W, I. Marín leg., CNIN 4595. ♀ México, CDMX, Coyoacán, Copilco. Av. Universidad 2016, 04/10/2020, 19°20’16.1376” N, 99°10’59.2392” W, A. Zaldívar-Riverón leg., CNIN 4598. ♀ México, CDMX, Azcapotzalco, metro Ferrería, 10/01/2021, 19°29’25.3752” N, 99°10’26.832” W, A. Zaldívar-Riverón leg., CNIN 4601. Three ♀ México, Estado de México, Ciudad Satélite, 20/10/2020, Jardín de casa. E. Pérez-Silva leg., DNA voucher number CNIN 4515-4517 (GenBank accession nos OQ525953-54, 56). ♀ México, Hidalgo, Tlanchinol, La Cabaña, 20°59’25.3392” N, 98°39’46.7388” W, Claudio leg., DNA voucher number CNIN 4154. ♀ México, Hidalgo, Tlanchinol, La cabaña, 20°59’25.3392” N, 98°39’46.7388” W, Claudio leg., DNA voucher number CNIN 4156. ♀ México, Puebla, Chapulco, 05/11/2016, 18°37’10.9992” N, 97°24’38.0016” W, CNIN 3988. ♀ México, Morelos, Tepoztlán, 06/05/2018, 18°59’24” N, 99°06’36” W, J. Gutiérrez-Rodríguez leg., CNIN 3969 (GenBank accession no. OQ525959).
Other examined material. Thirteen nymphs. Nymph, México, CDMX, Mixquic, 04/03/2013, 19°19’46.3332” N, 99°09’32.4252” W, V. de Jesús-Bonilla leg., DNA voucher number CNIN 3914. Nymph, México, CDMX, CU, 29/09/2012, 19°19’37.51” N, 99°10’28.89” W, M. García-París & N. Percino leg., DNA voucher number CNIN 3916 (GenBank accession no. MZ313410). Nymph, México, CDMX, Estacionamiento Instituto de Biología CU, 28/05/2016, 19°19’16.1328” N, 99°11’40.0272” W, M. García-París leg., DNA voucher number CNIN 3917. Nymph, México, CDMX, San Nicolás Totolapan, Ajusco, 16/08/2009, 19°17’33.432” N, 99°14’58.92” W, M. García-París & N. Percino leg., DNA voucher number CNIN 3919. Two nymphs, México, CDMX, El Pedregal de San Ángel, 20/07/1968, 19°18’53.0388” N, 99°11’34.6128” W, DNA voucher number CNIN 3936-3937. Nymph, México, CDMX, Del. Iztapalapa. Col. Lomas Estrella, 05/03/2018, 19°18’58.2264” N, 99°05’25.4148” W, V. Salinas leg., DNA voucher number CNIN 4540. Nymph, México, CDMX, Col. Copilco-Universidad, Av. Universidad 2016, 05/01/2018, 19°36’18.9” N, 99°07’39.4495” W, A. Zaldívar-Riverón leg., CNIN 4558. Two nymphs, México, Hidalgo, Tlanchinol, La cabaña, 20°59’25.3392” N, 98°39’46.7388” W, Claudio leg., DNA voucher number CNIN 4152 (GenBank accession no. OOP699671). Nymph, México, Hidalgo, Tlanchinol, La cabaña, 20°59’25.3392” N, 98°39’46.7388” W, Claudio leg., DNA voucher number CNIN 4155. Nymph, México, Morelos, Santo Domingo de Ocotitlan, 14/10/2017, 19°00’36” N, 99°03’0” W, K. López-Estrada & J. Gutiérrez-Rodríguez leg., DNA vouher no. CNIN3642 (GenBank accession no. MZ313391). Nymph, México, Morelos, Santo Domingo de Ocotitlan, 14/10/2017, 19°00’36” N, 99°03’0” W, K. López-Estrada & J. Gutiérrez-Rodríguez leg., DNA vouher no. CNIN3643 (GenBank accession no. MZ313392).
Geographic distribution. Our records indicate that S. chilango is geographically distributed across the Valley of Mexico, including Estado de México, Puebla, northwestern Morelos and most part of Mexico City. Moreover, there is an isolated record of this species in Tlanchinol, northeastern Hidalgo, which suggests that the distribution of this species extends to the southern part of the Sierra Madre Oriental morphotectonic province.
Etymology. The name of this new species refers to Mexican slang demonym for the inhabitants of Mexico City.
Remarks
This species is frequently found on the ground and under stones and logs. It is easy to find at twilight or after a heavy rain. Some mounted specimens with laterally flattened abdomen have the subgenital plate folded over the keel, giving these individuals a square subgenital plate in ventral view.
Stenopelmatus purepecha Suastes-Jiménez, Barrientos-Lozano, Gutiérrez-Rodríguez & Zaldívar-Riverón, sp. nov.
(Figs. 7A, B, 9C, 11A-D)
http://zoobank.org/urn:lsid:zoobank.org:act:629FDDEA-A806-4D38-B3E4-4FD2B002E08B
Diagnosis. Males. The only adult male that we could examine for this new species can be distinguished from those of the remaining species of the talpa species group by having the combination of the following morphological features: 1) subgenital plate with an irregular “trapezoid-like” shape in lateral view (Fig. 11C) (similar in S. chilango and S. tlaxcalli, Figs. 10C and 13C, respectively; “square-like” with a distal truncated edge in S. talpa and S. typhlops, Figs. 4D and 8C, respectively; “rectangle-like” with a distal truncated edge in S. sierragordaensis, Fig. 12C); and 2) ventral lobes of male internal genitalia in form of an isosceles triangle with a sharp apex in ventral view (Fig. 7A) (with form of an equilateral triangle with a sharp apex in S. typhlops and S. sierragordaensis, Fig. 6C and E, respectively; irregular triangle, concave proximally and sharp apically in S. talpa, Fig. 6A; irregular triangle, slightly concave proximally with sharp apex in S. chilango, Fig. 7C and D, respectively; sub-rectangular and folded in S. tlaxcalli, Fig. 7E, F). Moreover, adult males and females of S. purepecha can also be distinguished from those of the remaining species by having anterior and posterior margin of metasternum about the same size in ventral view (Fig. 11D) (posterior side distinctly shorter than anterior in the remaining species, Figs. 4C, 8D, 10D, 12D, 13D); and abdominal sternites with an uniform dark red color, stripes indistinct (Fig. 11D) (variable in color but always with black or reddish stripes in the remaining species; Figs. 4C, 8D, 10D, 12D, and 13D, respectively).

Description. Males. Measurements. Body length (from anterior margin of vertex to end of the abdomen): 33 mm. Pronotum length, along midline: 5.0 mm. Anterior, mid, and posterior femora length: 8.0, 6.0, 12 mm, respectively. Body color: head, legs and thorax a dark red to dark reddish orange, abdominal tergites black, sternites dark red without stripes. Head: head hypognate, relatively large compared to body size (0.38 times length of remaining part of body), with an oval shape that becomes narrower near the jaws’ area (Fig. 11A). Antennae long, thin, easily brittle, composed of 80 or more segments. Head vertex strongly convex. Fastigium of vertex absent. Fastigium frontalis with an “omega-like” pattern (Ω), slightly flattened and with elongated opening lines curved downwards. Eyes relatively small, oval or rhomboid in shape, situated close to and almost at the same level as the antennae insertions (Fig. 11A). Forehead slightly bulging; genae with no apparent divisions. Thorax: pronotum convex in lateral view, anterior margin slightly concave in dorsal view, with short and thin setae. Lateral lobe with a bisinuate lower edge and larger in the horizontal axis; the anterior angle has almost a right angle and a rounded corner in lateral view. Lateral margins of pronotum smooth in dorsal view, with a deep furrow parallel to the anterior border and marked and shallow furrow running along midline of pronotum, without overlapping with other furrows (Fig. 11B). Anterior-medial part of the pronotum with a pair of shallow grooves, 1 on each side of the median line and apparently symmetrical; grooves at an approximate 45º angle. Furrow distinct and shallow, parallel to posterior margin of pronotum, straight (Fig. 11B). Legs: anterior femora dorsally thickened, with inner side straight and the outer curved; anterior tibiae with 3 spurs and 4 calcars; middle femora flat; medium tibiae with 8 calcars and 2 spurs; posterior femora thicker than remaining ones, with a lateral depression along its entire length; rear tibia with 5 inner and 3 outer spines, 6 calcars and 2 spurs. The left rear tibia of the holotype has a different arrangement of the spines, not observed in any other individual of the species: 5 inner, 2 outer spines and 3 outer small and blunt spines, 6 calcars and 2 spurs (Fig. 5D). Abdomen (terminalia): cerci proximally thick, tapering towards the apex, ending in a fine point (Fig. 11C). Two hooks near the cerci. Lateral lobes of internal genitalia visible from outside, near the subgenital plate in lateral view. Supra-anal plate with a “triangle-like” shape, distinctly biconcave proximally. Subgenital plate with an irregular “hexagon-like” shape in ventral view (Fig. 11D), and an irregular “trapezoid-like” shape in lateral view (Fig. 11C). Male internal genitalia: internal genitalia longer than wide. Ventral lobes in the form of an isosceles triangle with sharp apex in ventral view; emargination between the ventral lobes in a deep V shape (Fig. 7A). Lateral lobes bent inwards almost at a right angle, proximally broad, rounded apices in dorsal view (Fig. 7A). Distance between the lateral lobes at approximately its mid-length smaller than in the remaining species of the S. talpa complex (Fig. 7B).
Females. Similar to males. Body length (from anterior margin of vertex to end of the abdomen): 42 mm. Pronotum length, along midline: 8 mm. Anterior, mid, and posterior femora length: 10, 9.3, 14 mm, respectively. Distance from apex of ovipositor to apex of subgenital plate in lateral view: 14.7 mm. Legs: similar to males. At least one of the rear tibiae has a smaller spine or a cuticular growth that does not become a spine, almost as long as wide with a blunt tip. It is usually present on the outer side of the tibia. Abdomen (terminalia): supra-anal plate with a “half-moon-like” shape, proximally concave (9C). Subgenital plate with an “isosceles triangle” shape in ventral view, and a “right triangle” shape in lateral view (Fig. 11E, F). Ovipositor strongly curved at a 90º angle, claw-shaped (Fig. 11F).

Taxonomic summary
Holotype. Adult male. México, Michoacán, Morelia, Cuanajillo, “El Águila”, 08/09/2013, 19°37’15.1860” N, 101°21’55.98” W, A. Pineda leg. DNA voucher CNIN3933.
Paratypes. One adult female and 2 nymph females. ♀ México, Michoacán, Ario de Rosales, 23/11/2006, 19°12’20.7144” N, 101°42’15.4152” W, J. J. T. C. leg., DNA voucher number CNIN 3934 (GenBank accession no. OP778774). Two nymphs, ♀ ♂, México, Michoacán, Tingambato, 20/12/2017, 19°25’50.0016” N, 101°51’24.9984” W, G. Beltrán & J. Gutiérrez-Rodríguez leg., DNA voucher number CNIN3736 (GenBak accession no. OP778772).
Geographic distribution. Stenopelmatus purepecha occurs in pine and pine-oak forests at elevations close to 2,000 m of altitude (but see below) along the central portion of the state of Michoacán, including localities in Morelia, Ario de Rosales and Tingambato.
Etymology. We named this species after the ethnic group, the Purepechas, that inhabits the Mexican state of Michoacán, where its type material was collected.
Remarks
Our DNA barcoding results indicated that there are 2 species of the S. talpa species group in Michoacán. Our species assignment to S. purepecha of the adult male holotype and the sequences adult female paratype (CNIN3736) was based on 2 shared external morphological features, anterior and posterior margin of metasternum about the same size (Fig. 11D) and the abdominal sternites with a uniform dark red color, without distinct stripes. These 2 features are not present in the nymphs from Tingambato, Michoacán, one of which was sequenced and was genetically similar to the above female paratype. We could not describe the second species from Michoacán (Stenopelmatus sp. 1) since we only had an adult female and some nymphs with a different degree of development. Further collect of adult males will help to describe this species. Apparently, there is not a clear geographic pattern between the distribution of the above 2 species. A male nymph (CNIN 3741) that was genetically (COI) similar to the specimens belonging to the undescribed species from the state of Michoacán (Stenopelmatus sp. 1) was collected in a much lower altitude near Apatzingán (300 m of altitude). Further collects of adult specimens from this locality will help to elucidate the species assignment of this population.
Stenopelmatus sierragordaensis Suastes-Jiménez, Barrientos-Lozano, Gutiérrez-Rodríguez & Zaldívar-Riverón, sp. nov.
(Figs. 5F, 6E, F, 12A-D)
http://zoobank.org/urn:lsid:zoobank.org:act:8D293ECA-052C-4971-84C1-F4CF8090120F
Diagnosis. Males of S. sierragordaensis can be distinguished from the remaining species of the talpa species group by having the following combination of morphological features: 1) abdominal sternites dark brown with a black stripe pattern (Fig. 12D) (dark red with black stripes in S. typhlops, Fig. 8D; dark red without stripes in S. purepecha, Fig. 11D; light brown to brown usually with stripes in S. talpa, Fig. 4C; brown to light brown with dark stripes in S. chilango and S. tlaxcalli, Figs. 10D and 13D, respectively); 2) ventral lobes of male internal genitalia in form of equilateral triangle with a sharp apex (Fig. 6E) (similar in S. typhlops, Figs 6C, D; irregular triangle, considerably concave proximally with a sharp apex in S. talpa, Fig. 6A, B; isosceles triangle with a sharp apex in S. purepecha, Fig. 7A, B; irregular triangle, slightly concave proximally with sharp apex in S. chilango, Fig. 7C, D; sub-rectangular and folded in S. tlaxcalli, Figs. 7E, F); 3) subgenital plate with a “rectangular-like” shape and a truncated edge in lateral view (Fig. 12C) (with a “square-like” shape and a truncated corner in S. talpa and S. typhlops, irregular trapezoid-like in S. chilango, S. purepecha, and S. tlaxcalli, Figs. 10C, 11C, and 13C, respectively).
Description. Males. Measurements. Body length (from anterior margin of vertex to end of the abdomen): 29.5 mm. Pronotum length, along midline: 6.0 mm. Anterior, mid, and posterior femora length: 7.1, 7.5, 12.3 mm, respectively. Body color: head, legs and thorax dark red to dark reddish orange, abdominal tergites black, sternites dark brown with apical, thin, black stripes. Head: head hypognate, relatively large compared to the size of the body (0.35 times length of remaining part of body), with an oval shape that becomes narrower near the jaws’area (Fig. 12A). Antennae long, thin, easily brittle, composed of 75 or more segments. Vertex strongly convex. Fastigium of vertex absent. Fastigium frontalis with an “omega-like” pattern (Ω), slightly flattened and with elongated opening lines curved downwards. Eyes relatively small, oval or rhomboid in shape, situated close to and almost at the same level as the antennae insertions (Fig. 12A). Forehead slightly bulging; genae with no apparent divisions. Thorax: pronotum almost straight or very slightly convex in lateral view, anterior margin slightly concave in dorsal view, with short and thin setae. Lateral lobe with a bisinuate lower edge and larger in the horizontal axis; anterior angle with almost having a right angle, and with a rounded corner in lateral view. Lateral margins of pronotum smooth in dorsal view, with a deep furrow parallel to the anterior border and marked and shallow furrow running along midline of pronotum, without overlapping with other furrows (Fig. 12B). Anterior-medial part of the pronotum with a pair of shallow grooves, 1 on each side of the median line and apparently symmetrical; grooves at an approximate 45º angle. Furrow distinct and shallow, parallel to posterior margin of pronotum, slightly bisinuate rear margin (Fig. 12B). Legs: anterior femora dorsally thickened, with inner side straight and the outer curved; anterior tibiae with 3 spurs and 4 calcars; middle femora flat; medium tibiae with 8 calcars and 2 spurs; posterior femora thicker than remaining ones, with a lateral depression along its entire length; rear tibia with 5 inner and 3 outer spines, 6 calcars and 2 spurs (Fig. 5F). Abdomen (terminalia): cerci proximally thicker, tapering toward the apex, ending in a fine point. Lateral lobes of male internal genitalia protruding and visible in dorsal and lateral view (Fig. 12C). Supra-anal plate with a “half-moon-like” shape, with its tips pointing proximally. Subgenital plate with an irregular “hexagon-like” shape, or an isosceles trapeze with twisted lateral sides, in ventral view (Fig. 12D), and truncated “rectangle-like” shape in lateral view (Fig. 12C). Male internal genitalia: almost as long as wide. Ventral lobes in form of an equilateral triangle with a sharp apex in ventral view; emargination between ventral lobes of male internal genitalia with a V shape (Fig. 6E). Lateral lobes bent inwards almost at an obtuse angle in dorsal view (Fig. 6E, F).
Females. Similar to males. Body length (from anterior margin of vertex to end of the abdomen): 34.5 mm. Pronotum length, along midline: 6.3 mm. Anterior, mid, and posterior femora length: 8.6, 8.3, 14.7 mm, respectively. Distance from apex of ovipositor to apex of subgenital plate in lateral view: 4.2 mm. Supra-anal plate with a “half-moon-like” shape, proximally concave. Subgenital plate with an “isosceles triangle” shape in ventral view, with a “right triangle” shape in lateral view (Fig. 12E, F). Ovipositor strongly curved at a 90º angle, claw-shaped (Fig. 12F).

Taxonomic summary
Holotype. Adult male. México, Querétaro, Km 5 desvío a el Madroño, Valle de Guadalupe, 15/06/2013, 21°17’59.6148” N, 99°10’40.3572” W, A. Zaldívar-Riverón, H. Clebsch, C. Pedraza-Lara & G. Aguilar leg., DNA voucher number CNIN 3646 (GenBank accession no. OP778768).
Paratype. One adult female. ♀ México, Querétaro, el Pilón, Estación Valle Verde, 31/07/2019, K. López-Estrada, J. Gutiérrez-Rodríguez, M. Suates-Jiménez leg., DNA voucher number CNIN 4348.
Geographic distribution. This species is only known to occur in pine-oak forest areas in the northeast portion of the Sierra Gorda Biosphere Reserve in the estate of Querétaro, Mexico.
Etymology. We named this species after the name of the natural protected area where it occurs in the state of Querétaro, the Sierra Gorda Biosphere Reserve.
Stenopelmatus tlaxcalli Suastes-Jiménez, Barrientos-Lozano & Zaldívar-Riverón, sp. nov.
(Figs. 5C, 7E, F, 13A-D)
http://zoobank.org/urn:lsid:zoobank.org:act:BC964E86-37EC-4E9F-9BC5-0B20712F14C5
Diagnosis. Adult males of S. tlaxcalli can be distinguished from the remaining members of the S. talpa species group by: 1) presence of 4 well-developed, sharp external spines on at least one of the posterior tibiae (usually 3 well-developed, sharp external spines in the remaining species, if there are 4 spines, then the fourth is always blunt); and 2) ventral lobes of male internal genitalia sub-rectangular and folded (Fig. 7E) (irregular triangle, slightly concave proximally with sharp apex in S. chilango, Fig. 7C; irregular triangle, considerably concave proximally with a sharp apex in S. talpa, Fig. 6A; equilateral triangle with a sharp apex in S. typhylops and S. sierragordaensis, Fig. 6C and E, respectively; isosceles triangle with a sharp apex in S. purepecha, Fig. 7A).
Description. Males. Measurements: body length (from anterior margin of vertex to end of the abdomen): 33-35.2 mm. Pronotum length: 8.0 mm. Anterior, mid and posterior femora length: 9.5-10, 9, 14 mm, respectively. Body color: head, legs and thorax dark reddish orange; abdominal tergites black, sternites with a pattern of reddish or reddish-yellow and black stripes; black, thin stripes, defined and regular in size. Head: hypognate, relatively large compared to the size of the body (0.45 times length of remaining part of body), with an oval shape that becomes narrower near the jaws’area (Fig. 12A). Antennae almost as long as body, thin, easily brittle, slender and filamentous; composed of 80 or more segments. Vertex strongly convex. Fastigium of vertex absent. Fastigium frontalis with an “omega-like” pattern (Ω), slightly flattened and with elongated opening lines curved downwards and with the presence of a furrow (Fig. 13A). Eyes relatively small, oval or rhomboid in shape, situated close to and almost at the same level as the antennae insertions (Fig. 13A). Thorax: pronotum convex in lateral view, anterior margin slightly concave in dorsal view, with short and thin setae. Lateral lobe with a bisinuate lower edge and larger in the horizontal axis; the anterior angle has almost a right angle and a rounded corner in lateral view. Lateral margins of pronotum smooth in dorsal view, with a deep furrow parallel to the anterior border and marked and shallow furrow running along midline of pronotum, without overlapping with other furrows (Fig. 13B). Anterior-medial part of the pronotum with a pair of shallow and poorly distinct grooves on each side of the median line and apparently symmetrical; grooves at an approximate 45º angle and a small discoloration in the furrows. Furrow distinct and shallow, parallel to posterior margin of pronotum, straight. Legs: anterior femora dorsally thickened, with inner side straight and the outer curved; anterior tibiae with 3 spurs and 4 calcars; middle femora flat; medium tibiae with 8 calcars and 2 spurs; posterior femora thicker than remaining ones, with a lateral depression along its length; posterior tibiae with 5 inner and 3 or 4 outer spines, 6 calcars and 2 spurs. Most calcars and internal spines are significantly larger than calcars and external spines (Fig. 5C). Abdomen (terminalia): cerci proximally broad, tapering towards the apex. A pair of hooks near the cerci, in the proximal area of the supra-anal plate (Fig. 13D). Supra-anal plate with a “triangle-like” shape, proximally concave (Fig. 13D). Subgenital plate with a very marked keel, irregular hexagon-like shape in ventral view, and an irregular “trapezoid-like” shape in lateral view (Fig. 13C, D). Male internal genitalia: almost as long as wide. Ventral lobes sub-rectangular, folded in ventral view. Emargination between the ventral lobes with a “U”-shape in ventral view (Fig. 7E). Lateral lobes bent inward almost at a right angle, the apical portion rounded in dorsal view (Fig. 7F).
Females. Unknown.
Taxonomic summary
Holotype. Adult male. México, Tlaxcala, 50 m del Río Zahuapán, 2,234 m, 05/08/2021, 19°19’21” N, 98°14’38.688” W, DNA voucher number CNIN 4608, GenBank accession no. OQ525960.
Paratype. One adult male. ♂ México, Tlaxcala, 50 m del Río Zahuapán, 2,234 m of altitude 05/08/2021, 19°19’21” N, 98°14’38.688” W, DNA voucher number CNIN 4609 (GenBank accession no. OQ525961).
Other examined material. Nymph, México, Estado de México, Nepantla, Tepetlixpa, 2017, 18°58’49” N, 98°50’0.2688” W, DNA voucher number CNIN 3651 (GenBank accession no. MZ313398).
Geographic distribution. The only records for this species include a locality near the Zahuapan River in the state of Tlaxcala, and another in Nepantla, municipality of Tepetlixpa, in the Estado of México.
Etymology. The name of this species refers to the Nahuatl name for tortilla, and also refers to the Mexican state of its type locality, Tlaxcala.
Remarks
Specimens of S. tlaxcalli have been collected at approximately 2,000 m of altitude. Adults of this species are morphologically similar to those of S. talpa, though they consistently differ genetically (COI) and in their male internal genitalia.
Other examined material
Stenopelmatus barcode sp. 1. Two adult females and 4 nymphs. ♀ México, Michoacán, Morelia, Ichaqueo, 08/05/2014, 19°34’20.514” N, 101°08’25.6416” W, L. Escalante-Jiménez leg., DNA voucher number CNIN 3737 (GenBank accession no. MZ313405). Nymph, México, Michoacán, Apatzingán, 29/07/2009, 19°05’01.5072” N, 102°21’13.0716” W, M. Rosas-Anaya leg., DNA voucher number CNIN 3741 (GenBank accession no. OP429107). ♀ México, Michoacán, Nuevo Parangaricutiro, carretera al Paracutín, cerca de San Juan Nuevo, municipio de Nuevo Parangaricutiro, 12/07/2009, 19°26’50.1432” N, 102°09’27.486” W, DNA voucher number CNIN 3744. Nymph, México, Michoacán, Nuevo Parangaricutiro, San Juan Nuevo, Bosque Certificado San Juan Nuevo, municipio de Nuevo Parangaricutiro, 12/07/2009, 19°24’56.5308” N, 102°07’50.61” W, J. Ponce leg., DNA voucher number CNIN 3931. Nymph, México, Michoacán, Nuevo Parangaricutiro, San Juan Nuevo, Bosque Certificado San Juan Nuevo, municipio de Nuevo Parangaricutiro, 12/07/2009, 19°24’56.5308” N, 102°07’50.61” W, J. Ponce leg., DNA voucher number CNIN 3932.
Stenopelmatus barcode sp. 2. Nymph ♀, México, Veracruz, Orizaba, 25/01/2016, 18°51’38.3” N, 97°07’42” W, E. Recuero leg., CNIN 3639 (GenBank accession no. OP778767).
Stenopelmatus barcode sp. 3. Two adult females and 3 nymphs. ♀ México, Puebla, Zacapoaxtla, Cascada La Gloria, 19/10/2017, 19°54’30.8016” N, 97°37’04.1016” W, A. Zaldívar-Riverón & J. Gutiérrez-Rodríguez leg., DNA vouher no. CNIN 3660. ♀ México, Puebla, San Juan, Libres, 25/11/2017, 19°27’06.9192” N, 97°42’19.4112” W, L. Herrera leg. GenBank accession no. MZ313392, CNIN 3655. Nymph, México, Puebla, Zacapoaxtla, Cascada La Gloria, 19/10/2017, 19°54’30.8016” N, 97°37’04.1016” W, A. Zaldívar-Riverón & J. Gutiérrez-Rodríguez leg., CNIN 3661. Nymph, México, Puebla, Zacapoaxtla, Cascada La Gloria, 19/10/2017, 19°54’30.8016” N, 97°37’04.1016” W, A. Zaldívar-Riverón & J. Gutiérrez-Rodríguez leg., CNIN 3662 (GenBank accession no. OP699674). Nymph, México, Puebla, Xochiapulco, Cascada la Gloria Atzaban, 19/10/2017, 19°54’30.8016” N, 97°37’04.1016” W, DNA voucher no. CNIN 4528.
Key to described species belonging to the S. talpa species group.
1. Adults with mostly black body color…………………………………….. S. typhlops Rehn
– Adults with reddish to orange thorax, legs and head, black to dark red abdominal tergites…………………………………….. 2
2. Adult males with sternites 8 and 9 (subgenital plate) not overlapping in lateral view; subgenital plate with “square-like” shape and truncated distal truncated edge in lateral view, slightly elongated laterally in ventral view. Ventral lobes of male internal genitalia in form of an irregular triangle, considerably concave proximally with a sharp apex in ventral view…………………………………….. S. talpa Burmeister
– Adult males with sternites 8 and 9 overlapping in lateral view; subgenital plate rectangular or trapezoidal-shaped in lateral view (visibly longer than wide); not elongated laterally in ventral view. Ventral lobes of male internal genitalia never in form of an irregular triangle nor considerably concave…………………………………….. 3
3. Subgenital plate of adult males with a “rectangular-like” shape and a truncated edge in lateral view. Ventral lobes of male internal genitalia in the shape of an equilateral triangle with a sharp apex…………………………………….. S. sierragordaensis Suastes-Jiménez, Barrientos-Lozano, Gutiérrez-Rodríguez & Zaldívar-Riverón sp. nov.
– Subgenital plate of adult males with an irregular trapezoid-like shape and a truncated edge in lateral view. Ventral lobes of male internal genitalia without an equilateral triangle-like shape nor a sharp apex…………………………………….. 4
4. Abdominal sternites of adults with a uniform dark red color, stripes indistinct. Posterior margin of metasternum of adults about the same size in ventral view…………………………………….. S. purepecha Suastes-Jiménez, Barrientos-Lozano, Gutiérrez-Rodríguez & Zaldívar-Riverón sp. nov.
– Abdominal sternites of adults with black or reddish stripes. Posterior margin of metasternum of adults distinctly shorter than anterior…………………………………….. 5
5. Adult males with 4 well-developed, sharp external spines on at least one of the posterior tibiae. Ventral lobes of male internal genitalia sub-rectangular and folded…………………………………….. S. tlaxcalli Suastes-Jiménez, Barrientos-Lozano & Zaldívar-Riverón sp. nov.
– Adult males with 3 well-developed, sharp external spines. Ventral lobes of male internal genitalia in form of an irregular triangle, slightly concave proximally with sharp apex…………………………………….. S. chilango Suastes-Jiménez, Barrientos-Lozano & Zaldívar-Riverón sp. nov.
Discussion
Despite being commonly known in Mexico by the local people where it occurs, the taxonomy of genus Stenopelmatus, as currently known, has been largely overlooked. This study represents an important contribution to formally describe the species diversity within one of its recently proposed species groups, the S. talpa species group. This monophyletic group is distributed along the Trans-Mexican Volcanic Belt and adjacent morphotectonic provinces, where it apparently originated and subsequently diversified following an east-central pattern, having its earliest divergence during the late Pliocene to early Pleistocene 3.55 to 1.58 Mya (Ruiz-Mendoza et al., 2023).
Our study confirmed that the members of the S. talpa species group possess a considerably conserved external morphology, with the color pattern and male and female terminalia being the only character systems with species-level diagnostic features. In contrast, the male internal genitalia, which has been traditionally neglected within Stenopelmatidae, provided most of the morphological features that helped to discriminate species in the group. Our results therefore highlight the taxonomic importance of this structure for Stenopelmatus and probably the family in general. Previous to our study, only the internal male genitalia of S. sartorianus Saussure was described and illustrated in detail (Gorochov, 2021). According to this figure, the male genitalia of this species appears to be similar to those observed within the S. talpa species group. Further work is necessary to reveal the existence of variation in the latter character system at different taxonomic levels within the Stenopelmatidae.
The uncorrected COI pairwise distances observed between the species of the S. talpa species group were generally higher than 2% except for 3 cases of specimens belonging to S. purepecha, S. talpa, and S. tlaxcalli, though 2 of them were only marginally lower (1.9%). Thus, our results support the use of the DNA barcode as a complementary tool to delimit species within Orthoptera, though it is important to consider that some phenomena such as incomplete lineage sorting and hybridization events can lead to taxonomic incongruence (Hawlitschek et al., 2017).
Due to their fossorial habits, members of Stenopelmatus are difficult to observe in the field, and thus the knowledge of their biology and ecology is very poor. Weissman et al. (2021) proposed the calling drums as an important feature to delimit species in Stenopelmatus. Unfortunately, we were not able to record calling drums for the specimens that we collected. Weissman et al. (2021) reported the calling drum of the neotype (adult male) of S. talpa, which consisted of short, irregular series of 2 to 6 drums over 1.6 s. Additional collects of specimens from the different species belonging to the S. talpa species group will help to know the interspecific variation of this behavioral feature.
Acknowledgments
We thank S. Guzmán for taking all the digital pictures; G. Aguilar, A. Almas-Altamirano, G. Beltrán, H. Clebsch, L. Escalante-Jiménez, F. Figueroa, M. García-París, K. Guevara, A. Hernando, L. Herrera, V. de Jesús Bonilla, K. López-Estrada, I. Marín, R. MacGregor, N. Mora-Ramírez, C. Pedraza-Lara, N. Percino, E. Pérez-Silva, A. Pineda, J. Ponce, E. Recuero, M. Rosas-Anaya, and V. Salinas for helping to collect some of the examined specimens; J. E. Uribe Arboleda for designing the primers for the S. talpa species group; the community of La Encarnación (Zimapán, Hidalgo) for their assistance during our fieldtrips to this locality; L. Márquez, N. López and A. Jiménez for their assistance in the laboratory (LANABIO); and Cristina Mayorga for her assistance at the CNIN IB-UNAM. M. Suastes-Jiménez was supported by a MSc scholarship given by the Consejo Nacional de Ciencia y Tecnología (Conacyt, Mexico). Thanks to the Posgrado en Ciencias Biológicas, UNAM, for its support during his MSc studies. A. Zaldívar-Riverón was funded by grants given by UNAM-DGAPA (PAPIIT Convocatoria 2019, Proyecto IN201119; Convocatoria 2022, Proyecto IN201622), to the Dirección General de Cómputo y de Tecnologías de Información y Comunicación, Universidad Nacional Autónoma de México (DGTIC, UNAM; LANCADUNAM- DGTIC-339), and the Consejo Nacional de Ciencia y Tecnología (Conacyt), México (convocatoria Ciencia de Frontera, Proyecto number 58548). J. Gutiérrez-Rodríguez was supported by a “Doctores Junta de Andalucía” postdoctoral fellowship (DOC_00668, FEDER EU/Consejería de Economía, Conocimiento, Empresas y Universidad, Junta de Andalucía).
References
Burmeister, H. (1838). Handbuch der Entomologie. Besondere Entomologie; Kaukerfe, Gymnognatha (vulgo:Orthoptera), 2, 721.
De Jesús-Bonilla, V. S., Barrientos-Lozano, L., & Zaldívar-Riverón, A. (2017). Sequence-based species delineation and molecular phylogenetics of the transitional Nearctic–Neotropical grasshopper genus Taeniopoda (Orthoptera, Romaleidae). Systematics and Biodiversity, 15, 600–617. http://dx.doi.org/10.1080/14772000.2017.1313792
Gorochov, A. V. (2021). The Families Stenopelmatidae and Anostostomatidae (Orthoptera). 1. Higher classification, new and little known taxa. Entomological Review, 100, 1106–1151. https://doi.org/10.1134/S0013873820080084
Gutiérrez-Rodríguez, J., Zaldívar-Riverón, A., Weissman, D. B., & Vandergast, A. G. (2022). Extensive species diversification and marked geographic phylogenetic structure in the Mesoamerican genus Stenopelmatus (Orthoptera: Stenopelmatidae: Stenopelmatinae) revealed by mitochondrial and nuclear 3RAD data. Invertebrate Systematics, 36, 1–21. https://doi.org/10.1071/IS21022
Hebert, P. D., Cywinska, A., Ball, S. L., & deWaard, J. R. (2003). Biological identifications through DNA barcodes. Proceedings of the Royal Society of London. Series B: Biological Sciences, 270, 313–321. https://doi.org/10.1098/rspb.2002.2218
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S. et al. (2012). Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics, 28, 1647–1649. https://doi.org/10.1093/bioinformatics/bts199
Padial, J. M., Miralles, A., De la Riva, I., & Vences, M. (2010). The integrative future of taxonomy. Frontiers in Zoology, 7, 16. https://doi.org/10.1186/1742-9994-7-16
Rocha-Sánchez, A. Y., Almaguer-Sierra, P., & Barrientos-Lozano, L. (2018). Procedimiento básico para la extracción y preparación de genitalia interna en Tettigoniidae (Orthoptera: Ensifera). Tecnointelecto, 15, 11–18.
Ruiz-Mendoza, P. X., Jasso-Martínez, J. M., Gutiérrez-Rodríguez, J., Samacá-Sáenz, E., & Zaldívar-Riverón, A. (2023). Mitochondrial genome characterization and mitogenome phylogenetics in the central Mexican Stenopelmatus talpa complex (Orthoptera: Stenopelmatidae: Stenopelmatini). Revista Mexicana de Biodiversidad, 94, e945094. https://doi.org/10.22201/ib.20078706e.2023.94.5094
Sánchez-Xolalpa, D. A., Álvarez, H. A., De la Torre-Anzúres, J., & Jiménez-García, D. (2017). Morphometry, behavior, and ecology of the Jerusalem cricket, Stenopelmatus talpa. Southwestern Entomologist, 42, 745–752. https://doi.org/10.3958/059.042.0313
Swofford, D. L. (2002). PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4. Sinauer Associates, Sunderland, Mass.
Weissman, D. B. (2001). Chapter 3: North and Central America Jerusalem crickets (Orthoptera: Stenopelmatidae): taxonomy, distribution, life cycle, ecology and related biology of the American species. In Field, L. H. (Eds.), The biology of wetas, king crickets and their allies (pp. 57–72). New York: CABI International.
Weissman, D. B. (2005). Jerusalem! Cricket? (Orthoptera: Stenopelmatidae: Stenopelmatus); origins of a common name. American Entomologist, 51, 138–139. https://doi.org/10.1093/ae/51.3.138
Weissman, D. B., Vandergast, A. D., & Ueshima, N. (2008). Jerusalem Crickets (Orthoptera: Stenopelmatidae). In J. L. Capinera (Ed.), Encyclopedia of Entomology (pp. 2054–2061). Dordrecht: Springer. https://doi.org/10.1007/978-1-4020-6359-6_1650
Weissman, D. B., Vandergast, A. G., Song, H., Shin, S., McKenna, D. D., & Ueshima, N. (2021). Generic relationships of New World Jerusalem crickets (Orthoptera: Stenopelmatoidea: Stenopelmatinae), including all known species of Stenopelmatus. Zootaxa, 4917, 1–122. https://doi.org/10.11646/zootaxa.4917.1.1


