
Mónica Orduña-Villaseñor a, David Valenzuela-Galván b,
Jorge E. Schondube a, *
a Universidad Nacional Autónoma de México, Instituto de Investigaciones en Ecosistemas y Sustentabilidad, Campus Morelia, Antigua carretera a Pátzcuaro Núm. 8701, Col. Ex-Hacienda de San José de la Huerta, 58190 Morelia, Michoacán, México
b Universidad Autónoma del Estado de Morelos, Centro de Investigación en Biodiversidad y Conservación, Av. Universidad Núm. 1001, Col. Chamilpa, 62209 Cuernavaca, Morelos, México
*Autor para correspondencia: chon@cieco.unam.mx (J.E. Schondube)
Recibido: 12 noviembre 2021; aceptado: 23 septiembre 2022
Resumen
Los gatos y perros domésticos son las mascotas preferidas del hombre, por lo que se han convertido en especies invasoras y los carnívoros más abundantes del planeta. A pesar de su buena relación con nuestra especie, tienen un impacto ecológico negativo con consecuencias para las políticas de conservación. Los efectos que estos animales tienen sobre la fauna silvestre han sido estudiados principalmente en países con baja biodiversidad de vertebrados (EUA, Nueva Zelanda y Europa), y salvo por algunos trabajos realizados en Australia, Brasil, y México, no contamos con información sobre sus efectos en países con alta biodiversidad. Los efectos negativos que ambas especies tienen sobre la biodiversidad incluyen la depredación y competencia con fauna nativa, su hibridación con especies filogenéticamente cercanas o poblaciones silvestres de su propia especie, y la transmisión de enfermedades a la fauna silvestre y a los humanos. La falta de información sobre los efectos de depredadores introducidos a nivel continental y la falsa idea de que los gatos y perros son ambientalmente inocuos, vuelve crucial aumentar la investigación sobre estas especies en países megadiversos para proponer estrategias de manejo que fomenten la conservación de fauna nativa.
Palabras clave: Especies invasoras; Manejo de mascotas; Carnívoros; Conservación de vertebrados; Animales domésticos; Animales ferales
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Your best friends can be your worst enemies: impact of domestic cats and dogs in megadiverse countries
Abstract
Domestic cats and dogs are the most beloved human pets. As a result, they have become globally invasive species and are the most abundant carnivores in the world. Although they have a good relationship with humans, domestic cats and dogs have a negative ecological impact with significant consequences for conservation policies worldwide. The effects of these animals on wildlife have been studied only in a few countries, with most studies carried out in countries or regions with low vertebrate biodiversity (USA, New Zealand, and Europe). Except for a few studies carried out in Australia, Brazil and Mexico, we have no information on the effects of these predators in megadiverse countries. The main negative environmental effects of both species include predation of vertebrates and invertebrates, competition for food and space with native fauna, hybridization with phylogenetically close species or wild populations of their species, and transmission of diseases to wildlife and humans. The lack of information on the effects that introduced predators have at continental scales and the false perception of the human population that cats and dogs do not adverse impacts on the environment difficult their management and control. It is crucial to increase the number of studies of these two species in megadiverse countries to propose adequate management strategies that promote the conservation of native fauna.
Keywords: Invasive species; Pet management; Carnivores; Vertebrate conservation; Domestic animals; Feral animals
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Introducción
Los gatos (Felis silvestris catus) y los perros domésticos (Canis lupus familiaris) son las mascotas favoritas del humano. La domesticación de los gatos tuvo su origen en Asia Menor y Egipto hace unos 9,000-10,000 años (Driscoll et al., 2007), mientras que la de los perros ocurrió en Europa y el este de Asia hace unos 40,000 años aproximadamente (Frantz et al., 2016). Los gatos fueron domesticados con el objetivo de eliminar roedores y otras plagas de casas y graneros (Driscoll et al., 2007, 2009; Frantz et al., 2016; Vigne, 2004), mientras que los perros han sido utilizados como animales de compañía, protección, pastoreo y caza (Driscoll et al., 2009; Frantz et al., 2016; Gompper, 2014a). En la actualidad, su cercana asociación con los humanos los ha convertido en los carnívoros más abundantes del planeta (APPA, 2017; Peterson et al., 2012; Wierzbowska et al., 2016).
Su expansión por el mundo es un reflejo de los movimientos de las poblaciones humanas en los últimos 15,000 años. Por el papel que estas especies juegan como animales domésticos y por sus diferentes capacidades de dispersión, la expansión de los rangos de distribución de gatos y perros ha variado en el tiempo y el espacio. Los perros acompañaron a los humanos en sus patrones migratorios tempranos, llegando al este de Asia hace unos 12,500 años, a Norteamérica hace unos 8,500 años (Jensen, 2007), a Australia hace unos 5,000 años, a América Central y del Sur hace aproximadamente 3,500 años (Gompper, 2014a) y a Madagascar hace menos de 2,500 años (Jensen, 2007). Mientras que los gatos han tenido una expansión más reciente, moviéndose con los colonizadores europeos a diferentes partes del mundo a partir del siglo XV. De este modo los gatos probablemente llegaron al Nuevo Mundo en el segundo viaje de Colón entre 1493 y 1495 (Frantz et al., 2016; Marra y Santella, 2016). A partir de la llegada de ambas especies a las diferentes regiones del planeta, su presencia comenzó a desatar importantes cambios ecológicos (Gompper, 2014a; Marra y Santella, 2016).
Impacto de los gatos y perros sobre la biodiversidad
Gatos y perros como mascotas
Debido a los usos que los de nuestra especie han dado a los gatos y perros como animales domésticos, el proceso de domesticación de estas especies implicó que se seleccionaran individuos que fueran buenos cazadores, mostrándose al mismo tiempo menos tímidos y agresivos con los humanos (Driscoll et al., 2007; Vigne, 2004). Sin embargo, a partir de siglo XIII en el caso de los perros y del siglo XIX con los gatos, se empezó una selección basada en características funcionales y estéticas (Frantz et al., 2016; Gompper, 2014b; O’Brien y Johnson, 2007). Este cambio en el proceso de domesticación permitió una nueva relación entre estos carnívoros y nuestra especie, cambiando su papel de ser únicamente animales de trabajo, a servir también como animales de compañía, sin que en este proceso se perdiera la capacidad para sobrevivir de forma independientes a los humanos en la mayoría de las razas (Gompper, 2014a; Hughes y Macdonald, 2013; Jensen, 2007). Este último proceso de domesticación generó una explosión en el tamaño de las poblaciones de gatos y perros a nivel mundial. Hoy en día, se estima que hay aproximadamente 600 millones de gatos (Dauphine y Cooper, 2009; Gompper, 2014a; O’Brien y Johnson, 2007) y entre 700 y 1,000 millones de perros en nuestro planeta (Gompper, 2014b; Hughes y Macdonald, 2013). Al evaluar los efectos ecológicos de gatos y perros es importante considerar que ambas especies pueden encontrarse tanto como mascotas domésticas, como animales callejeros, o ferales. Además, debemos tomar en cuenta que a los individuos con dueño suele permitírseles libre movimiento fuera de sus hogares (APPA, 2017; Dauphine y Cooper, 2009; Hughes y Macdonald, 2013; Pacheco, 2003). Tan solo en EUA, se ha calculado que hay cerca de 94 millones de gatos y 90 millones de perros con dueño (APPA, 2017; Hughes y Macdonald, 2013) y que a 65% de los gatos con dueño se les permite libre movimiento fuera de sus hogares (APPA, 2017; Duffy y Capece, 2012). En el caso de los perros con dueño, 75% de los individuos a nivel mundial tienen libre movimiento fuera de sus hogares (Hughes y Macdonald, 2013). Esto sucede sobre todo en países con poco o nulo control sobre el manejo de mascotas, como es el caso de México (APPA, 2017; Gompper, 2014b; Hughes y Macdonald, 2013).
Debido a la relación que gatos y perros tienen con los humanos, se han establecido tanto en hábitats urbanos, como en áreas rurales y zonas con baja o nula perturbación humana, por lo que se han convertido en especies exóticas e invasoras en muchas regiones de nuestro planeta (Doherty et al., 2015; Young et al., 2017). Hoy en día está claro que estas 2 especies tienen multiples efectos negativos sobre un gran número de especies nativas (Doherty, Dickman, Glen et al., 2017; Loss et al., 2013; Loyd et al., 2013; Young et al., 2011). La magnitud de sus efectos depende del estado de conservación en que se encuentran los sitios que invaden, de su manejo y del grado de la dependencia que los individuos de diferentes razas tengan con el ser humano (Gompper, 2014a, b; Marra y Santella, 2016; Spotte, 2014; Vanak y Gompper, 2010). Sin embargo, conocer sus efectos negativos puede ser difícil, tanto por la cercanía emocional que tenemos con ellos, como por el tiempo que ha pasado desde su llegada a las diferentes regiones del planeta donde están presentes (Driscoll et al., 2007, 2009; Gompper, 2014b). Esto limita nuestra capacidad para reconocer los cambios en el medio ambiente que han sido generados por estas mascotas, volviéndose crucial entender su ecología para poder valorar sus efectos sobre las especies nativas.
Es relevante notar que la forma en que los humanos manejamos a los gatos y perros domésticos afecta de forma notable su ecología y la forma en que interactúan con su hábitat y otras especies animales (Doherty, Dickman, Glen et al., 2017; Jensen, 2007; Loss et al., 2013; Vanak et al., 2009). De este modo, para su estudio, a los individuos de ambas especies se les ha clasificado de acuerdo con su forma de manejo y el tipo de asociación que tienen con los humanos (Brickner, 2003; Doherty, Dickman, Glen et al., 2017; Loss et al., 2013; Vanak et al., 2014; Weston et al., 2014; Wierzbowska et al., 2016). Las categorías de manejo son: a) gatos y perros con dueño sin libre acceso a la calle (total restricción espacial con control de sus movimientos por los propietarios); b) gatos y perros con dueño y de vida libre (de semi restricción y/o supervisión semi dependiente, a los que se les permite el uso de espacios exteriores a la casa de sus dueños); c) gatos y perros extraviados, abandonados y/o callejeros, a los que no se les provee de cuidados, no son agresivos con el humano y son alimentados ocasionalmente o cazan para alimentarse; y d) gatos y perros ferales o asilvestrados (sin dueño, sin restricción de movimiento y con total independencia alimentaria de los humanos). Estos últimos pueden ser muy agresivos con el humano, totalmente independientes de él y generalmente son descendientes de animales extraviados o abandonados, y están sujetos a la selección natural (Brickner, 2003; Vanak y Gompper, 2009; Vanak et al., 2014; Weston et al., 2014). Para los fines de este trabajo dividiremos a los gatos y perros en 2 grupos: individuos con dueño (categorías a y b) e individuos ferales (conjuntando las categorías c y d).
Gatos y perros domésticos como especies invasoras
Como especies exóticas invasoras en diferentes regiones de nuestro planeta, tanto los gatos como los perros domésticos tienen efectos nocivos sobre la fauna silvestre nativa tanto dentro de ecosistemas naturales, como en zonas rurales y urbanas (Doherty et al., 2016, Doherty, Dickman, Johnson et al., 2017; Gompper, 2014a; Loss et al., 2013). Podemos asumir que en Medio Oriente, Europa y en grandes zonas de Eurasia, los efectos de los individuos domésticos han sido menores por la presencia de sus parientes silvestres: el gato salvaje africano (Felis silvestris lybica), el gato silvestre europeo (Felis silvestris silvestris; Marra y Santella, 2016; Nowak, 2005; Vigne, 2004) y el lobo (Canis lupus; Frantz et al., 2016; Nowak, 2005). Como resultado de la presencia de estas especies silvestres, en estas áreas geográficas las presas estaban acostumbradas a este tipo de depredadores, lo que ha permitido que eviten con mayor eficiencia ser cazados por gatos y perros domésticos (Donfrancesco et al., 2019; Gompper, 2014a; Vanak y Gompper, 2009).
Actualmente, el gato y el perro son considerados como 2 de las especies invasoras de vertebrados con más efectos negativos a nivel global (Brickner, 2003; Gompper, 2014a; Loyd et al., 2013; Young et al., 2011). Según la “Base de Datos Global de Especies Invasoras” (GISD por sus siglas en inglés; GISD, 2017), el gato se encuentra entre las 100 peores especies invasoras del planeta, ocupando el lugar 38 y es el principal responsable planetario de la extinción de especies de aves (33 especies desde el siglo XVII; GISD, 2017; Medina et al., 2011; Winter, 2006). Se ha encontrado que los gatos cazan más de 1,000 especies de animales en todo el mundo (Lepczyk et al., 2015). Adicionalmente, los gatos son la principal amenaza potencial para 8% de las especies de aves, mamíferos y reptiles en peligro de extinción (Medina et al., 2011). Específicamente para el caso de aves insulares, después de los roedores exóticos, los gatos son los segundos depredadores en importancia, atribuyéndoles casi 26% de las extinciones de especies dentro de este grupo (Bonnaud et al., 2011; Doherty et al., 2016; Loss et al., 2013; Yip et al., 2014).
Aunque el perro doméstico no ha sido incluido dentro de la lista de las 100 peores especies invasoras de la GISD, hay evidencia contundente de sus impactos negativos sobre la fauna silvestre nativa (Doherty et al., 2016; Doherty, Dickman, Glen et al., 2017; Hughes y Macdonald, 2013; Young et al., 2011). Se ha encontrado que los perros tienen un gran impacto como depredadores, con 79% de las publicaciones sobre la ecología de perros domésticos indicando su papel como importantes cazadores de fauna silvestre (Doherty, Dickman, Glen et al., 2017; Hughes y Macdonald, 2013; Spotte, 2014). Con base en esta información, los perros son considerados como una amenaza potencial para 188 especies amenazadas de vertebrados en todo el mundo, las cuales incluyen a 96 especies de mamíferos (de 33 familias), 78 especies de aves (de 25 familias), 22 especies de reptiles (de 10 familias) y 3 especies de anfibios (Doherty, Dickman, Glen et al., 2017). De estas especies amenazadas, 87 están catalogadas como vulnerables, 71 en peligro de extinción y 30 en peligro crítico (Doherty, Dickman, Glen et al., 2017). Además, los perros domésticos han contribuido a la extinción de 11 especies de vertebrados (ver Doherty, Dickman, Glen et al., 2017 y trabajos citados en él).
Efectos ecológicos de los gatos y los perros domésticos
Entre los principales efectos ecológicos negativos generados por los gatos y perros domésticos, encontramos la depredación de diversas especies de fauna silvestre nativa, la competencia interespecífica o de interferencia con otras especies de carnívoros y la hibridación con otras especies de felinos y cánidos silvestres (Biró et al., 2005; Gompper, 2014a; Oliveira et al., 2008; Vanak y Gompper, 2009). Adicionalmente, pueden modificar la conducta de otras especies a las cuales no depredan y con las que no compiten (e.g., los perros persiguen y asustan a todo tipo de fauna) y pueden transmitir enfermedades y parásitos a la fauna silvestre, e incluso al ser humano (Acosta-Jamett, 2009; Acosta-Jamett et al., 2011; Doherty, Dickman, Glen et al., 2017; Gompper, 2014a; Lenth et al., 2008; Vanak y Gompper, 2009; Weston et al., 2014).
Además de los efectos como depredadores que principalmente tienen los individuos ferales de ambas especies, en años recientes se ha podido medir el impacto de depredación de los gatos y perros con dueño a los que se les permite libre movimiento fuera de sus hogares (Loss et al., 2013; Loyd et al., 2013; Wierzbowska et al., 2016; Woods et al., 2003). Estos animales, a pesar de ser alimentados por sus dueños, siguen teniendo efectos negativos sobre las comunidades de fauna silvestre. Por ejemplo, un solo individuo de gato con dueño lleva a su hogar en promedio 13.4 presas por año (van Heezik et al., 2010). Este número subestima sus efectos como depredadores, ya que los gatos domésticos con dueño abandonan o consumen en el sitio de caza 49% de sus presas (Loyd et al., 2013). La diversidad de especies nativas que son depredadas por estas mascotas varía entre 6 y 93 especies de vertebrados e invertebrados por gato por año (Barratt, 1998; Loyd et al., 2013; Orduña-Villaseñor, 2015; Woods et al., 2003). Se ha estimado que los gatos con dueño son responsables de hasta 16% del total de la depredación causada por esta especie en EUA (Loss et al., 2013). Los gatos de libre movimiento con dueño suelen tener un mayor impacto sobre reptiles, mamíferos y aves en términos del número de individuos depredados (Loss y Marra, 2017; Loyd et al., 2013; Orduña-Villaseñor, 2015), pero considerando el número de especies que son presas, el grupo más afectados son las aves, seguidas de los mamíferos (Barratt, 1997, 1998; Loyd et al., 2013; Murphy et al., 2019; Woods et al., 2003). Del papel de los perros domésticos con dueño como depredadores sabemos muy poco. La mayoría de los trabajos sobre su impacto se han enfocado a evaluar su depredación sobre especies particulares de animales (e.g., venado bura: Odocoileus hemionus; venado cola blanca: Odocoileus virginianus; Lowry y McArthur, 1978) o sobre grupos taxonómicos particulares (ungulados, lagomorfos; Silva-Rodríguez y Sieving, 2011, 2012; Wierzbowska et al., 2016), aunque hay información que indica que pueden ser importantes depredadores de aves (obs. pers.; Doherty, Dickman, Glen et al., 2017; Gompper, 2014a).
Por otro lado, un efecto ecológico que ha sido poco estudiado es el papel que estas 2 especies tienen como reservorios y transmisores de enfermedades. Esto se debe a que los perros y gatos domésticos interactúan tanto con el ser humano como con diferentes especies de fauna silvestre, fungiendo como importantes vectores de enfermedades. (material suplementario, apéndice 1). Entre estas encontramos la enfermedad de la fiebre por arañazo del gato (causada por la bacteria Bartonella henselae; Lepczyk et al., 2015), la rabia (causada por un virus del género Lyssavirus; Dyer et al., 2014; Otranto et al., 2015), la salmonelosis (Salmonella sp.; Baneth et al., 2016; Lepczyk et al., 2015), la toxoplasmosis (Toxoplasma gondii; Baneth et al., 2016; Lepczyk et al., 2015; Watts y Benson, 2016), el virus de la inmunodeficiencia felina (VIF; Baneth et al., 2016; Biezus et al., 2019; Lepczyk et al., 2015), el parvovirus (VPC; Acosta-Jamett, Cunningham et al., 2015; Acosta-Jamett, Surot et al., 2015; Blancher, 2013; Otranto et al., 2015) y el moquillo canino (VDC; Acosta-Jamett et al., 2011; Acosta-Jamett, Cunningham et al., 2015; Otranto et al., 2015).
En relación con la transmisión de enfermedades a la fauna silvestre, se ha documentado la transmisión de T. gondii del gato doméstico a individuos de gato montés (Lynx rufus; Alvarado-Esquivel et al., 2013) y jaguarundi (Herpailiurus jagouroundi; Bevins et al., 2012). Otro ejemplo es la transmisión del virus del moquillo canino entre perros domésticos y el lobo etíope (Canis simensis; Daszak, 2000; material suplementario, apéndice 1). Adicionalmente, hay reportes de gatos y perros que han contraído enfermedades y las han trasmitido a los humanos, como es el caso de la rabia, la toxocariasis (infección parasitaria por larvas de Toxocara catis y T. canis) y la toxoplasmosis (Baneth et al., 2016; Lepczyk et al., 2015). Por ejemplo, en 99% de los casos de rabia en humanos, el virus es transmitido por perros domésticos (OMS, 2019). Adicionalmente, los humanos pueden transmitir enfermedades tanto a gatos como a perros, como ha ocurrido con la enfermedad COVID 19 (Alberto-Orlando et al., 2022; Neira et al., 2021). Dada la alta densidad global de estas 2 especies y su continua invasión de ecosistemas naturales, el riesgo de que los gatos y perros funcionen como reservorios para la transmisión de nuevas enfermedades entre la fauna silvestre y los humanos es alto (Banks y Bryant, 2007; Gingold et al., 2009; Paniz-Mondolfi et al., 2019; Parsons et al., 2016; Ribeiro et al., 2019; Silva-Rodríguez y Sieving, 2011, 2012; Soto y Palomares, 2015; Weston et al., 2014; material suplementario, apéndice 1).
Cuando el gato y el perro doméstico invaden hábitats donde hay especies nativas de felinos y cánidos, pueden competir con ellas por presas y otros recursos (Gehrt et al., 2013; Hernandez et al., 2018; Lenth et al., 2008; Vanak et al., 2014; Vanak y Gompper, 2009). El efecto de esta competencia tiende a magnificarse debido a las altas densidades poblacionales que estos 2 carnívoros pueden alcanzar y a que ambas especies suelen verse beneficiadas por las actividades humanas (APPA, 2017; Gompper, 2014b; Hughes y Macdonald, 2013). Además, se ha detectado la hibridación de gatos y perros con especies nativas. Aunque en condiciones silvestres es poco común, tanto el gato como el perro doméstico, son capaces de generar híbridos con especies nativas de felinos y cánidos, y también pueden cruzarse con individuos de las poblaciones silvestres de sus especies originarias (Felis silvestris y Canis lupus; Anderson et al., 2009; Donfrancesco et al., 2019; Li et al., 2016; Nussberger et al., 2013; VonHoldt et al., 2013). Esto último, amenaza la integridad genética de las poblaciones silvestres de gatos silvestres y lobos, muchas de las cuales están fuertemente amenazadas (Adams et al., 2007; Biró et al., 2005; Donfrancesco et al., 2019; Oliveira et al., 2008; Verardi et al., 2006). Por ejemplo, se ha documentado que el gato doméstico se ha entrecruzado con poblaciones de gato silvestre (Felis s. silvestris y Felis s. libyca) en Hungría, Escocia, Italia, Alemania y en la península Ibérica (Adams et al., 2007; Le Roux et al., 2015; Li et al., 2016; O’Brien et al., 2009; Oliveira et al., 2008). E incluso se han detectado híbridos del gato doméstico y el gato de la selva o gato rojo (Felis chaus) en Egipto (Li et al., 2016). En el caso del perro doméstico, por su interacción y cercanía filogenética con otras especies de cánidos puede llegar a hibridarse, habiendo reportes de híbridos de perro doméstico con lobos (Anderson et al., 2009; Pilot et al., 2021; Popova y Zlatanova, 2020; VonHoldt et al., 2013), lobos etíopes (Gottelli et al., 1994, 2013; Leonard et al., 2014), coyotes (Canis latrans; Leonard et al., 2014; Weeks et al., 1990) y chacales dorados (Canis aureus; Galov et al., 2015; Leonard et al., 2014).
Finalmente, aunque contamos con poca información sobre cómo los gatos domésticos cambian la conducta de otros animales, este tema ha sido estudiado más ampliamente en perros (Banks y Bryant, 2007; Gingold et al., 2009; Parsons et al., 2016; Ribeiro et al., 2019; Silva-Rodríguez y Sieving, 2011, 2012; Soto y Palomares, 2015; Weston et al., 2014). En el caso de los gatos sabemos que sus marcas de orina pueden modificar la conducta de sus presas o de otros carnívoros medianos, quienes evitan estar activos en zonas marcadas (Ancillotto et al., 2019; Lepczyk et al., 2015). En el caso de los perros, el efecto sobre otras especies está asociado tanto a sus marcas olfativas, como a sus ladridos y a persecuciones (Banks y Bryant, 2007; Silva-Rodríguez y Sieving, 2012). Estas actividades pueden generar la disminución de fauna silvestre en zonas por las cuales hayan transitado perros, tanto solos, como acompañando a humanos que realizaban actividades recreativas (Parsons et al., 2016; Silva-Rodríguez y Sieving, 2011, 2012; Soto y Palomares, 2015; Weston et al., 2014). Vale la pena destacar la necesidad de conducir más trabajos sobre la ecología del miedo asociada a estas 2 especies.
Efectos de los gatos y perros domésticos a nivel mundial, con énfasis en países megadiversos
Los efectos ecológicos negativos de gatos y perros domésticos deben de ser mayores en países megadiversos que en países de baja diversidad de vertebrados por varias razones. Los factores que pueden agravar el daño que causan estas 2 especies dentro de los países megadiversos son tanto de tipo socioeconómico, como político y ecológicos (Dirzo et al., 2014; Doherty et al., 2016; Gompper, 2014b; Taeihagh, 2017). Hay que considerar que la mayoría de los países megadiversos, son naciones en vías de desarrollo con importantes problemas socioeconómicos, vacíos legales y de cumplimiento de leyes para el manejo adecuado de mascotas y especies invasoras, altas tasas de deforestación y en muchos casos, un rápido crecimiento de su frontera agrícola/ganadera (Dirzo et al., 2014; Doherty et al., 2015, 2016; Gompper, 2014a; Taeihagh, 2017).
La mayor parte de los estudios sobre la ecología de gatos y perros domésticos se han llevado a cabo en países templados de América del Norte y Europa. En general estos países tienen una baja diversidad de vertebrados (Doherty, Dickman, Glen et al., 2017; Llorente-bousquets y Ocegueda, 2008; Mittermeier et al., 1997; Pimm et al., 2014). Esto es importante de enfatizar, debido a que el comportamiento y los efectos de estas 2 especies invasoras pueden ser distintos en países megadiversos. Debido a que sólo nos interesa discutir el efecto de los gatos y perros sobre la fauna nativa (Doherty, Dickman, Glen et al., 2017; Pimm et al., 2014), para este trabajo definimos países megadiversos como el grupo de naciones poseedoras de la mayor cantidad de diversidad de vertebrados a nivel mundial (modificado de: Conabio, 2016; Llorente-bousquets y Ocegueda, 2008; Marchese, 2015; Mittermeier et al., 1997). De este modo, no incluimos en esta categoría a algunos países considerados como megadiversos por su alta diversidad de plantas, tales como China y EUA. Los países megadiversos considerados en esta revisión son 15. En África se encuentran: Madagascar, República Democrática del Congo y Sudáfrica; en América: Brasil, Colombia, Ecuador, México, Perú y Venezuela; en Asia: Filipinas, India, Indonesia y Malasia; y en Oceanía: Australia y Papúa Nueva Guinea (Conabio, 2016; Llorente-Bousquets y Ocegueda, 2008; Pierpaoli et al., 2003). La mayor parte de estos países megadiversos y los más importantes en términos del número de especies de vertebrados, se encuentran en latitudes tropicales (Conabio, 2016; Mittermeier et al., 1997), donde las invasiones de gatos y perros han sido más recientes, o donde ha habido presencia de perros manteniendo densidades poblacionales relativamente bajas (Driscoll et al., 2007, 2009; Driscoll y Macdonald, 2010; Frantz et al., 2016).
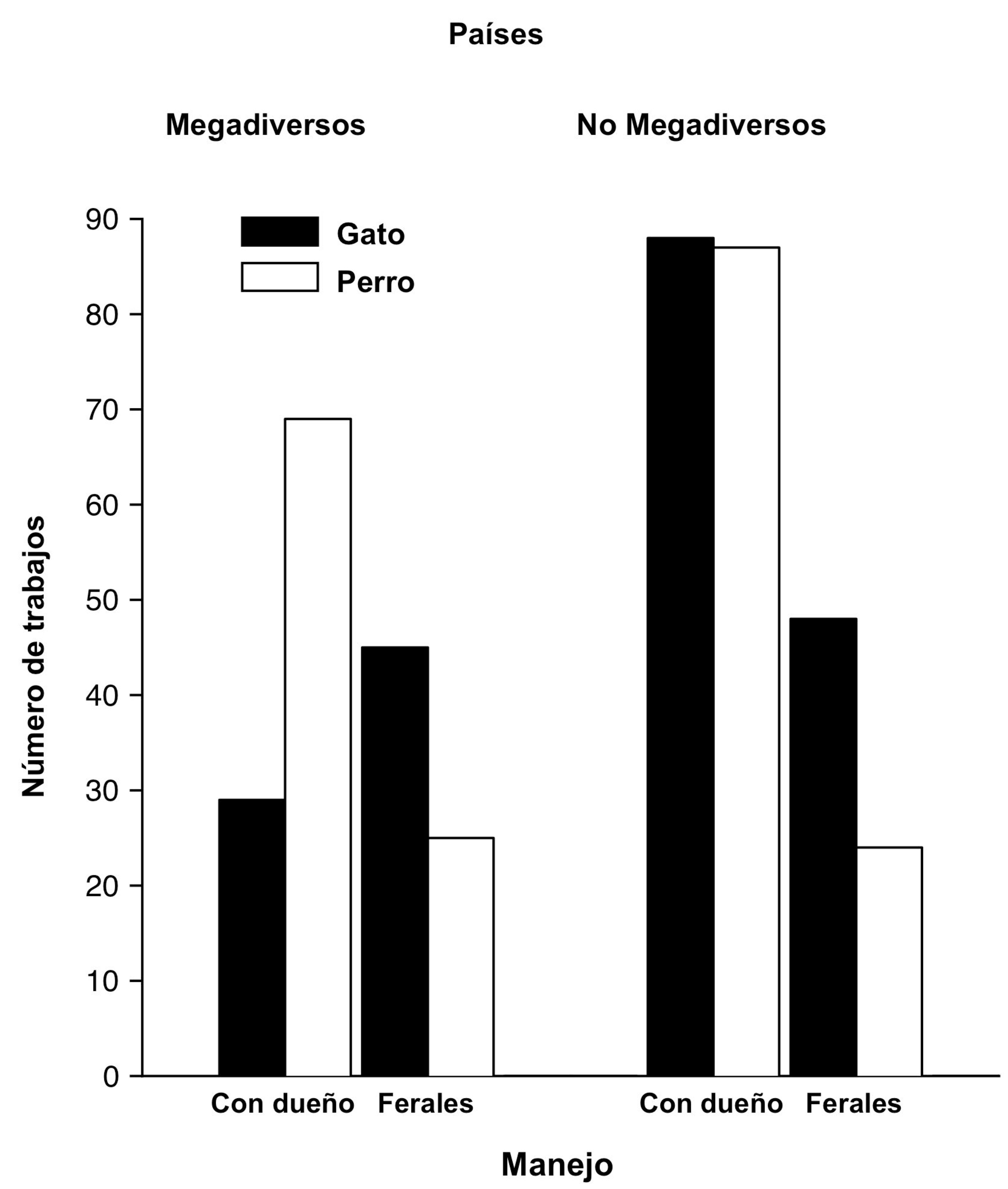
Se llevó a cabo una búsqueda en la literatura (Web of Knowledge, Science Direct, Google Scholar y Mendeley) con los términos: domestic cat, domestic dog, feral cat, feral dog, Felis catus, Canis lupus familiaris, free ranging cat y free ranging dog. Esta búsqueda nos permitió localizar un total de 199 estudios relacionados con la ecología de gatos y 196 trabajos sobre la ecología de los perros (fig. 1, material suplementario, apéndices 2, 3). Para los trabajos con gatos, 134 fueron realizados en países no megadiversos y los 65 restantes en países megadiversos. Para los países no megadiversos representados en la muestra (más de 20 países), la mayoría de los trabajos fueron llevados a cabo con gatos con dueños (88 trabajos; fig. 1), habiendo menos trabajos con gatos ferales (48; material suplementario, apéndice 2). Mientras que en los países megadiversos (6 países; Australia, Brasil, Filipinas, México, Sudáfrica y Venezuela), tenemos mejor conocimiento de lo que sucede con los gatos ferales (45 trabajos) que con los gatos con dueños (29 trabajos; material suplementario, apéndice 2). Para los perros domésticos, 103 publicaciones fueron generadas en países no megadiversos (más de 20 países; 87 trabajos con perros con dueño y 24 con perros ferales; material suplementario, apéndice 3). Los estudios llevados a cabo en países megadiversos con esta especie fueron 89, de los cuales 69 fueron para perros con dueño (realizados en Australia, Brasil, Colombia, Ecuador, India, México, Perú, Venezuela y Sudáfrica) y 25 con perros ferales (llevados a cabo en Australia, Brasil, Ecuador, India, Sudáfrica y México; material suplementario, apéndice 3).
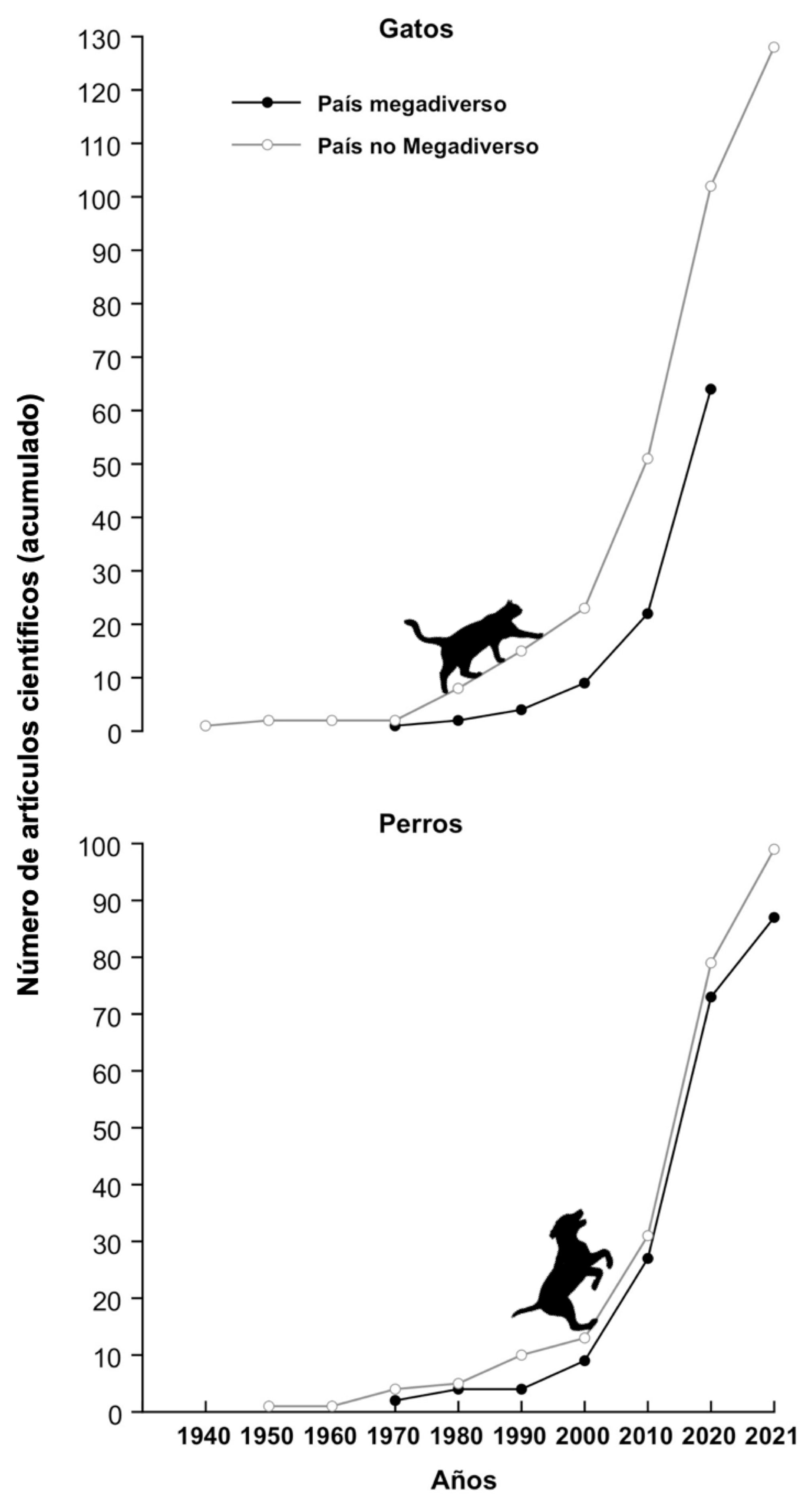
En países no megadiversos los temas más estudiados con gatos y perros domésticos han sido la depredación de fauna nativa, seguido por la competencia con otras especies de carnívoros y su impacto como reservorios de parásitos y enfermedades (material suplementario: apéndices 2, 3). En estos países existen pocos estudios sobre cómo cambian la conducta de otras especies de fauna, o sobre su hibridación con otras especies (material suplementario, apéndice 3) (Doherty et al., 2016; Doherty, Dickman, Glen et al., 2017; Loss et al., 2013). Para el caso particular de los gatos se han hecho más estudios sobre su impacto como depredadores, siendo la mayoría de estos sobre gatos con dueño. Mientras que en los países megadiversos se han estudiado más los efectos de depredación causados por individuos ferales (material suplementario, apéndice 2). Para el caso de los perros en países no megadiversos se ha estudiado más el efecto que tienen como reservorios de enfermedades, seguido por su papel como depredadores y sus efectos a través de la competencia (figs. 2, 4; material suplementario, apéndice 3) (Doherty, Dickman, Glen et al., 2017; Hughes y Macdonald, 2013). Mientras que en países megadiversos los trabajos con esta especie se han enfocado principalmente a entender su papel como depredadores, aunque hay un creciente interés en su papel como reservorios de enfermedades (figs. 2, 4) (Acosta-Jamett et al., 2011; Hughes y Macdonald, 2013; Loyd et al., 2013; Silva-Rodríguez et al., 2010).
¿Qué sabemos sobre los efectos ecológicos de gatos y perros en América?
En el continente americano la mayoría de los estudios tanto de gatos como de perros se han desarrollado sobre todo en 3 países: EUA (38 de gatos y 29 de perros), México (12 y 22 estudios de gatos y perros, respectivamente) y Brasil (10 y 19 estudios; tabla 1, fig. 3). Los gatos han sido relativamente poco estudiados en el continente (tabla 1, fig. 3). En estos estudios, los gatos domésticos con dueño han recibido más atención que los gatos ferales (tabla 1). A pesar de que el número de estudios sobre gatos domésticos se ha incrementado de forma importante en la última década, éstos siguen siendo muy pocos a nivel continental y no nos permiten entender el verdadero impacto que los gatos domésticos, tanto ferales como con dueño, han tenido en las masas continentales del planeta. Las zonas geográficas del continente que presentan pocos estudios o que carecen de información son las áreas tropicales que tienen la mayor biodiversidad, e incluyen la parte continental de México, Brasil, y a países como Colombia, Ecuador, Perú y Venezuela, para los cuales existe poca información publicada (Aldaz et al., 2015; Duque-García et al., 2017; Oliver et al., 2015; Raynor et al., 2020; Zapata-Ríos y Branch, 2018).
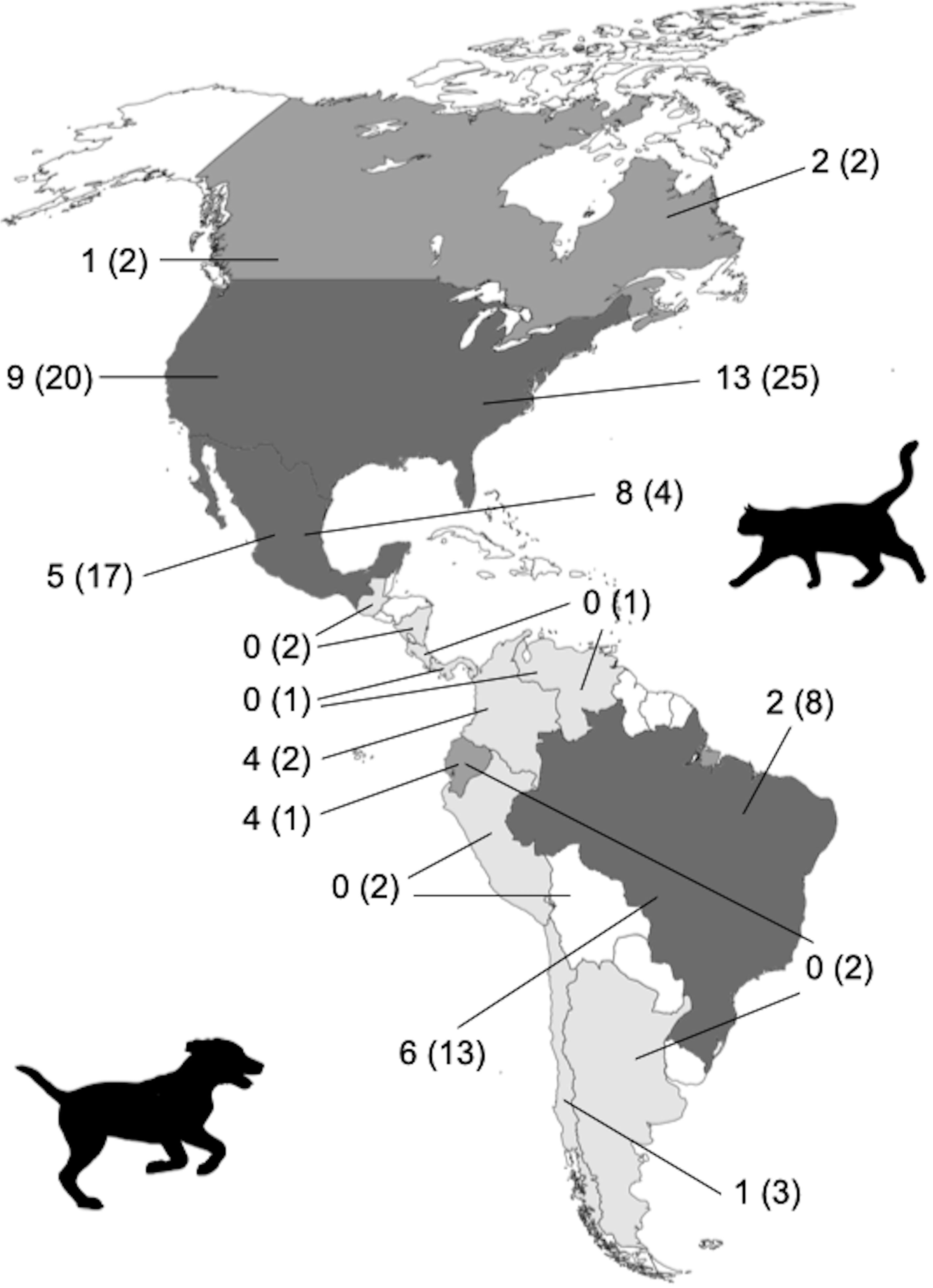
A nivel continental los perros han sido más estudiados que los gatos (112 trabajos). Estos trabajos representan estudios llevados a cabo en catorce países de América (tabla 1). Habiendo un mayor número de estudios en EUA, México, Brasil, Chile, Colombia y Ecuador (tabla 1, fig. 3). A diferencia de los gatos, los perros han sido más estudiados en las zonas tropicales del continente, aunque no hay estudios para varios países (tabla 1, fig. 3). La mayoría de los trabajos con perros se han enfocado en individuos domésticos con dueño, habiendo un menor número de trabajos con perros ferales (80 y 32 respectivamente; tabla 1, fig. 3). Al igual que con los gatos, el tema menos estudiado en los perros es el de hibridación con fauna silvestre. Aun así, sabemos que su hibridación con otros cánidos amenaza la integridad genética de especies como el lobo gris (Canis lupus; Valadez et al., 2002; VonHoldt et al., 2013) y el coyote (Canis latrans) en la parte norte del continente (Bee y Hall, 1951; Weeks et al., 1990). Siendo este un tema crucial que debe de ser abordado en América del Sur, donde existe una alta diversidad de cánidos nativos.
Los efectos de gatos y perros domésticos en México
En México, los gatos domésticos se encuentran asociados a los asentamientos humanos. Por lo tanto, su distribución en el país es un reflejo de la distribución de las zonas urbanas. Álvarez-Romero y colaboradores (2008), utilizando información sobre el ámbito hogareño y movimiento diario de individuos ferales y callejeros tomado de la literatura, propusieron que el área de impacto de los gatos de las zonas urbanas se puede extender por un radio aproximado de 5 km alrededor de cada núcleo poblacional (Álvarez-Romero et al., 2008; Calhoon y Haspel, 1989; Nowak, 2005). De este modo, dada la distribución de las zonas urbanas en México, los gatos se encuentran prácticamente en todos los tipos de vegetación del país. Adicionalmente su presencia es particularmente importante en diversas islas, con la mayor parte de los estudios sobre sus efectos como depredadores habiéndose llevado a cabo con individuos ferales en islas oceánicas de nuestro país (Aguirre-Muñoz et al., 2008, 2011; Donlan et al., 2000; Luna-Mendoza et al., 2011; March y Martínez 2007; Ortiz-Alcaraz et al., 2017).
Los resultados de estos trabajos indican una disminución en el tamaño poblacional y la extinción de especies de tamaño pequeño y mediano de reptiles, aves y mamíferos. En relación con los reptiles, los gatos han reducido la población de una especie de lagartija endémica a la Isla Socorro del archipiélago de Revillagigedo (Urosaurus auriculatus), representando esta especie una parte importante de la dieta de los gatos ferales que ahí habitan (Ortiz-Alcaraz et al., 2017). En el caso de las aves, Wood y colaboradores (2002), reportan que los gatos están directamente relacionados con la extirpación de al menos 14 poblaciones de diferentes especies y subespecies de aves marinas tan sólo en islas del noroeste de México. Además, los gatos son considerados como el factor más importante de riesgo para las poblaciones de aves acuáticas de las islas Ángel de la Guarda, San Marcos, Carmen, Santa Catalina y Cerralvo (Aguirre-Muñoz et al., 2008), todas localizadas en el golfo de California. Es crucial mencionar que los gatos han causado una drástica disminución en las poblaciones y la desaparición de la pardela mexicana (Puffinus opisthomelas), la alcita de Cassin (Ptychoramphus aleuticus) y el mérgulo de Xantus (Endomychura hypoleuca) (Keitt et al., 2002; McChesney y Tershy, 1998). Finalmente, la extinción total del petrel de Guadalupe (Oceanodroma macrodactyla), de una subespecie endémica de gorrión (Aimophila ruficeps sanctorum) de la isla Todos Santos (Howell y Webb, 1995; McChesney y Tershy, 1998; Mellink, 1993) y la desaparición en vida silvestre de la paloma de Socorro (Zenaida graysoni; Howell y Webb, 1995; Keitt y Tershy, 2003), también están asociadas a la presencia de gatos.
Tabla 1
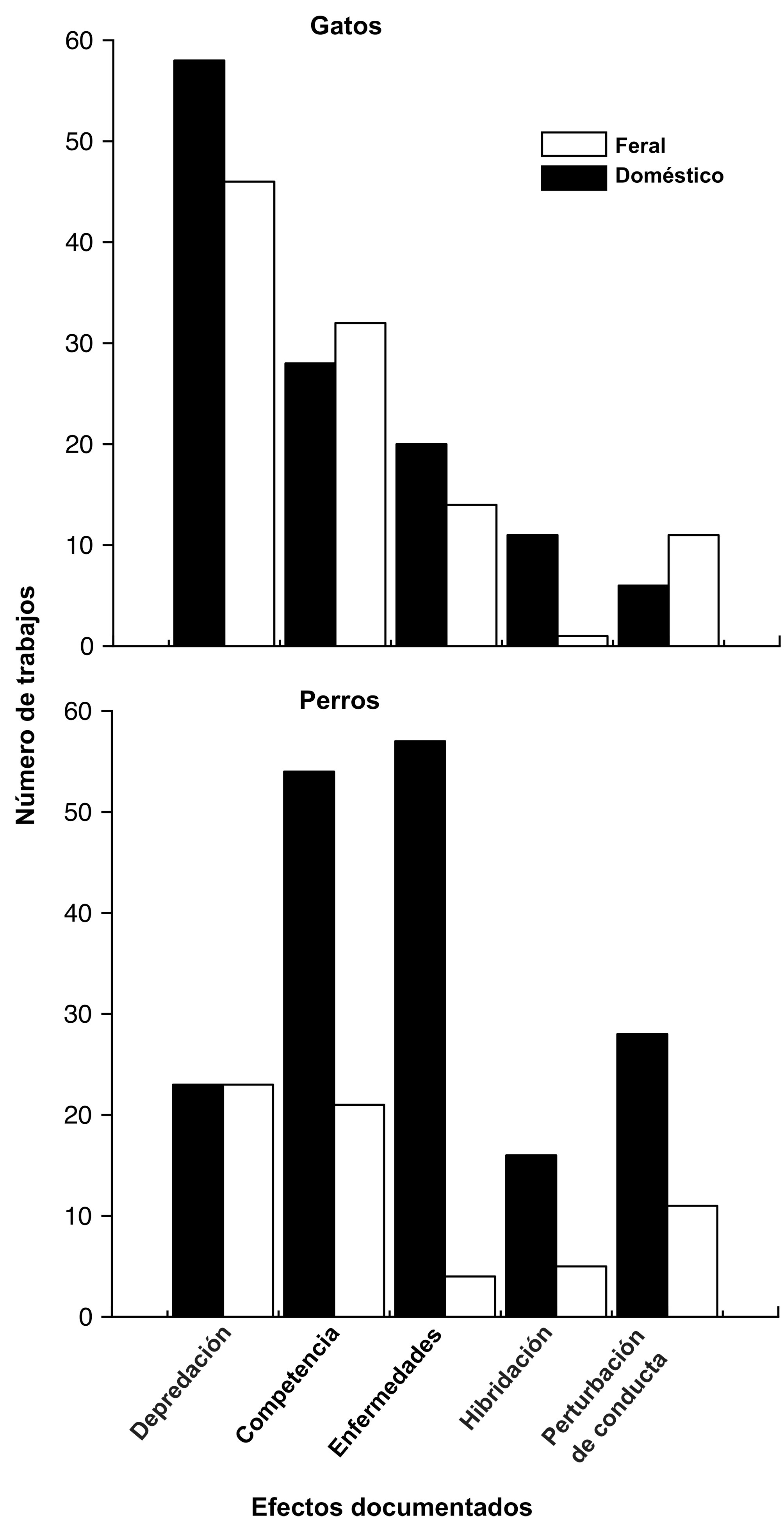
Número de estudios de gatos y perros en el continente americano ordenados según su efecto ecológico. Los trabajos fueron publicados en el periodo comprendido entre 1944 y 2021 en el caso de los gatos, y entre 1951 y 2021 en el caso de los perros (ver metodología para detalles del protocolo de búsqueda de información).
| Tipo | Efecto ecológico | Con dueño | Ferales |
| Gato | Depredación | 22 | 13 |
| Competencia | 13 | 6 | |
| Enfermedades | 12 | 6 | |
| Hibridación | 0 | 0 | |
| Perturbación del comportamiento de otras especies | 3 | 5 | |
| Total: 80 | 50 | 30 | |
| Perro | Depredación | 15 | 11 |
| Competencia | 25 | 15 | |
| Enfermedades | 33 | 5 | |
| Hibridación | 5 | 0 | |
| Perturbación del comportamiento de otras especies | 19 | 8 | |
| Total: 136 | 97 | 39 |
En el caso de mamíferos insulares mexicanos, el gato ha sido considerado un riesgo negativo para las poblaciones de especies endémicas de roedores, tales como Chaetodipus anthonyi, Peromyscus interparietalis y Neotoma bryanti; y es responsable de la extinción de 2 especies endémicas de rata (N. anthonyi y N. martinensis; Mellink, 1992; Rodríguez-Moreno et al., 2007; Vázquez-Domínguez et al., 2004). Además, los gatos son la causa más probable de la extinción del ratón Peromyscus guardia (Mellink et al., 2002; Vázquez-Domínguez et al., 2004) y de la disminución de las poblaciones del conejo en la Isla Cedros (Sylvilagus bachmani cerrosensis; Álvarez-Romero et al., 2008).
La presencia de gatos ferales en islas oceánicas es muy importante por las condiciones ambientales de las mismas y el alto endemismo de especies no voladoras generado por los procesos de evolución por aislamiento que se dan en ellas (Álvarez-Romero et al., 2008; Keitt y Tershy, 2003; March y Martínez, 2007; Tershy et al., 2002). Debido a esto, en nuestro país ha habido un gran esfuerzo para erradicar a los gatos en islas oceánicas (Aguirre-Muñoz et al., 2008; Luna-Mendoza et al., 2011; March y Martínez, 2007; Ortiz-Alcaraz et al., 2017). En este sentido, se ha logrado la erradicación de gatos ferales en las islas Asunción, Carmen, Catalina, Coronados Norte, Coronados, Estanque, Isabel, Mejía, Monserrat, Natividad, Partida Sur, Rasa, San Francisco, San Jerónimo, San Marín, San Roque y las islas de Todos Santos Norte y Sur (Aguirre-Muñoz y Mendoza, 2009; Aguirre-Muñoz et al., 2008; Arnaud et al., 1993; Donlan et al., 2000; Keitt et al., 2002). Sin embargo, aún hay poblaciones ferales de gatos en algunas islas del océano Pacífico (Cedros y Guadalupe) y del golfo de California (Ángel de la Guarda, Cerralvo y Espíritu Santo; Aguirre-Muñoz y Mendoza, 2009; Arnaud et al., 1993; Keitt et al., 2002, 2005; Medina et al., 2011). Aunque se sabe poco acerca de la presencia de gatos en islas e islotes del golfo de México, es muy probable que haya individuos de gatos ferales en islas donde hay actividades humanas frecuentes. Adicionalmente se les puede encontrar en islas del caribe mexicano como Cozumel e Isla Mujeres (Aguirre-Muñoz et al., 2011; Álvarez-Romero et al., 2008).
Aunque los efectos del gato doméstico sobre la fauna insular en nuestro país están relativamente bien documentados, sabemos muy poco de la ecología y los efectos tanto de los gatos ferales cómo de individuos con dueño en el México continental. Sobre todo, hay poca información de lo que ocurre en las zonas urbanas y los hábitats naturales y agrícolas que los rodean, a pesar de la alta densidad que los gatos pueden alcanzar en estos sitios (Hernández-Cortazar et al., 2015; Orduña-Villaseñor, 2015; Ortega-Pacheco et al., 2007; Pacheco, 2003). Por la información existente para otros países podemos suponer que los gatos deben tener importantes efectos sobre diversas especies de reptiles, anfibios, aves y mamíferos (Baker et al., 2008; Barratt, 1997; Legge et al., 2017; Loyd et al., 2013). El impacto por depredación de los gatos en el México continental solo se ha estudiado con gatos domésticos con dueño de vida libre en la ciudad de Morelia, Michoacán (Orduña-Villaseñor, 2015) y Xalapa, Veracruz (Mella-Méndez, 2019). En estos estudios se reporta el consumo de 40 especies de vertebrados para Morelia y de 47 especies de vertebrados para Xalapa. El gato doméstico debe también considerarse un competidor potencial con otros carnívoros nativos del país, sin embargo, no existe información sobre este tema. Finalmente, los gatos interactúan con individuos de fauna silvestre lo que los vuelve potenciales reservorios de enfermedades. Por ejemplo, en Durango se estudió a gatos con dueño y callejeros, detectando toxoplasmosis, enfermedad que se ha encontrado también en otras especies silvestres de mamíferos presentes en México como el tlacuache norteño (Didelphis virginiana), el ardillón mexicano (Otospermophilus variegatus), el cacomixtle norteño (Bassariscus astutus), el zorrillo manchado occidental (Spilogale gracilis) y la comadreja cola larga (Mustela frenata; Alvarado-Esquivel et al., 2014; Dubey et al., 2007; García-Aguilar y Gallo-Reynoso, 2012; Hernández-Cortazar et al., 2015).
De forma similar a los gatos, los perros domésticos, sobre todo los individuos ferales, son importantes depredadores de fauna silvestre e incluso llegan a atacar ganado y personas (Cuarón et al., 2009; Mellink, 1993; Weston et al., 2014). En nuestro país han sido considerados como un importante factor de riesgo para las poblaciones de fauna en islas oceánicas (Aguirre-Muñoz et al., 2008; Donlan et al., 2000; Ortiz-Alcaraz et al., 2017; Wood et al., 2002). De este modo, se ha documentado su papel negativo sobre las poblaciones de aves marinas y pinnípedos en varias islas (García-Aguilar y Gallo-Reynoso, 2012; Ortiz-Alcaraz et al., 2017; Rodríguez-Estrella et al., 1991) y se le asocia con las reducciones poblacionales del conejo (Sylvilagus bachmani cerrosensis) y del venado bura (Odocoileus hemionus cerrosensis) de Isla Cedros (Baja California), siendo ambas subespecies endémicas en peligro de extinción (García-Aguilar y Gallo-Reynoso, 2012; Mellink, 1993). En esa misma isla también se ha reportado depredación de lobos y elefantes marinos del norte (Zalophus californianus y Mirounga angustirostris) por perros ferales, representando estas especies hasta 85% de la dieta de los perros (García-Aguilar y Gallo-Reynoso, 2012), lo que pone en riesgo sus poblaciones locales (García-Aguilar y Gallo-Reynoso, 2012). Adicionalmente se ha documentado el impacto de perros en otras islas, con manadas de perros ferales mostrando una alta agresividad a los humanos y a la fauna silvestre en Isla Guadalupe (Keitt et al., 2005; Mellink, 1993). En Cozumel se ha documentado que depredan al mapache pigmeo (Procyon pygmaeus) especie de carnívoro endémico de esta isla mexicana y considerada en peligro crítico de extinción. También se ha documentado que esta especie endémica ha estado en contacto con agentes infecciosos causantes de rabia, moquillo canino y toxoplasmosis, transmitidos por perros y gatos domésticos y ferales, que potencialmente pueden causar epizootias en sus poblaciones con efectos negativos para su conservación (Cuarón et al., 2016; Weber, 2010).
La presencia de perros ferales también es motivo de preocupación en algunas regiones y áreas naturales del México continental. Por ejemplo, se han registrado perros ferales en por lo menos 31 áreas naturales protegidas del país (Tershy et al., 2002). Sin embargo, los efectos de estas poblaciones sobre la fauna silvestre han sido poco estudiados. Adicionalmente a los daños que causan a la fauna silvestre, uno de los efectos más perjudiciales de esta especie para el ser humano ha sido la depredación de ganado. Estos ataques se dan tanto por perros ferales, como por perros de campesinos en algunas zonas rurales del país, siendo sus principales presas las aves de corral, chivos, borregos, e incluso el ganado vacuno (Álvarez-Romero et al., 2008; Cuarón et al., 2016; Romero-López et al., 2008).
Otro de los efectos negativos que pueden tener los perros domésticos sobre la fauna silvestre en nuestro país es la competencia con depredadores nativos (Mauti et al., 2017; Vanak et al., 2014; Vanak y Gompper, 2009, 2010). Debido a que este tema no ha sido estudiado en nuestro país, presentamos aquí algunas ideas que consideramos deben ser exploradas en futuros proyectos de investigación. Los perros domésticos, por su tamaño, sociabilidad, plasticidad en su dieta y asociación con el humano tienen la capacidad para desplazar a especies como coyotes, zorras, gato montés y un número importante de especies de carnívoros medianos y pequeños nativos a nuestro país (Gompper, 2014a; Vanak y Gompper, 2009). En países donde no hay poblaciones de perros callejeros se pueden encontrar poblaciones importantes de carnívoros silvestres como coyotes, zorro rojo (Vulpes vulpes), mapaches (Procyon lotor) e incluso osos negros (Ursus americanus), en y alrededor de zonas urbanas (Crooks y Soulé, 1999; Lenth et al., 2008; Wierzbowska et al., 2016). La ausencia de éstos y otras especies de carnívoros en ciudades de México puede estar relacionada con las altas densidades de perros callejeros y perros con dueño a los que se les permite libre movimiento fuera de sus hogares. Adicionalmente, las poblaciones de perros ferales en diferentes partes del país podrían ser un obstáculo importante para los proyectos de conservación y reintroducción de especies de carnívoros como el lobo mexicano (Beschta y Ripple, 2016; Newsome et al., 2015; Ripple et al., 2015).
Los perros son excelentes reservorios de parásitos y enfermedades (Alvarado-Esquivel et al., 2014; Cortés-Calva et al., 2013; Dubey et al., 2007; Fishbein et al., 1992; Suzán y Ceballos, 2005). Aunque este es un tema de estudio relativamente nuevo a nivel mundial, en México ya se ha estudiado su papel como reservorios de enfermedades para la fauna silvestre. Por ejemplo, recientemente se estudiaron los endoparásitos de perros ferales en algunas ciudades del país (Dubey et al., 2007; Jarquín-Díaz et al., 2016) y se ha visto la prevalencia de algunas enfermedades tanto en perros domésticos, como en otras especies de mamíferos silvestres con las que estos interactúan (i.e., tlacuache, ardillón mexicano, cacomixtle, zorrillo moteado y comadreja; Suzán y Ceballos, 2005; Weber, 2010). Entre las enfermedades estudiadas se encuentran el parvovirus (Moreno, 2016; Suzán y Ceballos, 2005), la rabia (Fishbein et al., 1992; Flores-Ibarra y Estrella-Valenzuela, 2004), el virus del moquillo canino (Almuna, 2016; Moreno, 2016), y las causadas por Neospora avinum (Dubey et al., 2007), T. gondii (Alvarado-Esquivel et al., 2014; Cedillo-Peláez et al., 2012; Dubey et al., 2007) y Hepatozoon canis (Jarquín-Díaz et al., 2016). A pesar de que estos estudios se han llevado a cabo en diferentes regiones de México, aún falta mucho por estudiar sobre este tema y sobre el papel que los perros domésticos pueden tener para trasmitir enfermedades a la fauna silvestre y a la población humana de nuestro país.
Como ya mencionamos anteriormente, aunque no es frecuente que ocurra en condiciones silvestres, se ha documentado la hibridación de perros con especies de cánidos silvestres como lobos y coyotes (Bee y Hall, 1951; Leonard et al., 2014; Valadez et al., 2002; VonHoldt et al., 2013; Weeks et al., 1990). Aunque las posibilidades de que ésto ocurra son bajas, en nuestro país se han encontrado rastros arqueológicos de un híbrido de lobo gris con perro doméstico en la ciudad de Teotihuacán (Valadez et al., 2002). El hecho de que ya ha ocurrido hibridación entre perros y lobos en México es alarmante debido a que uno de los efectos negativos de la hibridación entre el lobo gris y el perro en Norteamérica fue un decremento de la densidad poblacional del lobo (Beschta y Ripple, 2016; Newsome et al., 2015; Ripple et al., 2015). Debido a que en la actualidad el lobo mexicano está siendo reintroducido en diferentes regiones del país, la hibridación entre los individuos reintroducidos y perros, aumentaría la vulnerabilidad de las nuevas poblaciones y su integridad genética a futuro (Hailer y Leonard, 2008; Leonard et al., 2014; Randi, 2008).
Los efectos de los perros domésticos con dueño para nuestro país han sido poco estudiados, sin embargo, la mayoría de las publicaciones sobre perros que son mascotas han sido sobre su demografía, documentando la estructura de edades y densidad poblacional en diferentes ciudades del país (Kisiel et al., 2016; Orihuela y Solano, 1995; Ortega-Pacheco et al., 2007; Romero-López et al., 2008; Ruiz-Izaguirre et al., 2012, 2014). Estos estudios muestran principalmente 2 cosas, la alta densidad de perros que hay en México y la falta de un manejo adecuado de ellos tanto por sus dueños como por las autoridades. Por ejemplo, se ha documentado que entre 60 y 65% de las personas que viven en ciudades mexicanas son dueños de perros (Kisiel et al., 2016; Ortega-Pacheco et al., 2007). Mientras que en zonas rurales el porcentaje de personas con perros varía entre 84 y 100% (Kisiel et al., 2016; Orihuela y Solano, 1995; Ruiz-Izaguirre et al., 2014). Adicionalmente, la densidad de perros en diferentes regiones de México llega a ser muy alta. Se han estimado densidades en algunas ciudades del país que van desde los 75 hasta arriba de 1,000 perros/km2 (Orihuela y Solano, 1995; Ortega-Pacheco et al., 2007). A esto debemos sumarle que a un alto porcentaje de estos perros se les permite libre movimiento fuera de sus hogares (aproximadamente a 75%; Gompper, 2014b; Hughes y Macdonald, 2013; Kisiel et al., 2016; Ortega-Pacheco et al., 2007; Romero-López et al., 2008). Estos altos números sugieren que estudiar la ecología de los perros domésticos con dueño es crucial para establecer normas de manejo adecuadas que ayuden a reducir los impactos de los perros, tanto en el medio ambiente como en la salud humana.
¿Podemos esperar un efecto ambiental mayor de los gatos y perros domésticos en países megadiversos?
Aunque los primeros estudios de ecología de gatos y perros domésticos con dueños se comenzaron en EUA en 1941 y 1951 respectivamente (Bee y Hall, 1951; McMurry y Sperry, 1941), e India en 1975 (Oppenheimer y Oppenheimer, 1975), el impacto negativo de estas especies domésticas no fue reconocido hasta finales de la última década del siglo pasado (Barratt, 1997, 1998; Coleman et al., 1997; Weber y Dailly, 1998). A partir de ese momento, aumentó la cantidad de trabajos con estas 2 especies, sobre todo con los gatos, tanto en EUA (Coleman et al., 1997; Coleman y Temple, 1993; Crooks y Soulé, 1999) como en Australia (Barratt, 1997, 1998; Meek, 1999) y Europa (Ciucci y Boitani, 1998). Este incremento en el número de publicaciones sobre ambas especies continúa acelerándose en la década actual (fig. 1; Doherty et al., 2016; Doherty, Dickman, Glen et al., 2017; Gompper, 2014a; Hughes y Macdonald, 2013).
Asociado a las condiciones socioeconómicas, en la mayoría de los países megadiversos, principalmente en los que se encuentran en vías de desarrollo, no existe un manejo y control adecuado sobre especies invasoras más allá de malezas de importancia para la agricultura, algunos invertebrados y vertebrados en islas (Aguirre-Muñoz y Mendoza, 2009; Loss y Marra, 2017; March y Martínez 2007; Pimm et al., 2014; Simberloff, 2013). Además, el control de mascotas con dueño o de individuos ferales generalmente es muy reducido y de existir se limita a ciudades de gran tamaño (e.g., perreras), islas o algunas áreas naturales protegidas (Kikillus et al., 2017; Lilith et al., 2006; Marra y Santella, 2016). Incluso en países desarrollados, aunque los perros tienden a ser mejor manejados, con reglas de esterilización y control reproductivo, y sin que se les permita libre movimiento fuera de sus hogares, las restricciones de libre movimiento para los gatos y el control de heces de perros pueden ser inexistentes o limitada (Coleman et al., 1997; Duffy y Capece, 2012; Peterson et al., 2012). Adicionalmente en países en vías de desarrollo la población humana suele tener gatos y perros principalmente con fines utilitarios, como erradicación de roedores o protección. Estos animales suelen recibir menos cuidados que los individuos considerados como mascotas en países desarrollados y se les permite mayor movimiento fuera de los hogares de sus dueños (Gompper, 2014a; Hughes y Macdonald, 2013; Pacheco, 2003). Esto provoca que tanto gatos como perros domésticos puedan generar poblaciones ferales más fácilmente en la mayoría de los países en vías de desarrollo (Gompper, 2014b; Rinzin et al., 2016; Taeihagh, 2017).
A estas notables diferencias en el manejo que reciben los gatos y perros domésticos entre los países ricos y los países en vías de desarrollo debemos sumarle los procesos de deterioro ambiental que existen asociados al desarrollo (Gompper, 2014b; Peterson et al., 2012; Pimm et al., 2014; Taeihagh, 2017). En muchos países megadiversos tropicales existen altas tasas de deforestación y fragmentación de los hábitats nativos (Dirzo et al., 2014; Taeihagh, 2017). Este cambio de uso del suelo reduce la presencia de especies nativas de carnívoros que pueden controlar, a través de la depredación y la competencia, a los gatos y perros domésticos, además de permitir una mayor movilidad de estas 2 especies dentro de los hábitats nativos (Bellard et al., 2014; Young et al., 2017). Esta perturbación antropogénica también permite explosiones poblacionales de especies tanto nativas como invasoras que sirven como alimento a estos 2 carnívoros. Cabe resaltar el papel de la fragmentación de los hábitats naturales, que al aislar parches de vegetación nativa y aumentar las zonas de borde entre estos y las zonas agrícolas/ganaderas, aumenta la probabilidad de que especies como los gatos y perros invadan hábitats con poco uso humano (Crooks y Soulé, 1999; Manor y Saltz, 2004; Marks y Duncan, 2009; Ordeñana et al., 2010; Torres y Prado, 2010). Adicionalmente, la fragmentación reduce el hábitat disponible para las especies de vertebrados nativos, exponiéndolos más a la depredación (Crooks y Soulé, 1999; Vanak et al., 2014; Vanak y Gompper, 2010) y a los otros efectos negativos generados por los gatos y los perros (Doherty et al., 2015, 2016; Doherty, Dickman, Glen et al., 2017; Kays y DeWan, 2004).
Desde la perspectiva de los factores ecológicos que pueden aumentar los daños causados por gatos y perros domésticos debemos considerar cómo los patrones de diversidad pueden afectar la interacción entre especies invasoras y especies nativas. Sobre todo, debemos considerar la forma en la cual gatos y perros domésticos impactan a la fauna nativa. A continuación, discutimos como la alta diversidad de vertebrados de los países megadiversos, puede modificar el efecto de gatos y perros domésticos como depredadores y como reservorios de enfermedades. Simplemente por la diversidad de especies, funciones ecológicas y taxones que existen en países megadiversos tropicales, los gatos y perros domésticos deben interactuar con un mayor número de especies, con diferentes roles funcionales y con más grupos de organismos que en países no megadiversos localizados en zonas templadas (Bellard et al., 2014; Gómez-Ortiz y Moreno, 2017; Oliver et al., 2015; Young et al., 2017). Esto les permitirá depredar y competir con un mayor número de organismos que en países no megadiversos. Sin embargo, esta es una generalización que no ofrece mucha información sobre sus potenciales efectos.
En países megadiversos tropicales la mayoría de las comunidades de vertebrados tienen estructuras complejas, con unas pocas especies presentando abundancias altas y el resto de las especies presentando abundancias bajas (Dirzo et al., 2014; Marchese, 2015; Morgan et al., 2009; Pimm et al., 2014). Aunque en algunos casos, depredadores como gatos y perros suelen consumir presas con relación a su abundancia (Gompper, 2014a; Marra y Santella, 2016; Wierzbowska et al., 2012, 2016), se ha observado que los gatos en particular suelen cazar cualquier especie de tamaño adecuado a sus capacidades como depredador, sin importar su abundancia (Baker et al., 2008; McGregor et al., 2015; Spotte, 2014). De este modo, aunque estos depredadores tengan un mayor efecto sobre las especies comunes, pueden tener un efecto más alto de lo esperado sobre algunas especies raras, sobre todo aquellas que no son buenas escapando o escondiéndose de depredadores con las características de gatos y perros (Brickner, 2003; Doherty et al., 2016; Lilith et al., 2008; Loyd et al., 2013). Adicionalmente, en zonas tropicales, y en particular en países con una alta diversidad beta como México (Gómez-Ortiz y Moreno, 2017; Marchese, 2015; Molsher et al., 2005; Newsome et al., 2015; Pimm et al., 2014), es común encontrar un gran número de especies de vertebrados que presentan rangos de distribución geográfica pequeños (Marchese, 2015; Young et al., 2017), por lo cual la posibilidad de que sus poblaciones sean afectadas por un depredador invasor son mayores. Esto hace que aumente su riesgo de extinción de forma similar a lo que sucede en ambientes insulares (Medina et al., 2011; Oliver et al., 2015; Pimm et al., 2014).
También hay que considerar el efecto que los cambios en la estructura de comunidades de animales silvestres, causados por la depredación de la fauna nativa por gatos y perros domésticos, pueden tener a nivel de interacciones entre especies en países megadiversos. Por ejemplo, se ha visto que en países tropicales las redes de interacciones mutualistas entre animales y plantas (polinización y dispersión de semillas) están más anidadas, son más especializadas y menos robustas que en ecosistemas templados (Aizen et al., 2002; Dalsgaard et al., 2011; Kay y Schemske, 2004; Olesen, 2002; Schemske et al., 2009). De modo que la desaparición de especies, o la reducción del tamaño poblacional de polinizadores y dispersores de semillas causada por gatos y perros pueden tener un efecto mayor en los países tropicales megadiversos (Bellard et al., 2014; Mittermeier et al., 1997; Olesen, 2002; Schemske et al., 2009). Esto puede generar efectos cascada y cambios en la estructura de la vegetación, como los que han causado depredadores en otras partes del mundo. Ejemplo de esto es el de la extirpación de los lobos en Yellowstone y su posterior re-introducción (Beschta y Ripple, 2016; Newsome et al., 2015; Ripple et al., 2015), las cuales afectaron notoriamente las poblaciones de varias especies de plantas y vertebrados (Beschta y Ripple, 2016, 2019; Ripple et al., 2015).
Con relación a la transmisión de patógenos, tanto los gatos como los perros domésticos con dueño o ferales deben ser considerados como importantes reservorios potenciales de enfermedades. Sin embargo, la relación que hay entre especies invasoras y los efectos que estas tienen en la transmisión de enfermedades a huéspedes nativos, sobre todo en el caso de las enfermedades emergentes, ha sido un tema muy poco estudiado (Daszak, 2000; Medina-Vogel, 2010; Young et al., 2017). Se sabe que la introducción de especies exóticas en ecosistemas nativos favorece la propagación de enfermedades (Doherty et al., 2015; Pimm et al., 2014; Young et al., 2017), pudiendo tener implicaciones económicas, de salud pública y de conservación de la biodiversidad (Hooker y Innes, 1995; March y Martínez, 2007; Pimentel et al., 2001, 2005). Por ejemplo, 49% de las enfermedades humanas conocidas son de origen zoonótico y un alto porcentaje de las enfermedades infecciosas emergentes que nos pueden afectar (73%), también tienen un origen en la fauna silvestre y pueden saltar a nuestra especie desde animales domésticos (Daszak, 2000; Daszak et al., 2001; Vargas et al., 2004; Young et al., 2017). Adicionalmente, las enfermedades infecciosas son una de las 5 causas principales de extinción de especies animales a nivel mundial (Bellard et al., 2016; Dirzo et al., 2014; Pimm et al., 2014; Taeihagh, 2017; Young et al., 2017).
Actualmente, no está claro cómo las especies invasoras y su interacción con la biodiversidad afectan la dinámica de las enfermedades infecciosas (Pimm et al., 2014; Young et al., 2017). Sin embargo, hay evidencia de que las especies invasoras juegan papeles únicos en la transmisión de enfermedades que difieren de los que tienen las especies nativas (fig. 4; Doherty et al., 2015; Pimm et al., 2014; Young et al., 2017). Especies invasoras como los gatos y los perros domésticos pueden tanto reducir el número de las especies que actúan como reservorios de enfermedades por medio de la depredación o la competencia, como incrementarlo al volverse ellos mismos nuevos reservorios. En el caso de estas 2 especies, tanto su alto potencial reproductivo como su alta eficacia al cazar, les confiere un papel muy importante tanto en disminuir el número de hospederos nativos, como en convertirse en vectores potenciales (Elton, 1958; March y Martínez, 2007; Simberloff, 2013).
Por lo tanto, la poca evidencia que hay sobre este tema sugiere que tanto los gatos como los perros domésticos podrían prestar un servicio ecológico reduciendo la tasa de propagación de enfermedades infecciosas al reducir el número de especies de hospederos naturales y/o disminuir las abundancias de especies comunes dentro de comunidades con una gran diversidad de especies (Young et al., 2017). Sin embargo, esto implica que estas 2 especies de depredadores hayan cambiado de forma drástica las comunidades de vertebrados nativos, lo que puede tener un alto costo funcional y de conservación (Doherty et al., 2015, 2016). Además, en el caso de que una enfermedad local pueda infectarlos, dada su abundancia, podrían facilitar la ocurrencia de epidemias que infecten a otras especies de félidos y cánidos, otros mamíferos, o incluso a la población humana local (Acosta-Jamett, Cunningham et al., 2015; Aguirre, 2009; Baneth et al., 2016; Bevins et al., 2012; Otranto et al., 2015).
¿Tienen los gatos y perros domésticos algún efecto positivo en los sitos que invaden?
Es importante considerar que los gatos y los perros no son las únicas especies invasoras de vertebrados asociadas a los humanos. Debemos tomar en cuenta a otras especies como las ratas (Rattus rattus y R. norvegicus; Cox et al., 2000; Pimentel et al., 2000; Faulquier et al., 2009; Grant-Hoffman et al., 2010; Neinavaz et al., 2013), ratones (Mus musculus; Angel et al., 2009; Bolton et al., 2014; Harper, 2010), mangostas (Herpestes auropunctatus; Ćirović et al., 2011; Doherty et al., 2016), comadrejas (Mustela erminea; Doherty et al., 2016; Simberloff, 2013), cabras, puercos, e incluso aves (Passer domesticus, Columba livia, Myiopsitta monachus, Streptopelia decaocto por nombrar algunas; Aguirre-Muñoz, 2009; March y Martínez, 2007; Simberloff, 2013). Estas especies, que también pueden tener grandes costos ecológicos, económicos y sociales (Pimentel et al., 2005; Simberloff, 2013; Young et al., 2017), al llegar a ser abundantes, pueden funcionar como importantes presas de los gatos y perros domésticos.
Existe evidencia que muestra que los gatos domésticos cazan a un gran número de individuos de especies invasoras plaga de roedores y aves (Flux, 2007; Loss et al., 2013; Loyd et al., 2013; Orduña-Villaseñor, 2015; van Heezik et al., 2010). Por lo cual, además de considerar los daños que generan como depredadores sobre la fauna nativa, también debemos tomar en cuenta que estas especies, bajo ciertas condiciones, pueden estar prestando un servicio ecosistémico crucial a través del control de plagas, cazando cantidades considerables de ratones comunes (M. musculus), ratas (R. rattus y R. norvegicus), gorriones comunes (P. domesticus) y palomas europeas (C. livia). Todas estas especies invasoras presentan altas densidades, sobre todo en zonas urbanas (Grant-Hoffman et al., 2010; Hetmański y Wołk, 2005; MacGregor-Fors et al., 2009; Sacchi et al., 2002).
Aunque no tenemos información sobre el balance que existe entre el costo y el beneficio que puedan ofertar los gatos y los perros como controles de otras especies invasoras, es importante considerar que si reducimos el tamaño de las poblaciones de gatos y perros, debemos también manejar a las plagas controladas por estos depredadores (Mahlaba et al., 2017; Orduña-Villaseñor, 2015; Strauss et al., 2006). Este es un tema que requiere ser estudiado y que consideramos crucial para llevar a cabo un buen manejo de especies de depredadores invasores en el futuro cercano.
Conclusiones
Los gatos y perros domésticos son especies invasoras exóticas en la mayor parte de nuestro planeta. A pesar de que la ecología de estas especies ha sido poco estudiada, sobre todo en países megadiversos, es claro que ambas especies de carnívoros tienen profundos efectos negativos sobre el medio ambiente y la salud humana. La literatura existente nos permite empezar a conocer su impacto como depredadores de fauna nativa y como reservorios de enfermedades, aunque otros de sus efectos, como los impactos que su presencia genera en el comportamiento de otros vertebrados, su papel como competidores de otros carnívoros y el potencial que tienen de hibridarse con especies filogenéticamente cercanas a ellos, son básicamente desconocidos y requieren ser explorados. Sin embargo, la historia de nuestra relación con estas especies domésticas dificulta que los humanos las percibamos como la grave amenaza ambiental que son. Esto plantea importantes retos para el manejo adecuado de gatos y perros.
Dada la información existente, consideramos que gatos y perros deben de ser considerados dentro de planes de manejo a diferentes escalas geográficas para reducir sus efectos ambientales negativos, sobre todo en países megadiversos. Actualmente, ha sucedido solo en Australia (Bellard et al., 2016; Legge et al., 2017; Moseby et al., 2015). En el caso de nuestro país, tanto la alta diversidad β, como el alto número de especies endémicas de diferentes grupos de vertebrados, nos vuelve más susceptibles a los efectos negativos por depredación de ambas especies y nos obliga a considerar acciones de manejo en el corto plazo. Consideramos que para lograr un manejo adecuado de gatos y perros domésticos es necesario tener una estrategia que incluya acciones simultáneas de investigación, educación y manejo. Requerimos investigación sobre la ecología de ambas especies, sobre todo de individuos con dueño en la masa continental de nuestro país. Además de generar campañas masivas de educación sobre este tema, donde se resalten tanto los costos ambientales que tienen ambas especies, como la responsabilidad que tenemos como dueños de los efectos negativos asociados al mal manejo de estas mascotas. Estas campañas deben de nutrirse de la información generada por los proyectos de investigación para presentar información local con la que el público se identifique. Finalmente, necesitamos desarrollar una estrategia multisectorial para la aplicación de acciones de manejo en campo. Esta estrategia requiere tanto de la participación de investigadores, ONG y de la sociedad civil, como de actores gubernamentales. El gobierno debe actuar a nivel municipal para tener un mayor control de mascotas con dueño e individuos callejeros, y a nivel estatal y federal para el manejo de individuos ferales. Para lograr ésto, es necesario que haya un plan de manejo de fauna invasora a nivel gobierno, donde se señalen las instancias responsables de llevar a cabo las acciones de manejo.
Agradecimientos
El presente trabajo se realizó con el apoyo del proyecto PAPIIT IN207119. Agradecemos a Nicoletta Righini, Leticia Mirón y Pilar Rodriguez-Moreno por sus observaciones y comentarios a nuestro escrito. Agradecemos también a Edgar Pérez Negrón Souza por los diferentes apoyos prestados durante la elaboración de este artículo. El mismo constituye un requisito para la obtención del grado de Doctor en Ciencias Biológicas, del Posgrado en Ciencias Biológicas de la Universidad Nacional Autónoma de México (PCBIOL-UNAM).
Referencias
Acosta-Jamett, G. (2009). The role of domestic dogs in diseases of significance to humans and wildlife health in central Chile (Tesis doctoral). The University of Edinburgh. The Institute of Zoology, London.
Acosta-Jamett, G., Chalmers, W. S. K., Cunningham, A. A., Cleaveland, S., Handel, I. G. y Bronsvoort, B. M. C. (2011). Urban domestic dog populations as a source of canine distemper virus for wild carnivores in the Coquimbo region of Chile. Veterinary Microbiology, 152, 247–257. https://doi.org/10.1016/j.vetmic.2011.05.008
Acosta-Jamett, G., Cunningham, A. A., Bronsvoort, B. M. de C. y Cleaveland, S. (2015). Serosurvey of canine distemper virus and canine parvovirus in wild canids and domestic dogs at the rural interface in the Coquimbo Region, Chile. European Journal of Wildlife Research, 61, 329–332. https://doi.org/10.1007/s10344-014-0886-0
Acosta-Jamett, G., Surot, D., Cortes, M., Marambio, V., Valenzuela, C., Vallverdu, A. y Ward, M. P. (2015). Epidemiology of canine distemper and canine parvovirus in domestic dogs in urban and rural areas of the Araucania region in Chile. Veterinary Microbiology, 178, 260–264. https://doi.org/10.1016/j.vetmic.2015.05.012
Adams, J. R., Lucash, C., Schutte, L. y Waits, L. P. (2007). Locating hybrid individuals in the red wolf (Canis rufus) experimental population area using a spatially targeted sampling strategy and faecal DNA genotyping. Molecular Ecology, 16, 1823–1834. https://doi.org/10.1111/j.1365-294X.2007.03270.x
Aguirre-Muñoz, A., Croll, D. A., Donlan, C. J., Henry, R. W., Hermosillo, M. A., Howald, G. R. et al. (2008). High-impact conservation: invasive mammal eradications from the Islands of Western México. AMBIO: A Journal of the Human Environment, 37, 101–107. https://doi.org/10.1579/0044-7447(2008)37[101:HCIMEF]2.0.CO;2
Aguirre-Muñoz, A., Mendoza A. R., Arredondo P. B., Arriaga C. L., Campos G. E. et al. (2009). Especies exóticas invasoras: impactos sobre las poblaciones de flora y fauna, los procesos ecológicos y la economía. En Conabio (Ed.), Capital natural de México, Vol. II. Estado de conservación y tendencias de cambio, Capítulo 6 (pp. 277–318). Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Aguirre-Muñoz, A., Samaniego-Herrera, A., Luna-Mendoza, L., Ortiz-Alcaraz, A., Rodríguez-Malagón, M., Méndez-Sánchez, F. et al. (2011). Island restoration in Mexico: ecological outcomes after systematic eradications of invasive mammals. En C. R. Veitch, M. N. Clout y D. R. Towns (Eds.), 2011. Island invasives: eradication and management. IUCN (pp. 250–258). Gland, Switzerland: IUCN.
Aguirre, A. A. (2009). Wild canids as sentinels of ecological health: a conservation medicine perspective. Parasites and Vectors, 2 (S7), 1–8. https://doi.org/10.1186/1756-3305-2-S1-S7
Aizen, M. A., Vázquez, D. P. y Smith-Ramírez, C. (2002). Historia natural y conservación de los mutualismos planta-animal del bosque templado de Sudamérica austral. Revista Chilena de Historia Natural, 75, 79–97.
Alberto-Orlando, S., Calderón, J. L., León-Sosa, A., Patiño, L., Zambrano-Alvarado, M. N., Pasquel-Villa, L. D. et al. (2022). SARS-CoV-2 transmission from infected owner to household dogs and cats is associated with food sharing. International Journal of Infectious Diseases, 122, 295–299. https://doi.org/10.1016/j.ijid.2022.05.049
Almuna, M. R. (2016). Factores de riesgo asociados a tasas de infección de distemper canino en perro doméstico (Canis familiaris) y carnívoros silvestres de la Reserva de la Biosfera de Janos, Chihuahua, México (Tesis de maestría). Universidad de Chile/ UNAM/ Facultad de Ciencias Veterinarias y Pecuarias Magister en Ciencias Animales y Veterinarias.
Alvarado-Esquivel, C., Gayosso-Domínguez, E. A., Villena, I. y Dubey, J. P. (2013). Seroprevalence of Toxoplasma gondii infection in captive mammals in three zoos in Mexico City, Mexico. Journal of Zoo and Wildlife Medicine, 44, 803–806. https://doi.org/10.1638/2013-0032.1
Alvarado-Esquivel, C., Romero-Salas, D., Cruz-Romero, A., García-Vázquez, Z., Peniche-Cardeña, Á., Ibarra-Priego, N. et al. (2014). High prevalence of Toxoplasma gondii antibodies in dogs in Veracruz, Mexico. BMC Veterinary Research, 10, 4–7. https://doi.org/10.1186/s12917-014-0191-x
Álvarez-Romero, J. G., Medellín, R. A., Oliveras de Ita, A., Gómez de Silva, H. y Sánchez, O. (2008a). Canis lupus (Linnaeus 1758); Sinón. C. familiaris. Perro doméstico En J. G. Álvarez-Romero, R. A. Medellín, A. Oliveras de Ita, H. Gómez de Silva y O. Sánchez (Eds.), Animales exóticos en México: una amenaza para la biodiversidad. (pp. 35–38). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Instituto de Ecología, UNAM/ Secretaría de Medio Ambiente y Recursos Naturales.
Álvarez-Romero, J. G., Medellín, R. A., Oliveras de Ita, A., Gómez de Silva, H. y Sánchez, O. (2008b). Felis silvestris (Schreber 1775); Sinón. F. catus, gato doméstico En J. G. Álvarez-Romero, R. A. Medellín, A. Oliveras de Ita, H. Gómez de Silva y O. Sánchez (Eds.), Animales exóticos en México: una amenaza para la biodiversidad. (pp. 44–48). México D.F.: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/ Instituto de Ecología, UNAM/ Secretaría de Medio Ambiente y Recursos Naturales.
Ancillotto, L., Venturi, G. y Russo, D. (2019). Presence of humans and domestic cats affects bat behaviour in an urban nursery of greater horseshoe bats (Rhinolophus ferrumequinum). Behavioural Processes, 164, 4–9. https://doi.org/10.1016/j.beproc.2019.04.003
Anderson, T. M., VonHoldt, B. M., Candille, S. I., Musiani, M., Greco, C., Stahler. et al. (2009). Molecular and evolutionary history of melanism in North American gray wolves. Science, 323, 1339–1343. https://doi.org/10.1126/science.1165448
APPA, (2017). American pet products association. Pet industry market size and ownership statistics. Recuperado el 20 marzo, 2017 de: URL http://www.americanpetproducts.org/press_industrytrends.asp
Arnaud, G., Rodríguez, A., Ortega-Rubio, A. y Álvarez-Cárdenas, S. (1993). Predation by cats on the unique endemic lizard of Socorro Island (Urosaurus auriculatus), Revillagigedo, Mexico. Ohio Journal of Science, 93, 101–104.
Baker, P. J., Molony, S. E., Stone, E., Cuthill, I. C. y Harris, S. (2008). Cats about town: is predation by free-ranging pet cats Felis catus likely to affect urban bird populations? Ibis, 150, 86–99. https://doi.org/10.1111/j.1474-919X.2008.00836.x
Baneth, G., Thamsborg, S. M., Otranto, D., Guillot, J., Blaga, R., Deplazes, P. et al. (2016). Major parasitic zoonoses associated with dogs and cats in Europe. Journal of Comparative Pathology, 155, S54–S74. https://doi.org/10.1016/j.jcpa.2015.10.179
Banks, P. B. y Bryant, J. V. (2007). Four-legged friend or foe? dog walking displaces native birds from natural areas. Biology Letters, 3, 611–3. https://doi.org/10.1098/rsbl.2007.0374
Barratt, D. G. (1997). Predation by house cats, Felis catus (L.), in Canberra, Australia. I. Prey composition and preference. Wildlife Research, 24, 263–277.
Barratt, D. G. (1998). Predation by house cats, Felis catus (L.), in Canberra, Australia. II. Factors affecting the amount of prey caught and estimates of the impact on wildlife. Wildlife Research, 25, 475–487.
Bee, J. W. y Hall, E. R. (1951). An instance of coyote-dog hybridization. Transaction of the Kansas Academy of Science, 54, 73–77.
Bellard, C., Genovesi, P. y Jeschke, J. M. (2016). Global patterns in threats to vertebrates by biological invasions. Proceedings of the Royal Society B: Biological Sciences, 283, 20152454. https://doi.org/10.1098/rspb.2015.2454
Bellard, C., Leclerc, C., Leroy, B., Bakkenes, M., Veloz, S., Thuiller, W. y Courchamp, F. (2014). Vulnerability of biodiversity hotspots to global change. Global Ecology and Biogeography, 23, 1376–1386. https://doi.org/10.1111/geb.12228
Beschta, R. L. y Ripple, W. J. (2016). Riparian vegetation recovery in Yellowstone: The first two decades after wolf reintroduction. Biological Conservation, 198, 93–103. https://doi.org/10.1016/j.biocon.2016.03.031
Beschta, R. L. y Ripple, W. J. (2019). Can large carnivores change streams via a trophic cascade? Ecohydrology, 12, 1-13 https://doi.org/10.1002/eco.2048
Bevins, S. N., Carver, S., Boydston, E. E., Lyren, L. M., Alldredge, M., Logan, K. A., Riley, S. et al. (2012). Three pathogens in sympatric populations of pumas, bobcats, and domestic cats: Implications for infectious disease transmission. Plos One, 7, 1–10. https://doi.org/10.1371/journal.pone.0031403
Biezus, G., Machado, G., Ferian, P. E., da Costa, U. M., Pereira, L. H. H. da S., Withoeft, J. A. et al. (2019). Prevalence of and factors associated with feline leukemia virus (FeLV) and feline immunodeficiency virus (FIV) in cats of the state of Santa Catarina, Brazil. Comparative Immunology, Microbiology and Infectious Diseases, 63, 17–21. https://doi.org/10.1016/j.cimid.2018.12.004
Biró, Z., Lanszki, J., Szemethy, L., Heltai, M. y Randi, E. (2005). Feeding habits of feral domestic cats (Felis catus), wild cats (Felis silvestris) and their hybrids: trophic niche overlap among cat groups in Hungary. Journal of Zoology, 266, 187–196. https://doi.org/10.1017/S0952836905006771
Blancher, P. (2013). Estimated number of birds killed by house cats (Felis catus) in Canada. Avian Conservation and Ecology, 8, 3. https://doi.org/10.5751/ACE-00557-080203
Bolton, M., Stanbury, A., Baylis, A. M. M. y Cuthbert, R. (2014). Impact of introduced house mice (Mus musculus) on burrowing seabirds on Steeple Jason and Grand Jason Islands, Falklands, South Atlantic. Polar Biology, 37, 1659–1668. https://doi.org/10.1007/s00300-014-1554-2
Bonnaud, E., Medina, F. M., Vidal, E., Nogales, M., Tershy, B., Zavaleta, E., Donlan, C. J. et al. (2011). The diet of feral cats on islands: a review and a call for more studies. Biological Invasions, 13, 581–603. https://doi.org/10.1007/s10530-010-9851-3
Brickner, I. (2003). The impact of domestic cat (Felis catus) on wildlife welfare and conservation: a literature review. With a situation summary from Israel. Tel Aviv University Report. Recuperado el 10 noviembre, 2017 de: https://www.tau.ac.il/lifesci/zoology/members/yom-tov/inbal/cats.pdf
Calhoon, R. E. y Haspel, C. (1989). Urban cat populations compared by season, subhabitat and supplemental feeding. Journal of Animal Ecology, 58, 321–328. https://doi.org/10.2307/5003
Cedillo-Peláez, C. Díaz-Figueroa, I. D., Jiménez-Seres, M. I., Sánchez-Hernández, G. y Correa, D. (2012). Frequency of antibodies to Toxoplasma gondii in stray dogs of Oaxaca, México. Journal of Parasitology, 98, 871–872. https://doi.org/10.1645/GE-3095.1
Ćirović, D., Raković, M., Milenković, M. y Paunović, M. (2011). Small indian mongoose Herpestes auropunctatus (Herpestidae, Carnivora): An invasive species in Montenegro. Biological Invasions, 13, 393–399. https://doi.org/10.1007/s10530-010-9831-7
Ciucci, P. y Boitani, L. (1998). Wolf and dog depredation on livestock in central Italy. Wildlife Society Bulletin, 26, 504–514.
Coleman, J. S. y Temple, S. A. (1993). Rural residents’ free-ranging domestic cats: a survey. Wildlife Society Bulletin, 21, 381–390. http://www.jstor.org/stable/3783408
Coleman, J. S., Temple, S. A. y Craven, S. R. (1997). Cats and wildlife: a conservation dilemma. 1997 USFWS and University of Wisconsin Extension Report 1-4 University of Wisconsin, Cooperative Extension Publications, Room 170, 630 W. Mifflin St. Madison, WI.
Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad). (2016). Sistema de información sobre especies invasoras en México. Recuperado el 29 noviembre, 2017 de: http://www.biodiversidad.gob.mx/invasoras%0A
Cortés-Calva, P., Gallo-Reynoso, J. P., Delgadillo-Rodríguez, J., Lorenzo, C. y Álvarez-Castañeda, S. T. (2013). The effect of feral dogs and other alien species on native mammals of Isla de Cedros, México. Natural Areas Journal, 33, 466–473. https://doi.org/http://dx.doi.org/10.3375/043.033.0410
Cox, M. P. G., Dickman, C. R. y Cox, W. G. (2000). Use of habitat by the black rat (Rattus rattus) at North Head, New South Wales: an observational and experimental study. Austral Ecology, 25, 375–385. https://doi.org/10.1111/j.1442-9993.2000.tb00043.x
Crooks, K. R. y Soulé, M. E. (1999). Mesopredator release and avifaunal extinctions in a fragmented system. Nature, 400, 563–566. https://doi.org/10.1038/23028
Cuarón, A. D., de Grammont, P. C. y McFad-den, K. (2016). Procyon pygmaeus. The IUCN Red list of threatened species 2016. Recuperado el 25 mayo, 2018 de: URL http://www.iucnredlist.org/details/18267/0
Cuarón, A. D., Valenzuela-Galván, D., García-Vasco, D., Copa, M. E., Bautista, S., Mena, H. et al. (2009). Conservation of the endemic dwarf carnivores of Cozumel Island, Mexico. Small Carnivore Conservation, 41, 15–21.
Dalsgaard, B., Magård, E., Fjeldså, J., González, A. M. M., Rahbek, C., Olesen, J. M. et al. (2011). Specialization in plant-hummingbird networks is associated with species richness, contemporary precipitation and quaternary climate-change velocity. Plos One, 6, 1–7. https://doi.org/10.1371/journal.pone.0025891
Daszak, P. (2000). Emerging infectious diseases of wildlife-threats to biodiversity and human health. Science, 287, 443–449. https://doi.org/10.1126/science.287.5452.443
Daszak, P., Cunningham, A. A. y Hyatt, A. D. (2001). Anthropogenic environmental change and the emergence of infectious diseases in wildlife. Acta Tropica, 78, 103–116.
Dauphine, N. y Cooper, R. (2009). Impacts of free-ranging domestic cats (Felis catus) on birds in the United States: a review of recent research with conservation and management recommendations. Proceedings of the Fourth International Partners in Flight Conference: Tundra to Tropic, 205–219.
Dirzo, R., Young, H. S., Galetti, M., Ceballos, G., Isaac, N. J. B. y Collen, B. (2014). Defaunation in the Anthropocene. Science, 345, 401–406. https://doi.org/10.1126/science.1251817
Doherty, T. S., Dickman, C. R., Nimmo, D. G. y Ritchie, E. G. (2015). Multiple threats, or multiplying the threats? Interactions between invasive predators and other ecological disturbances. Biological Conservation, 190, 60–68. https://doi.org/10.1016/j.biocon.2015.05.013
Doherty, T. S., Glen, A. S., Nimmo, D. G., Ritchie, E. G. y Dickman, C. R. (2016). Invasive predators and global biodiversity loss. Proceedings of the National Academy of Sciences, 113, 11261–11265. https://doi.org/10.1073/pnas.1602480113
Doherty, T. S., Dickman, C. R., Glen, A. S., Newsome, T. M., Nimmo, D. G., Ritchie, E. G. et al. (2017). The global impacts of domestic dogs on threatened vertebrates. Biological Conservation, 210, 56–59. https://doi.org/10.1016/j.biocon.2017.04.007
Doherty, T. S., Dickman, C. R., Johnson, C. N., Legge, S. M., Ritchie, E. G. y Woinarski, J. C. Z. (2017). Impacts and management of feral cats Felis catus in Australia. Mammal Review, 47, 83–97. https://doi.org/10.1111/mam.12080
Donfrancesco, V., Ciucci, P., Salvatori, V., Benson, D., Andersen, L. W., Bassi, E. et al. (2019). Unravelling the scientific debate on how to address wolf-dog hybridization in Europe. Frontiers in Ecology and Evolution, 7, 1–13. https://doi.org/10.3389/fevo.2019.175
Donlan, C. J., Tershy, B. R., Keitt, B. S., Wood, B., Sánchez, J. Á., Croll, D. A. et al. (2000). Island conservation action in Northwest México. En D. H. Browne, H. Chaney y K. Mitchell (Eds.), Proceedings of the fifth California Islands symposium (pp. 330–338). Santa Barbara, CA: Santa Barbara Museum of Natural History.
Driscoll, C. A. y Macdonald, D. W. (2010). Top dogs: wolf domestication and wealth. Journal of Biology, 9, 10.1-6 https://doi.org/10.1186/jbiol226
Driscoll, C. A., Macdonald, D. W. y O’Brien, S. J. (2009). From wild animals to domestic pets, an evolutionary view of domestication. Proceedings of the National Academy of Sciences, 106, 9971–9978. https://doi.org/10.1073/pnas.0901586106
Driscoll, C. A., Menotti-Raymond, M., Roca, A. L., Hupe, K., Johnson, W. E., Geffen, E. et al. (2007). The near Eastern origin of cat domestication. Science, 317, 519–523. https://doi.org/10.1126/science.1139518
Dubey, J. P., Alvarado-Esquivel, C., Liesenfeld, O., Herrera-Flores, R. G., Ramírez-Sánchez, B. E., González-Herrera, A. et al. (2007). Neospora caninum and Toxoplasma gondii antibodies in dogs from Durango city, Mexico. The Journal of Parasitology, 93, 1033–1035. https://doi.org/10.1645/GE-1281R.1
Duffy, D. C. y Capece, P. (2012). Biology and impacts of pacific island invasive species. 7. The domestic cat (Felis catus). Pacific Science, 66, 173–212. https://doi.org/10.2984/66.2.7
Duque-García, Y., Echeverri-Zuluaga, M., Trejos-Suárez, J. y Ruiz-Sáenz, J. (2017). Prevalence and molecular epidemiology of canine parvovirus 2 in diarrheic dogs in Colombia, South America: a possible new CPV-2a is emerging? Veterinary Microbiology, 201, 56–61. https://doi.org/10.1016/j.vetmic.2016.12.039
Dyer, J. L., Yager, P., Orciari, L., Greenberg, L., Wallace, R., Hanlon, C. A. et al. (2014). Rabies surveillance in the United States during 2013. Journal of the American Veterinary Medical Association, 245, 1111–1123. https://doi.org/10.2460/javma.245.10.1111
Elton, C. S. (1958). The ecology of invasions by animals and plants. The University of Chicago Press, Chicago and London, USA.
Faulquier, L., Fontaine, R., Vidal, E., Salamolard, M. y Le Corre, M. (2009). Feral cats Felis catus threaten the endangered endemic barau’s petrel Pterodroma baraui at Reunion Island (Western Indian Ocean). Waterbirds, 32, 330–336. https://doi.org/10.1675/063.032.0213
Fishbein, D. B., Frontini, M. G., Dobbins, J. G., Flores, C. E., Quiroz, H. G., Gaméz R. J. J. et al. (1992). Prevention of canine rabies in rural México: an epidemiologic study of vaccination campaigns. American Journal of Tropical Medicine and Hygiene, 47, 317–327. doi: 10.4269/ajtmh.1992.47.317. PMID: 1524145.
Flores-Ibarra, M. y Estrella-Valenzuela, G. (2004). Canine ecology and socioeconomic factors associated with dogs unvaccinated against rabies in a Mexican city across the US-Mexico border. Preventive Veterinary Medicine, 62, 79–87. https://doi.org/10.1016/j.prevetmed.2003.10.002
Flux, J. E. C. (2007). Seventeen years of predation by one suburban cat in New Zealand. New Zealand Journal of Zoology, 34, 289–296. https://doi.org/10.1080/03014220709510087
Frantz, L. A. F., Mullin, V. E., Pionnier-Capitan, M., Lebrasseur O., Ollivier, M., Perri, A. et al. (2016). Genomic and archaeological evidence suggests a dual origin of domestic dogs. Science, 352, 1228–1231. https://doi.org/10.1126/science.aaf3161
Galov, A., Fabbri, E., Caniglia, R., Arbanasić, H., Lapalombella, S., Florijančić, T. et al. (2015). First evidence of hybridization between golden jackal (Canis aureus) and domestic dog (Canis familiaris) as revealed by genetic markers. Royal Society Open Science, 2, 1–14. https://doi.org/http://dx.doi.org/10.1098/rsos.150450
García-Aguilar, M. C. y Gallo-Reynoso, J. P. (2012). Perros ferales en la isla de Cedros, Baja California, México: una posible amenaza para los pinnípedos. Revista Mexicana de Biodiversidad, 83, 785–789. https://doi.org/10.7550/rmb.24851
Gehrt, S. D., Wilson, E. C., Brown, J. L. y Anchor, C. (2013). Population ecology of free-roaming cats and interference competition by coyotes in urban parks. Plos One 8, 1–11. https://doi.org/10.1371/journal.pone.0075718
Gingold, G., Yom-Tov, Y., Kronfeld-Schor, N. y Geffen, E. (2009). Effect of guard dogs on the behavior and reproduction of gazelles in cattle enclosures on the Golan Heights. Animal Conservation, 12, 155–162. https://doi.org/10.1111/j.1469-1795.2009.00235.x
GISD, (2017). Global Invasive Species Database 2017. Recuperado 20 noviembre, 2017 de: URL http://www.iucngisd.org/gisd
Gómez-Ortiz, Y. y Moreno, C. E. (2017). La diversidad funcional en comunidades animales: una revisión que hace énfasis en los vertebrados. Animal Biodiversity and Conservation, 40, 165–174.
Gompper, M. E. (2014a). Free-ranging dogs and Wildlife Conservation. Oxford University Press, University of Missouri, New York, USA.
Gompper, M. E. (2014b). The dog-human-wildlife interface: assessing the scope of the problem. The demographics and ownership of free-ranging dogs. En: E. M. Gompper (Ed.), Free-ranging dogs and Wildlife Conservation (pp. 9–54). Oxford University Press, University of Missouri, New York, USA.
Gottelli, D., Sillero-Zubiri, C., Applebaum, G. D., Roy, M. S., Girman, D. J., García-Moreno, J. et al. (1994). Molecular genetics of the most endangered canid: the Ethiopian wolf Canis simensis. Molecular Ecology, 3, 301–312. https://doi.org/10.1111/j.1365-294X.1994.tb00070.x
Gottelli, D., Sillero-Zubiri, C., Marino, J., Funk, S. M. y Wang, J. (2013). Genetic structure and patterns of gene flow among populations of the endangered Ethiopian wolf. Animal Conservation, 16, 234–247. https://doi.org/10.1111/j.1469-1795.2012.00591.x
Grant-Hoffman, M. N., Mulder, C. P. y Bellingham, P. J. (2010). Invasive rats alter woody seedling composition on seabird-dominated islands in New Zealand. Oecologia, 163, 449–460. https://doi.org/10.1007/s00442-009-1523-6
Hailer, F. y Leonard, J. A., (2008). Hybridization among three native North American Canis species in a region of natural sympatry. Plos One, 3, e3333. https://doi.org/10.1371/journal.pone.0003333
Harper, G. A. (2010). Habitat use by mice during winter on subantarctic Auckland Island. New Zealand Journal of Ecology, 34, 262–264.
Hernández-Cortazar, I., Acosta-Viana, K. Y., Ortega-Pacheco, A., Guzman-Marin, E. del S., Aguilar-Caballero, A. J. y Jimenéz-Coello, M. (2015). Toxoplasmosis in Mexico: epidemiological situation in humans and animals. Revista do Instituto de Medicina Tropical de Sao Paulo, 57, 93–103. https://doi.org/10.1590/S0036-46652015000200001
Hernández, S. M., Loyd, K. A. T., Newton, A. N., Gallagher, M. C., Carswell, B. L. y Abernathy, K. J. (2018). Activity patterns and interspecific interactions of free-roaming, domestic cats in managed trap-neuter-return colonies. Applied Animal Behaviour Science, 202, 63-68. https://doi.org/10.1016/j.applanim.2018.01.014
Hetmański, T. y Wołk, E. (2005). The effect of environmental factors and nesting conditions on clutch overlap in the feral pigeon Columba livia f. urbana (Gm.). Polish Journal of Ecology, 53, 523–534.
Hooker, S. y Innes, J. (1995). Ranging behaviour of forest-dwelling ship rats, Rattus rattus, and effects of poisoning with brodifacoum. New Zealand Journal of Zoology, 22, 291–304. https://doi.org/10.1080/03014223.1995.9518044
Howell, S. N. G. y Webb, S. (1995). A guide to the birds of Mexico and Northern Central America. Oxford, CA: University Press.
Hughes, J. y Macdonald, D. W. (2013). A review of the interactions between free-roaming domestic dogs and wildlife. Biological Conservation, 157, 341–351. https://doi.org/10.1016/j.biocon.2012.07.005
Jarquín-Díaz, V. H., Barbachano-Guerrero, A., Maldonado-Rodríguez, R., Vásquez-Aguilar, A. A. y Aguilar-Faisal, J. L. (2016). First molecular evidence of Hepatozoon canis in domestic dogs and ticks in fragmented rainforest areas in Mexico. Veterinary Parasitology: Regional Studies and Reports, 6, 4–8. https://doi.org/10.1016/j.vprsr.2016.11.001
Jensen, P. (2007). The behavioural biology of dogs. Massachusetts: CAB Intern. https://doi.org/10.1086/527644
Kay, K. M. y Schemske, D. W. (2004). Geographic patterns in plant-pollinator mutualistic networks: Comment. Ecology, 85, 875-878. https://doi.org/10.1890/03-3016
Kays, R. W. y DeWan, A. A. (2004). Ecological impact of inside/outside house cats around a suburban nature preserve. Animal Conservation, 7, 273–283. https://doi.org/10.1017/S1367943004001489
Keitt, B. S., Wilcox, C., Tershy, B. R., Croll, D. A. y Donlan, C. J. (2002). The effect of feral cats on the population viability of black-vented shearwaters (Puffinus opisthomelas) on Natividad Island, Mexico. Animal Conservation, 5, 217–223. https://doi.org/10.1017/S1367943002002263
Keitt, B. S. y Tershy, B. R. (2003). Cat eradication significantly decreases shearwater mortality. Animal Conservation, 6, 307–308. https://doi.org/10.1017/S1367943003003378
Keitt, B., Henry, R. W., Aguirre, M. A., García, C., Luna, M. L., Hermosillo, M. Á. et al. (2005). El impacto de los gatos introducidos (Felis catus) en el ecosistema de Isla Gudalupe. En K. Santos del Prado y E. Peters (Eds.), Isla Guadalupe: restauración y conservación (pp. 219–322). México D.F.: Secretaría de Medio Ambiente y Recursos Naturales (Semarnat)/ Instituto Nacional de Ecología (INE)/ Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE)/ Grupo de Ecología y Conservación de islas (GECI)-Secretaría de Marina-Armada.
Kikillus, H. K., Chambers, G. K., Farnworth, M. J. y Hare, K. M. (2017). Research challenges and conservation implications for urban cat management in New Zealand. Pacific Conservation Biology, 23, 15–24. https://doi.org/10.1071/PC16022
Kisiel, L. M., Jones-Bitton, A., Sargeant, J. M., Coe, J. B., Flockhart, D. T, Reynoso-Palomar, A. et al. (2016). Owned dog ecology and demography in Villa de Tezontepec, Hidalgo, Mexico. Preventive Veterinary Medicine, 135, 37–46. https://doi.org/10.1016/j.prevetmed.2016.10.021
Le Roux, J. J., Foxcroft, L. C., Herbst, M. y Macfadyen, S. (2015). Genetic analysis shows low levels of hybridization between African wildcats (Felis silvestris lybica) and domestic cats (F. s. catus) in South Africa. Ecology and Evolution, 5, 288–299. https://doi.org/10.1002/ece3.1275
Legge, S., Murphy, B. P., McGregor, H., Woinarski, J. C. Z., Augusteyn, J., Ballard, G. et al. (2017). Enumerating a continental-scale threat: How many feral cats are in Australia? Biological Conservation, 206, 293–303. https://doi.org/10.1016/j.biocon.2016.11.032
Lenth, B. E., Knight, R. L. y Brennan, M. E. (2008). The effects of dogs on wildlife communities. Natural Areas Journal, 28, 218–227. https://doi.org/10.3375/0885-8608(2008)28[218:
TEODOW]2.0.CO;2
Leonard, J. A., Echegaray, J., Randi, E. y Carles, V. (2014). Impact of hybridization with domestic dogs on the conservation of wild canids. En M. E. Gompper, (Ed.), Free-ranging dogs and Wildlife Conservation (pp. 170–184). New York: Oxford University Press.
Lepczyk, C. A., Lohr, C. A. y Duffy, D. C. (2015). A review of cat behavior in relation to disease risk and management options. Applied Animal Behaviour Science, 173, 29–39. https://doi.org/10.1016/j.applanim.2015.07.002
Li, G., Davis, B. W., Eizirik, E. y Murphy, W. J. (2016). Phylogenomic evidence for ancient hybridization in the genomes of living cats (Felidae). Genome Research, 26, 1–11. https://doi.org/10.1101/gr.186668.114
Lilith, M., Calver, M. y Garkaklis, M. (2008). Roaming habits of pet cats on the suburban fringe in Perth, Western Australia: What size buffer zone is needed to protect wildlife in reserves? Australian Zoologist, 34, 65–72. https://doi.org/10.7882/FS.2008.011
Lilith, M., Calver, M., Styles, I. y Garkaklis, M. (2006). Protecting wildlife from predation by owned domestic cats: Application of a precautionary approach to the acceptability of proposed cat regulations. Austral Ecology, 31, 176–189. https://doi.org/10.1111/j.1442-9993.2006.01582.x
Llorente-Bousquets, J. y Ocegueda, S. (2008). Estado del conocimiento de la biota. En Capital natural de México, Vol. I. Conocimiento actual de la biodiversidad (pp. 283–322). México D.F.: Conabio.
Loss, S. R. y Marra, P. P. (2017). Population impacts of free-ranging domestic cats on mainland vertebrates. Frontiers in Ecology and the Environment, 15, 502–509. https://doi.org/10.1002/fee.1633
Loss, S. R., Will, T. y Marra, P. P. (2013). The impact of free-ranging domestic cats on wildlife of the United States. Nature Communications, 4, 1396. https://doi.org/10.1038/ncomms2380
Lowry, D. A. y McArthur, K. L. (1978). Domestic dogs as predators on deer. Wildlife Society Bulletin, 6, 38–39.
Loyd, K. A. T., Hernández, S. M., Carroll, J. P., Abernathy, K. J. y Marshall, G. J. (2013). Quantifying free-roaming domestic cat predation using animal-borne video cameras. Biological Conservation, 160, 183–189. https://doi.org/10.1016/j.bio
con.2013.01.008
Luna-Mendoza, L., Barredo-Barberena, J. M., Hernández-Montoya, J. C., Aguirre-Muñoz, A., Méndez-Sánchez, F. A, Ortiz-Alcaraz, A. et al. (2011). Planning for the eradication of feral cats on Guadalupe Island, México: home range, diet, and bait acceptance. En C. R. Veitch, M. N. Clout y D. R. Towns (Eds.), Island invasives: eradication and management (pp. 192–197). Gland, Switzerland and The Centre for Biodiversity and Biosecurity (CBB), Auckland, New Zealand: IUCN.
MacGregor-Fors, I., Morales-Pérez, L., Quesada, J. y Schondube, J. E. (2009). Relationship between the presence of house sparrows (Passer domesticus) and Neotropical bird community structure and diversity. Biological Invasions, 12, 87–96. https://doi.org/10.1007/s10530-009-9432-5
Mahlaba, T. A. M., Monadjem, A., McCleery, R. y Belmain, S. R. (2017). Domestic cats and dogs create a landscape of fear for pest rodents around rural homesteads. Plos One, 12, 1–9. https://doi.org/10.1371/journal.pone.0171593
Manor, R. y Saltz, D. (2004). The impact of free-roaming dogs on gazelle kid/female ratio in a fragmented area. Biological Conservation, 119, 231–236. https://doi.org/10.1016/j.biocon.
2003.11.005
March, M. I. J. y Martínez, J. M. (2007). Especies invasoras de alto impacto a la biodiversidad. Prioridades en México. Jiutepec, Morelos, México: Instituto Mexicano de Tecnología del Agua. IMTA, Conabio, GECI, AridAmérica, The Nature Conservancy.
Marchese, C., (2015). Biodiversity hotspots: A shortcut for a more complicated concept. Global Ecology and Conservation, 3, 297–309. https://doi.org/10.1016/j.gecco.2014.12.008
Marks, B. K. y Duncan, R. S. (2009). Use of forest edges by free-ranging cats and dogs in an urban forest fragment. Southeastern Naturalist, 8, 427–436. https://doi.org/10.1656/
058.008.0305
Marra, P. P. y Santella, C. (2016). Cat wars: the devastating consequences of a cuddly killer. N. Jersey: Princeton University Press. https://doi.org/10.1111/jofo.12191
Mauti, S., Traoré, A., Sery, A., Bryssinckx, W., Hattendorf, J. y Zinsstag, J. (2017). First study on domestic dog ecology, demographic structure and dynamics in Bamako, Mali. Preventive Veterinary Medicine, 146, 44–51. https://doi.org/10.1016/j.prevetmed.2017.07.009
McChesney, G. J. y Tershy, B. R. (1998). History and status of introduced mammals and impacts to breeding seabirds on the California Channel and Northwestern Baja California Islands. Colonial Waterbirds, 21, 335–347.
McGregor, H., Legge, S., Jones, M. E. y Johnson, C. N. (2015). Feral cats are better killers in open habitats, revealed by animal-borne video. Plos One, 10, 1–12. https://doi.org/10.1371/journal.pone.0133915
McMurry, F. B. y Sperry, C. C. (1941). Food of feral house cats in Oklahoma, a progress report. Journal of Mammalogy, 22, 185–190.
Medina, F. M., Bonnaud, E., Vidal, E., Tershy, B. R., Zavaleta, E. S., Donlan, C. J. et al. (2011). A global review of the impacts of invasive cats on island endangered vertebrates. Global Change Biology 17, 3503–3510. https://doi.org/
10.1111/j.1365-2486.2011.02464.x
Medina-Vogel, G. (2010). Ecología de enfermedades infecciosas emergentes y conservación de especies silvestres. Ecology of emerging infectious diseases and wild species conservation. Archivos de Medicina Veterinaria, 42, 11–24. https://doi.org/10.4067/S0301-732X2010000100003
Meek, P. D. (1999). The movement, roaming behaviour and home range of free-roaming domestic dogs, Canis lupus familiaris, in coastal New South Wales. Wildlife Research, 26, 847. https://doi.org/10.1071/WR97101
Mella-Méndez, I. (2019). Depredación de fauna silvestre por gatos domésticos (Felis catus) en la ciudad de Xalapa, Veracruz, México (Tesis de maestría en Neuroetología). Veracruz: Universidad Veracruzana, Instituto de Neuroetología.
Mellink, E. (1993). Biological conservation of Isla de Cedros, Baja California, Mexico: assessing multiple threats. Biodiversity and Conservation, 2, 62–69.
Mellink, E. (1992). The status of Neotoma anthonyi (Rodentia, Muridae, Cricetinae) of Todos Santos Islands, Baja California, México. Bulletin Southern California Academy of Sciences, 91, 137–140.
Mellink, E., Ceballos, G. y Luévano, J. (2002). Population demise and extinction threat of the Angel de la Guarda deer mouse (Peromyscus guardia). Biological Conservation, 108, 107–111. https://doi.org/10.1016/S0006-3207(02)00095-2
Mittermeier, R. A., Robles-Gil, P. y Mittermeier, C. G. (1997). Megadiverse countries. Megadiversity. Earth’s Biologically Wealthiest Nations. CEMEX/Agrupaciaon Sierra Madre. Recuperado el 10 junio, 2017 de: https://www.biodiversitya-z.org/content/megadiverse-countries.pdf
Molsher, R., Dickman, C., Newsome, A. y Müller, W. (2005). Home ranges of feral cats (Felis catus) in Central-Western New South Wales, Australia. Wildlife Research, 32, 587–595. https://doi.org/http://dx.doi.org/10.1071/WR04093
Moreno, O. K. (2016). Estudio serológico y molecular de distemper y parvovirus canino en comunidades de carnívoros de la Reserva de la Biósfera de Janos, Chihuahua (Tesis de maestría en Ciencias de la producción y de la salud animal). Universidad Nacional Autónoma de México, Ciudad de México.
Morgan, S. A., Hansen, C. M., Ross, J. G., Hickling, G. J., Ogilvie, S. C. y Paterson, A. M. (2009). Urban cat (Felis catus) movement and predation activity associated with a Wetland Reserve in New Zealand. Wildlife Research, 36, 574–580. https://doi.org/10.1071/WR09023
Moseby, K. E., Peacock, D. E. y Read, J. L. (2015). Catastrophic cat predation: A call for predator profiling in wildlife protection programs. Biological Conservation, 191, 331–340. https://doi.org/10.1016/j.biocon.2015.07.026
Murphy, B. P., Woolley, L. A., Geyle, H. M., Legge, S. M., Palmer, R., Dickman, C. R. et al. (2019). Introduced cats (Felis catus) eating a continental fauna: the number of mammals killed in Australia. Biological Conservation, 237, 28–40. https://doi.org/10.1016/j.biocon.2019.06.013
Neinavaz, E., Barati, A., Brown, J. L., Etezadifar, F. y Emami, B. (2013). Effects of nest characteristics and black rat Rattus rattus predation on daily survival rates of great egret Ardea alba nests in mangrove forest in the Hara Biosphere Reserve, the Persian Gulf. Wildlife Biology, 19, 240–247. https://doi.org/10.2981/12-087
Neira, V., Brito, B., Agüero, B., Berrios, F., Valdés V., Gutierrez, A. et al. (2021). A household case evidences shorter shedding of SARS-CoV-2 in naturally infected cats compared to their human owners. Emerging Microbes and Infections, 10, 376–83. https://doi.org/10.1080/22221751.2020.1863132
Newsome, T. M., Dellinger, J. A., Pavey, C. R., Ripple, W. J., Shores, C. R., Wirsing, A. J. et al. (2015). The ecological effects of providing resource subsidies to predators. Global Ecology and Biogeography, 24, 1–11. https://doi.org/10.1111/geb.12236
Nowak, R. M. (2005). Walker’s carnivores of the world. Baltimore, Londres: The Johns Hopkins University Press.
Nussberger, B., Greminger, M. P., Grossen, C., Keller, L. F. y Wandeler, P. (2013). Development of SNP markers identifying European wildcats, domestic cats, and their admixed progeny. Molecular Ecology Resources, 13, 447–460. https://doi.org/10.1111/1755-0998.12075
O’Brien, J., Devillard, S., Say, L., Vanthomme, H., Léger, F., Ruette, S. y Pontier, D. (2009). Preserving genetic integrity in a hybridising world: are European wildcats (Felis silvestris silvestris) in eastern France distinct from sympatric feral domestic cats? Biodiversity and Conservation, 18, 2351–2360. https://doi.org/10.1007/s10531-009-9592-8
O’Brien, S. J. y Johnson, W. E. (2007). The evolution of cats. Scientific American, 297, 68–75. https://doi.org/10.1038/scientificamerican0707-68
Olesen, J. M. (2002). Geographic patterns in plant-pollinator mutualistic networks. Ecology, 83, 2416-2424. DOI:10.2307/307180383
Oliveira, R., Godinho, R., Randi, E. y Alves, P. C. (2008). Hybridization versus conservation: are domestic cats threatening the genetic integrity of wildcats (Felis silvestris silvestris) in Iberian Peninsula? Philosophical Transactions of the Royal Society B. Biological Sciences, 363, 2953–2961. https://doi.org/10.1098/rstb.2008.0052
Oliver, T. H., Heard, M. S., Isaac, N. J. B., Roy, D. B., Procter, D., Eigenbrod, F. et al. (2015). Biodiversity and resilience of ecosystem functions. Trends in Ecology and Evolution, 30, 673–684. https://doi.org/10.1016/j.tree.2015.08.009
OMS (Organización Mundial de la Salud). (2019). Rabia. Recuperado el 26 agosto, 2019 de: https://www.who.int/es/news-room/fact-sheets/detail/rabies
Oppenheimer, E. C. y Oppenheimer, J. R. (1975). Certain behavioral features in the pariah dog (Canis familiaris) in West Bengal. Applied Animal Ethology, 2, 81–92. https://doi.org/10.1016/0304-3762(75)90067-X
Ordeñana, M. A., Crooks, K. R., Boydston, E. E., Fisher, R. N., Lyren, L. M., Siudyla, S. et al. (2010). Effects of urbanization on carnivore species distribution and richness. Journal of Mammalogy, 91, 1322–1331. https://doi.org/10.1644/09-MAMM-A-312.1
Orduña-Villaseñor, M. V. (2015). Dieta de gato doméstico Felis silvestris catus en el Municipio de Morelia, Michoacán (Tesis de maestría). Centro de Investigaciones en Ecosistemas, Ecología. Posgrado en Ciencias Biológicas, Universidad Nacional Autónoma de México. Ciudad de México, México.
Orihuela, T. A. y Solano, V. J. (1995). Demographics of the
owned dog population in Miacatlán, Mor. Mexico. Anthrozoos, VIII, 171–175.
Ortega-Pacheco, A., Rodríguez-Buenfil, J. C., Bolio-González, M. E., Sauri-Arceo, C. H., Jiménez-Coello, M. y Forsberg, C. L. (2007). A survey of dog populations in urban and rural areas of Yucatan, Mexico. Anthrozoos, 20, 261–274. https://doi.org/10.2752/089279307X224809
Ortiz-Alcaraz, A., Aguirre-Muñoz, A., Arnaud-Franco, G., Galina-Tessaro, P., Rojas-Mayoral, E., Méndez-Sánchez, F. et al. (2017). Progress in the eradication of the feral cat (Felis catus) and recovery of the native fauna on Socorro Island, Revillagigedo Archipelago, Mexico. Therya, 8, 3–9. https://doi.org/10.12933/therya-17-425
Otranto, D., Cantacessi, C., Pfeffer, M., Dantas-torres, F., Brianti, E., Deplazes, P. et al. (2015). The role of wild canids and felids in spreading parasites to dogs and cats in Europe Part I: Protozoa and tick-borne agents. Veterinary Parasitology, 213, 12–23. https://doi.org/10.1016/j.vetpar.2015.04.022
Pacheco, R. A. (2003). Mascotas en los hogares: enfermedades de los niños adquiridas por convivencia con animales. Enfermedades Infecciosas y Microbiologia, 23, 137–148.
Paniz Mondolfi, A. E., Colmenares, G. A., Mendoza, P. Y., Hernández-Pereira, C. E., Medina, C., Vargas, F. et al. (2019). Autochthonous cutaneous leishmaniasis in urban domestic animals (Felis catus / Canis lupus familiaris) from central-western Venezuela. Acta Tropica, 191, 252–260. https://doi.org/10.1016/j.actatropica.2019.01.006
Parsons, A. W., Bland, C., Forrester, T., Baker-Whatton, M. C., Schuttler, S. G., McShea, W. J. et al. (2016). The ecological impact of humans and dogs on wildlife in protected areas in eastern North America. Biological Conservation, 203, 75–88. https://doi.org/10.1016/j.biocon.2016.09.001
Peterson, M. N., Hartis, B., Rodriguez, S., Green, M. y Lepczyk, C. A. (2012). Opinions from the front lines of cat colony management conflict. Plos One, 7, 44616. https://doi.org/10.1371/journal.pone.0044616
Pierpaoli, M., Birò, Z. S., Herrmann, M., Hupe, K., Fernandes, M., Ragni, B. et al. (2003). Genetic distinction of wildcat (Felis silvestris) populations in Europe, and hybridization with domestic cats in Hungary. Molecular Ecology, 12, 2585–2598. https://doi.org/10.1046/j.1365-294X.2003.01939.x
Pilot, M., Moura, A. E., Okhlopkov, I. M., Mamaev, N. V., Manaseryan, N. H., Hayrapetyan, V. et al. (2021). Human-modified canids in human-modified landscapes: The evolutionary consequences of hybridization for grey wolves and free-ranging domestic dogs. Evolutionary Applications, 14, 2433–2456. https://doi.org/10.1111/eva.13257
Pimentel, D., Lach, L., Zuniga, R. y Morrison D. (2000). Environmental and economic costs of nonindigenous species in the United States. Bioscience, 50, 53–65.
Pimentel, D., McNair, S., Janecka, J., Wightman, J., Simmonds, C., O’Connell, C. et al. (2001). Economic and environmental threats of alien plant, animal, and microbe invasions. Agriculture, Ecosystems and Environment, 84, 1–20. https://doi.org/10.1016/S0167-8809(00)00178-X
Pimentel, D., Zuniga, R. y Morrison, D. (2005). Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecological Economics, 52, 273–288. https://doi.org/10.1016/j.ecolecon.2004.10.002
Pimm, S. L., Jenkins, C. N., Abell, R., Brooks, T. M., Gittleman, J. L., Joppa, L. N. et al. (2014). The biodiversity of species and their rates of extinction, distribution, and protection. Science, 344, 1246752–1246752. https://doi.org/10.1126/science.1246752
Popova, E. y Zlatanova, D. (2020). Living a dog’s life: A putative gray wolf in a feral dog group. Mammalia, 84, 115–120. https://doi.org/10.1515/mammalia-2019-0010
Randi, E. (2008). Detecting hybridization between wild species and their domesticated relatives. Molecular Ecology, 17, 285–293. https://doi.org/10.1111/j.1365-294X.2007.03417.x
Raynor, B., De la Puente-León, M., Johnson, A., Díaz, E. W., Levy, M. Z., Recuenco, S. E. et al. (2020). Movement patterns of free-roaming dogs on heterogeneous urban landscapes: Implications for rabies control. Preventive Veterinary Medicine, 178, 104978. https://doi.org/10.1016/j.prevetmed.2020.104978
Ribeiro, F. S., Nichols, E., Morato, R. G., Metzger, J. P. y Pardini, R. (2019). Disturbance or propagule pressure? Unravelling the drivers and mapping the intensity of invasion of free-ranging dogs across the Atlantic forest hotspot. Diversity and Distributions, 25, 191–204. https://doi.org/10.1111/ddi.12845
Rinzin, K., Tenzin, T. y Robertson, I. (2016). Size and demography pattern of the domestic dog population in Bhutan: implications for dog population management and disease control. Preventive Veterinary Medicine, 126, 39–47. https://doi.org/10.1016/j.prevetmed.2016.01.030
Ripple, W. J., Beschta, R. L. y Painter, L. E. (2015). Trophic cascades from wolves to alders in Yellowstone. Forest Ecology and Management, 354, 254–260. https://doi.org/10.1016/j.foreco.2015.06.007
Rodríguez-Estrella, R., Arnaud, G., Álvarez-Cardenas, S. y Rodríguez, A. (1991). Predation by Feral Cats on Birds at Isla Socorro, Mexico. Western Birds, 22, 141–143.
Rodríguez-Moreno, A., Gustavo, A. y Bernie, T. (2007). Impacto de la erradicacion del gato (Felis catus), en dos roedores endémicos de la isla Coronados, golfo De California, México. Acta Zoológica Mexicana, 23, 1–13. https://doi.org/10.21829/azm.2007.231552
Romero-López, J. A., Jaramillo-Arango, C. J., Martínez-Maya, J. J., Álvarez Peralta, E. y Terrones, C. R. (2008). Study of the population structure of dogs in a political district in Mexico City. Journal of Animal and Veterinary Advances, 7, 1352–1357. https://medwelljournals.com/abstract/?doi=javaa.2008.1352.1357
Ruiz-Izaguirre, E. y Eilers, C. H. A. M. (2012). Perceptions of village dogs by villagers and tourists in the coastal region of rural Oaxaca, Mexico. Anthrozoos, 25, 75–91. https://doi.org/10.2752/175303712X13240472427555
Ruiz-Izaguirre, E., Eilers, K. H. A. M., Bokkers, E. A. M., Ortolani, A., Ortega-Pacheco, A. y de Boer, I. J. M. (2014). Human-dog interactions and behavioural responses of village dogs in coastal villages in Michoacán, Mexico. Applied Animal Behaviour Science, 154, 57–65. https://doi.org/10.1016/j.applanim.2014.02.002
Sacchi, R., Gentilli, A., Razzetti, E. y Barbieri, F. (2002). Effects of building features on density and flock distribution of feral pigeons Columba livia var. domestica in an urban environment. Canadian Journal of Zoology, 80, 48–54. https://doi.org/10.1139/z01-202
Schemske, D. W., Mittelbach, G. G., Cornell, H. V., Sobel, J. M. y Roy, K. (2009). Is there a latitudinal gradient in the importance of biotic interactions? Annual Review of Ecology, Evolution, and Systematics, 40, 245–269. https://doi.org/10.1146/annurev.ecolsys.39.110707.173430
Silva-Rodríguez, E. A., Ortega-Solís, G. R. y Jiménez, J. E. (2010). Conservation and ecological implications of the use of space by chilla foxes and free-ranging dogs in a human-dominated landscape in southern Chile. Austral Ecology, 35, 765–777. https://doi.org/10.1111/j.1442-9993.2009.02083.x
Silva-Rodríguez, E. A. y Sieving, K. E. (2012). Domestic dogs shape the landscape-scale distribution of a threatened forest ungulate. Biological Conservation, 150, 103–110. https://doi.org/10.1016/j.biocon.2012.03.008
Silva-Rodríguez, E. A. y Sieving, K. E. (2011). Influence of care of domestic carnivores on their predation on vertebrates. Conservation Biology, 25, 808–815. https://doi.org/10.1111/j.1523-1739.2011.01690.x
Simberloff, D. (2013). Invasive species what everyone needs to know. Oxford: Universidad de New York.
Soto, C. A. y Palomares, F. (2015). Human-related factors regulate the presence of domestic dogs in protected areas. Oryx, 49, 254–260. https://doi.org/10.1017/S0030605313000604
Spotte, S. (2014). Free-ranging cats: behavior, ecology, management. New Jersey: John Wiley and Sons. https://doi.org/10.1002/9781118884058
Strauss, S. Y., Webb, C. O. y Salamin, N. (2006). Exotic taxa less related to native species are more invasive. Proceedings of the National Academy of Sciences, 103, 5841–5845. https://doi.org/10.1073/pnas.0508073103
Suzán, G. y Ceballos, G. (2005). The role of feral mammals on wildlife infectious disease prevalence in two nature reserves within Mexico City limits. Journal of Zoo and Wildlife Medicine, 36, 479–484. https://doi.org/10.1638/04-078.1
Taeihagh, A. (2017). Crowdsourcing, sharing economies and development. Journal of Developing Societies, 33, 191–222. https://doi.org/10.1177/0169796X17710072
Tershy, B. R., Donlan, C. J., Keitt, B. S., Croll, D. A, Sanchez, J. A, Wood, B. et al. (2002). Island conservation in north-west Mexico: a conservation model integrating research, education and exotic mammal eradication. En C. R. Veitch y M. N. Clout (Eds.), Turning the tide: the eradication of invasive species (pp. 293–300). Gland, Switzerland and Cambridge, UK: IUCN/ SSC Invasive Species Specialist Group.
Torres, P. C. y Prado, P. I. (2010). Domestic dogs in a fragmented landscape in the Brazilian Atlantic Forest: abundance, habitat use and caring by owners. Brazilian Journal of Biology, 70, 987–994. https://doi.org/10.1590/S1519-69842010000500010
Valadez, R., Rodríguez, B., Manzanilla, L. y Tejeda, S. (2002). Dog-wolf hybrid biotype reconstruction from the archaeological city of Teotihuacan in Prehispanic Central Mexico. En L. M. Snyder y E. A. Moore (Eds.), Dogs and people in social, working, economic or symbolic interaction. 9th ICAZ Conference, Durham (pp. 120–130). México D.F.
van Heezik, Y., Smyth, A., Adams, A. y Gordon, J. (2010). Do domestic cats impose an unsustainable harvest on urban bird populations? Biological Conservation, 143, 121–130. https://doi.org/10.1016/j.biocon.2009.09.013
Vanak, A. T., Dickman, C. R., Silva-Rodríguez, E. A., James R. A. Butler y Ritchie E. G. (2014). Chapter 3 Top-dogs and under-dogs: competition between dogs and sympatric carnivores, En M. E. Gompper (Ed.), Free-ranging dogs and Wildlife Conservation (pp. 69–93) Columbia, MO: University of Missouri.
Vanak, A. T. y Gompper, M. E. (2009). Dogs Canis familiaris as carnivores: Their role and function in intraguild competition. Mammal Review, 39, 265–283. https://doi.org/10.1111/j.1365-2907.2009.00148.x
Vanak, A. T. y Gompper, M. E. (2010). Interference competition at the landscape level: The effect of free-ranging dogs on a native mesocarnivore. Journal of Applied Ecology, 47, 1225–1232. https://doi.org/10.1111/j.1365-2664.2010.01870.x
Vanak, A. T., Thaker, M. y Gompper, M. E. (2009). Experimental examination of behavioural interactions between free-ranging wild and domestic canids. Behavioral Ecology and Sociobiology, 64, 279–287. https://doi.org/10.1007/s00265-009-0845-z
Vargas, C., Soto, L., Rojas, M. E. y Breña, P. (2004). Enfermedades de mascotas en humanos. Revisión actualizada pet diseases in humans. Updated review. Revista de la Sociedad Peruana de Medicina Interna, 17, 17–26.
Vázquez-Domínguez, E., Ceballos, G. y Cruzado, J. (2004). Extirpation of an insular subspecies by a single introduced cat: The case of the endemic deer mouse Peromyscus guardia on Estanque Island, Mexico. Oryx, 38, 347–350. https://doi.org/10.1017/S0030605304000602
Verardi, A., Lucchini, V. y Randi, E. (2006). Detecting introgressive hybridization between free-ranging domestic dogs and wild wolves (Canis lupus) by admixture linkage disequilibrium analysis. Molecular Ecology, 15, 2845–2855. https://doi.org/10.1111/j.1365-294X.2006.02995.x
Vigne, J. D. (2004). Early taming of the cat in Cyprus. Science, 304, 259–259. https://doi.org/10.1126/science.1095335
VonHoldt, B. M., Pollinger, J. P., Earl, D. A., Parker, H. G., Ostrander, E. A. y Wayne, R. K. (2013). Identification of recent hybridization between gray wolves and domesticated dogs by SNP genotyping. Mammalian Genome, 24, 80–88. https://doi.org/10.1007/s00335-012-9432-0
Watts, D. E. y Benson, A. M. (2016). Prevalence of antibodies for selected canine pathogens among wolves (Canis lupus) from the Alaska Peninsula, USA. Journal of Wildlife Diseases, 52, 506–515. https://doi.org/10.7589/2015-06-140
Weber, J. M. y Dailly, L. (1998). Food habits and ranging behaviour of a group of farm cats (Felis catus) in a Swiss mountainous area. Journal of Zoology, 245, 234–237. https://doi.org/10.1017/S0952836998286092
Weber, M. (2010). Perros (Canis lupus familiaris) y gatos (Felis catus) ferales en la Reserva de la Biosfera Los Petenes, Campeche, México: diagnóstico, efectos en la fauna nativa y perspectivas de control. Informe de Proyecto. Campeche: PNUD-Conanp-SDP.
Weeks, J. L., Tori, G. M. y Shieldcastle, M. C. (1990). Coyotes (Canis latrans) in Ohio. Ohio Journal of Science, 90, 142–145.
Weston, M. A., Fitzsimons, J. A., Wescott, G., Miller, K. K., Ekanayake, K. B. y Schneider, T. (2014). Bark in the park: A review of domestic dogs in parks. Environmental Management, 54, 373–382. https://doi.org/10.1007/s00267-014-0311-1
Wierzbowska, I. A., Hędrzak, M., Popczyk, B., Okarma, H. y Crooks, K. R. (2016). Predation of wildlife by free-ranging domestic dogs in Polish hunting grounds and potential competition with the grey wolf. Biological Conservation, 201, 1–9. https://doi.org/10.1016/j.biocon.2016.06.016
Wierzbowska, I. A., Olko, J., Hedrzak, M. y Crooks, K. R. (2012). Free-ranging domestic cats reduce the effective protected area of a Polish national park. Mammalian Biology, 77, 204–210. https://doi.org/10.1016/j.mambio.2012.01.004
Winter, L. (2006). Impacts of feral and free-ranging cats on bird species of conservation concern. Other Publications in Wildlife Management, 28. DigitalCommons@ University of Nebraska – Lincoln. Recuperado el 22 marzo, 2012 de: https://digitalcommons.unl.edu/icwdmother/28; http://www.abcbirds.org/newsandre-ports/NFWF.pdf
Wood, B., Tershy, B. R., Hermosillo, M. A, Donlan, C. J., Sánchez, J. A, Keitt, B. S. et al. (2002). Removing cats from islands in north-west Mexico. En C. R. Veitch y M. N. Clout (Eds.), Turning the tide: the eradication of invasive species (pp. 374–380). Gland, Switzerland and Cambridge, UK: IUCN/ SSC Invasive species specialist group.
Woods, M., Mcdonald, R. A. y Harris, S. (2003). Predation of wildlife by domestic cats Felis catus in Great Britain. Mammal Review, 33, 174–188. https://doi.org/10.1046/j.1365-2907.2003.00017.x
Yip, S. J. S., Dickman, C. R., Denny, E. A. y Cronin, G. M. (2014). Diet of the feral cat, Felis catus, in central Australian grassland habitats: Do cat attributes influence what they eat? Acta Theriologica, 59, 263–270. https://doi.org/10.1007/s13364-013-0166-5
Young, H. S., Parker, I. M., Gilbert, G. S., Guerra, A. S. y Nunn, C. L. (2017). Introduced species, disease ecology, and biodiversity-disease relationships. Trends in Ecology and Evolution, 32, 41–54. https://doi.org/10.1016/j.tree.2016.09.008
Young, J. K., Olson, K. A., Reading, R. P., Amgalanbaatar, S. y Berger, J. (2011). Is wildlife going to the dogs? impacts of feral and free-roaming dogs on wildlife populations. Bioscience, 61, 125–132. https://doi.org/10.1525/bio.2011.61.2.7
Zapata-Ríos, G. y Branch, L. C. (2018). Mammalian carnivore occupancy is inversely related to presence of domestic dogs in the high Andes of Ecuador. Plos One, 13, 0192346. https://doi.org/10.1371/journal.pone.0192346


