Masa relativa de la camada en Phrynosoma orbiculare (Squamata: Phrynosomatidae) en el Parque Estatal Sierra Morelos, Toluca, Estado de México
Orlando Suárez-Rodríguez a, *, Gabriel Suárez-Varón a, Fernanda Chávez-Siles a, Fernanda Pérez-Arriaga a, Gabriel Andrade-Soto a, Lizet Aguilar-Isaac a, Wade C. Sherbrooke b y Oswaldo Hernández-Gallegos a
a Laboratorio de Herpetología, Facultad de Ciencias, Universidad Autónoma del Estado de México, Instituto Literario #100 Centro, 50000 Toluca, Estado de México, México
b Southwestern Research Station, American Museum of Natural History, P.O. Box 16553, Portal, Arizona, 85632 USA
*Autor para correspondencia: biol.osr.herp@gmail.com (O. Suárez-Rodríguez)
Resumen
El tamaño y masa relativa de la camada y las particularidades de las crías son características importantes de historia de vida. El presente estudio aporta información sobre el tamaño y masa relativa de la camada y características de las crías de Phrynosoma orbiculare en el Parque Estatal Sierra Morelos, Toluca, Estado de México (± 2,700 m snm). Las hembras gestantes tuvieron una LHC de 75.2 ± 2.3 mm (n = 14), con un tamaño de camada de 8.3 ± 0.6 crías y una masa relativa de la camada de 27.2 ± 1.5 g. Las hembras de mayor talla y peso tuvieron un mayor número de crías, una situación común en lagartijas de ambientes templados; sin embargo, las hembras más ligeras tuvieron una mayor masa relativa de la camada que hembras de mayor peso; ambos resultados han sido registrados en lagartijas, incluyendo especies del género Phrynosoma. Las crías tuvieron una LHC de 26.1 ± 0.1 mm y un peso de 1.1 ± 0.01 g (sin diferencia sexual) y existe una variación baja en ambas características; la proporción sexual fue 1:1.
Palabras clave:
Reproducción; Tamaño de camada; Características de historia de vida; Proporción sexual
© 2018 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Effect of grazing on a nocturnal rodent community in grasslands of Perote Valley, Veracruz
Abstract
Clutch size, relative clutch mass and traits of hatchlings are important life history traits. The present study provides information on clutch size, relative clutch mass and traits of hatchlings of a population of Phrynosoma orbiculare occurring in the Parque Estatal Sierra Morelos, Toluca, Estado de México (± 2,700 m asl). Gravid females (n = 14) having SVL of 75.2 ± 2.3 mm produced clutch sizes of 8.3 ± 0.6 hatchlings, and relative clutch masses of 27.2 ± 1.5 g. Females of greater size and weight had higher numbers of live-born offspring, a common situation in lizards of temperate habitats, whereas lighter weight females had higher relative clutch mass than heavier females; both results have been recorded in lizards, including species of Phrynosoma. Neonates had SVL of 26.1 ± 0.1 mm, and weights of 1.1 ± 0.01 g (without sexual difference), we found a very low variation in both traits; the sex ratio was 1:1.
Keywords:
Reproduction; Clutch size; Life history traits; Sex ratio
Introducción
En México, los estudios que abordan la masa relativa de la camada se han realizado en diferentes familias de lagartijas, entre las que se encuentran: Phrynosomatidae (Sceloporus, Bastiaans et al., 2013; Rodríguez-Romero et al., 2002, 2004, 2005), Iguanidae (Ctenosaura, Castro-Franco et al., 2011), Polychrotidae (Anolis, Ramírez-Bautista y Vitt, 1997), Scincidae (Plestiodon, Muñoz-Nolasco, 2015), Teiidae (Aspidoscelis, Ramírez-Bautista et al., 2000; Rodríguez-Romero et al., 2005), Xantusiidae (Lepidophyma, Rodríguez-Romero et al., 2005) y Xenosauridae (Xenosaurus, Zamora-Abrego et al., 2007).
Se ha documentado dentro del género Phrynosoma una inversión reproductora elevada en comparación con otras especies (Barbault, 1988; Pianka, 1981, 1986), lo cual aparentemente obedece a su forma corporal robusta (Pianka, 1981) y a las características asociadas a su modo de forrajeo pasivo (Vitt y Price, 1982). Este género incluye especies ovíparas y vivíparas; las 7 especies vivíparas pertenecen a 2 clados: Tapaja y Brevicauda (Leaché y McGuire, 2006; Nieto-Montes de Oca et al., 2014); dentro del clado Tapaja se reconocen 4 especies: P. hernandesi, P. douglasii, P. ditmarsi y P. orbiculare (Nieto-Montes de Oca et al., 2014).
La lagartija P. orbiculare es de forrajeo pasivo, endémica de México, se reproduce durante otoño e invierno, habita en bosques de pino-encino y áreas de matorral semiárido, en altitudes que oscilan entre 1,500 y 3,400 m snm (Hodges, 2002; Lemos-Espinal y Smith, 2007; Méndez-de la Cruz et al., 2003; Sherbrooke, 2003). Su amplia distribución geográfica (Bryson et al., 2012) (desde Chihuahua hasta el sur de Puebla y Guerrero, así como del sur de Nuevo León al Eje Neovolcánico Transversal y la Meseta Central Mexicana), puede promover variación en características reproductoras (Du et al., 2005) y sería muy importante analizar las causas de dichas variaciones. Sin embargo, los estudios relacionados con su tamaño y masa relativa de la camada son escasos y en su mayoría anecdóticos, con tamaños de muestra limitados (Davis y Smith, 1953; Goldberg, 2011; Hernández-Ibarra et al., 2000; Lemos-Espinal y Smith, 2007; Vázquez-Díaz y Quintero-Díaz, 2005) y cuando incluyen una muestra representativa los ejemplares corresponden a diferentes poblaciones, elevaciones y tipos de vegetación (Hodges, 2004; Pianka y Parker, 1975), lo cual puede promover variaciones significativas en atributos de historia de vida. Vinculado a esta situación, en estudios sobre atributos reproductores se ha fortalecido la idea que la unidad de interés principal debe ser la población más que la especie (Du et al., 2005). El presente estudio incorpora información del tamaño, masa relativa y características de la camada de P. orbiculare en una población ubicada en el Parque Estatal Sierra Morelos, Toluca, Estado de México. Estos resultados se comparan con diversas especies de lagartijas, particularmente pertenecientes al género Phrynosoma del clado Tapaja.
Materiales y métodos
Los datos del tamaño y masa relativa de la camada, LHC y peso de las crías se analizaron mediante una regresión lineal, utilizando la LHC, PT y PA de las hembras como variables independientes. Se comparó la LHC y el peso de las crías por sexo con una t de student. Con una prueba de X2 (Zar, 2010) se analizó la proporción sexual de las crías. Todos los datos fueron registrados como media ± error estándar, intervalo, coeficiente de variación (C.V. = desviación estándar/media y el resultado expresado en porcentaje) y tamaño de muestra; se consideró un alfa del 0.05. Los análisis estadísticos se realizaron en Statgraphics Centurion XVI.I®.
Resultados
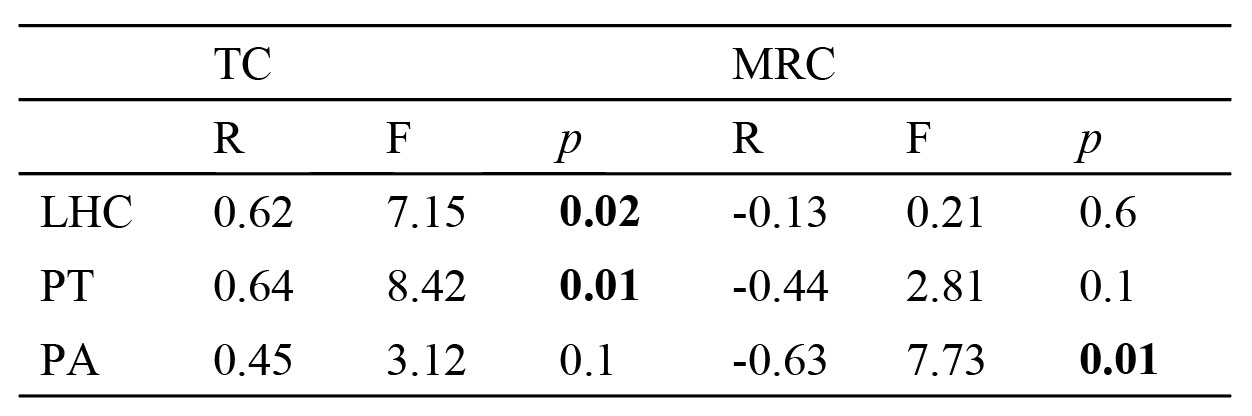
Valores de R, F y p para los análisis de regresión simple de la longitud hocico-cloaca (LHC), peso total (PT) y peso absoluto (PA) de las hembras de Phrynosoma orbiculare con el tamaño de camada (TC) y la masa relativa de la camada (MRC) en el Parque Estatal Sierra Morelos, Toluca, Estado de México. Donde R = coeficiente de correlación, F = prueba de F y p = valor de probabilidad. Los valores en negritas indican relaciones significativas.
Discusión
Los resultados de este estudio difieren de los registrados previamente por Pianka y Parker (1975) para P. orbiculare (especialmente en la MRC), ya que las hembras del PESM tienen una MRC promedio de más del doble (27.2 vs. 12.8 [que resulta muy bajo para el género cuyos valores son ≥ 20], respectivamente), un número de crías ligeramente menor (8.3 crías vs. 9.4 crías, respectivamente) y una maduración sexual a una menor talla (51 mm de LHC vs. 67 mm de LHC, respectivamente). Bryson et al. (2012) encontraron que P. orbiculare está ampliamente asociada a bosques de pino-encino en elevaciones ≥ 1,900 m; mientras que según Montanucci (1981) y Sherbrooke (2003) esta lagartija habita también en matorral semiárido y en pastizales. Como se enunció previamente, la muestra de P. orbiculare utilizada por Pianka y Parker (1975) fue conformada por individuos de diferentes localidades (i.e., = diferentes ambientes). Dicha heterogeneidad ambiental en la muestra de Pianka y Parker (1975), posiblemente repercute en la diferencia en el grado de mirmecofagia. Rojas-Hernández et al. (2016) registraron una marcada mirmecofagia en P. orbiculare del PESM (92.2% y 72.8%, en número y en volumen, respectivamente), el cual es mayor al grado de mirmecofagia registrado por Pianka y Parker (1975; 67.5% y 25.9%, en número y en volumen, respectivamente). Es dificil saber cuánta de la diferencia en la MRC y dieta es debida a un aspecto biológico y cuánta es debida a la mezcla de diferentes poblaciones (o algún otro aspecto metodológico). Sin embargo, un aspecto adicional (no excluyente) puede ser el principal responsable de la diferencia en la MRC entre ambos estudios. Mientras que Pianka y Parker (1975) utilizaron ejemplares depositados en museos, los datos del presente estudio fueron obtenidos de individuos vivos. En este sentido se ha registrado que los ejemplares de museo pueden ganar o perder peso de forma significativa y con ello afectar sustancialmente el valor de la MRC (Martin, 1978).
Los valores de LHC y de TC de las hembras de P. orbiculare del PESM sexualmente maduras, son las más bajos cuando los comparamos con los del resto de las especies del clado Tapaja (tabla 2) y en general con las especies del género Phrynosoma (tabla 11 en Pianka y Parker, 1975); cabe señalar que el 92.9% de las hembras de P. orbiculare maduran a una LHC ≥ 66 mm, la cual es muy similar a las especies dentro del clado y del género (tabla 2; tabla 11 en Pianka y Parker, 1975). Con excepción de la hembra prematura de 51 mm de LHC, una LHC > 66 mm es alcanzada durante su segundo año de vida en hembras de P. orbiculare (Hernández-Navarrete et al., 2016), lo cual coincide con la edad de madurez sexual de las especies dentro del género Phrynosoma incluyendo aquéllas del clado Tapaja (tabla 11 en Pianka y Parker, 1975), cuya tasa de crecimiento impide la madurez sexual y reproducción en el mismo año de su nacimiento (Hernández-Navarrete et al., 2016), contrario a lo que sucede en otras lagartijas vivíparas de ambiente templado en el centro de México (Rodríguez-Romero et al., 2011).
Longitud hocico-cloaca (LHC) a la madurez sexual y atributos reproductores de las hembras del género Phrynosoma del clado Tapaja. Las especies están ordenadas de acuerdo con la filogenia propuesta por Nieto-Montes de Oca et al. (2014). ND = datos no disponibles. MRC obtenida por el método de Tinkle (1972)* y de acuerdo con Cuellar (1984)**; *** = P. hernandesi; **** = P. douglassi + P. hernandesi.
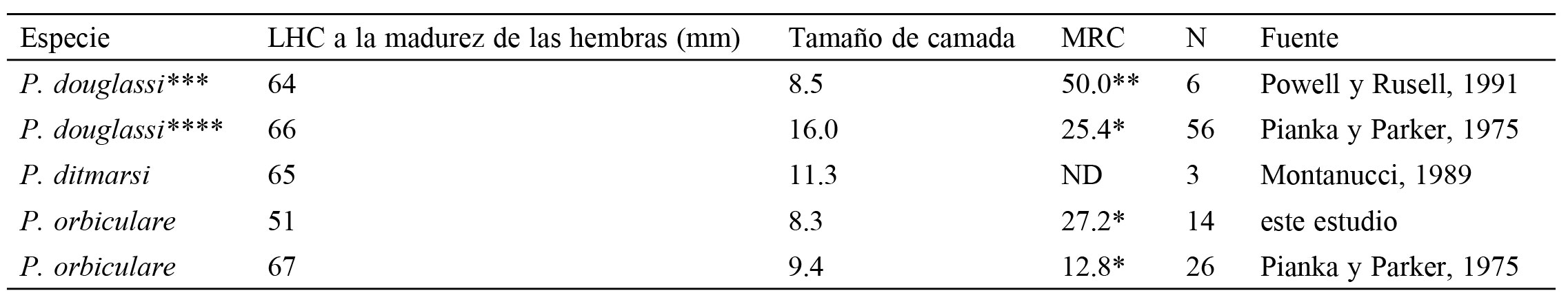
Las especies del género Phrynosoma si bien difieren en LHC, longitud de la cola, número, longitud y arreglo de los cuernos, además de su distribución geográfica (Luxbacher y Knouft, 2009), estas comparten un conjunto de características que las hacen altamente conspicuas entre las lagartijas: 1) dieta a base de hormigas, 2) una forma corporal muy similar (redonda y aplanada), 3) presencia de espinas, 4) coloración críptica, 5) estómago grande, 6) movimientos lentos, 7) modo de forrajeo pasivo, 8) cavidad abdominal amplia y 9) tamaño de camada o nidada grandes (Pianka y Parker, 1975; Pianka, 1981; Barbault, 1988; Sherbrooke, 2003). La MRC promedio (27.2) de las hembras de P. orbiculare del PESM se encuentra en el intervalo del clado Tapaja y del género Phrynosoma (tabla 2; tabla 11 en Pianka y Parker, 1975); se considera alta respecto a la MRC de especies de forrajeo amplio (promedio = 19.0, Vitt y Price, 1982) y es similar a la de especies de forrajeo pasivo (promedio = 25.9, Vitt y Price, 1982), cuya crípsis y cantidad baja de movimiento moldean su MRC (Vitt y Price, 1982). La MRC elevada de P. orbiculare, la especie más primitiva del género (Presh, 1969), forma parte del conjunto de características anatómicas y ecológicas que se encuentran interrelacionadas (Pianka, 1981; Sherbrooke, 2003) y que le han permitido sobrevivir y reproducirse exitosamente (Sherbrooke, 2003) desde su origen en el Mioceno (Bryson et al.,
2012).
Proporción sexual y características de las crías. Al igual que en P. douglasii (= hernandesi) (Powell y Russell, 1991), los resultados de este estudio indican que la proporción sexual al nacimiento de P. orbiculare fue de 1:1. Dicha proporción representa una estrategia evolutivamente estable con la mayor adecuación, donde si todos los individuos de la población la adoptan y en presencia de la selección natural, no puede ser sustituida por una estrategia alterna (Futuyma, 2006). Como sucede en otra especie de Phrynosoma (Powell y Russell, 1991), la proporción sexual de P. orbiculare en el PESM varía marcadamente entre camadas (de 0.14 a 0.7, proporción de machos en la población), pero la población mantiene dicha estrategia; el conocimiento de la proporción sexual en los individuos adultos de P. orbiculare del PESM ayudará a comprender mejor este aspecto demográfico.
La LHC y peso de las crías resultaron ser las características con menor variación; adicionalmente, ambas se mantienen estables independientemente del TC. Como se ha sugerido y registrado previamente (Rodríguez-Romero et al., 2002; Smith y Fretwell, 1974), la evidencia en P. orbiculare del PESM indica una optimización en el tamaño y peso de las crías; la ausencia de un trueque entre el tamaño y número de crías es un patrón común en poblaciones animales (Stearns, 1992). Cabe señalar que las crías de P. orbiculare en el PESM aparentemente son más grandes y más pesadas que otras crías de especies vivíparas del género Phrynosoma (Golberg, 1971; Powell y Russell, 1991), lo cual puede promover que la MRC se mantenga elevada a pesar de la disminución en el TC.
Agradecimientos
Referencias
Barbault, R. (1988). Body size, ecological constraints and the evolution of life-history strategies. En M. Hetch, B. Wallace y G. Prance (Eds.), Evolutive biology (pp. 261–286). Nueva York: Springer-Verlag.
Bastiaans, E., Méndez-de la Cruz, F., Rodríguez-Hernández, K., Flores-Aguirre, C. y Sinervo, B. (2013). Female reproductive investment in the mezquite lizard (Sceloporus grammicus) species complex (Squamata: Phrynosomatidae). Southwestern Naturalist, 58, 335–343.
Beltrán-Sánchez, E., Mendoza-Quijano, F. y Sherbrooke, W. C. (2005). Aspectos reproductivos del camaleón vivíparo Phrynosoma taurus (Sauria: Phrynosomatidae) de Zumpango del Río, Guerrero. Boletín de la Sociedad Herpetológica, 13, 37–41.
Bryson, R. W., García-Vázquez, U. O. y Riddle, B. R. (2012). Diversification in the Mexican horned lizard Phrynosoma orbiculare across a dynamic landscape. Molecular Phylogenetics and Evolution, 62, 87–96.
Castro-Franco, R., Bustos-Zagal, M. G. y Méndez-de la Cruz, F. R. (2011). Variation in parental investment and relative clutch mass of the spiny-tail iguana, Ctenosaura pectinata (Squamata: Iguanidae) in central México. Revista Mexicana de Biodiversidad, 82, 199–204.
Cuellar, O. (1984). Reproduction in a parthenogenetic lizard: with a discussion of optimal clutch size and a critique of the clutch weight/body weight ratio. American Midland Naturalist, 111, 242–258.
Davis, W. B. y Smith, H. M. (1953). Lizards and turtles of the Mexican state of Morelos. Herpetologica, 9, 100–108.
Dougthy, P. (1997). The effects of fixed clutch size on lizard life-histories: reproduction in the Australian Velvet Gecko, Oeodura lesueuri. Journal of Herpetology, 31, 266–272.
Du, W. G., Ji, X., Zhang, Y. P., Xu, X. F. y Shine, R. (2005). Identifying sources of variation in reproductive and life history traits among five populations of a Chinese lizard (Takydromus septentrionalis, Lacertidae). Biological Journal of Linnean Society, 85, 443–453.
Dunham, A. E., Grant, W. y Overall, K. L. (1989). Interfaces between biophysical and physiological ecology and the population ecology of terrestrial vertebrate ectotherms. Physiological Zoology, 62, 335–355.
Fitch, H. S. (1970). Reproductive cycles in lizards and snakes. Miscellaneous Publication, University of Kansas, Museum of Natural History, 52, 1–247.
Futuyma, D. J. (2006). Evolution. Massachusetts: Sinauer Associates.
Gobierno del Estado de México. (2013). Resumen ejecutivo del programa de conservación y manejo del Parque Estatal Sierra Morelos, Estado de México. Gaceta del Gobierno, 45, 1–32.
Goldberg, S. R. (1971). Reproduction in the short-horned lizard Phrynosoma douglassi in Arizona. Herpetologica, 27, 311–314.
Goldberg, S. R. (2011). Notes on reproduction of the Mexican-Plateau horned lizard, Phrynosoma orbiculare (Squamata: Phrynosomatidae). Bulletin of the Chicago Herpetological Society, 46, 61–62.
Hernández-Ibarra, X., Ramírez-Bautista, A. y Torres-Cervantes, R. (2000). Phrynosoma orbiculare orbiculare. Notes of life history. Herpetological Review, 31, 175–176.
Hernández-Navarrete, D., Suárez-Rodríguez, O., Suárez-Varón, G., Pérez-Pérez, A. y Hernández-Gallegos, O. (2016). Crecimiento corporal de Phrynosoma orbiculzare (Squamata: Phrynosomatidae). Memorias de la XIV Reunión Nacional de Herpetología, Nayarit, México. Tepic: Sociedad Herpetológica Mexicana A.C.
Hodges, W. L. (2002). Phrynosoma systematics, comparative reproductive ecology, and conservation of a Texas Native (Tesis doctoral). Austin: University of Texas.
Hodges, W. L. (2004). Evolution of viviparity in horned lizards (Phrynosoma). Testing the cold climate hypothesis. Journal of Evolutionary Biology, 17, 1230–1237.
Leaché, A. D. y McGuire, J. A. (2006). Phylogenetic relationships of horned lizards (Phrynosoma) based on nuclear and mitochondrial data: evidence for a misleading mitochondrial gene tree. Molecular Phylogenetics and Evolution, 39, 628–644.
Lemos-Espinal, J. A. y Smith, H. M. (2007). Anfibios y reptiles del estado de Coahuila, México / Amphibians and reptiles of the state of Coahuila, México. México, D.F.: UNAM-Conabio (Universidad Nacional Autónoma de México, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad).
Luxbacher, A. M. y Knouft, J. H. (2009). Assessing concurrent patterns of environmental niche and morphological evolution among species of horned lizards (Phrynosoma). Journal of Evolutionary Biology, 22, 1669–1678.
Martin, R. F. (1978). Clutch weight/total body weight ratios of lizards (Reptilia, Lacertilia, Iguanidae): preservative induced variation. Journal of Herpetology, 12, 248–251.
Marzluf, J. M., Bowman, R. y Donnelly, R. (2001). Avian ecology and conservation in an urbanized world. Seattle, Washington: Kluwer Academic Publishers.
Méndez-de la Cruz, F. R., Guillette, L. J., Jr. y Villagrán-Santa Cruz, M. (1993). Differential atresia of ovarian follicles and its effect on the clutch size of two populations of the viviparous lizard Sceloporus mucronatus. Functional Ecology, 7, 535–540.
Méndez-de la Cruz, F., Hernández-Gallegos, O. y Rodríguez-Romero, F. (2003). Phrynosoma orbiculare. Elaboración de fichas de 5 especies de lacertilios: PROY-NOM-059ECOL-2000. Departamento de Zoología, Instituto de Biología, Universidad Nacional Autónoma de México. Bases de datos SNIB-Conabio. Proyecto W033. México. D.F.
Montanucci, R. (1981). Habitat separation between Phrynosoma douglassi and P. orbiculare. (Lacertilia: Iguanidae) in Mexico. Copeia, 1981, 147–153.
Montanucci, R. (1989). Maintenance and propagation of horned lizards (Phrynosoma) in captivity. Bulletin Chicago Herpetological Society, 24, 229–238.
Muñoz-Nolasco, F. J. (2015). Esfuerzo reproductor y ecología térmica de dos especies vivíparas del género Plestiodon (Squamata: Scincidae) de México (Tesis). Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara. Zapopan, Jalisco.
Nieto-Montes de Oca, A., Arenas-Moreno, D., Beltrán-Sánchez, E. y Leaché, A. D. (2014). A new species of horned lizard (genus Phrynosoma) from Guerrero, México, with an updated multilocus phylogeny. Herpetologica, 70, 241–257.
Pianka, E. R. (1981). Resource acquisition and allocation among animals. En C. Townsend y P. Calow (Eds.), Physiological ecology: an evolutionary approach to resource use (pp. 300–314). Sunderland, MA: Sinauer Associates.
Pianka, E. R. (1986). Ecology and natural history of desert lizards. Analysis of the ecological niche and community structure. Princeton, New Jersey: Princeton University Press.
Pianka, E. y Parker, W. S. (1975). Ecology of horned lizards: a review with special reference to Phrynosoma platyrhinos. Copeia, 1975, 141–162.
Powell, L. y Russell, A. P. (1991). Parturition and clutch characteristics of short-horned lizards (Phrynosoma douglasii brevirostre) from Alberta. Canadian Journal of Zoology, 69, 2759–2764.
Presch, W. (1969). Evolutionary osteology and relationships of the horned lizard genus Phrynosoma (Family Iguanidae). Copeia, 1969, 250–275.
Ramírez-Bautista, A., Balderas-Valdivia, C. y Vitt, L. J. (2000). Reproductive ecology of the whiptail lizard Cnemidophorus lineatissimus (Squamata: Teiidae) in a tropical dry forest. Copeia, 2000, 712–722.
Ramírez-Bautista, A. y Vitt, L. J. (1997). Reproduction in the lizard Anolis nebulosus (Polychrotidae) from the Pacific coast of México. Herpetologica, 53, 423–431.
Rodríguez-Romero, F., Méndez-de la Cruz, F., García-Collazo, R. y Villagrán-Santa Cruz, M. (2002). Comparación del esfuerzo reproductor en dos especies hermanas del género Sceloporus (Sauria: Phrynosomatidae) con diferente modo reproductor. Acta Zoológica Mexicana, 85, 181–188.
Rodríguez-Romero, F., Méndez-de la Cruz, F. R. y López-González, L. (2005). Análisis comparado del esfuerzo reproductor en algunos lacertilios mexicanos de ambientes tropical y templado. Revista de la Sociedad Mexicana de Historia Natural, 2, 168–177.
Rodríguez-Romero, F., Smith, G. R., Cuellar, O. y Méndez-de la Cruz, F. R. (2004). Reproductive traits of a high elevation viviparous lizard Sceloporus bicanthalis (Lacertilia: Phrynosomatidae) from Mexico. Journal of Herpetology, 38, 438–443.
Rodríguez-Romero, F., Smith, G. R., Méndez-Sánchez, F., Hernández-Gallegos, O., Sánchez-Nava, P. y Méndez-de la Cruz, F. R. (2011). Demography of a semelparous, high-elevation population of Sceloporus bicanthalis (Lacertilia: Phrynosomatidae) from the Nevado de Toluca volcano, Mexico. The Southwestern Naturalist, 56, 71–77.
Rojas-Hernández, U., Salas-Mercado, G. y Hernández-Gallegos, O. (2016). Dieta de Phrynosoma orbiculare (Squamata: Phrynosomatidae). Memorias de la XIV Reunión Nacional de Herpetología, Nayarit, México. Tepic: Sociedad Herpetológica Mexicana A.C.
Santos-Bibiano, R. (2015). Aspectos reproductores de Phrynosoma sherbrookei (Sauria: Phrynosomatidae) en la localidad de Tenexatlajco, Chilapa de Alvarez, Guerrero, México (Tesis). Unidad Académica de Ciencias Químico-Biológicas, Universidad Autónoma de Guerrero, Chilpancingo, Guerrero, México.
Sherbrooke, W. C. (2003). Introduction to horned lizards of North America. California Natural History Guides No. 64. Berkeley: University of California Press.
Shine, R. (1992). Relative clutch mass and body shape in lizards and snakes: is reproductive investment constrained or optimized? Evolution, 46, 828–833.
Smith, C. C. y Fretwell, S. D. (1974). The optimal balance between size and number of offspring. The American Naturalist, 108, 499–506.
Stearns, S. C. (1976). Life-history tactics: a review of the ideas. Quarterly Review of Biology, 51, 3–47.
Stearns, S. C. (1992). The evolution of life histories. New York: Oxford University Press.
Suárez-Rodríguez, O. (2014). Tasa de adquisición de temperatura por medio de diferentes modos de transmisión y su relación con la forma del cuerpo en Phrynosoma orbiculare. (Tesis) de Maestría. Universidad Autónoma del Estado de México. Toluca, Estado de México, México.
Tinkle, D. W. (1967). The life and demography of the side-blotched lizard, Uta stansburiana. University of Michigan, Museum of Zoology. Miscelaneous Publications, 132, 21–182.
Tinkle, D. W. (1972). The dynamics of a Utah population of Sceloporus undulatus. Herpetologica, 28, 351–359.
Tinkle, D. W. y Handley, N. F. (1973). Reproductive effort and winter activity in the viviparous montane lizard Sceloporus jarrovi. Copeia, 1973, 272–276.
Tinkle, D. W., Wilbur, H. M. y Tilley, S. G. (1970). Evolutionary strategies in lizard reproduction. Evolution, 24, 55–74.
Vázquez-Díaz, J. y Quintero-Díaz, G. E. (2005). Anfibios y reptiles de Aguascalientes. México, D.F.: Conabio (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad).
Vitt, L. J. (1974). Reproductive effort and energy comparisons of adults, eggs, and neonates of Gerrhonotus coeruleus principis. Journal of Herpetology, 8, 165–168.
Vitt, L. J. y Congdon, J. D. (1978). Body shape, reproductive effort and relative clutch mass in lizards: resolution of a paradox. American Naturalist, 112, 595–608.
Vitt, L. J. y Price, H. J. (1982). Ecological and evolutionary determinants of relative clutch mass in lizards, Herpetologica, 38, 237–255.
Zamora-Ábrego, J. G., Zúñiga-Vega, J. J. y Nieto-Montes de Oca, A. (2007). Variation in reproductive traits within the lizard genus Xenosaurus. Journal of Herpetology, 41, 630–637.
Zamudio, K. R. y Parra-Olea, G. (2000). Reproductive mode and female reproductive cycle of two endemic Mexican horned lizards (Phrynosoma taurus and Phrynosoma braconnieri). Copeia, 2000, 222–229.
Zar, J. H. (2010). Biostatistical Analysis. Englewood Cliffs, Nueva Jersey: Prentice-Hall.


