Edna Rodríguez-Sánchez a, b, Laura J. Giraldo-Kalil a, Juan Núñez-Farfán a, *
a Universidad Nacional Autónoma de México, Instituto de Ecología, Departamento de Ecología Evolutiva, Circuito Exterior s/n anexo al Jardín Botánico, Ciudad Universitaria, Delegación Coyoacán, 04510 Mexico City, Mexico
b Universidad Nacional Autónoma de México, Instituto de Geología, Departamento de Paleontología, Circuito de Investigación, Ciudad Universitaria, Delegación Coyoacán, 04510 Mexico City, Mexico
*Corresponding author: farfan@unam.mx (J. Núñez-Farfán)
Received: 22 September 2021; accepted: 24 February 2022
Abstract
Besides recruitment and dispersal, fruits are key resources for the maintenance of insect communities. This study is focused on the insects inhabiting the fruits of 4 wild Lauraceae species. Although the trees of this family are important elements of tropical forests, their interaction with insects, especially in association with fruits, remains poorly studied in wild tree species. Our study aims to characterize the diversity of insects associated with fruits of Damburneya ambigens, Damburneya gentlei, Damburneya salicifolia, and Nectandra turbacensis, in the rainforest of Los Tuxtlas, Veracruz. We present an illustrated taxonomic list of species, annotated with a comprehensive review of the insects’ natural history and their interactions with Lauraceae species. We reared 54 insect species from approximately 6,500 fruits, some of which represent potential new species and records for Mexico. Insect species diversity was high and differed between Lauraceae species. The reared insects comprise a wide variety of distributional ranges, feeding types, and habitats. This research provides novel information about the interactions among insects and fruits of Lauraceae and the complexity of their trophic networks in tropical rainforests. Furthermore, it evidences the importance of wild fruits as resources for insect communities.
Keywords: Damburneya; Nectandra; Neotropical; Parasitoids; Saprophagous; Seed predators; Tropical rainforest
© 2022 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Diversidad de insectos asociados a los frutos de cuatro especies arbóreas de Lauraceae de la región de Los Tuxtlas, México: un listado taxonómico anotado e ilustrado
Resumen
Además del reclutamiento y dispersión de las plantas, los frutos son recursos esenciales para el mantenimiento de las comunidades de insectos. Este estudio se enfoca en insectos que habitan frutos de 4 especies silvestres de Lauraceae. Aunque los árboles de esta familia son elementos importantes en los bosques tropicales, sus interacciones con insectos, aún están poco estudiadas para especies silvestres. Nuestro objetivo fue caracterizar la diversidad de insectos asociados con frutos de Damburneya ambigens, Damburneya gentlei, Damburneya salicifolia y Nectandra turbacensis en la selva tropical de Los Tuxtlas. Presentamos un listado taxonómico ilustrado y anotado con una revisión exhaustiva de la historia natural de los insectos y su interacción con especies de Lauraceae. Criamos 54 especies de insectos de aproximadamente 6,500 frutos, mismas que podrían corresponder a nuevas especies y registros nuevos para México. La diversidad de especies de insectos fue alta y divergió entre especies arbóreas. Los insectos colectados presentan gran variedad de rangos de distribución, hábitos alimenticios y hábitats. Esta investigación aporta información novedosa sobre las interacciones entre insectos y frutos de Lauraceae y la complejidad de sus redes tróficas en selvas tropicales. Asimismo, evidencia la importancia de los frutos como recursos para las comunidades de insectos.
Palabras clave: Damburneya; Nectandra; Neotropical; Parasitoides; Saprófagos; Depredadores de semillas; Selva alta perennifolia
© 2022 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
By carrying the plants’ seeds and determining their dispersal, fruits play a central role in plant recruitment, population dynamics, and genetic structure. Fruits constitute a key resource in the maintenance of insect communities. Fruits and seeds are important food sources and at the same time provide shelter, mating, oviposition, and growth sites for insects. Hence, fruit and seed attributes can largely impact insect fitness. Moreover, the fruits frequently support very complex trophic interactions involving dispersers, insect seed predators (pre- and post-dispersal), pulp feeders, parasites, and even pathogens. Altogether, such interactions impact plant fitness and recruitment (Sallabanks & Courtney, 1992).
Our study focused on the insect fauna inhabiting the fruits of Lauraceae trees, which are drupes or one-seeded berries characterized by their fleshy mesocarp (Rohwer, 1993b). Lauraceae is a diverse family that comprises about 50 genera and 2,500-3,000 species widely distributed in tropical and subtropical ecosystems (Rohwer, 1993b). It is among the most common tree families in wet forests in the Neotropical region (Gentry, 1988; van der Werff & Richter, 1996). Despite its ecological importance, the family remains poorly studied (Lorea-Hernández, 2002; van der Werff & Richter, 1996). This is also evidenced in the little knowledge about reproductive biology and biotic interactions with insects like pollinators (Rohwer, 1993b), herbivores, frugivores and seed predators. Also, there is a clear lack of information about the ecology of insect-fruit interaction and its consequences on wild Lauraceae populations. Although there are several reports of insects associated with neotropical Lauraceae fruits, most of them are restricted to pest insects of commercial fruits such as avocado (Persea americana; Coria-Ávalos, 1999; Hoddle & Hoddle, 2012; Manrique et al., 2014; Muñiz, 1970).
We aimed to characterize the diversity of insects associated with fruits of 4 tree species of Lauraceae in the genera Damburneya Raf. and Nectandra Rol. ex Rottb., in the Biosphere Reserve of Los Tuxtlas. This is the natural protected area that hosts the largest number of Lauraceae species in Mexico (Lorea-Hernández, 2002). Unfortunately, more than 80% of the original rainforest cover has been removed by deforestation in this region, causing a dramatic landscape fragmentation (Dirzo et al., 2007), and this trend is expected to persist in the next years (Von Thaden et al., 2018). The situation is particularly worrying because most of the Mexican Lauraceae species are likely to be under some degree of extinction threat due to habitat loss, and the difficulty of most of them to establish under disturbance conditions or secondary vegetation (Lorea-Hernández, 2002).
To our knowledge, very few works have recorded the insects associated with Lauraceae fruits of the genera Damburneya and Nectandra, especially regarding insect species diversity (Andrade-Lara, 1989; Atkinson & Equihua-Martínez, 1985b; Downey, 2018; Link & Link, 2008; Riccardi & Rodríguez-Sánchez, 2021; Rodríguez, 2009; Sánchez-Garduño, 1995). Considering the accelerated rainforest loss in Los Tuxtlas region, and the scarcity of information of the impact of biotic interactions on an ecologically important group such as Lauraceae, there is an increasing need to study the diversity of insects associated with these plants. Here, we report the surprising diversity of insects associated with fruits of D. ambigens, D. gentlei, D. salicifolia and N. turbacensis, at Los Tuxtlas region in an illustrated taxonomic list. We annotated the list with a detailed bibliographic review to describe and discuss aspects of natural history.
Materials and methods
This research is focused on wild Nectandra and Damburneya species; the latter genus was recently reinstated and separated from Nectandra by Trofimov et al. (2016). Individuals of these genera present numerous small flowers and low fruiting in relation to the flower production (1 per 1,000 flowers; Rohwer, 1993b); however, thousands of fruits are produced per tree. Fruiting can be annual or biannual (Ibarra-Manríquez & Sinaca-Colín, 1995), and fruit maturation takes several weeks to months (Rohwer, 1993b). Fruits are one-seeded berries, green when immature and blackish purple at maturity. Each fruit has a characteristic red cupule at the base and contains one recalcitrant seed that occupies most of the fruit volume when mature and germinates soon after the fruit falls from the tree (Chávez-Pesqueira & Núñez-Farfán, 2016; Rohwer, 1993b). The pulp has a large amount of lipids (Stiles, 1993) and the fruits are consumed by insects, monkeys (Dirzo et al., 1997), and birds, which are the main dispersers (Rohwer, 1993a). Pre-dispersal seed predation by curculionid beetles and other insects is common and can harm a large proportion of young fruits (Rohwer, 1993b). Moreover, rodents can act both as post-dispersal seed predators or secondary dispersers (Dirzo et al., 1997; Rohwer, 1993a).
Here we focused on the fruits of D. ambigens, D. gentlei, D. salicifolia, and N. turbacensis, which are relatively common trees in the tropical rainforest of the study area and are locally employed for timber extraction (Ibarra-Manríquez et al., 1997). The fruits of these species vary in size (from 1 to 2 cm), and form (from elliptic to rounded, Rohwer, 1993b). A detailed description of tree species, fruit shape, and size is shown in Supplementary material table S1 and figure 1.
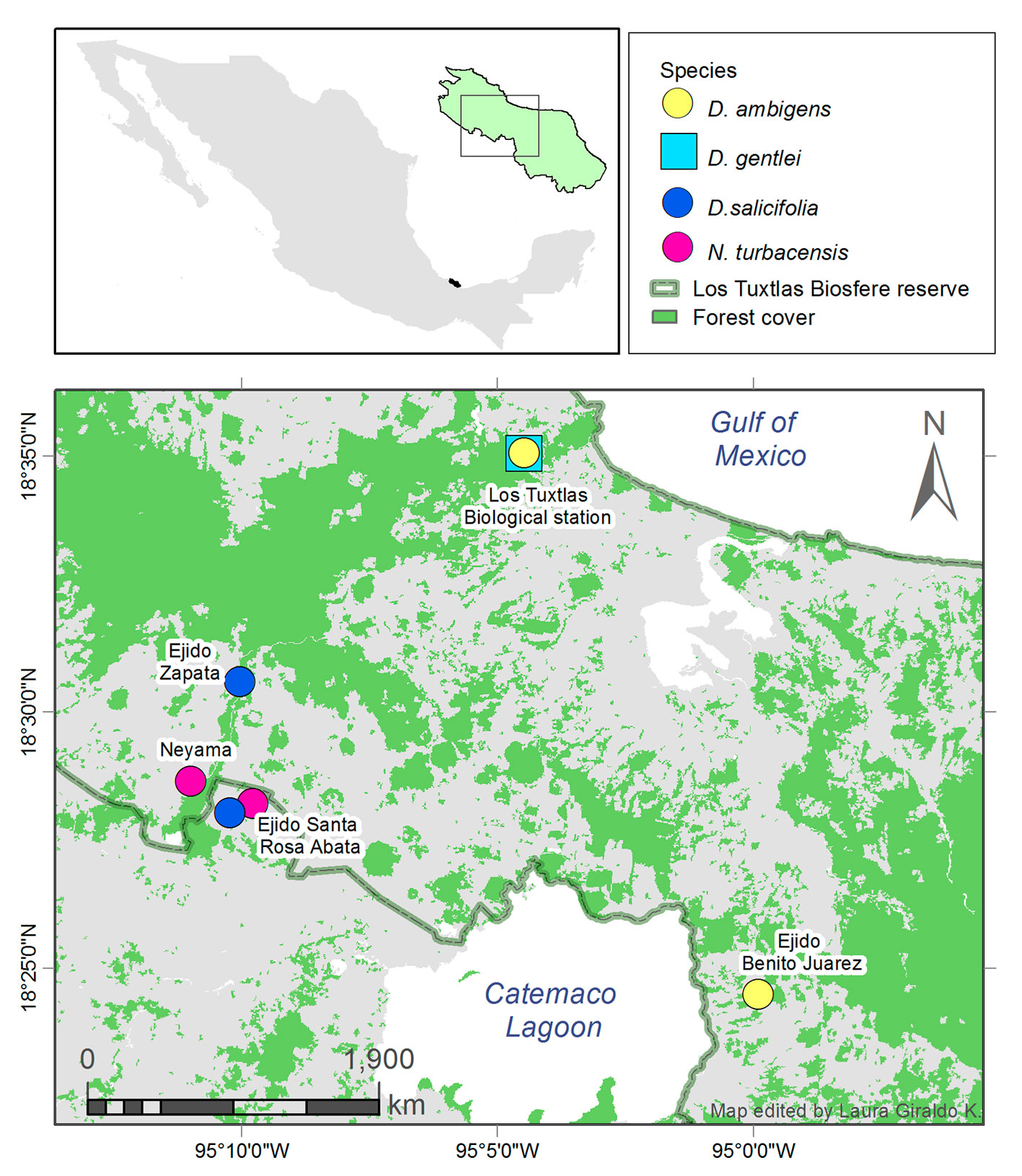
This study was conducted in the tropical rainforest of Los Tuxtlas Biosphere Reserve, Veracruz, Mexico, in the mountain range of the San Martín volcano and nearby areas. The site has an elevation gradient from sea level up to 1,700 m asl and a humid tropical climate. Mean annual temperature ranges from 24 °C to 27 °C and mean annual precipitation ranges from 4,000 to 6,000 mm according to elevation (Gutíerrez-García & Ricker, 2011). The rainfall is concentrated in the rainy season, between June and February.
Fruits from D. ambigens, D. gentlei, D. salicifolia, and N. turbacensis trees were collected between October 2016 and 2018. The sampling was performed in different populations according to the availability of fruits and the distribution of the tree species. Sampling sites were located within the Biosphere Reserve of Los Tuxtlas and contiguous zones, including forest fragments, forest edges, and nearby principal rural roads (Fig. 2; Table 1). We sampled around 10 trees per species and population, except for D. gentlei, because there were very few fruiting trees of this species (Table 1).
Approximately 6,500 fruits were collected, and the sample size was at least 50 fruits per tree. As the height of D. ambigens and D. gentlei make the fruits inaccessible, the fruits of these species were collected from the ground. The fruits of D. salicifolia and N. turbacensis were collected directly from the branches.
The fruits were placed in plastic containers, covered with a fine mesh, and deposited for 2 months in a growth chamber at 27/20 °C with a 12-hr light/dark cycle. The emerged adult insects were recorded and collected for 2 months, and once the rearing period ended, we opened the fruits to collect the remaining individuals. The adult insects were preserved and determined with the support of specialists and the use of taxonomic keys. Representative specimens were mounted, photographed, and deposited at the Colección Nacional de Insectos (CNIN) and Colección Entomológica at Estación Biológica de Los Tuxtlas (Instituto de Biología, UNAM). Collection numbers are shown in Supplementary material table S2.
Insect species and abundance are listed in alphabetic order. Furthermore, Hill numbers were used to calculate diversity. Insect-species richness, Hill diversity, and sample coverage were calculated for each Lauraceae species to characterize the diversity of insects associated with fruits and the sample completeness with the R platform (R Core Team, 2021) using the “entropart” package (Marcon & Hérault, 2015).
The Hill numbers integrate the richness and abundance of species and show the effective species within communities using q values that represent abundance sensitivity. We calculated diversity using q = 1 which proportionally weights the species by their abundance (Shannon diversity equivalence), and q = 2 which proportionally weights the species by their squared abundance (Simpson diversity equivalence). These values range from 0 to species richness; if the later value is reached, it would indicate that the community is equally represented by the individuals of each species (Chao et al., 2020; Roswell et al., 2021). Insect abundance was analyzed for each tree species. Moreover, sample coverage was calculated by weighting species by their abundances using Zhang and Huang’s (2007) method, to show the proportion of individuals within the community belonging to the species of the current sample. Coverage values range from 0 to 1, where 1 indicates absolute completeness (Chao et al., 2020; Roswell et al., 2021). On the other hand, as there were very few D. gentlei trees (n = 2), this species was not included in diversity metrics calculation.
Table 1
Collection data of Lauraceae fruits sampled at Los Tuxtlas region.
| Lauraceae species | Population | Latitude | Longitude | Altitude
(m asl) |
Collection date | No. trees | No. Fruits |
| D. ambigens | EBT Tux. | 18.586712 | -95.077173 | 187 | Sep, 2018 | 9 | 936 |
| B. Juárez | 18.404641 | -94.997894 | 624 | Oct, 2018 | 10 | 714 | |
| D. gentlei | EBT Tux. | 18.583465 | -95.075192 | 175 | Sep, 2018 | 2 | 133 |
| D. salicifolia | Zapata | 18.448984 | -95.049031 | 405.71 | Oct, 2016 | 16 | ca. 800 |
| Sta. Rosa | 18.467034 | -95.170503 | 551.1 | Ago, 2017 | 12 | ca. 600 | |
| Neyama | 18.47424 | -95.182429 | 575.12 | Ago, 2018 | 10 | 980 | |
| N. turbacensis | Sta. Rosa | 18.46482 | -95.171356 | 550.86 | Ago, 2017 | 16 | ca. 800 |
| Neyama | 18.477074 | -95.183467 | 579.84 | Ago, 2017 | 11 | ca. 550 | |
| Neyama | 18.47424 | -95.182429 | 575.12 | Ago, 2018 | 10 | 944 |
The taxonomic list includes pictures of the species and a comprehensive bibliographic review including geographic, taxonomic, and ecological information of each registered insect taxon. The ecological information provided is focused on the insect-plant association, especially with Lauraceae plants. Furthermore, we include our analysis and observations on natural history of the collected species. The bibliographic search was performed between January 2018 and June 2021 using Scholar Google and Scopus (Supplementary material table S3).
Results
We reared a total of 54 insect species, including Coleoptera (21 spp.), Diptera (11 spp.), Hymenoptera (16 spp.), Lepidoptera (5 spp.), and Thysanoptera (1 sp.) (collection number on Supplementary material Table 2). We were not able to determine damaged or incomplete individuals to species level, and sometimes neither to genera. The insects showed a differential occurrence and abundance between the tree species. For example, Neosilba sp. (Diptera: Lonchaeidae) is the only species shared between the 4 Lauraceae species. Furthermore, several species occurred just 1 or 2 times, while others such as Bracon sp. 1 (Hymenoptera: Braconidae), Heilipus albomaculatus, Pagiocerus frontalis (Coleoptera: Curculionidae), Neosilba sp. (Diptera: Lonchaeidae), and Stenoma catenifer (Lepidoptera: Oecophoridae) were very abundant. Pagiocerus frontalis is by far the most abundant species, although it was absent in D. gentlei (Table 2). It is worth stressing that, since fruits were collected directly from the branches, all the insects reared from D. salicifolia and N. turbacensis fruits constitute pre-dispersal associations.
To characterize the richness and diversity of insects associated with the fruits of each Lauraceae species, we calculated sample coverage and several diversity metrics (Table 3). The sample coverage was high (> 90%; Table 3), indicating that our sampling provides a good representation of the species within the community of insects associated with Nectandra and Damburneya fruits. On the other hand, N. turbacensis had the highest insect richness with 37 recorded species, as well as the highest diversity (q = 1) and the lowest dominance (q = 2), followed by D. ambigens and D. salicifolia, respectively. Nectandra turbacensis was 70-80% more diverse than the other tree species. Furthermore, the 3 species showed an intermediate to high dominance with very low values of effective species for q = 2 (Table 3). In contrast, D. gentlei had only 2 associated insect species (Heilipus albomacultus and Neosilba sp.), likely because of the low tree sampling.
Below, we provide an illustrated and annotated taxonomic list. It compiles and describes the relevant published information and our observations on the natural history of the collected insect species in D. ambigens, D. gentlei, D. salicifolia, and N. turbacensis fruits. We emphasized the existing information regarding insect association with Lauraceae species. A general synthesis of this information is provided in Supplementary material table S2, including novelties on insect-plant associations and new records for Mexico.

Coleoptera
Curculionidae
Anchonus Schönherr, 1825
The genus contains more than 100 registered species distributed in America (Zimmerman, 1964). There are 15 registered neotropical species (Thomas & O’Brien, 1999), but only 2 are recorded in Mexico (A. abnormis and A. fraterculus; Morrone, 1999). However, according to Morrone (2014) Anchonus (Fig. 3A) is one of the Curculionidae genera that contains a great part of endemic species of Mexico.
Anchonus species are typically recorded as associated with wood (Arnett et al., 2002; Thomas & O’Brien, 1999), but they have also been reported as flightless ground-dweller beetles found in driftwood, decaying wood, and ground vegetation (Zimmerman, 1964). This contrasts with our report in Damburneya ambigens fruits; Anchonus sp. is probably a seed predator or saprophagous, but unfortunately, we do not have precise observations of its feeding habits.
Coccotrypes cyperi Beeson, 1929
Like other species of the genus, C. cyperi is known for its invasive potential (Haack & Rabaglia, 2013; Fig. 4A). This Asian species is widely distributed in the paleotropics and it has been described as an adventive species in the neotropics (Atkinson & Peck, 1994). However, there are reports of this species in primary old-growth tropical forest in Central America (Kirkendall & Ødegaard, 2007).
Table 2.
List of insects associated with Damburneya and Nectandra fruits. Taxa are presented in alphabetical order. The abundance of insect species is provided according to Lauraceae host tree. In the case of Bracon sp. 4, a single individual was found but the plant from which it emerged is unknown, it might be Damburneya salicifolia or Nectandra turbacensis. Abbreviations: AW = associated with wood, Fr = frugivorous, Fu = fungivorous, Ga = gall inducers or inquilines, Pa = parasite, Pd = parasitoid, Ph = phytophagous, Pl = pollinators, Po = polyphagous, Pr = predator, Sa = saprophagous, Sc = scavenger, SP = seed predator, ? = uncertain data.
| Insect | D. ambigens | D. gentlei | D. salicifolia | N. turbacensis | Feeding types |
| Coleoptera | |||||
| Curculionidae | |||||
| Anchonus sp. | 1 | AW, Sa?, SP? | |||
| Coccotrypes cyperi | 1 | AW, SP | |||
| Conotrachelus serpentinus | 1 | SP | |||
| Heilipus albomaculatus | 260 | 3 | SP | ||
| Heilipus draco | 2 | SP | |||
| Heilipus guttiger | 5 | 2 | SP | ||
| Hypothenemus interstitialis | 3 | Po | |||
| Pagiocerus frontalis | 7,764 | 5,121 | 209 | SP | |
| Laemophloeidae | |||||
| Cryptolestes sp. | 1 | AW, Fr, Pr | |||
| Nitidulidae | |||||
| Carpophilus maculatus | 2 | 2 | Pl, Sa | ||
| Carpophilus sp. 1 | 1 | 1 | Pl, Sa | ||
| Carpophilus sp. 2 | 1 | Pl, Sa | |||
| Stelidota sp. 1 | 1 | Sa | |||
| Stelidota sp. 2 | 1 | Sa | |||
| Stelidota sp. 3 | 2 | Sa | |||
| Stelidota sp. 4 | 1 | Sa | |||
| Stelidota sp. 5 | 1 | Sa | |||
| Stelidota sp. 6 | 1 | Sa | |||
| Stelidota sp. 7 | 1 | Sa | |||
| Stelidota sp. 8 | 6 | Sa | |||
| Staphylinidae | |||||
| Aleocharinae sp. | 1 | Ph, Pl Pr, Pd | |||
| Diptera | |||||
| Chloropidae | |||||
| Apallates sp. | 1 | Sa | |||
| Chaetochlorops inquilinus | 1 | Sa | |||
| Conioscinella sp. | 1 | 7 | Sa, Pr, Ph | ||
| Drosophilidae | |||||
| sp. 1 | 1 | Fr | |||
| Lauxaniidae | |||||
| sp. 1 | 1 | Pl, Sa | |||
| Lonchaeidae | |||||
| Neosilba sp. | 22 | 4 | 55 | 16 | Fr, Pl |
| Table 2. Continued | |||||
| Insect | D. ambigens | D. gentlei | D. salicifolia | N. turbacensis | Feeding types |
| Muscidae | |||||
| Atherigona orientalis | 2 | Ph, Po, Sa | |||
| Phoridae | |||||
| sp. 1 | 1 | Pa, Sa | |||
| sp. 2 | 1 | Pa, Sa | |||
| Stratiomyidae | |||||
| Ptecticus sackenii | 4 | Fr, Sa | |||
| Tachinidae | |||||
| sp. 1 | 6 | Pa, Pd | |||
| Hymenoptera | |||||
| Braconidae | |||||
| Apanteles sp. 1 | 1 | 1 | Pd | ||
| Apanteles sp. 2 | 2 | Pd | |||
| Apanteles sp. 3 | 1 | Pd | |||
| Apanteles sp. 4 | 1 | Pd | |||
| Aridelus sp. | 1 | Pd | |||
| Bracon sp. 1 | 1 | 16 | 2 | Pd, Ph | |
| Bracon sp. 2 | 1 | Pd, Ph | |||
| Bracon sp. 3 | 1 | Pd, Ph | |||
| Bracon sp. 4 | ? | ? | Pd, Ph | ||
| Eulophidae | |||||
| Galeopsomyia fausta | 7 | Pd | |||
| Eurytomidae | |||||
| Eurytoma sp. | 6 | Pd, Ph, Pr | |||
| Sycophila sp. | 3 | Ga, Pd, Ph, SP | |||
| Figitidae | |||||
| Aganaspis pelleranoi | 1 | Pd | |||
| Formicidae | |||||
| Mycocepurus goeldii | 1 | Fu | |||
| Pheidole sp. | 1 | Pr, Sc, SP | |||
| Ichneumonidae | |||||
| sp. 1 | 1 | Pd | |||
| Lepidoptera | |||||
| sp. 1 | 5 | 8 | – | ||
| sp. 2 | 1 | – | |||
| sp. 3 | 1 | – | |||
| sp. 4 | 20 | – | |||
| Oecophoridae | |||||
| Stenoma catenifer | 5 | 4 | 1 | SP | |
| Thysanoptera | |||||
| Phlaeothripidae | |||||
| sp. 1 | 1 | Fu, Ga, Ph, Pr |
Table 3
Diversity metrics for insect species associated with Nectandra and Damburneya fruits. The data are presented according to tree species. Values of richness, diversity q = 1 (diversity), diversity q = 2 (dominance), and sample coverage are provided.
| Lauraceae | Richness | Diversity
q = 1 |
Diversity
q = 2 |
Sample coverage |
| D. ambigens | 26 | 3.53 | 2.55 | 0.998 |
| D. salicifolia | 13 | 2.61 | 1.63 | 0.999 |
| N. turbacensis | 37 | 12.66 | 7.02 | 0.943 |
Coccotrypes cyperi breeds on large-seeded fruits reports comprise large seeds of fallen fruits of several palms and woody plants including the Lauraceae Persea americana, and other wild and commercial species such as Cynometra hemitobophylla, Mangifera indica, Mammea americana, Orbignya oleifera, Phytelephas macrocarpa, Theobroma cacao (Wood, 1982), Terminalia myriocarpa, Swietenia macrophylla (Wood & Bright, 1992), Spondias mombin (Atkinson & Equihua-Martínez, 1985b), among many others. Detailed list of records can be found in Atkinson (2020). Nevertheless, C. cyperi probably can breed in all parts of the trees, like twigs, wood (Haack & Rabaglia, 2013), and petioles (Kirkendall et al., 2015). The species of Coccotrypes are haplodiploid (Kirkendall & Ødegaard, 2007) and reproduce by inbred mating: females reproduce with their siblings and emerge to colonize a new plant structure (Wood, 1982).
In Mexico, C. cyperi has been reported in cacao crops (Equihua-Martínez, 1992; Pérez-De La Cruz et al., 2009), and in tropical rainforests (Pérez-De la Cruz et al., 2015, 2016). The tropical rainforest at Los Tuxtlas is very likely an appropriate environment for C. cyperi. Thus, its occurrence in the area is not surprising. The specimen found in D. ambigens was very likely predating on seeds. To our knowledge, this is the first record of C. cyperi in a wild Lauraceae tree species, and specifically in Damburneya since other known records from the family are restricted to P. americana.
Conotrachelus serpentinus (Klug, 1829)
Conotrachelus serpentinus (Fig. 3B) has been recorded from Florida to Colombia, preying seeds of various Persea species like P. americana, P. gratissima, and other native Lauraceae species (Coria-Ávalos, 1999; Muñiz, 1970; Whitehead, 1979). Nevertheless, it is not considered a pest of economic importance (Whitehead, 1979). Individuals of Conotrachelus genus had already been registered in Brazil preying seeds of Nectandra megapotamica (Link & Link, 2008) and in Los Tuxtlas region predating seeds of other plant families like Chrysobalanaceae, Fabaceae, Malvaceae, Meliaceae and Polygonaceae (Andrade-Lara, 1989).
Heilipus albomaculatus Champion, 1902
Heilipus albomaculatus (Fig. 3C, D) is distributed in Mexico, Belize, and Guatemala (Wibmer & O’Brien, 1986). Larvae have been recorded attacking the base of Persea americana trunks (Cervantes & Huacuja, 2017). Moreover, Sánchez-Garduño (1995) reported H. albomaculatus as the main pre-dispersal seed predator of Damburneya ambigens seeds in Los Tuxtlas region. As Sánchez-Garduño (1995), we generally found 1 individual per seed; however, we observed 2 individuals per seed once in approximately 260 fruits surveyed. The larva feeds and pupates on the seed, and later insect emerges as an adult. Predation incidence is ca. 19%; nevertheless, the larvae do not necessarily consume the whole seed, and in some cases, predation can facilitate the germination (Sánchez-Garduño, 1995). In contrast, we registered an incidence of H. albomaculatus of 16% and 2% in D. ambigens and D. gentlei seeds, respectively.
Heilipus draco Fabricius, 1801
The distribution of H. draco (Fig. 3E, F) includes Mexico, Guatemala, El Salvador, Nicaragua, Costa Rica, Panama, Cuba, Trinidad and Tobago, Colombia Venezuela, Brazil, Peru, Bolivia, and Paraguay (Díaz-Grisales et al., 2021; Janzen, 1987; Morrone, 1999; Wibmer & O’Brien, 1986). It has been recorded in deciduous and semi-deciduous rain forest, associated with fruits of the following Lauraceae species: Beilschmiedia pendula, Ocotea cernua, O. oblonga, O. puberula, Nectandra cissiflora, N. lineata (Downey, 2018), and O. veraguensis (Janzen, 1987). Downey (2018) reported that H. draco damages between 0.05 and 3.4% of wild Lauraceae seeds in Barro Colorado island in Panama.
Heilipus guttiger Champion, 1902
Heilipus guttiger (Fig. 3 G, H) has been recorded in Mexico, Guatemala, Panama, and Colombia (Díaz-Grisales et al., 2021; Wibmer & O’Brien, 1986). There is limited information about this species, and to our knowledge, this is the first report of the host plants to which it is associated (D. salicifolia and N. turbacensis). We observed 1 larva per seed, the larvae feed and pupate on the seeds, and later insects emerges as adults. In some cases, we found live larvae after 2 months of rearing when we opened the fruits; we put them back to the growth chamber to let them complete their life cycle.
Hypothenemus interstitialis Hopkins, 1915
Hypothenemus interstitialis (Fig. 4B) has a circumtropical distribution (Noguera-Martínez & Atkinson, 1990), and has been reported in several countries of North, Central, and South America as well as in the Caribbean Antilles, inhabiting dry and humid ecosystems in lowland areas (Atkinson, 2020). This myelophagous and polyphagous beetle has been reported to feed on 33 plant families and more than 90 species, including 7 Lauraceae species of Nectandra, Ocotea, Persea, and Sassafras (Atkinson, 2020). The insects of the genera Hypothenemus reproduce by endogamic polygyny and arrhenotokic parthenogenesis, thus, the flightless males are likely haploid (Wood, 1982). Despite the females make galleries to oviposit around a dozen eggs or less, the life cycle is short, and several generations can coexist within the same material leading to large populations (Wood, 1982).
There are several previous reports in Mexico, including unidentified Lauraceae and other plant species in Jalisco (Burgos-Solorio & Equihua-Martínez, 2007). Gerónimo-Torres et al. (2015) reported very few individuals in the mangrove ecosystem in Tabasco. Similarly, Falcón-Brindis et al. (2018) reported few individuals of H. interstitialis in tropical rainforests of Tabasco; however, its abundance increased in disturbed areas such as grasslands and forest crops that are likely advantageous for generalist insects (Falcón-Brindis et al., 2018). This beetle has been reported in Campeche in secondary tropical semideciduous forests in several plant families, being avocado the only Lauraceae (Estrada & Atkinson, 1988). Furthermore, H. interstitialis has been reported in an unidentified Nectandra in Veracruz (Atkinson & Equihua-Martínez, 1985b); thus, despite the very few reports, its occurrence feeding on D. ambigens fruits at Los Tuxtlas is not surprising.
Pagiocerus frontalis Fabricius, 1801
The pre-dispersal seed predator P. frontalis (Fig. 4C) occurs from the USA to South America (Atkinson & Equihua-Martínez, 1985a; Wood, 1982). This species feeds on large hard seeds and the fruit tissue that covers them (spermatophagy; Kirkendall et al., 2015). Particularly, it is associated with many Lauraceae genera (Atkinson & Peck, 1994; Kirkendall et al., 2015), including Damburneya (Sánchez-Garduño, 1995), Beilschmiedia, Nectandra (Downey, 2018), Ocotea (Downey, 2018; Wood, 1982), and Persea (Atkinson & Equihua-Martínez, 1985a; Atkinson & Peck, 1994; Wood, 1982). Nevertheless, this insect has been reported in other plant families like Annonaceae, Boraginaceae (Eidt-Wendt & Schulz, 1990), Poaceae (Castro-Ccoscco & Mejía-Espinoza, 2011; Okello et al., 1996), and Rubiaceae (Kirkendall et al., 2015). Pagiocerus frontalis can become a dominant predator (Downey, 2018), and is a pest of corn crops. Furthermore, it can settle down and harm a wide variety of substrates such as fruits, coffee grains, and even plastic (Castro-Ccoscco & Mejía-Espinoza, 2011; Eidt-Wendt & Schulz, 1990; Kirkendall et al., 2015; Wood, 1982). This beetle is widely distributed in Mexico, and it has been collected on Lauraceae trees of Persea and Nectandra (Atkinson & Equihua-Martínez, 1985a). Atkinson (2020) provided a detailed list of records for this species.
In a study in the Los Tuxtlas region, Sánchez-Garduño (1995) registered dozens of these beetles per seed of D. ambigens. The larvae preyed and formed galleries in 11% of the seeds, emerging later from the fruits as adults. Nevertheless, as H. albomacultus (see above), P. frontalis did not always consume the whole seed; and in some cases, predation facilitates seed germination.
Pagiocerus frontalis is known to attack exposed seeds of fallen fruits (Kirkendall et al., 2015); however, we reared individuals from fruits collected from the ground and from tree branches. Besides D. ambigens, we also found this beetle in D. salicifolia and N. turbacensis seeds. Additionally, it was the most abundant insect species and the most harmful to the seeds.


Laemophloeidae
Cryptolestes Ganglbauer, 1899
Cryptolestes (Fig. 4D) is hard to distinguish from other related genera because of its problematic taxonomy. It comprises several secondary pest species associated with stored grains and other products. Six species are reported in America and have a nearly cosmopolitan distribution; in contrast, there are approximately 13 non-economic species in America, all of them barely known (Thomas, 1988). They usually occur under the bark of hardwood logs and are likely fungivores; however, some species can feed on scale insects (Thomas, 1988). On the other hand, some species have been found in stored grains in Mexico. For example, C. pusillus has been reported on stored maize (Callejas-Chavero et al., 2019; Córdova et al., 2011). Moreover, C. ferrugineus has been found in stored sorghum, wheat, barley, and pinto beans; also, other unidentified species was reported to feed on stored chili (Córdova et al., 2011; Wong et al., 1992). There are also registers of other poorly known species in a few taxonomic studies (Thomas, 1988, 2002). To our knowledge there is no information about the association of these beetles with wild plant species; hence, this study could be the first report of this kind. Furthermore, Loschiavo and Sinha (1966) reported the fungi of stored seeds as a source of food for C. ferrugineus, thus we cannot rule out the fungi as the food source of Cryptolestes found in N. turbacensis seeds.
Nitidulidae
Carpophilus Stephens, 1830
Carpophilus (Fig. 5) contains over 200 species (Brown et al., 2012) and at least 33 occur in Mexico (Blackwelder, 1945; Williams et al., 1983). This genus has many taxonomic problems and probably it is not a monophyletic group (Brown et al., 2012). The distribution of Carpophilus is almost cosmopolitan (Dobson, 1954), although most species are from tropical and subtropical regions (Prado, 1987; Williams et al., 1983). Some Carpophillus are pollinators of Annonaceae (Brown et al., 2012; Prado, 1987) and Calycanthaceae flowers (Brown et al., 2012; Williams et al., 2008), but most species feed on a variety of fruits (James & Vogele, 2000; James et al., 1995; Williams et al., 1983), grains and stored products (Brown et al., 2012; James et al., 1995; Prado, 1987; Rodríguez-Del Bosque et al., 1998). The individuals quickly break down dried fruits and drupes (James et al., 1995; James & Vogele, 2000; Prado, 1987; Rodríguez-Del Bosque et al., 1998; Williams et al., 1983). According to Williams et al. (1983) this is the most economically important genus of Nitidulidae since approximately 16 species are considered pests (Dobson, 1954), and in some cases they are vector of plant pathogenic fungi (Rodríguez-Del Bosque et al., 1998). There are few parasitoid wasps known to attack Carpophilus larvae, including species of Anisopteromalus, Zeteticontus, and Pseudisobrachium genera (Williams et al., 1984).
Regarding associations with Lauraceae, Link and Link (2008) reported low infestation of 3 unidentified species of Carpophilus on Nectadra megapotamica fruits in Brazil. We reared 2 unidentified species of Carpophilus from D. salicifolia and N. turbacensis fruits (Fig. 5B, C). They show differences in body size and punctuation of mesosternal disc, prosternum, propleuron and elytra. Considering the known feeding habits, the reared species were likely consuming the pulp of the fruits or decaying rests of the seeds.
Carpophilus maculatus Murray, 1864
Carpophilus maculatus (Fig. 5A) has a wide geographic distribution and is common in crops in the tropical zones of South America (Brown et al., 2012). It has also been registered in USA (Ohio; Williams et al., 1992), Asia, and Australia (Brown et al., 2012). From our knowledge, this is the first record for Mexico. Carpophilus maculatus is a stored products pest (Dobson, 1954), but it also pollinates Annonaceae species and associates with fruits of many plant families like Anacardiaceae, Apocynaceae, Araceae, Arecoidae, Bromeliaceae, Caricaceae, Convolvulaceae, Curcubitaceae, Dioscoreaceae, Fabaceae, Malvaceae, Moraceae, Musaceae, Myrtaceae, Oxalidaceae, Pandanaceae, Piperaceae, Poaceae, Rubiaceae, Rutaceae, Sapotaceae, and Solanaceae (Brown, 2009). Nevertheless, there are no reports of its association of C. maculatus with Lauraceae fruits until now.
Stelidota Erichson, 1843
The genus Stelidota (Fig. 6) is distributed worldwide (except in Africa), especially in tropical regions (Ford, 1996). Stelidota comprises about 50 species worldwide, 30 species occur in the Neotropical region, and 8 has been registered in Mexico: S. alternans, S. championi, S. ferruginea, S. germinata, S. octomaculata, S. rubripes, S. solitaria, and S. strigosa (Blackwelder, 1945; Weiss & Williams, 1980; Williams et al., 1989). Some parasitoid wasps have been reported attacking Stelidota adults (Microctonus nitiduldis), and larvae (Serphus obsoletus and various species of Proctotrupidae; Weiss & Williams, 1980; Williams et al., 1984)
Like most nitidulids, Stelidota is saprophagous of fruits and flowers of Araceae, Bromeliaceae, Calycanthaceae, Curcubitaceae, Ebenaceae, Ericaceae, Fagaceae, Malvaceae, Moraceae, Poaceae, Rosaceae, Rutaceae, Sapindaceae, Solanaceae, and others plant families (Weber & Connell, 1975; Williams et al., 2008). The Stelidota beetles reared from D. ambigens are likely the first records of the association of this genus with Lauraceae. Surprisingly, we registered 8 species that mainly show differences in mandible shape, elytra form and color pattern, and body size. Considering the known feeding habits, the reared species were likely consuming the pulp of the fruits or decaying rests of the seeds.
Staphylinidae
Aleocharinae
Aleocharinae (Fig. 4E) is the most diverse subfamily of Staphylinidae with approximately 12,000 species; its distribution is cosmopolitan, and the species of this group are generally abundant (Navarrete-Heredia et al., 2002). It is the third richest subfamily in Mexico comprising 234 species recorded; however, it remains poorly studied (Navarrete-Heredia & Newton, 2014).
Aleocharines have many feeding habits, and could be phytophagous, pollinators, inquilines, generalist predators, and parasitoids (Navarrete-Heredia et al., 2002; Sayers et al., 2019). However, they are characterized by their habitat specialization: Charoxus is the only genus that has been reported to be associated with fruits (Ficus spp.), and Aleochara the only parasitoid genus, in this case of Diptera pupae (Navarrete-Heredia et al., 2002). From our knowledge, this is the first reported association of Aleocharinae with Lauraceae plants; however, we did not observe its feeding type (Fig. 4E). Regarding the Staphylinidae family, there are only 2 reported Lauraceae associations with Lindera spp. flowers (Dupont & Kato, 1999; Tokumoto et al., 2019) and P. americana wood (Peña et al., 2015).

Diptera
Chloropidae
Apallates Sabrosky, 1980
The genus Apallates (Fig. 7A) comprises more than 30 species (Evenhuis & Pape, 2020a), widely distributed from North America to the Neotropical region (Sabrosky, 1987). Furthermore, there are records from immigrant Apallates in the Oceanian region (Nartshuk, 2012). Apallates flies use dead bodies of invertebrate during their life cycle (Nartshuk, 2014), and are very likely saprophagous. Interestingly, Wolda and Sabrosky (1986) reported 2 species of Apallates visiting flowers of Aristolochia, a plant genus known for attracting Chloropid and other Dipteran pollinators with strong carrion-like odors, that can even mimic recently dead insects (Oelschlägel et al., 2015). We only found an Apallates specimen within N. turbacensis fruits, probably because the parental fly oviposited on the fruit attracted by the smell of dead insects coming from inside. The literature about the genus Apallates is very scarce, and to our knowledge, this is the first record of these flies within a Lauraceae fruit.
Chaetochlorops inquilinus Coquillett, 1989
The genera Chaetochlorops is distributed in the Nearctic and Neotropical regions (Nartshuk, 2012) and comprises 3 species (Evenhuis & Pape, 2020b). The literature about the genus and C. inquilinus (Fig. 7B) is very scarce; however, the distribution of this fly is well known for central and eastern United States (Sabrosky, 1950). Most specimens are reared rather than collected in the field, and the source of food of the larvae is not completely clear. Several observations of larvae on plants previously injured by other insects and decaying plant material suggest that the larvae are very likely saprophagous, parasitic, or predaceous. Interestingly, these flies have even been reported in Cecidomyiidae (Diptera) galls (Sabrosky, 1950). Furthermore, there are several reports of pupae and larvae of these chloropids recovered from larvae of plant pests including curculionids such as Conotrachelus and moths belonging to Tortricidae, Olethreutidae (Sabrosky, 1950), and Pyralidae families (Neunzig, 1972; Sabrosky, 1950). These observations are interesting regarding our findings. We reared only 1 adult specimen of C. inquilinus from a fruit of N. turbacensis. Unfortunately, we were not able to determine the food source of the larvae with direct observations, and we cannot provide more precise information than the already known. The reared specimen is the first report in wild Lauraceae fruits; furthermore, the occurrence of seed predators and frugivore insects in the same group of fruits suggest a secondary invader lifestyle of C. inquilinus. More details on this specimen can be found in Riccardi and Rodríguez-Sánchez (2021).

Conioscinella sp. Duda, 1929
The genus Conioscinella (Fig. 7C) is distributed all over the world (Nartshuk, 2012). It comprises about 150 species (Evenhuis & Pape, 2020b); furthermore, it is a very complex group with taxonomical problems, and would likely be separated into multiple genera (Wheeler, 2010). The larvae of Conioscinella flies can feed on a wide variety of sources. Some of them are saprophagous and consume dead invertebrate bodies or decaying plant tissues (Nartshuk, 2014). Interestingly, female flies are strongly attracted to scents secreted by plant bugs (Miridae), which act as chemical clues to find and eat recently injured or dead bugs (Zhang & Aldrich, 2004). They can also be generalist scavengers, as demonstrated by Norrbom (1983), who reared Conioscinella flies from dead horseshoe crabs. Larvae are also known as predators of gall former insects associated with grasses (Nartshuk, 2014), and predators of arthropod eggs such as arachnids (Gillung & Borkent, 2017). The specimens of Conioscinella reared from D. ambigens and N. turbacensis fruits were probably saprophagous; however, we cannot rule out the possibility that they were feeding on the fruit decaying tissue. These are likely the first reports of this genera associated with Lauraceae trees.
Drosophilidae
This family has more than 4,400 species distributed all over the world (Bächli, 2020). Drosophilid flies exhibit a wide range of food sources such as fruits, fungi, and flowers (Merritt et al., 2009). They can also feed on tree sap and decomposing organic matter such as fruits and plant tissue. Furthermore, they can prey other invertebrates and be commensals of arthropods such as crabs (Merritt et al., 2009) and spiders (Young, 1982). Despite the common use of drosophilids as experimental model organisms, the ecology of this groups is scarce, especially for immature stages (Valadão et al., 2019).
Drosophilid flies can inhabit a wide variety of habitats in the neotropics including forests and other natural vegetation areas, as well as plantations, gardens, and urban areas (Valadão et al., 2019). There are 100 species of fruit-breeding drosophilid flies in the Neotropics associated with 180 host plant species, most of them characterized by the production of fleshy fruits (Valadão et al., 2019). Drosophilid flies are considered generalist; however, host selection is hypothesized to respond to a certain level of specialization related to the microbiome present in the host plant fruits, which could stablish mutualistic interactions with the flies (Valadão et al., 2019).
Some reports of drosophilid flies associated with Lauraceae include pollinators of Lindera flowers (Dupont & Kato, 1999), as well as nectarivores (adults) and phytophagous (larvae) of Litsea flowers (Kato, 2000). Moreover, fruit feeders have been reported for Lindera (Lee et al., 2015; Van Klinken & Walter, 2001), Cryptocarya (Montgomery, 1975; Van Klinken & Walter, 2001), Endiandra, and Litsea (Van Klinken & Walter, 2001), as well as for the Neotropical species Nectandra megapotamica (Link & Link, 2008), N. grandiflora, Persea americana, Cinnamomum sp., and Ocotea sp., (Gottschalk, 2008; Valadão et al., 2019). Here, we report a drosophilid fly reared from D. salicifolia fruits, which to our knowledge is first register for the fruits of this species and for the genus Damburneya (image not shown).
Lauxaniidae
The family is extremely diverse worldwide, especially in tropical regions (Brown et al., 2010). Of the 94 genera worldwide, 84 genera and 400 species occur in Neotropical region, and 57 genera and 200 species occur in Mexico, Central America, and Caribbean islands. However, it is possible that only a small part of this diversity is known (Brown et al., 2010; Fig. 8A).
This is mostly a saprophagous family, although there are some pollinator species. The existence of phytophagous larvae has been suggested, but there are not accurate observations to corroborate it (Brown et al., 2010; Miller, 1977). Individuals have been reported in bird nests, peaty soil, rotting straw, rotten wood, fallen leaves, decaying vegetation, decaying fruits of Citrus spp., galls of Viola spp. (Miller, 1977), and as pollinators of Lauraceae species such as Lindera erythrocarpa (Dupont & Kato, 1999) and Ocotea catharinensis (Montagna et al., 2018). To our knowledge, this is the first report of a Lauxaniidae species associate with Lauraceae fruits.
Lonchaeidae
Neosilba McAlpine, 1962
Neosilba (Fig. 8B) is an American genus that comprises 40 described species; however, there are still many undescribed species (Galeano-Olaya & Canal, 2012; Martins de Almeida et al., 2019). Around 21 species occur in the Neotropical region (Uchoa, 2012), and 10 species in Mexico and Central America (Brown et al., 2010).
Neosilba flies are mainly frugivorous, but they can also feed on other vegetal tissues (Galeano-Olaya & Canal, 2012; Uchoa, 2012). The larvae can be first or second fruits invaders (Martins de Almeida et al., 2019; McAlpine & Steyskal, 1982), and they can feed on many native and exotic plants specially in the Neotropical region (Costa, 2005; McAlpine & Steyskal, 1982; Raga et al., 2015; Saavedra-Díaz et al., 2017; Uchoa, 2012). Furthermore, many species are polyphagous (Uchoa, 2012). Some species are important pests of commercial fruits worldwide, causing putrefaction indirectly (Martins de Almeida et al., 2019).
Regarding the association with Lauraceae plants, Neosilba can widely attack Persea americana (avocado) fruits, establishing 14 larvae per fruit (Raga et al., 2015; Martins de Almeida et al., 2019). Some of the species that attack avocado fruits are N. batesi, N. certa, N. glaberrima, N. pendula, N. parva, and N. zadolicha (Martins de Almeida et al., 2019; McAlpine & Steyskal, 1982; Raga et al., 2015; Uchoa, 2012). In addition, N. bifida has also been reported in Cinnamomum triplinerve fruits, but in a low proportion (7 larvae in a tree sample; Saavedra-Díaz et al., 2017). It is noteworthy that Neosilba sp. was the only insect species recorded in all the 4 tree species sampled in this study. We mainly observed 1 larva per fruit and occasionally 2. The larvae pupate outside the fruits.
Some parasitoid wasps have been reported attacking Neosilba larvae, including Braconidae (Doryctobracon areolatus, D. crawfordi, Microcasis sp., Phaenocarpa pericaro, Utetes anastrephanae) and Figitidae (Aganaspis nordlanderi, A. pelleranoi, Trybliographa nordlanderi, Lopheucoila anastrephae) families (Costa, 2005; Ruiz-Hurtado et al., 2013; Saavedra-Díaz et al., 2017). We reared A. pelleranoi, 1 of these parasitoid species from N. turbacensis fruits (Table 2).
Muscidae
Atherigona orientalis Schiner,1868
Among the approximately 195 Atherigona species worldwide (Brown et al., 2010), only A. orientalis and A. reversura occur in America (Grzywacz & Pape, 2014). Atherigona orientalis (Fig. 7D) distributes in tropical and subtropical regions worldwide (Pont & Magpayo, 1995; Suh & Kwon, 2016). Larvae are polyphagous (Suh & Kwon, 2016), and have been described as saprophagous or phytophagous (Couri & De Araújo, 1992; Grzywacz & Pape, 2014; Ribeiro et al., 2016). The eggs are usually deposited in feces, vertebrate, or invertebrate carrion, and decaying or living plant tissue such as fruits (Couri & De Araújo, 1992; Grzywacz & Pape, 2014; Pont & Magpayo, 1995; Suh & Kwon, 2016). Furthermore, this species is considered of forensic and sanitary importance, since it is a pathogen vector (Grzywacz & Pape, 2014; Ribeiro et al., 2016).
Atherigona orientalis can be primary or secondary invader of plants and is considered pest of some Poaceae and Solanaceae species (Couri & De Araújo, 1992; Ribeiro et al., 2016; Suh & Kwon, 2016). It is difficult to determine whether the individuals are saprophagous or phytophagous within the fruits (Suh & Kwon, 2016). Atherigona orientalis has been recorded in fruits of several plants including Persea americana and other plants of Amaryllidaceae, Apiaceae, Brassicaceae, Curcubitaceae, Fabaceae, Lauraceae, Poaceae, Moraceae, Myrtaceae, Rosaceae, Rutaceae, and Solanaceae families (Ribeiro et al., 2016; Suh & Kwon, 2016). We are not certain about the food source of the A. orientalis individuals reared from N. turbacensis fruits.
Phoridae
The family comprises more than 4,000 species distributed worldwide (Pape & Thompson, 2020; Fig. 7E, F). These flies have a wide range of food sources like decaying organic matter such as plants and corpses, as well as fungi, and flowers. Furthermore, they commonly are found in dump places, burrows, and nests of birds, bees, wasps, and termites (Peterson, 1987). There are reports of larvae feeding on seed capsules, feces, gastropods, and a wide variety of insects. Also, they can parasite insects and other arthropods, and even cause diseases to humans (Peterson, 1987). To our knowledge, there are few known reports of phorid flies associated with Lauraceae plants. For example, Megaselia scalaris larvae are parasitoids of Isognathus caricae (Lepidoptera) that infest avocado fruits and, interestingly, M. scalaris also feeds on the pulp (Souza et al., 2020). Moreover, phorids are also pollinators of wild Lindera species (Dupont & Kato, 1999). We just reared 2 phorid flies from D. ambigens and N. turbacensis fruits (Fig. 7E, F), that were likely parasitoids of Lepidoptera and other insect species or were feeding on fruit or insect decaying matter.
Stratiomyidae
Ptecticus sackenii Williston, 1885
Ptecticus sackenii (Fig. 8C) is distributed in Canada, United States, and Mexico, with some reports in Nicaragua and Costa Rica (GBIF Secretariat, 2021). The literature about this species is very scarce, the reports include larvae found on ripening (Cordero-Jenkins et al., 1990), rotting, or fermenting fruits of several plant species (Woodley, 2009), other rotting plant material, and fungi (McFadden, 1967).
Our records of specimens on the fruits of D. ambigens are not surprising, since the flies likely consumed rotting material within the fruits. The adults emerged at the later stage of the rearing period. According to Hauser, M. (comm. Pers, 2019) it is very unlikely that the larvae were feeding on the seeds. Furthermore, it is worth mentioning that the pupae were attached to the outer surface of D. ambigens fruits and have about the same size as the diameter of the fruits. We have not found any other report of P. sackenii in other Lauraceae species. However, Basset et al. (2019) found several individuals of 4 undetermined species of Ptecticus in fruits of a Lauraceae species in a tropical rainforest in Thailand.
Tachinidae
The family Tachinidae is one of the most diverse within Diptera and comprises about 1,501 genera and 8,500 species. This family has a worldwide distribution, but the higher diversity of species and genera occurs in the Neotropical region (O’Hara, 2014; Fig. 8D). The group has had several taxonomic classification issues that have been reviewed and analyzed in detail by O’Hara (2013).
Tachinid flies are parasites and parasitoids, larvae are commonly endoparasites of insects and other arthropods including arachnids and centipedes. Most of tachinid species, depend on Lepidoptera larvae (Wood, 1987). Furthermore, larvae and adults of Coleoptera are also hosts of several tachinid genera. They can attack wood and soil beetles like Scarabeids, Cerambicids, and Elaterids; and can also parasitize weevils even when hidden within fruits, stems, or other plant structures (Wood, 1987). In addition, other insects such as Orthopteroids, Dictyipteroids, Dermaptera, Hymenoptera, and even some Diptera can be parasitized by tachinids. Moreover, these flies obtain sugar by visiting the flowers of several plant families
(Wood, 1987).
To our knowledge, most of the insects that feed on Lauraceae and are parasitized by tachinids belong to Lepidoptera. The tachinid parasitoid reports include lepidoptera hosts found in fruits of Cinnamomum (Kan et al., 2003), feeding on leaves of Ocotea veraguensis (Corrales & Epstein, 1997), Nectandra megapotamica (Specht et al., 2014), Nectandra sinuata (Myshondt, 1975), and feeding on unspecified structures of avocado (González-Herrera & Soto-Rodríguez, 1998). Furthermore, tachinids have also been reported as floral visitors of several Lauraceae including avocado (Carabalí-Banguero et al., 2018; Castañeda-Vildózola et al., 1999; Wysoki et al., 2002), Sassafras albidum (Tooker et al., 2006), and Ocotea catarinensis (Montana, 2018). Moreover, tachinids are also pollinators of Laurus azorica (Forfang & Olesen, 1998).
We hypothesize that the individuals reared from D. ambigens fruits (Fig. 8D) were likely parasitoids of Lepidoptera. However, we do not rule out that these flies could also parasitize Coleoptera species.
Hymenoptera
Braconidae
Apanteles Foerster, 1862
Apanteles is a polyphyletic genus that comprises approximately 1,300 endoparasitoid species worldwide (Le Masurier, 1987; Mason, 1981; Fig. 9A-D). These wasps attack most Lepidoptera species. Furthermore, they are gregarious or solitary, and can be polyphagous or monophagous (Le Masurier, 1987; Mason, 1981; Wharton et al., 1997; Whitfield, 1995). Apanteles has been registered in many plant families (Rodriguez, 2009), including several records in Lauraceae fruits of species such as Beilchemiedia alloiophylla, Cinnamomum triplinerva, Nectandra purpurea, Ocotea dendrodaphne, O. veraguensis, Persea americana, P. schiedeana, and other unidentified Persea species. In these fruits, the wasps attack Saturniidae (Gupta et al., 2016) and Oecophoridae moths, particularly Stenoma species (Boscán de Martínez & Godoy, 1982; Hoddle & Hoddle, 2008c, 2012; Rodriguez, 2009).
Apanteles species can be dominant gregarious parasitoids of Stenoma catenifer in avocado (P. americana), especially in countries like Guatemala, Venezuela, and Peru (Hoddle & Hoddle, 2012). Some studies reported parasitoid attack rates from 12 to 60% and registered 6-9 pupae per host, causing the death of 12 to 37% of the S. catenifer larvae (Hoddle & Hoddle, 2008a, 2012; Hoddle et al., 2011).
We reared 4 Apanteles species (Fig. 9A-D), which present differences in mesosoma sculpture, petiole shape, and hypopygium flexibility. These individuals formed white “cottony” pupae outside the fruits. Also, we reared individuals of Apanteles sp. and Stenoma catenifer in D. salicifolia and N. turbacensis fruits. Hence, it is very likely that the wasp parasitizes this moth and/or the other Lepidoptera species reared (Table 2).
Aridelus Marshall, 1887
The genus consists of 46 valid species worldwide and 4 described species in the Neotropical region (Campos, 2001; Lee et al., 2017). Aridelus species are koinobiont endoparasitoids of Hemiptera, mainly of Pentatomidae species (Campos, 2001; Lee et al., 2017; Maeto & Kudo, 1992; Wharton et al., 1997). The individual reared from N. turbacensis (Fig. 9E) was probably associated with a host not reported so far or not collected in this study; also, its association with the fruits could be casual.
Bracon Fabricius, 1804
Bracon is one of the richest and most common genera of the Braconidae family, it contains around 1,000 described species worldwide and 14 valid species in Mexico (Fernández & Sharkey, 2006; Yu, 2016; Fig. 9F-H). Nevertheless, both morphologic and phylogenetic data suggest that the genus is paraphyletic. Bracon species are mostly idiobiont, solitary or gregarious, specialist or generalist ectoparasitoids of Coleoptera, Diptera, Hymenoptera (sawflies), and Lepidoptera larvae (Campos, 2001; Shaw & Huddleston, 1991; Wharton et al., 1997). However, 3 phytophagous species have been reported in association with fruits and leaf galls of Burseraceae species (Flores et al., 2007; Perioto et al., 2011; Ranjith et al., 2016).
Particularly in avocado fruits (Persea americana), Bracon species have been recorded attacking the curculionids Conotrachelus persea (Becerril-Garduño, 2017) and Heilipus lauri (Castañeda-Vildózola et al., 2017). Bracon is very likely the only recorded parasitoid genus of Heilipus beetles (Castañeda-Vildózola et al., 2017). We reared 4 Bracon species that show clear morphological differences (Fig. 9F-H; Bracon sp. 4 is not shown). These wasps likely attacked curculionid beetles or other insect groups. However, we recently described 2 new species from the collected specimens: Bracon laurae and Bracon rosamondae, (here referred as Bracon sp 1.) which are very likely phytophagous, a comprehensive and detailed description can be found in Rodríguez-Sánchez et al. (2022).


Eulophidae
Galeopsomyia fausta LaSalle, 1997
Galeopsomyia fausta (Fig. 10A) is an idiobiont ectoparasitoid and parthenogenetic wasp distributed from Mexico to Argentina (LaSalle & Peña, 1997; Llácer et al., 2005). Galeopsomyia species are parasitoid of galls inductors, except for G. fausta which has been registered attacking Phyllocnistis citrella (Lepidoptera) larvae and pupae (LaSalle & Peña, 1997). Phyllocnistis citrella is a citrus miner, originally from India and very recently introduced in America; therefore, no native host of G. fausta is known (LaSalle & Peña, 1997; Llácer et al., 2005; Ruiz et al., 2001).
The specimens of G. fausta that we reared from N. turbacensis fruits likely attacked any of the Lepidoptera species within the fruits. This could be the first evidence of a native host and native plant with which it is associated. Interestingly, this is not the first report of a Galeopsomyia species associated with fruits, since Perioto et al. (2009) found and described G. ituana in Ilex affinis seeds (Aquifoliaceae) from Brazil.
Eurytomidae
Eurytoma Illiger, 1807
The genus comprises about 700 species worldwide, 84 of which occur in the Neotropical region (Gates et al., 2008; Fig. 10B). Eurytoma larvae present many feeding habits. They can be entomophagous of Coleoptera, Diptera, Hymenoptera, and Lepidoptera species, also they can be phytophagous inside galls induced by other insects. Several species are egg, larval and pupal parasitoids of phytophagous and predator species of Hymenoptera, Coleoptera, Diptera, Hemiptera, and Lepidoptera. Some Eurytoma, are even hyperparasitoids of Chalcidoidea superfamily and Braconidae family (Bugbee, 1967; Burks, 1971; Gómez et al., 2011; Zerova & Fursov, 1991). Interestingly, sometimes Eurytoma parasitoids complete their development by feeding on plant tissues (Burks, 1971; Zerova & Fursov, 1991). However, only the hosts of approximately 28 Neotropical species are known (Gates et al., 2008).
Phytophagous Eurytoma associated with seeds have been reported for plants of Apiaceae, Fabaceae, and Rosaceae families (Gómez et al., 2011). On the other hand, Bugbee (1967) reported several insect hosts like Apanteles solitarium, Bracon cephi, and beetles of Curculionidae family in North America. We hypothesize a parasitoid lifestyle of Eurytoma wasps which likely attacked other insects within N. turbacensis fruits. To our knowledge, this is the first report of Eurytoma associated with Lauraceae species.
Sycophila Walker, 1871
Sycophila contains 117 described species worldwide, although there could be cryptic variation (Li et al., 2010; Fig. 10C). These wasps are koinobiont endoparasitoids of eggs and larvae of phytophagous, gall inducers, or inquilines (Diptera, Lepidoptera and Hymenoptera; Gibernau et al., 2002; Gómez et al., 2013; Hanson & Nishida, 2014). However, there are also phytophagous, seed predators, and galls inducers or inquilines Sycophila species (Gibernau et al., 2002; Lotfalizadeh & Gharali, 2007).

Sycophila spp. are associated with many plants, but the most common are the species of Ficus and Quercus (Gómez et al., 2013; Lotfalizadeh & Gharali, 2007). To our knowledge, the individuals of Sycophila reared from N. turbacensis fruits are the first specimens of this genus reported in association with Lauraceae plants. As Sycophila exhibits a wide variety of feeding habits, we have no certainty of its food source. However, we do not rule out that the individuals could have attacked Eurytoma wasps that also occur within N. turbacensis fruits, as suggested by Leite (2014). Furthermore, they could also feed on the seeds, as demonstrated by Perioto et al. (2009) who registered Sycophila wasps feeding on Ilex affinis seeds from Brazil.

Figitidae
Aganaspis pelleranoi Brèthes, 1924
Aganaspis genus was created by Lin in 1987 to include 4 Asian species (A. daci, A. ocellata, A. contracta, and A. major). Later Nordlander suggested that Ganaspis pelleranoi should be placed in Aganaspis genus, and 2 more American species (A. nordlanderi and A. alujai) were added, expanding the genus to 7 species (Díaz et al., 2006; Ovruski et al., 2007). However, Díaz et al. (2006) argued that A. pelleranoi should go back to Ganaspis genus; therefore, the taxonomy of this species remains problematic. Aganaspis pelleranoi (Fig. 10D) has a Neotropical distribution, occurring from Mexico to Argentina (Ovruski et al., 2007). It is a solitary koinobiont endoparasitoid of Tephritidae (Anastrepha spp., Ceratitis capitata and Rhagolethis turpinae) and Loncheidae (Neosilba spp.) flies (Aluja et al., 2009; Díaz et al., 2006; Guimarães et al., 2003).
Aganaspis pelleranoi individuals have been recorded in a variety of fruits, especially in Myrtaceae and Rutaceae fruits, commercial orchards, and areas with native trees. Although they can attack their hosts within fruits on the trees, they frequently attack on fallen fruits. Individuals settle on the pulp attracted by fruit volatiles (Aluja et al., 1998, 2009; Gonçalves et al., 2013). On the other hand, A. pelleranoi is a potential biological control of some Tephritidae and Loncheidae flies that are pests of commercial fruits (Ovruski et al., 2007). Interestingly, this wasp may enter in diapause (Aluja et al., 1998). From our knowledge, the individual reared from N. turbacensis fruits is the first report of an association of this wasp with Lauraceae fruits. In this case it is very likely that the wasp was attacking Neosilba sp. flies that were also reared from these fruits (Table 2).
Formicidae
Mycocepurus goeldii Forel, 1893
Mycocepurus goeldii has only been recorded in Guyana, Paraguay, Brazil, and Argentina, where it commonly occurs (Mackay et al., 2004; Wild, 2007). Therefore, the specimen that we found is probably the first report of this species for Mexico (Fig. 11A). To our knowledge, M. smithii is the only species of the genus reported so far at Los Tuxtlas rainforest (Quiroz-Robledo & Valenzuela, 1995).
Mycocepurus species are fungus-growing ants with diurnal foraging activity; they collect different vegetal material from several plant families such as flowers and fruit pulp, which they use as a fungal-culturing substrate (Leal & Oliveira, 1998). Mycocepurus ants can even promote the germination of some plants by removing the pulp (Oliveira et al., 1995), but there are also records of endosperm removal from the seeds of some plant species (Leal & Oliveira, 2000). Furthermore, they can also act as seed dispersers (Christianini et al., 2007). Mycocepurus species also forage on corpses and insect feces of Lepidoptera and Coleoptera. The foraging activity depends on the nest location and its vicinity to resources and always occurs on the ground (Leal & Oliveira, 2000). Particularly, M. goeldii has been observed in Brazil foraging on the ground of fields and never climbing up on the plants. The workers collect Baccharis dracunculifolia (Asteraceae) and Schinus terebinthifolius (Anacardiaceae) flowers, Bidens pilosus (Asteraceae) seeds, and caterpillar droppings (Kempf, 1963).
Leal and Oliveira (1998) reported Mycocepurus sp. ants foraging on Ocotea fruits, and they likely use many more Lauraceae fruits as a resource. We found a single individual of M. goeldii from D. ambigens fruits collected from the ground. It probably was foraging the pulp, insect feces, or corpses inside the seeds.
Pheidole Westwood, 1839
Pheidole is the largest ant genus containing 8% of the species. This monophyletic and hyperdiverse genus comprises 1,124 valid species, but hundreds remain to be discovered; estimations point nearly 1500 species (Moreau, 2008; Sarnat et al., 2015; Wilson, 2003; Fig. 11B). Pheidole distributes worldwide in all biomes and a wide range of environmental conditions. This genus originated in America, where there are 624 species, the greatest species richness reported (Moreau, 2008; Wilson, 2003). In addition, individuals encapsulated in amber have been recorded from Late Eocene in Colorado and Miocene in the Dominican Republic and Mexico (Chiapas; Moreau, 2008; Varela-Hernández & Riquelme, 2021). Species richness has a positive correlation with temperature, precipitation, and area (Economo et al., 2015). This genus is dominant in the number of colonies, workers, and biomass in tropical regions (Economo et al., 2015; Wilson, 2003). In Mexico, 132 species have been recorded, 22 in the state of Veracruz and 4 in Los Tuxtlas rainforest (P. scabriventris, P. psilogaster, P. mooreorum, and P. tuxtlasana; Vásquez-Bolaños, 2011; Wilson, 2003).
Pheidole workers are dimorphic in size, degree of morphological specialization, behavior, and numerical representation. Minor workers forage and perform quotidian tasks within the nest, and large-headed majors specialize in seed milling, abdominal food storage, and defense (Moreau, 2008; Wilson, 1984). These ants are scavengers, predators, and seed-predators. Seeds are often stored in granaries within the ant nest, leading to seed dispersal, predation, and even germination. This behavior is a widespread trait in Pheidole that likely evolved multiple times and may have influenced the genus radiation, taking advantage of a resource that other ants cannot exploit. However, the life history of many species remains unknown (Moreau, 2008).
Although Pheidole ants do not usually remove strong seeds (Moreau, 2008), their association with Lauraceae has been reported. An individual was observed visiting the coccid Bombacoccus aguacatae on Persea americana stems (Kondo, 2010). Furthermore, several species of Pheidole were recorded inspecting and cleaning seeds through pulp removal on the spot and removing seeds of Ocotea pulchella and Ocotea spectabilis (Christianini et al., 2007). We only found 1 individual of Pheidole among thousands of D. ambigens fruits, but this record is not surprising, since fruits rich in lipids attract Pheidole ants (Moreau, 2008).
Ichneumonidae
This is one of the most diverse families of insects, it contains 1,538 genera and over 24,000 described species worldwide, although at least 100,000 species are estimated (Ruiz-Cancino et al., 2014). Around 7,400 species are distributed in Neotropical region, and approximately 1,300 species and 343 genera are distributed in Mexico, of which 580 are endemic (Fernández & Sharkey, 2006; Khalaim et al., 2018; Ruiz-Cancino et al., 2014). This monophyletic family consist of solitary or gregarious, idiobiont or kionobiont, ecto or endoparasitoids, and even hyperpasitoids. They attack larvae and pupae of holometabolous insects like Lepidoptera, Coleoptera, Diptera and Hymenoptera (Fernández & Sharkey, 2006; Ruiz-Cancino et al., 2014). We were unable to properly identify the specimen reared from D. ambigens fruits because it was deteriorated (Fig. 10E).
Lepidoptera
We found 5 morphospecies of microlepidoptera in Nectandra and Damburneya fruits (Fig. 12). All of them pupated outside the fruits and were likely seed predators or frugivores; however, we only were able to determine the taxonomical identity of 1 species (Stenoma catenifer, Fig. 12E, see description below) because most of the specimens were deteriorated or had a very small size.
Thus, here we summarize the relevant references for Lepidoptera associated with fruits and seeds of Lauraceae species. Most of them are moths, a group with a complex taxonomy, usually difficult for non-specialists (Hoddle & Parra, 2013). It is common to find descriptions of new species or taxonomical works during a bibliographic search, especially belonging to Tortricidae, a family typically associated with avocado orchards (Brown & Hoddle, 2010; Hoddle & Hoddle, 2008c). The attack of lepidopteran larvae is easily detected in the fruits by the accumulation of feces on the fruit surface, the presence of holes and tunnels, and the liberation of perseitol exudates (Brown & Hoddle, 2010).
Several Tortricids reported for avocado and other Lauraceae includes species of Histura (Brown & Hoddle, 2010), Cryptaspasma (Brown & Brown, 2004), Amorbia (Phillips-Rodriguez & Powell, 2007), Netechma (Hoddle & Hoddle, 2008c), Argyrotaenia, Polyortha (Hoddle & Parra, 2013), and Anacrusis, among others (Brown et al., 2014). Furthermore, there have also been reports of other families in avocado orchards such as Noctuidae (Euxoa and Micrathetis), Coleophoridae (Holcocera; Adamski & Hoddle, 2009; Hoddle & Hoddle, 2008b), and Oecophoridae (Stenoma and Antaeotricha; Hoddle & Parra, 2013). At least 111 species of Lepidoptera associated with avocado crops have been reported around the world; however, the knowledge about native Lauraceae that can be potential hosts of these groups is quite scarce (Hoddle & Parra, 2013). For example, Rodriguez (2009) studied in detail the association of Microgastrinae (Hymenoptera) and Lepidoptera in many plant families of Costa Rica. He reported the presence of Arctiidae, Lasiocampidae, Mimallonidae, Oecophoridae (before Elachisitidae), and Saturniidae individuals associated with unidentified Lauraceae species.
Oecophoridae
Stenoma catenifer Walsingham, 1912
Stenoma catenifer (Fig. 12E) is a specialist seed predator of Lauraceae species distributed from Mexico to South America (Hoddle & Hoddle, 2008a). It is a well-known pest of avocado crops (P. americana), although it has also been reported in Beilschmiedia sp., Persea schiedeana (Cervantes, 1999; Royals et al., 2016), Chlorocardium rodiei (Cervantes, 1999), Nectandra megapotamica, and Cinnamomum camphora (Link & Link, 2008). A more detailed review of this information can be found in Hoddle and Parra (2013).
Besides seeds, S. catenifer can also feed on young branches and stems, fruit pedicels, and pulp, causing premature fruit drop and even the death of small trees (Hoddle & Hoddle, 2008b, c). The number of larvae per fruit and the infestation percentage could vary between Lauraceae species from about 3 to10% in Chlorocardium rodiei (Cervantes et al., 1999), 1.5 to 45-95% in P. americana (Hoddle & Hoddle, 2008a) and from 1 to less than 5% in both Cinnamomum camphora and Nectandra megapotamica (Hoddle & Parra, 2013; Link & Link, 2008). In this study, we registered only 1 larva per fruit and an infestation of less than 1%. Interestingly, Cervantes et al. (1999) observed larvae feeding on the top part of the fruits, without causing any harm to the seed embryo. Many eggs are deposited on the pedicel or fruit surface (Hohmann et al., 2003), 11-20 days later the larvae pupate inside the fruits or buried in the ground, and 8-20 days later emerge as adults (Cervantes, 1999; Hoddle & Hoddle, 2008b).
Furthermore, many parasitoids have been reported to attack S. catenifer larvae, belonging to Chrysodoria genus (Diptera) and several genera of Hymenoptera: Apanteles, Brachycyrtus, Chelonus, Dolichogenidea, Eudeleboea, Hymenochaonia, Hypomicrogaster, Macrocentrus, Pristeromerus, Pseudophanertoma, Trichogramma and Xiphosomella (see more detailed information in Hoddle & Hoddle, 2012).
As S. catenifer is frequently associated with Lauraceae fruits, its occurrence in D. ambigens, D. salicifolia and N. turbacensis could be expected, although surprisingly it has not been reported before in the fruits of neither of these species.

Thysanoptera
Phlaeothripidae
The family Phlaeothripidae (suborder Tubulifera) includes about 400 genera and more than 3,550 species, most of them within the subfamily Phaleothripinae (Mound, 2013). Phlaeothripids occur in the tropics and have varied feeding habits (Thrips Wiki Contributors, 2020; Fig. 13). They can feed on litter fungi (Mound, 1977), and can be phytophagous, predators, and even gall formers and inquilines in several plant families (López-Núñez et al., 2019).
Most of the thrips associated with Lauraceae have been reported on avocado orchards and several species are economically important pests (Hoddle et al., 2002). Many of the reports belong to phytophagous thrips, specially of the Thripidae family. However, there are also several records in avocado of phytophagous Phlaeothripidae, such as the pest Pseudophilothrips perseae, or predator species of Lepthothrips (Cambero-Campos et al., 2011; Sánchez-Roncancio et al., 2001) like L. mcconnelli attacking other pest thrips (Hoddle et al., 2003; Johansen et al., 1999). Furthermore, Hoddle and Mound (2020) reported species of other 7 Phlaeothripidae genera in avocado crops in Tanzania in different plant structures. On the other hand, Androthrips ramachandrai has been found in the wood of avocado trees and is very likely a natural enemy of the ambrosia beetles that cause the laurel wilt disease (Peña et al., 2015). Moreover, the thrips of the Phlaeothripid family can induce galls in leaves of wild Lauraceae, like Nectandra cuspidata (Silva et al., 2018).
The food source of the individual found in D. ambigens (Fig. 13) is uncertain, it was likely predator of other insects that inhabited the fruits or feed on the pulp. However, the presence of this insect could be casual. We do not even rule out the possibility that this undetermined specimen could be feeding on the litter because the fruits of D. ambigens were collected from the ground.
Discussion
Highlights of the association of insects and Lauraceae fruits. Our study aimed to characterize the diversity of insects inhabiting the fruits of 4 wild Lauraceae species at Los Tuxtlas region. Here, we provide novel and surprising evidence of the complexity of insect communities occurring inside the fruits of D. ambigens, D. gentlei, D. salicifolia, and N. turbacensis. We reared 54 insect species from approximately 6,500 Damburneya and Nectandra fruits, in Los Tuxtlas region (Table 2), which contrast with the 2 previously reported species (P. frontalis and H. albomaculatus associated with D. ambigens; Sánchez-Garduño, 1995). The sampling represented most of the expected insect species through a high sample completeness of D. ambigens, D. salicifolia, and N. turbacensis fruits (Table 3). Conversely, the sampling of D. gentlei was deficient, and it is required a sampling of at least 10 trees to achieve a most complete representation of the insect fauna associated with the fruits of this tree species.

This research allowed us to describe 2 new species of wasp, which are very likely phytophagous, a group with very few reports worldwide (Rodríguez-Sánchez et al., 2022). Furthermore, based on our comprehensive bibliographic review, we provide the first report of the association of most of the studied insects with Damburneya and Nectandra fruits. Likewise, almost half of the insect species have never been reported in Lauraceae fruits, and H. guttiger and G. fausta are reported in association with a native host plant for the first time (see a synthesis of this data in Supplementary material table S2). Furthermore, the reared insects represent a great taxonomic, ecological, and lifestyle diversity that occurs naturally in a variety of geographical distribution ranges (Table 2; Table 3; see annotated taxonomic list section). About 55% of the species have a worldwide distribution, and 30% are restricted to the Neotropical region. It is worth highlighting that our records of C. maculatus and Mycocepurus goeldii represent potential new records for Mexico. Moreover, some of our records remain at the genus level and potentially represent new species as some of them belong to species-rich or poorly studied groups.
It must be noticed that some of the species recorded are known pests of commercial crops or stored products like P. frontalis, A. orientalis, and S. catenifer. Other unidentified species belong to genera with known pest species like Cryptolestes, Carpophilus, and Neosilba. Also, we recorded 1 specimen of C. cyperi, a potentially invasive species, and H. interstitialis, a species commonly associated with disturbed areas (see details in annotated taxonomic list). Future studies are needed to test if these insects are invasive species that migrate from cultivated plants or if they are naturally distributed in these and other wild Lauraceae plants. Upcoming research efforts must address the potential impact of these insect species in the regeneration of wild plants and in native insect communities and analyze its relationship with forest fragmentation, deforestation, and land-use change.

Insect diversity and ecology. There were differences in the ecological diversity of insects reared from the 4 Lauraceae species fruits (Table 3). Nectandra turbacensis showed the highest insect richness and diversity and the lowest dominance. On the other hand, even with a high sampling completeness, D. ambigens and D. salicifolia showed low values of insect diversity. These differences in insect dominance among tree species are likely explained by the great abundance of P. frontalis, which was especially high in D. ambigens (Table 2, Table 3). Moreover, many other interacting variables like genetic, epigenetic, and phenotypic variations, fruit characteristics (e.g., fruit and seed size), chemical ecology, insect behavior, insect dispersion, insect and plant phenology, population dynamics, and other intra and interspecific interactions could likely explain the insect diversity divergence between Lauraceae species (see some reviews in Burggren, 2017; Forister et al., 2012; Frago et al., 2012; Mitter et al., 1991; Scriber, 2010; Sharma et al., 2021; Szentesi & Jermy, 1990; Thompson et al., 1997). Species of Lauraceae at Los Tuxtlas vary in abundance and distribution along an altitudinal gradient (L. Giraldo-Kalil, personal observation); hence, insect diversity and distribution could likely be influenced by spatial variation of tree species.
The differences in insect diversity among Lauraceae species may also be related to the specificity of the insect species, which is partly determined by their feeding habits (Forister et al., 2012). The casual or unspecific associations are likely expected from generalist insects that inhabit a wide variety of environments, plant structures, and substrates (Forister et al., 2012). This partially supports our observations, since most of the rare species and those that occurred in low abundance could be considered generalists (Supplementary material table S2). Saprophagous insects, for example, can feed on decaying tissue of plants, animals, or other organisms. Also, polyphagous insects are expected to occur in several kinds of substrates and/or in association with several plant families. Conversely, predator and parasitoid insects are constrained by the presence of their insect preys and hosts, respectively, rather than by the plant itself. On the other hand, feeding habits do not necessarily determine specialization. For instance, P. frontalis, a seed predator, is not constrained to Lauraceae fruits and can attack fruits from at least 5 plant families. In contrast, some seed predators like Heilipus species, Conotrachelus, and Stenoma catenifer, which were relatively abundant in comparison with other species, are expected to have a certain degree of specialization to Lauraceae and perhaps to certain Damburneya and Nectandra species, as they only have been reported in association with this plant family (Supplementary material table S2).
The insect species reared from Lauraceae fruits encompass almost all the known feeding habits of insects. We recorded seed predators, phytophagous, insect predators, saprophagous, fungivores, parasites, parasitoids, and even some of the species are known to act as pollinators of several plant species. The wide variety of feeding habits that our study found is likely related with all the stages of fruit development: from seed predators typically expected at the first ripening stages, to saprophagous species more common at the most mature and rotting stages. Furthermore, it evidences the high complexity of the trophic interactions in the insect communities associated with wild Lauraceae fruits. All these feeding habits play an important role in the entire ecosystem, mainly through their interaction with plants (Weisser & Siemann, 2008). For example, seed predators maintain plant species diversity and regulate plant populations by impacting the density of conspecific seedlings and altering plant competition (Janzen, 1971). Phytophagous insects can directly affect carbon storage and plant resource allocation. Saprophagous and fungivores play a crucial role in nutrient cycling and act as ecosystem engineers. Moreover, the populations of all these insects are regulated by predators, parasites, and parasitoids (Weisser & Siemann, 2008). We highlight the need of more studies regarding the association of insects with fruits of wild plants and their role in the regulation of ecosystem processes, which are still scarce worldwide, especially in comparison to works involving cultivated plants and insect pests. This kind of studies are very important to understand how insect trophic networks contribute to the maintenance of insects and plant diversity, and to detect native hosts plants of insects (Copeland et al., 2009).
Lauraceae species exhibit short pulses of massive synchronous fruiting followed by periods with low or nil fruit production. This phenomenon known as masting has been hypothesized as an adaptive reproductive strategy to satiate seed predators when fruiting is abundant allowing a fraction of the seeds to escape from their attack and to diminish seed predators’ populations when fruiting is scarce (Janzen, 1971; Kelly & Sork, 2002). It remains unclear how specialist insects could manage the lack of fruits during such long periods, especially considering that the reproductive phenology of Nectandra and Damburneya is characterized by supra-annual events of flowering and fruiting (Ibarra-Manríquez et al., 1997). They likely move from one Lauraceae species to another during the year or migrate to find available fruits in other
areas. Further research is needed to assess how the reproductive phenology of Lauraceae species affects insect communities and populations.
We described a diverse insect community inhabiting the fruits of Lauraceae and intended to provide basic information for future insect assessment in the study area, a necessary task for conservation and monitoring efforts. Here, besides highlighting the great diversity and numerous ecological functions of insects associated with fruits of wild Lauraceae in a tropical rainforest, we must stress the impact of deforestation, habitat loss and fragmentation, and the reduction of Lauraceae populations on insect diversity. Unfortunately, the rainforest of Los Tuxtlas has been drastically deforested and fragmented since the 1970s, mainly due to livestock activity (Dirzo & García, 1992; Vega-Vela et al., 2018; Von Thaden et al., 2020). Moreover, deforestation is the main threat for the studied plant species (de Kok, 2020a, b, c, d) and likely for all Lauraceae in Mexico (Lorea-Hernández, 2002). Habitat loss is the prime extinction threat for insects, especially to those inhabiting tropical rainforests (Samways, 2007), and together with habitat fragmentation, it can modify dynamic processes of insect species, diminishing species richness and abundance, or changing community structure, ecological interactions, and ecological functions (Martinson & Fagan, 2014; Murphy et al., 2016). Insect response to fragmentation varies according to distinctive characteristics of the species (e.g., dispersion capacity, rarity, specificity, trophic level) and fragment properties (e.g., size, connectivity, type of vegetation), therefore it is important to collect and integrate information of both elements (Didham et al., 1996; Hunter, 2002; Martinson & Fagan, 2014; Murphy et al., 2016; Ruiz-Guerra et al., 2010; Tscharntke et al., 2002; Van Nouhuys, 2005). The loss of Lauraceae trees in the tropical rainforest would lead to several cascading negative effects, including the reduction of key resources like fruits, the decline of insect populations (particularly of specialist insects) associated with Lauraceae fruits, and the critical loss of interaction networks. We encourage future studies to assess the impact of insect-fruit interaction in the recruitment of Lauraceae and other wild tree species and the impact of forest fragmentation on insect communities.
Limitations of the study and recommendations. We must recognize some limitations in insect rearing that could affect the sampling and study of the insects. For example, we found several unidentified larvae that did not complete their development. Also, several reared specimens could not be determined to species level because they were damaged or because they belong to very species-rich or poorly studied groups. We recommend some improvements for future studies: First, individual storage of the fruits before maturation should be considered to accurately establish the insect associations. For that, several conditions are critical, including a suitable temperature, aeration, and a substrate that keeps the moisture as close to the natural conditions as possible to diminish mortality and guarantees successful insect development. Second, it should be considered the study of fruits at different stages of ripening to understand the phases of fruit colonization.
Concluding remarks. This study evidences the high complexity and diversity of insect communities inhabiting Damburneya and Nectandra fruits. We presented data about high species richness that contrast with previous reports for the studied species, even within the very same study area. Furthermore, the insect species exhibited most of the known insects feeding habits, showing the complex trophic interactions occurring within the fruits. Probably most of the species are not restricted to Lauraceae plants, except for some specialized seed predators. Furthermore, several species are pests of commercial crops that must be monitored to understand whether they have a detrimental effect on wild fruits. The fauna inhabiting inside plant structures like fruits is frequently unnoticed, and consequently, underestimated. This study demonstrated that besides their importance on plant recruitment, fruits provide vital resources for the maintenance of insect communities. We stress that the loss of Lauraceae trees would be highly detrimental for insect communities inhabiting the fruits of these plants not only because of the loss of species diversity but also because of the loss of complex trophic interactions. Furthermore, we highlight the need to study and monitor the diversity and natural history of the insect fauna associated with the fruits of Lauraceae and other wild plant families.
Acknowledgments
The authors thank the logistic support of Rosalinda Tapia during laboratory work and to the staff of the “Estación de Biología Tropical Los Tuxtlas (IB-UNAM)” in particular the director, Rosamond Coates-Lutes, during the fieldwork. To Santiago Sinaca Colín and Erli Velasco Sinaca for helping us explore the study area, locate the trees, and collect the fruits, and Andrea Alfaro, Fernando López, Iván Camargo, Ivan De la Cruz, and Lourdes Martínez for field assistance. To Adriana Pérez for her help during insect rearing and collection. Also, we acknowledge the specialists who helped us with the taxonomic determination: Andrew Cline, Gareth Powell, Macotulio Soto-Hernández, Sara López-Pérez, and Thomas Atkinson (Coleoptera); Claudio Barros de Caravalho, Francisco Dzul-Cahuich, Martin Hauser, Paula Riccardi, and Vicente Hernández (Diptera); Adolfo Ibarra (Lepidoptera); Alejandro Zaldívar-Riverón, Lucía Pérez-Benavidez, Sofía Fernández-Parriego, and Maximiliano Vázquez-Franco (Hymenoptera). We thank Cristina Mayorga for her help with specimen mounting and Rubén Pérez and Susana Guzmán-Gómez for assistance with insect photographing. To the anonymous reviewers that helped us improve our manuscript with their suggestions. LJGK acknowledges the grant (No. 463686) provided by the Consejo Nacional de Ciencia y Tecnología (CONACYT) for her Ph. D. studies. LJGK and ERS acknowledge the Programa de Posgrado en Ciencias Biológicas at the Universidad Nacional Autónoma de México for its support during their postgraduate studies. ERS thanks for the Scholarship from Sistema Nacional de Investigadores (SNI, CONACYT). This research is a result of the project “Ecología Evolutiva de especies simpátricas de Nectandra (Lauraceae) en una selva húmeda tropical”, funded by SEP-CONACYT (grant 284692 to JNF).
References
Adamski, D., & Hoddle, M. (2009). A new Holcocera Clemens from Guatemala and redescription of H. iceryaeella (Riley) from the United States (Lepidoptera: Coleophoridae: Blastobasinae: Holcocerini): Two congeners with incidental preference for avocado. Proceedings of the Entomological Society of Washington, 111, 254–262. https://doi.org/10.4289/0013-8797-111.1.254
Aluja, M., López, M., & Sivinski, J. (1998). Ecological evidence for diapause in four native and one exotic species of larval-pupal fruit fly (Diptera: Tephritidae) parasitoids in tropical environments. Annals of the Entomological Society of America, 91, 821–833. https://doi.org/10.1093/aesa/91.6.821
Aluja, M., Ovruski, S. M., Guillén, L., Oroño, L. E., & Sivinski, J. (2009). Comparison of the host searching and oviposition behaviors of the tephritid (Diptera) parasitoids Aganaspis pelleranoi and Odontosema anastrephae (Hymenoptera: Figitidae, Eucoilinae). Journal of Insect Behavior, 22, 423–451. https://doi.org/10.1007/s10905-009-9182-3
Andrade-Lara, O. (1989). Cultivo de insectos depredadores de propágulos de árboles tropicales en Los Tuxtlas, Veracruz (Bachelor´s Thesis). Universidad Nacional Autónoma de México. México, CDMX.
Arnett, R. H. J., Thomas, M. C., Skelley, P. E., & Frank, J. H. (2002). American Beetles, Volume II: Polyphaga: Scarabaeoidea through Curculionoidea. Florida: CRC Press.
Atkinson, T. H. (2020). Bark and Ambrosia Beetles. Retrieved 28 july, 2020 from: https://www.barkbeetles.info
Atkinson, T. H., & Equihua-Martínez, A. (1985a). Lista comentada de los coleopteros Scolytidae y Platypodidae del Valle de México. Folia Enomológica Mexicana, 65, 63–108.
Atkinson, T. H., & Equihua-Martínez, A. (1985b). Notes on the biology and distribution of mexican and central american Scolytidae (Coleoptera). II.Scolytinae: Cryphalini and Corthylini. The Coleopterists Bulletin, 39, 355–363.
Atkinson, T. H., & Peck, S. B. (1994). Annotated checklist of the bark and ambrosia beetles ( Coleoptera : Platypodidae and Scolytidae ) of tropical southern Florida. Florida Entomological Society, 77, 313–329.
Bächli, G. (2020). TaxoDros: the database on taxonomy of drosophilidae. Retrieved 06 october, 2020 from: https://www.taxodros.uzh.ch/
Basset, Y., Ctvrtecka, R., Dahl, C., Miller, S. E., Quicke, D. L. J., Segar, S. T. et al (2019). Insect assemblages attacking seeds and fruits in a rainforest in Thailand. Entomological Science, 22, 137–150. https://doi.org/10.1111/ens.12346
Becerril-Garduño, F. (2017). Evaluación de daño causado por el barrenador pequeño del fruto del aguacate (Conotrachelus perseae Barber) en Meyuca de Morelos, Coatepec Harinas, Estado de México (Bachelor´s Thesis). Universidad Autónoma del Estado de México. Mexico, Estado de México.
Blackwelder, R. E. (1945). Checklist of the coleopterous insects of Mexico, Central America, the West Indies, and South America, pt. 3. Bulletin of the United States National Museum, 185, 343–550. https://doi.org/10.5479/si.03629236.185.3
Boscán de Martínez, N., & Godoy, F. (1982). Apanteles sp (Hymenoptera: Braconidae) parásito del taladrador del fruto del aguacate Stenoma catenifer Walsingham (Lepidoptera: Stenomidae) en Venezuela. Agronomía Tropical, 32, 319–321.
Brown, B. V., Borkent, A., Cumming, J. M., Wood, D. M., Woodley, N. E., & Zumbado, M. (2010). Manual of Central American Diptera. Volume 2. Ottawa: NCR Research Press.
Brown, J. W., & Brown, R. L. (2004). A new species of Cryptaspasma Walsingham (Lepidoptera: Tortricidae: Olethreutinae) from Central America , the Caribbean , and southeastern United, with a catalog of the world fauna of Microcorsini. Proceedings of the Entomological Society of Washington, 106, 288–297.
Brown, J. W., & Hoddle, M. S. (2010). A new species of Histura Razowski (Lepidoptera: Tortricidae: Polyorthini) from Guatemala attacking avocados (Persea americana) (Lauraceae). Proceedings of the Entomological Society of Washington, 112, 10–21. https://doi.org/10.4289/0013-8797-112.1.10
Brown, J. W., Janzen, D. H., Hallwachs, W., Zahiri, R., Hajibabaei, M., & Hebert, P. D. N. (2014). Cracking complex taxonomy of Costa Rican moths: Anacrusis Zeller (Lepidoptera: Tortricidae: Tortricinae). Journal of the Lepidopterists’ Society, 68, 248–263. https://doi.org/10.18473/lepi.v68i4.a3
Brown, S. D. J. (2009). Molecular systematics and colour variation of Carpophilus species (Coleoptera: Nitidulidae) of the South Pacific (Master’s Thesis). Lincoln University. Pennsylvania, USA.
Brown, S. D. J., Armstrong, K. F., & Cruickshank, R. H. (2012). Molecular phylogenetics of a South Pacific sap beetle species complex (Carpophilus spp., Coleoptera: Nitidulidae). Molecular Phylogenetics and Evolution, 64, 428–440. https://doi.org/10.1016/j.ympev.2012.04.018
Bugbee, R. E. (1967). Revision of chalcid wasps of genus Eurytoma in America north of Mexico. Proceedings of the United States National Museum, 118, 433–552. https://doi.org/10.5479/si.00963801.118-3533.433
Burggren, W. W. (2017). Epigenetics in insects: mechanisms, phenotypes and ecological and evolutionary implications. In Verlinden, H. (Ed.), Advances in Insect Physiology, Vol. 53 (pp. 1–30). Academic Press. https://doi.org/10.1016/bs.aiip.2017.04.001
Burgos-Solorio, A., & Equihua-Martínez, A. (2007). Platypodidae y Scolytidae (Coloeptera) de Jalisco, México. Dugesiana, 14, 59–82.
Burks, B. D. (1971). A synopsis of the genera of the family Eurytomidae (Hymenoptera: Chalcidoidea). Transactions of the American Entomological Society, 97, 1–89.
Callejas-Chavero, A., Cornejo-Romero, A., Serrato-Díaz, A., & Rendón-Aguilar, B. (2019). Diversity and trophic structure of insects associated with grains of three maize landraces in San Agustín Loxicha, Oaxaca, Mexico. Entomological Science, 22, 42–47. https://doi.org/10.1111/ens.12335
Cambero-Campos, J., Johansen-Naime, R., García-Martínez, O., Cantu-Sifuentes, M., Cerna-Chavez, E., & Retana-Salazar, A. (2011). Especies depredadoras de trips (Thysanoptera) asociadas a huertas de aguacate en Nayarit, México. Acta Zoológica Mexicana, 27, 115–121. https://doi.org/10.21829/azm.2011.271738
Campos, D. F. (2001). Lista de los géneros de avispas parasitoides Braconidae (Hymenoptera: Ichneumonoidea) de la región Neotropical. Biota Colombiana, 2, 193–232.
Carabalí-Banguero, D., Montoya-Lerma, J., & Carabalí-Muñoz, A. (2018). Dípteros asociados a la floración del aguacate Persea americana Mill cv. Hass en Cauca, Colombia. Biota Colombiana, 19, 92–111. https://doi.org/10.21068/c2018v19n01a06
Castañeda-Vildózola, A., Equihua-Martínez, A., Valdés-Carrasco, J., Barrientos-Priego, A. F., Ish-Am, G., & Gazit, S. (1999). Insectos polinizadores del aguacatero en los estados de México y Michoacán. Revista Chapingo Serie Horticultura, 5, 129–136.
Castañeda-Vildózola, Á., Palacios-Torres, R. E., Sánchez-Pale, J. R., Franco-Mora, O., Valdez-Carrasco, J., & Equihua-Martínez, A. (2017). Sympatry of two species of Heilipus Germar, 1824 (Coleoptera: Curculionidae) infesting avocado (Persea americana Mill.) in central Mexico. The Coleopterists Bulletin, 71, 361–363. https://doi.org/10.1649/0010-065X-71.2.361
Castro-Ccoscco, R. M., & Mejía-Espinoza, K. (2011). Preferencia alimentaria de Pagiocerus frontalis en variedades de maíz en el Distrito de Coya – Provincia de Calca (Bachelor´s Thesis). Universidad Nacional de San Antonio Abad del Cusco. Peru, Cusco.
Cervantes-Mayagoitia, J. F., & Huacuja-Zamudio, A. H. (2017). Guía de los ácaros e insectos herbívoros de México, Vol. 3 Ácaros e insectos dendrófagos de importancia agrícola y forestal. Ciudad de México: Universidad Autónoma Metropolitana.
Cervantes-Peredo, L. (1999). The stenomatine moth, Stenoma catenifer Walsingham: a pre-dispersal seed predator of Greenheart (Chlorocardium rodiei (Schomb.) Rohwer, Richter & van der Werff) in Guyana. Journal of Natural History, 33, 531–542. https://doi.org/10.1080/002229399300236
Chao, A., Kubota, Y., Zelený, D., Chiu, C. H., Li, C. F., Kusumoto, B. et al. (2020). Quantifying sample completeness and comparing diversities among assemblages. Ecological Research, 35, 292–314. https://doi.org/10.1111/1440-1703.12102
Chávez-Pesqueira, M., & Núñez-Farfán, J. (2016). Habitat fragmentation changes the adaptive value of seed mass for the establishment of a tropical canopy tree. Biotropica, 48, 628–637. https://doi.org/10.1111/btp.12346
Christianini, A. V, Mayhé-Nunes, A. J., & Oliveira, P. S. (2007). The role of ants in the removal of non-myrmecochorous diaspores and seed germination in a neotropical savanna. Journal of Tropical Ecology, 23, 343–351. https://doi.org/10.1017/S0266467407004087
Comisión Nacional de Áreas Naturales Protegidas. (2017). Áreas Naturales Protegidas Federales de la República Mexicana. Digital map. Retrieved 05 july 2019 from: http://sig.conanp.gob.mx/website/pagsig/info_shape.htm
Copeland, R. S., Luke, Q., & Wharton, R. A. (2009). Insects reared from the wild fruits of Kenya. Journal of East African Natural History, 98, 11–66.
Cordero-Jenkins, L. E., Jirón, L. F., & Lezama, H. J. (1990). Notes on the biology and ecology of Ptecticus testaceus (Diptera: Stratiomyidae) a soldier fly associated with mango fruit (m . indica) in Costa Rica. Tropical Pest Management, 36, 285–286. https://doi.org/10.1080/09670879009371489
Córdova-Ballona, L., Sánchez-Soto, S., García-López, E., Ortíz-García, C. F., & Romero-Nápoles, J. (2011). Insectos asociados a alimentos vegetales deteriorables en tiendas de autoservicio, en Tabasco, México. Boletín del Museo de Entomología de la Universidad del Valle, 12, 25–32.
Coria-Ávalos, V. M. (1999). Ciclo de vida, fluctuacion poblacional y control del barrenador de la semilla del agucate (Conotrachelus perseae Barber, C. aguacatae B.) (Coleoptera: Curculionidae) en Ziracuateiro, Michoacán, México. Revista Chapingio Serie Horticultura, 5, 313–318.
Corrales, J. F., & Epstein, M. E. (1997). Review of Costa Rican Venadicodia, with descriptions of two new species and localities for V. ruthea (Lepidoptera: Limacodidae). Revista de Biología Tropical, 45, 1093–1105.
Costa, S. G. M. (2005). Himenópteros parasitóides de larvas frugívoras (Diptera: Tephritoidea) na Reserva Florestal Adolpho Ducke, Manaus, Amazonas, Brasil (Master’s Thesis). Universidade Federal Do Amazonas. Brazil, Manaus.
Couri, M. S., & De Araújo, P. F. (1992). The immature stages of Atherigona orientalis Schiner (Diptera: Muscidae). Proceedings of the Biological Society of Washington, 105, 490–493.
de Kok, R. (2020a). Damburneya ambigens. The IUCN Red List of Threatened Species 2020: e.T146610634A146651525. Retrieved 21 January 2021 from: https://dx.doi.org/10.2305/IUCN.UK.2020-1.RLTS.T146610634A146651525.en
de Kok, R. (2020b). Damburneya gentlei. The IUCN Red List of Threatened Species 2020: e.T146611071A146651540. Retrieved 21 January 2021 from: https://dx.doi.org/10.2305/IUCN.UK.2020-3.RLTS.T146611071A146651540.en
de Kok, R. (2020c). Damburneya salicifolia. The IUCN Red List of Threatened Species 2020:e.T146611340A146651565. Retrieved 21 January 2021 from: https://dx.doi.org/10.2305/IUCN.UK.2020-1.RLTS.T146611340A146651565.en
de Kok, R. (2021d). Nectandra turbacensis. The IUCN Red List of Threatened Species 2021: e.T143822730A143953306. https://dx.doi.org/10.2305/IUCN.UK.2021-1.RLTS.T143822730A143953306.en. [accessed 21 January 2021].
Díaz, N. B., Guimarães, J. A., & Gallardo, F. E. (2006). Systematic review of species of the genus Aganaspis Lin (Hymenoptera: Figitidae: Eucoilinae). Transactions of the American Entomological Society, 132, 271–277. https://doi.org/10.3157/0002-8320(2006)132[271:SROSOT]2.0.CO;2
Díaz-Grisales, V., Equihua-Martínez, A., Romero-Nápoles, J., Valdez-Carrasco, J., & Vallejo-Espinosa, L. F. (2021). Nuevos registros de especies y un hospedero vegetal del género Heilipus Germar (Curculionidae: Molytinae: Hylobiini) para Colombia. Acta Zoológica Mexicana (N.S.), 37, 1–12. https://doi.org/10.21829/azm.2021.3712334
Didham, R. K., Ghazoul, J., Stork, N. E., & Davis, A. J. (1996). Insects in fragmented forests: A functional approach. Trends in Ecology and Evolution, 11, 255–260. https://doi.org/10.1016/0169-5347(96)20047-3
Dirzo, R., & García, M. C. (1992). Rates of deforestation in Los Tuxtlas, a neotropical area in southeast Mexico. Conservation Biology, 6, 84–90. https://doi.org/10.1046/j.
1523-1739.1992.610084.x
Dirzo, R., Ibarra-Manríquez, G., & Sánchez-Garduño, C. M. (1997). Nectandra ambigens. In E. González-Soriano, R. Dirzo & R. C. Vogt (Eds.), Historia Natural de Los Tuxtlas (p. 647). Ciudad de México: Intituto de Biología, Instituto de Ecología, Universidad Nacional Autónoma de México/ Conabio.
Dirzo, R., Mendoza, E., & Ortíz, P. (2007). Size-related differential seed predation in a heavily defaunated Neotropical Rain Forest. Biotropica. 39, 355–362.
Dobson, R. M. (1954). The species of Carpophilus Stephens (Col. Nitidulidae) associated with Stored Products. Bulletin of Entomological Research, 45, 389–402. https://doi.org/10.1017/S0007485300027206
Downey, H. (2018). Insect-mediated indirect interactions in tropical forest plant communities (Doctoral Thesis). University of Oxford. England, Oxford.
Dupont, Y. L., & Kato, M. (1999). Phenology and flower-visiting entomofauna of six species of Lindera (Lauraceae) in Japan. Nordic Journal of Botany, 19, 707–718. https://doi.org/10.1111/j.1756-1051.1999.tb00680.x
Economo, E. P., Klimov, P., Sarnat, E. M., Guénard, B., Weiser, M. D., Lecroq, B. et al. (2015). Global phylogenetic structure of the hyperdiverse ant genus Pheidole reveals the repeated evolution of macroecological patterns. Proceedings of the Royal Society B: Biological Sciences, 282, 20141416. https://doi.org/10.1098/rspb.2014.1416
Eidt-Wendt, J., & Schulz, F. A. (1990). Studies on the biology and ecology of Pagiocerus frontalis (Fabricius) (Coleoptera: Scolytidae) infesting stored maize in Ecuador. In F. Fleurat-Lessard & P. Ducom (Eds.), Proceedings of the Fifth International Working Conferences on Stored-Product Protection (pp. 61–69). Bordeaux.
Equihua-Martínez, A. (1992). Coleopteros Scolytidae atraídos a trampas NTP-80 en el Soconusco, Chiapas, México. Folia Entomológica Mexicana, 84, 55–66.
Estrada-Valencia, A., & Atkinson, T. H. (1988). Scolytidae y Platypodidae (Coleoptera) de Escárcega, Campeche, México. Biogeografía, biología, importancia económica y una lista comentada de especies. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 58, 199–220.
Evenhuis, N., & Pape, T. (Eds.). (2020a). Apallates Sabrosky. Retrieved 07 october, 2020 from: http://www.diptera.org/Nomenclator/Details/264464
Evenhuis, N., & Pape, T. (Eds.). (2020b). Conioscinella Duda. Retrieved 06 october, 2020 from: http://www.diptera.org/Nomenclator/Details/253486
Falcón-Brindis, A., Pérez-de la Cruz, M., Mata-Zayas, E. E., De la Cruz-Pérez, A., Sánchez-Soto, S., & Burelo-Ramos, C. M. (2018). Scolytinae y Platypodinae (Coleoptera: Curculionidae) de Tabasco, México. Acta Zoológica Mexicana (N.S.), 34, 1–10. https://doi.org/10.21829/azm.2018.3412107
Fernández, M., & Sharkey, M. (2006). Introducción a los Hymenoptera de la Región Neotropical. In Universidad Nacional de Colombia (Vol. 1). https://doi.org/10.1017/CBO9781107415324.004
Flores, S., Nassar, J. M., & Quicke, D. L. J. (2007). Reproductive phenology and pre-dispersal seed-feeding in Protium tovarense (Burseraceae ), with a description of the first known phytophagous “Bracon” species (Hymenoptera: Braconidae: Braconinae). Journal of Natural History, 39, 3663–3685. https://doi.org/10.1080/00222930500392659
Ford, E. J. (1996). The Genus Stelidota Erichson in North America: a new species from Florida, new synonymy and lectotype designations (Coleoptera: Nitidulidae). The Coleopterists Bulletin, 50, 149–153.
Forfang, A. S., & Olesen, J. M. (1998). Male – biased sex ratio and promiscuous pollination in the dioecious island tree Laurus azorica (Lauraceae). Plant Systematics and Evolution, 212, 143–157.
Forister, M. L., Dyer, L. A., Singer, M. S., Stireman III, J. O., & Lill, J. T. (2012). Revisiting the evolution of ecological specialization, with emphasis on insect–plant interactions. Ecology, 93, 981–991. https://doi.org/10.1890/11-0650.1
Frago, E., Dicke, M., & Godfray, H. C. J. (2012). Insect symbionts as hidden players in insect-plant interactions. Trends in Ecology and Evolution, 27, 705–711. https://doi.org/10.1016/j.tree.2012.08.013
Galeano-Olaya, P. E., & Canal, N. A. (2012). New species of Neosilba McAlpine (Diptera: Lonchaeidae) and new records from Colombia. Papéis Avulsos de Zoologia, 52, 361–385. https://doi.org/10.1590/S0031-10492012021100001
Gates, M., Mena-Correa, J., Sivinski, J., Ramírez-Romero, R., Córdova-García, G., & Aluja, M. (2008). Description of the Immature Stages of Eurytoma sivinskii Gates and Grissell (Hymenoptera: Eurytomidae), an Ectoparasitoid of Anastrepha (Diptera: Tephritidae) Pupae in Mexico. Entomological News, 119, 354–360. https://doi.org/10.3157/0013-872x-119.4.354
GBIF Secretariat (2021). Ptecticus sackenii Williston, 1885 in GBIF Secretariat (2021). GBIF Backbone Taxonomy. Checklist dataset. Accessed via GBIF.org. Retrieved 08 august 2021 from: https://doi.org/10.15468/39omei
Gentry, A. H. (1988). Changes in plant community diversity and floristic composition on environmental and geographical gradients. Annals of the Missouri Botanical Garden, 75, 1–34.
Gerónimo-Torres, J. C., Pérez-De la Cruz, M., De la Cruz-Pérez, A., & Torres-De la Cruz, M. (2015). Scolytinae y Platypodinae ( Coleoptera : Curculionidae ) asociados a manglares de Tabasco, México. Revista Colombiana de Entomologia, 41, 257–261.
Gibernau, M., Albre, J., Dejean, A., & Barabé, D. (2002). Seed predation in Philodendron solimoesense (Araceae) by chalcid wasps (Hymenoptera). International Journal of Plant Sciences, 163, 1017–1023. https://doi.org/10.1086/342628
Gillung, J. P., & Borkent, C. J. (2017). Death comes on two wings: A review of dipteran natural enemies of arachnids. Journal of Arachnology, 45, 1–19. https://doi.org/10.1636/JoA-S-16-085.1
Gómez, J. F., Nieves-Aldrey, J. L., Nieves, M. H., & Stone, G. N. (2011). Comparative morphology and biology of terminalinstar larvae of some Eurytoma (Hymenoptera, Eurytomidae) species parasitoids of gall wasps (Hymenoptera, Cynipidae) in western Europe. Zoosystema, 33, 287–323. https://doi.org/10.5252/z2011n3a3
Gómez, J. F., Nieves-Aldrey, J. L., & Stone, G. N. (2013). On the morphology of the terminal-instar larvae of some European species of Sycophila (Hymenoptera: Eurytomidae) parasitoids of gall wasps (Hymenoptera: Cynipidae). Journal of Natural History, 47, 2937–2960. https://doi.org/10.1080/00222933.2013.791937
Gonçalves, R. S., Nava, D. E., Pereira, H. C., Lisbôa, H., Grützmacher, A. D., & Valgas, R. A. (2013). Biology and fertility life table of Aganaspis pelleranoi (Hymenoptera: Figitidae) in larvae of Anastrepha fraterculus and Ceratitis capitata (Diptera: Tephritidae). Annals of the Entomological Society of America, 106, 791–798. https://doi.org/10.1603/AN13044
González-Herrera, A., & Soto-Rodríguez, G. (1998). Bombyliidae y Tachinidae (Diptera) parasitoides de Lepidoptera en Costa Rica. Revista de Biología Tropical, 46, 1147–1148.
Gottschalk, M. S. (2008). Utilização de recursos tróficos por espécies neotropicais de Drosophilidae (Diptera) (Doctoral Thesis). Universidade Federal do Rio Grande do Sul. Brazil, Porto Alegre.
Grzywacz, A., & Pape, T. (2014). Larval morphology of Atherigona orientalis (Schiner) (Diptera: Muscidae) —a species of sanitary and forensic importance. Acta Tropica, 137, 174–184. https://doi.org/10.1016/j.actatropica.2014.05.018
Guimarães, J. A., Gallardo, F. E., Diaz, N. B., & Zucchi, R. A. (2003). Eucoilinae species (Hymenoptera: Cynipoidea: Figitidae) parasitoids of fruit-infesting dipterous larvae in Brazil: identity, geographical distribution and host associations. Zootaxa, 278, 1–23. https://doi.org/10.11646/zootaxa.278.1.1
Gupta, A., Das, A. K., Neog, K., & Verghese, A. (2016). First report of Cotesia dictyoplocae (Hymenoptera: Braconidae), a larval parasitoid of Antheraea assamensis (Lepidoptera: Saturniidae), from India. Florida Entomologist, 99, 541–543. https://doi.org/10.1653/024.099.0330
Gutíerrez-García, G., & Ricker, M. (2011). Climate and climate change in the region of Los Tuxtlas (Veracruz, Mexico): A statistical analysis. Atmósfera, 24, 347–373.
Haack, R. A., & Rabaglia, R. J. (2013). Exotic bark and ambrosia beetles in the USA: Potential and current invaders. In J. E. Peña (Ed.), Potential invasive pests of agricultural crops (pp. 48–74). Boston: CAB International.
Hanson, P., & Nishida, K. (2014). Insect galls of Costa Rica and their parasitoids. In G. Fernandes & J. Santos (Eds.), Neotropical insect galls (pp. 497–517). Dordrecht: Springer. https://doi.org/10.1007/978-94-017-8783-3_23
Hoddle, M. S., & Hoddle, C. D. (2008a). Aspects of the field ecology of Stenoma catenifer (Lepidoptera: Elachistidae) infesting hass avocados in Guatemala. Florida Entomologist, 91, 693–694. https://doi.org/10.1653/0015-4040-91.4.693
Hoddle, M. S., & Hoddle, C. D. (2008b). Bioecology of Stenoma catenifer (Lepidoptera: Elachistidae) and associated larval parasitoids reared from Hass avocados in Guatemala. Journal of Economic Entomology, 101, 692–698. https://doi.org/10.1603/0022-0493(2008)101[692:boscle]2.0.co;2
Hoddle, M. S., & Hoddle, C. D. (2008c). Lepidoptera and associated parasitoids attacking hass and non-Hass avocados in Guatemala. Journal of Economic Entomology, 101, 1310–1316. https://doi.org/10.1093/jee/101.4.1310
Hoddle, M. S., & Hoddle, C. D. (2012). Surveys for Stenoma catenifer (Lepidoptera: Elachistidae) and associated parasitoids infesting avocados in Perú. Journal of Economic Entomology, 105, 402–409. https://doi.org/10.1603/ec11414
Hoddle, M. S., Jetter, K. M., & Morse, J. G. (2003). The economic impact of Scirtothrips perseae Nakahara (Thysanoptera: Thripidae) on California avocado production. Crop Protection, 22, 485–493. https://doi.org/10.1016/S0261-2194(02)00199-0
Hoddle, M. S., Millar, J. G., Hoddle, C. D., Zou, Y., McElfresh, J. S., & Lesch, S. M. (2011). Field optimization of the sex pheromone of Stenoma catenifer (Lepidoptera: Elachistidae): evaluation of lure types, trap height, male flight distances, and number of traps needed per avocado orchard for detection. Bulletin of Entomological Research, 101, 145–152. https://doi.org/10.1017/s0007485310000301
Hoddle, M. S., Morse, J. G., Phillips, P. A., Faber, B. A., & Jetier, K. M. (2002). Avocado thrips: New challenge for growers. California Agriculture, 56, 103–107. https://doi.org/10.3733/ca.v056n03p103
Hoddle, M. S., & Mound, L. A. (2020). Surveys for Thrips (Thysanoptera) associated with avocados (Persea americana) in Mbeya, Tanzania. African Entomology, 28, 195–197. https://doi.org/10.4001/003.028.0195
Hoddle, M. S., & Parra, J. R. P. (2013). Potential lepidopteran pests associated with avocado fruit in parts of the home range of Persea americana. In J. E. Peña (Ed.), Potential invasive pests of agricultural crops (pp. 86–97). Boston: CABI. https://doi.org/10.1079/9781845938291.0086
Hohmann, C. L., Meneguim, A. M., Andrade, E. A., Novaes, T. G. de & Zandoná, C. (2003). The avocado fruit borer, Stenoma catenifer (wals.) (Lepidoptera: Elachistidae): egg and damage distribution and parasitism. Revista Brasileira de Fruticultura, 25, 432–435. https://doi.org/10.1590/S0100-29452003000300018
Hunter, M. D. (2002). Landscape structure, habitat fragmentation, and the ecology of insects. Agricultural and Forest Entomology, 4, 159–166. https://doi.org/10.1046/j.1461-9563.2002.00152.x
Ibarra-Manríquez, G., Ricker, M., Ángeles, G., Sinaca-Colín, S., & Sinaca-Colín, M. A. (1997). Useful plants of the Los Tuxtlas rain forest (Veracruz, Mexico): considerations of their market potential. Economic Botany, 51, 362–376. https://doi.org/10.2307/4255991
Ibarra-Manríquez, G., & Sinaca-Colín, S. (1995). Lista florística comentada de la Estación de Biología Tropical “Los Tuxtlas”, Veracruz, México. Revista de Biología Tropical, 43, 75–115.
James, D. G., Faulder, R. J., & Bartelt, R. J. (1995). Fauna and seasonal abundance of Carpophilus spp. (Coleoptera: Nitidulidae) in four stone fruit growing regions of southeastern Australia as determined by pheromone-trapping. Australian Journal of Entomology, 34, 327–333. https://doi.org/10.1111/j.1440-6055.1995.tb01350.x
James, D. G., & Vogele, B. (2000). Development and survivorship of Carpophilus hemipterus (L.), Carpophilus mutilatus Erichson and Carpophilus humeralis (F.) (Coleoptera: Nitidulidae) over a range of constant temperatures. Australian Journal of Entomology, 39, 180–184. https://doi.org/10.1046/j.1440-6055.2000.00163.x
Janzen, D. H. (1971). Seed predation by animals. Annual Review of Ecology and Systematics, 2, 465–492. https://doi.org/10.1146/annurev.es.02.110171.002341
Janzen, D. H. (1987). Insect diversity of a Costa Rican dry forest: why keep it, and how? Biological Journal of the Linnean Society, 30, 343–356. https://doi.org/10.1111/j.1095-8312.1987.tb00
307.x
Johansen, R., Mojica-Guzmán, A., & Ascención-Betanzos, G. (1999). Introducción al conocimiento de los insectos tisanópteros mexicanos, en el aguacatero (Persea americana Miller). Revista Chapingo Serie Horticultura, 5, 279–285.
Kan, E., Fukuhara, N., & Hidaka, T. (2003). Parasitism by tachinid parasitoids (Diptera: Tachinidae) in connection with their survival strategy. Applied Entomology and Zoology, 38, 131–140. https://doi.org/10.1303/aez.2003.131
Kato, M. (2000). Anthophilous insect community and plant-pollinator interactions on Amami Islands in the Ryukyu Archipelago , Japan. Contributions from the Biological Laboratory, Kyoto University, 29, 157–254.
Kelly, D., & Sork, V. L. (2002). Mast seeding in perennial plants: Why, how, where? Annual Reviews of Ecology and Systematics, 33, 427-447.
Kempf, W. W. (1963). A review of the ant genus Mycocepurus Forel, 1893 (Hymenoptera: Formicidae). Studia Entomologica, 6, 417–432.
Khalaim, A. I., Kasparyan, D. R., & López-Ortega, M. (2018). New records and descriptions of Ichneumonidae (Hymenoptera) from Mexico. Zootaxa, 4486, 1–30. https://doi.org/10.11646/zootaxa.4486.1.1
Kirkendall, L. R., Biedermann, P. H. W., & Jordal, B. H. (2015). Evolution and diversity of bark and Ambrosia beetles. In F. E. Vega, & R. W. Bierdermann (Eds.), Bark beetles (pp. 85–156). London: Academic Press. https://doi.org/10.1016/B978-0-12-417156-5.00003-4
Kirkendall, L. R., & Ødegaard, F. (2007). Ongoing invasions of old-growth tropical forests: Establishment of three incestuous beetle species in southern Central America (Curculionidae: Scolytinae). Zootaxa, 1588, 53–62. https://doi.org/10.11646/zootaxa.1588.1.3
Kondo, T. (2010). Description of a new coccid (Hemiptera, Coccidae) on avocado (Persea americana Mill.) from Colombia, South America. Zookeys, 42, 37–45. https://doi.org/10.3897/zookeys.42.377
LaSalle, J., & Peña, J. E. (1997). A New Species of Galeopsomyia (Hymenoptera: Eulophidae: Tetrastichinae): A fortuitous parasitoid of the citrus leafminer, Phyllocnistis citrella (Lepidoptera: Gracillariidae). The Florida Entomologist, 80, 461–470. https://doi.org/10.2307/3495611
Le Masurier, A. D. (1987). A comparative study of the relationship between host size and brood size in Apanteles spp. (Hymenoptera: Braconidae). Ecological Entomology, 12, 383–393. https://doi.org/10.1111/j.1365-2311.1987.tb01
019.x
Leal, I. R., & Oliveira, P. S. (1998). Interactions between fungus-growing ants (Attini), fruits and seeds in cerrado vegetation in southeast Brazil. Biotropica, 30, 170–178. https://doi.org/10.1111/j.1744-7429.1998.tb00052.x
Leal, I. R., & Oliveira, P. S. (2000). Foraging ecology of attine ants in a Neotropical savanna: Seasonal use of fungal substrate in the cerrado vegetation of Brazil. Insectes Sociaux, 47, 376–382. https://doi.org/10.1007/PL00001734
Lee, H. R., An, T. H., Ku, D. S., & Byun, B. K. (2017). Taxonomic revision of the genus Aridelus Marshall (Hymenoptera: Braconidae: Euphorinae) from Korea. Journal of Asia-Pacific Biodiversity, 10, 409–412. https://doi.org/10.1016/j.japb.2017.05.002
Lee, J. C., Dreves, A. M. Y. J., Cave, A. M., Kawai, S., Isaacs, R., Miller, J. C. et al. (2015). Infestation of wild and ornamental noncrop fruits by Drosophila suzukii (Diptera : Drosophilidae). Annals of the Entomological Society of America, 108, 117–129. https://doi.org/10.1093/aesa/sau014
Leite, G. L. D. (2014). Galling insects on Caryocar brasiliense Camb. (Caryocaraceae). In G. Fernandes, & J. Santos (Eds.), Neotropical insect galls (pp. 179–192). Dordrecht: Springer. https://doi.org/10.1007/978-94-017-8783-3_12
Li, Y., Zhou, X., Feng, G., Hu, H., Niu, L., Hebert, P. D. N. et al. (2010). COI and ITS2 sequences delimit species, reveal cryptic taxa and host specificity of fig-associated Sycophila (Hymenoptera, Eurytomidae). Molecular Ecology Resources, 10, 31–40. https://doi.org/10.1111/j.1755-0998.2009.02671.x
Link, D., & Link, F. M. (2008). Identificação de plantas hospedeiras da broca do abacate, Stenoma catenifer Walsingham (Lepidoptera: Elachistidae) no Rio Grande do Sul. Neotropical Entomology, 37, 342–344. https://doi.org/10.1590/S1519-566X2008000300016
Llácer, E., Urbaneja, A., Garrido, A., & Jacas, J. A. (2005). Morpho-
logy and development of immature stages of Galeopsomyia fausta (Hymenoptera: Eulophidae: Tetrastichinae). Annals of the Entomological Society of America, 98, 747–753. https://doi.org/10.1603/0013-8746(2005)098[0747:madois]2.0.co;2
López-Núñez, F. A., Ribeiro, S., Marchante, H., Heleno, R. H., & Marchante, E. (2019). Life inside a gall: diversity, phenology and structure of portuguese gall communities, their hosts, parasitoids and inquilines. Arthropod-Plant Interactions, 13, 477–488. https://doi.org/10.1007/s11829-018-9655-4
Lorea-Hernández, F. G. (2002). La familia Lauraceae en el sur de México: Diversidad, distribución y estado de conservación. Boletín de la Sociedad Botánica de México, 71, 59–70. https://doi.org/10.17129/botsci.1663
Loschiavo, S. R., & Sinha, R. N. (1966). Feeding, oviposition, and aggregation by the rusty grain beetle, Cryptolestes ferrugineus (Coleoptera: Cucujidae) on seed-borne fungi. Annals of the Entomological Society of America, 59, 578–585. https://doi.org/10.1093/aesa/59.3.578
Lotfalizadeh, H., & Gharali, B. (2007). Some notes on the genus Sycophila Walker, 1871 (Hymenoptera: Eurytomidae) in the west of Iran. Zoology in the Middle East, 40, 77–84. https://doi.org/10.1080/09397140.2007.10638207
Mackay, W. P., Maes, J.-M., Fernández, P. R., & Luna, G. (2004). The ants of north and central America: the genus Mycocepurus (Hymenoptera: Formicidae). Journal of Insect Science, 4, 1–7. https://doi.org/10.1673/031.004.2701
Maeto, K., & Kudo, S. (1992). A new Euphorine species of Aridelus (Hymenoptera, Braconidae) associated with a subsocial bug Elasmucha putoni (Heteroptera, Acanthosomatidae). Japanese Journal of Entomology, 60, 77–84.
Manrique B., M. B., Carabalí, A., Kondo, T., & Bacca, T. (2014). Biología del pasador del fruto del aguacate Stenoma catenifer Walsingham (Lepidoptera: Elachistidae) y búsqueda de sus posibles enemigos naturales. Boletín Científico Centro De Museos De Historia Natural. Universidad de Caldas, 18, 79–92.
Marcon, E., & Hérault, B. (2015). entropart: An R Package to Measure and Partition Diversity. Journal of Statistical Software, 67, 1–26. https://doi.org/10.18637/jss.v067.i08
Martins de Almeida, L. B., Coelho, J. B., Uchoa, M. A., & Gisloti, L. J. (2019). Diversity of fruit flies (Diptera: Tephritoidea) and their host plants in a conservation unit from midwestern Brazil. Florida Entomologist, 102, 562–570. https://doi.org/10.1653/024.102.0333
Martinson, H. M., & Fagan, W. F. (2014). Trophic disruption: a meta-analysis of how habitat fragmentation affects resource consumption in terrestrial arthropod systems. Ecology Letters, 17, 1178–1189. https://doi.org/10.1111/ele.12305
Mason, W. R. M. (1981). The polyphyletic nature of Apanteles Forester (Hymenoptera: Braconidae): a phylogeny and reclassification of Microgastrine. Memoirs of the Entomological Society of Canada, 113, 1–147. https://doi.org/10.4039/entm113115fv
McAlpine, J. F., & Steyskal, G. C. (1982). A revision of Neosilba McAlpine with a key to the world genera of Lonchaeidae (Diptera). The Canadian Entomologist, 114, 105–137. https://doi.org/10.4039/Ent114105-2
McFadden, M. W. (1967). Soldier Fly Larvae in America North of Mexico. Proceedings of the United States National Museum, 121, 1–72. https://doi.org/10.5479/si.00963801.121-3569.1
Merritt, R. W., Courtney, G. W., & Keiper, J. B. (2009). Diptera: (Flies, Mosquitoes, Midges, Gnats). In V. H. Resh, & R. T. Cardé (Eds.), Encyclopedia of insects (pp. 284–297). Amsterdam: Academic Press. https://doi.org/10.1016/B978-0-12-374144-8.00085-0
Miller, R. M. (1977). Ecology of Lauxaniidae (Diptera: Acalyptratae) I. old and new rearing records with biological notes and discussion. Annals of the Natal Museum, 23, 215–238.
Mitter, C., Farrell, B., & Futuyma, D. J. (1991). Phylogenetic studies of insect-plant interactions: Insights into the genesis of diversity. Trends in Ecology & Evolution, 6, 290–293. https://doi.org/10.1016/0169-5347(91)90007-K
Montagna, T., Silva, J. Z., Pikart, T. G., & Reis, M. S. (2018). Reproductive ecology of Ocotea catharinensis, an endangered tree species. Plant Biology, 20, 926–935. https://doi.org/10.1111/plb.12847
Montana, T. (2018). Fundamentos ecológicos e genéticos para a conservação de Ocotea catharinensis Mez (Doctoral Thesis). Universidade Federal de Santa Catarina. Brazil, Florianópolis.
Montgomery, S. (1975). Comparative Breeding site ecology and the adaptive radiation of picture-winged Drosophila (Diptera: Drosophilidae) in Hawaii. Proceedings of the Hawaiian Entomological Society, XXII, 65–103.
Moreau, C. S. (2008). Unraveling the evolutionary history of the hyperdiverse ant genus Pheidole (Hymenoptera: Formicidae). Molecular Phylogenetics and Evolution, 48, 224–239. https://doi.org/10.1016/j.ympev.2008.02.020
Morrone, J. J. (1999). Lista de las especies mexicanas de Molytinae (Coleoptera: Curculionidae). Dugesiana, 6, 51–67.
Morrone, J. J. (2014). Biodiversidad de Curculionoidea (Coleoptera) en México. Revista Mexicana de Biodiversidad, 85, 312–324. https://doi.org/10.7550/rmb.30038
Mound, L. A. (1977). Species diversity and the systematics of some New World leaf litter Thysanoptera (Phlaeothripinae; Glyptothripini). Systematic Entomology, 2, 225–244. https://doi.org/10.1111/j.1365-3113.1977.tb00371.x
Mound, L. A. (2013). Order Thysanoptera Haliday, 1836. Zootaxa, 3703, 49–50. https://doi.org/10.11646/zootaxa.3703.1.11
Muñiz, R. (1970). Estudio morfológico de dos especies de Conotrachelus que son plagas del aguacate (Persea gratissima Gaertn), en México. Revista de la Sociedad Mexicana de Historia Natural, 31, 289–338.
Murphy, S. M., Battocletti, A. H., Tinghitella, R. M., Wimp, G. M., & Ries, L. (2016). Complex community and evolutionary responses to habitat fragmentation and habitat edges: What can we learn from insect science? Current Opinion in Insect Science, 14, 61–65. https://doi.org/10.1016/j.cois.2016.01.007
Myshondt, A. (1975). Notes on the life cycle and natural history of butterflies of El Salvador. V. Anaea (Memphis) morvus boisduvali (Nymphalidae). Journal of the Lepidopterists’ Society, 29, 32–39.
Nartshuk, E. P. (2012). A check list of the world genera of the family Chloropidae (Diptera, Cyclorrhapha, Muscomorpha). Zootaxa, 3267, 1–43. https://doi.org/10.11646/zootaxa.3267.
1.1
Nartshuk, E. P. (2014). Grass-fly larvae (Diptera, Chloropidae): Diversity, habitats, and feeding specializations. Entomolo-
gical Review, 94, 514–525. https://doi.org/10.1134/S00138
7381404006X
Natural Resources Canada, USGS, INEGI, CONABIO & CONAFOR (2015). North American Land Change Monitoring System (NALCMS).. North American Land Cover 30-meter dataset. Digital map. Retrieved 22 January 2020 from: http://www.cec.org/north-american-environmental-atlas/land-cover-30m-2015-landsat-and-rapideye/
Navarrete-Heredia, J. L., & Newton, A. F. (2014). Biodiversidad de Staphylinidae (Insecta: Coleoptera) en México. Revista Mexicana de Biodiversidad, 85, 332–338. https://doi.org/10.7550/rmb.33488
Navarrete-Heredia, J., Newton, A., Thayer, M., Ashe, J., & Chandler, D. (2002). Guía ilustrada para los géneros de Staphylinidae (Coleoptera) de México. Guadalajara: Universidad de Guadalajara/ Conabio.
Neunzig, H. H. (1972). Taxonomy of Acrobasis larvae and pupae in Eastern North America (Lepidoptera: Pyralidae) (Technical). Washington D.C.: United States Department of Agriculture and North Carolina Agricultural Experiment Station.
Noguera-Martinez, F. A., & Atkinson, T. H. (1990). Biogeography and biology of bark and ambrosia beetles (Coleoptera: Scolytidae and Platypodidae) of a mesic montane forest in Mexico, with an annotated checklist of species. Annals of the Entomological Society of America, 83, 453–466. https://doi.org/10.1093/aesa/83.3.453
Norrbom, A. L. (1983). Four acalyptrate Diptera reared from dead horseshoe crabs. Entomological News, 94, 117–121.
O’Hara, J. (2013). History of tachinid classification (Diptera, Tachinidae). Zookeys, 316, 1–34. https://doi.org/10.3897/zookeys.316.5132
O’Hara, J. (2014). World genera of the Tachinidae (Diptera) and their regional occurrence. Version 8.0.
Oelschlägel, B., Nuss, M., Tschirnhaus, M., Pätzold, C., Neinhuis, C., Dötterl, S., & Wanke, S. (2015). The betrayed thief – the extraordinary strategy of Aristolochia rotunda to deceive its pollinators. New Phytologist, 206, 342–351. https://doi.org/10.1111/nph.13210
Okello, S., Reichmuth, C., & Schulz, F. A. (1996). Laboratory investigations on the developmental rate at low relative humidity and the developmental temperature limit of Pagiocerus frontalis (Fab.) (Col., Scolytidae) at high and low temperatures. Anzeiger für Schädlingskunde Pflanzenschutz, Umweltschutz, 69, 180–182.
Oliveira, P. S., Galetti, M., Pedroni, F., & Morellato, L. P. C. (1995). Seed cleaning by Mycocepurus goeldii Ants (Attini) facilitates germination in Hymenaea courbaril (Caesalpiniaceae). Biotropica, 27, 518. https://doi.org/10.2307/2388966
Ovruski, S. M., Wharton, R. A., Rull, J., & Guillén, L. (2007). Aganaspis alujai (Hymenoptera: Figitidae: Eucoilinae), a new species attacking Rhagoletis (Diptera: Tephritidae) in the Neotropical region. Florida Entomologist, 90, 626–634.
Pape, T., & Thompson, F. C. (2020). Systema Dipterorum. Retrieved 06 october, 2020 from: http://www.diptera.org
Peña, J. E., Weihman, S. W., McLean, S., Cave, R. D., Carrillo, D., Duncan, R. E. et al. (2015). Predators and parasitoids associated with Scolytinae in Persea species (Laurales: Lauraceae) and other Lauraceae in Florida and Taiwan. Florida Entomologist, 98, 903–910. https://doi.org/10.1653/024.098.0314
Pérez-De La Cruz, M., Equihua-Martínez, A., Romero-Nápoles, J., Sánchez-Soto, S., García-López, E., & Bravo-Mojica, H. (2009). Escolítidos (Coleoptera: Scolytidae) asociados al agroecosistema cacao en Tabasco, México. Neotropical Entomology, 38, 602–609. https://doi.org/10.1590/S1519-
566X2009000500007
Pérez-De la Cruz, M., Hernández-May, M. A., De la Cruz-Pérez, A., & Sánchez-Soto, S. (2016). Scolytinae y Platypodinae (Coleoptera: Curculionidae) de dos áreas de conservación en Tabasco, México. Revista de Biologia Tropical, 64, 319–326. https://doi.org/10.15517/rbt.v64i1.15931
Pérez-De la Cruz, M., Zavaleta-Bastar, P. G., & De la Cruz-Pérez, A. (2015). Aproximación al conocimiento de la diversidad de Scolytinae y Platypodinae (Coleoptera: Curculionidae) asociados a selvas de Tabasco, México. Entomotropica, 30, 201–211.
Perioto, N. W., Lara, R. I. R., & Costa, V. A. (2009). Parasitic wasps (Hymenoptera) associated with fruits of Ilex affinis Gardner (Aquifoliaceae) in Brazil with description of two new Chalcidoidea. Arquivos do Instituto Biológico, 76, 85–90.
Perioto, N. W., Lara, R. I. R., Ferreira, C. S., Rodrigues-Fernandes, D. R., Pedroso, E. D. C., Linhares-Volpe, H. X. et al. (2011). A new phytophagous Bracon Fabricius (Hymenoptera, Braconidae) associated with Protium ovatum Engl. (Burseraceae) fruits from Brazilian savannah. Zootaxa, 3000, 59–65. https://doi.org/10.11646/zootaxa.3000.1.4
Peterson, B. V. (1987). Phoridae. In J. F. McAlpine (Ed.), Manual of Nearctic Diptera, Volume 2 (pp. 689–712).Otawa: Research Branch Agriculture Canada.
Phillips-Rodriguez, E., & Powell, J. A. (2007). Phylogenetic relationships, systematics, and biology of the species of Amorbia Clemens (Lepidoptera: Tortricidae: Sparganothini). Zootaxa, 1670, 1–21. https://doi.org/10.11646/zootaxa.1670.
1.1
Pont, A. C., & Magpayo, F. R. (1995). Muscid shoot-flies of the Philippine Islands (Diptera: Muscidae, genus Atherigona Rondani). Bulletin of Entomological Research Supplement Series, 3, 1–121. https://doi.org/10.1017/S1367
426900000321
Prado, C. E. (1987). El género Carpophilus Stephens (Coleoptera: Nitidulidae) en Chile. Revista Chilena de Entomología, 15, 27–32.
Quiroz-Robledo, L., & Valenzuela-González, J. (1995). A comparison of ground ant communities in a tropical rainforest and adjacent grassland in Los Tuxtlas, Veracruz, Mexico. Southwestern Entomologist, 20, 203–2013.
R Core Team. (2021). R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
Raga, A., Souza-Filho, M. F. de, Strikis, P. C., & Marangoni-Montes, S. M. N. (2015). Lance fly (Diptera: Lonchaeidae) host plants in the State of São Paulo, Southeast Brazil. Entomotropica, 30, 57–68.
Ranjith, A. P., Quicke, D. L. J., Saleem, U. K. A., Butcher, B. A., Zaldívar-Riverón, A., & Nasser, M. (2016). Entomophytophagy (‘sequential predatory, then phytophagous behaviour’) in an Indian braconid ‘Parasitoid’ wasp (Hymenoptera): specialized larval morphology, biology and description of a new species. Plos One, 11, e0156997. https://doi.org/10.1371/journal.pone.0156997
Ribeiro, L. do P., Netto, A. C. M., Jochims, F., Haseyama, K. L. F., & de Carvalho, C. J. B. (2016). First record of Atherigona reversura Villeneuve (Diptera: Muscidae) feeding on Bermudagrass (Cynodon dactylon cv. Jiggs, Poaceae) in Brazil: morphological and molecular tools for identification. Revista Brasileira de Entomologia, 60, 270–274. https://doi.org/10.1016/j.rbe.2016.04.001
Riccardi, P. R., & Rodríguez-Sánchez, E. (2021). New records and notes on the morphology and biology of Chaetochlorops Malloch (Diptera: Chloropidae). Journal of Insect Biodiversity, 026, 060–065. https://doi.org/10.12976/jib/2021.26.2.5
Rodriguez, J. J. (2009). Integrating molecular, ecological and morphological data to explore the biodiversity of microgastrine parasitoid wasps (Doctoral Thesis). University of Illinois, USA.
Rodríguez-Del Bosque, L. A., Leos-Martínez, J., & Dowd, P. F. (1998). Effect of ear wounding and cultural practices on abundance of Carpophilus freemani (Coleoptera: Nitidulidae) and other microcoleopterans in maize in northeastern Mexico. Journal of Economic Entomology, 91, 796–801. https://doi.org/10.1093/jee/91.4.796
Rodríguez-Sánchez E., Giraldo-Kalil L. J., Quicke D. L. J., & Zaldívar-Riverón A. (2022) Two new species of the braconid wasp genus Bracon (Braconinae) from Los Tuxtlas region in Veracruz, Mexico, reared from fruits of three species of Lauraceae. Zootaxa, 5162, 67–77. https://doi.org/10.11646/zootaxa.5162.1.4
Rohwer, J. G. (1993a). Lauraceae: Nectandra. In Flora Neotropica, Vol. 60 (pp. 1–332). New York: New York Botanical Garden Press.
Rohwer, J. G. (1993b). Lauraceae. In K. Kubitzki, J. G. Rohwer, & Bittrich, V. (Eds.), The families and genera of vascular plants, Vol. 2 (pp. 366–391). Berlin: Springer-Verlag.
Roswell, M., Dushoff, J., & Winfree, R. (2021). A conceptual guide to measuring species diversity. Oikos, 130, 321-338. https://doi.org/10.1111/oik.07202
Royals, H. R., Gilligan, T. M., & Passoa, S. C. (2016). Screening aid: avocado seed moth, Stenoma catenifer Walsingham. Fort Collins: Identification Technology Program.
Ruiz-Cancino, E., Martínez-Bernal, C., Coronado-Blanco, J., Mateos-Crespo, J. R., & Peña, J. E. (2001). Himenopteros parasitoides de Phyllocnistis citrella Stainton (Lepidoptera: Gracillariidae) en Tamaulipas y Norte de Veracruz, México, con una clave para las especies. Folia Enomológica Mexicana, 40, 83–91.
Ruiz-Cancino, E., Rafaelevich-Kasparyan, D., González-Moreno, A., Khalaim, A. I., & Coronado-Blanco, J. M. (2014). Biodiversidad de Ichneumonidae (Hymenoptera) en México. Revista Mexicana de Biodiversidad, Supplem. 85, 385–391. https://doi.org/10.7550/rmb.32448
Ruiz-Guerra, B., Guevara, R., Mariano, N. A., & Dirzo, R. (2010). Insect herbivory declines with forest fragmentation and covaries with plant regeneration mode: Evidence from a Mexican tropical rain forest. Oikos, 119, 317–325. https://doi.org/10.1111/j.1600-0706.2009.17614.x
Ruiz-Hurtado, F. M., Ramírez-Quimbayo, J. H., Rojas-Plazas, B., Galeano-Olaya, E. P., & Canal, N. A. (2013). Diversidad de parasitoides (Hymenoptera) de moscas frugivoras (Díptera: Tephritoidea) en dos áreas cafeteras del departamento del Tolima, Colombia. Revista Tumbaga, 2, 29–53.
Saavedra-Díaz, J., Galeano-Olaya, P. E., & Canal, D., N. A. (2017). Relaciones ecológicas entre frutos hospederos, moscas frugívoras y parasitoides en un fragmento de bosque seco tropical. Revista de Ciencias Agrícolas, 34, 32–49. https://doi.org/10.22267/rcia.173401.61
Sabrosky, C. W. (1950). A synopsis of the chloropid genera Chaetochlorops and Eugaurax (Diptera). Journal of the Washington Academy of Sciences, 40, 183–188.
Sabrosky, C. W. (1987). Chloropidae. In J. F. McAlpine (Ed.), Manual of Nearctic Diptera, Volume 2 (pp. 1049–1068). Quebec: Research Branch Agriculture Canada.
Sallabanks, R., & Courtney, S. P. (1992). Frugivory, seed predation, and insect-vertebrate interactions. Annual reviews of Entomology, 37, 377–400.
Samways, M. A. (2007). Insect conservation: a synthetic management approach. Annual Review of Entomology, 52, 465–487.
Sánchez-Garduño, C. M. (1995). Ecología de la depredación predispersión en semillas de Nectandra ambigens en Los Tuxtlas (Bachelor’s Thesis). Universidad Nacional Autónoma de México. CDMX, Mexico.
Sánchez-Roncancio, M. Y., González-Hernández, H., Johansen-Naime, R., Mojica-Guzmán, A., & Anaya-Rosales, S. (2001). Trips (Insecta: Thysanoptera) asociados a frutales de los Estados de México y Morelos, México. Folia Entomológica Mexicana, 40, 169–187.
Sarnat, E. М., Fischer, G., Guénard, B., & Economo, E. P. (2015). Introduced Pheidole of the world: taxonomy, biology and distribution. Zookeys, 543, 1–109. https://doi.org/10.3897/zookeys.543.6050
Sayers, T. D. J., Steinbauer, M. J., & Miller, R. E. (2019). Visitor or vector? The extent of rove beetle (Coleoptera: Staphylinidae) pollination and floral interactions. Arthropod-Plant Interactions, 13, 685–701. https://doi.org/10.1007/s11829-019-09698-9
Scriber, J. M. (2010). Integrating ancient patterns and
current dynamics of insect-plant interactions: Taxonomic
and geographic variation in herbivore specialization. Insect Science, 17, 471–507. https://doi.org/10.1111/j.1744-7917.
2010.01357.x
Sharma, G., Malthankar, P. A., & Mathur, V. (2021). Insect-Plant Interactions: A Multilayered Relationship. Annals of the Entomological Society of America, 114, 1–16. https://doi.org/10.1093/aesa/saaa032
Shaw, M. R., & Huddleston, T. (1991). Classification and biology of braconid wasps (Hymenoptera: Braconidae), Vol. 7. London: Royal Entomological Society of London.
Silva, E. C., Santos, B. B., & Araújo, W. S. (2018). Insect gall occurrence in savanna and forest remnant sites of Hidrolândia, GO, Brazil Central. Papéis Avulsos de Zoologia, 58, e20185804. https://doi.org/10.11606/1807-0205/2018.58.04
Souza, T. S., Fernandes, V. J., Aguiar-Menezes, E. L., Resende, A. L. S., Pereira, T. P. L., Gazal, V. S. et al. (2020). Larvae of Megaselia Róndani, 1856 (Diptera: Phoridae) as parasitoid of Sphingidae (Lepidoptera) and its frugivory on avocado. Entomological Communications, 2, ec02020. https://doi.org/10.37486/2675-1305.ec02020
Specht, A., Iltchenco, J., Fronza, E., Roque-Specht, V., Luz, P., & Montezzano, D. (2014). Biological aspects of Tiracola grandirena (Herrich-Schäffer, 1868) (Lepidoptera: Noctuidae): a polyphagous armyworm. Brazilian Journal of Biology, 74, 205–211. https://doi.org/10.1590/1519-6984.12212
Stiles, E. W. (1993). The influence of pulp lipids on fruit preference by birds. Vegetatio, 107, 227–235.
Suh, S. J., & Kwon, Y. J. (2016). First finding of a quarantine pest, Atherigona (Acritochaeta) orientalis Schiner (Diptera: Muscidae), in Korea. Entomological Research, 46, 185–189. https://doi.org/10.1111/1748-5967.12161
Szentesi, Á., & Jermy, T. (1990). The Role of experience in host plant choice by phytophagous insects. In E. A. Bernays (Ed.), Insect-plant interactions (pp. 39–74). Boston: CRC Press.
Thomas, M. C. (1988). A revision of the New World Species of Cryptolestes Ganglbauer (Coleoptera: Cucujidae: Laemophloeinae). Insecta Mundi, 2, 43–65.
Thomas, M. C. (2002). Descriptions of four new species of Cryptolestes Ganglbauer , with a revised key to the New World species and notes on other species (Coleoptera: Laemophloeidae). Insecta Mundi, 16, 147–155.
Thomas, M. C., & O’Brien, C. W. (1999). On the genus Anchonus Schonherr in Florida (Coleoptera: Curculionidae). Insecta Mundi, 13, 229–233.
Thompson, J. N., Cunningham, B. M., Segraves, K. A., Althoff, D. M., & Wagner, D. (1997). Plant polyploidy and insect/plant interactions. American Naturalist, 150, 730–743. https://doi.org/10.1086/286091
Thrips Wiki Contributors. (2020). Thrips Wiki. Retrieved 11 february, 2020 from: https://thrips.info/wiki
Tokumoto, Y., Matsushita, M., Kishimoto-Yamada, K., Nikkeshi, A., Isogimi, T., & Nakagawa, M. (2019). Floral visitors and reproductive success in two sequentially flowering Lindera shrubs (Lauraceae) of central Japan. Journal of Forest Research, 24, 42–51. https://doi.org/10.1080/13416979.2018.1542764
Tooker, J. F., Hauser, M., & Hanks, L. M. (2006). Floral host plants of Syrphidae and Tachinidae (Diptera) of Central Illinois. Annals of the Entomological Society of America, 99, 96–112. https://doi.org/10.1603/0013-8746(2006)099[0096:FHPOSA]2.0.CO;2
Trofimov, D., Rudolph, B., & Rohwer, J. G. (2016). Phylogenetic study of the genus Nectandra (Lauraceae), and reinstatement of Damburneya. Taxon, 65, 980–996. https://doi.org/10.12705/655.3
Tscharntke, T., Steffan-Dewenter, I., Kruess, A., & Thies, C. (2002). Characteristics of insect populations on habitat fragments: A mini review. Ecological Research, 17, 229–239. https://doi.org/10.1046/j.1440-1703.2002.00482.x
Uchoa, M. A. (2012). Fruit flies (Diptera: Tephritoidea): biology, host plants, natural enemies, and the implications to their natu-
ral control. In S. Soloneski (Ed.), Integrated pest management and pest control – current and future tactics (pp. 271–300). London: IntechOpen. https://doi.org/10.5772/31613
Valadão, H., Proença, C. E. B., Kuhlmann, M. P., Harris, S. A., & Tidon, R. (2019). Fruit-breeding drosophilids (Diptera) in the Neotropics: playing the field and specialising in generalism? Ecological Entomology, 44, 721–737. https://doi.org/10.1111/een.12769
van der Werff, H., & Richter, H. G. (1996). Toward an improved classification of Lauraceae. Annals of the Missouri Botanical Garden, 83, 409–418. https://doi.org/10.2307/2399870
Van Klinken, R. D., & Walter, G. H. (2001). Larval hosts of
australian Drosophilidae (Diptera): A field survey in sub-
tropical and tropical Australia. Australian Journal of
Entomology, 40, 163–179. https://doi.org/10.1046/j.1440-
6055.2001.00221.x
Van Nouhuys, S. (2005). Effects of habitat fragmentation at different trophic levels in insect communities. Annales Zoologici Fennici, 42, 433–447.
Varela-Hernández, F., & Riquelme, F. (2021). A new ant species of the genus Pheidole Westwood, 18391 from miocene mexican amber. Southwestern Entomologist, 46, 75–82. https://doi.org/10.3958/059.046.0107
Vásquez-Bolaños, M. (2011). Lista de especies de hormigas (Hymenoptera: Formicidae) para México. Dugesiana, 18, 95–133.
Vega-Vela, V., Muñoz-Robles, C. A., Rodríguez-Luna, E., López-Acosta, J. C., & Serna-Lagunes, R. (2018). Análisis de la fragmentación del paisaje de la Reserva de la Biosfera Los Tuxtlas, Veracruz, México. Ecosistemas y Recursos Agropecuarios, 5, 227–238. https://doi.org/10.19136/era.a5n
l4.1442
Von Thaden, J. J., Laborde, J., Guevara, S., & Mokondoko-Delgadillo, P. (2020). Dinámica de los cambios en el uso del suelo y cobertura vegetal en la Reserva de la Biosfera Los Tuxtlas (2006-2016). Revista Mexicana de Biodiversidad, 91, 1–14. https://doi.org/10.22201/ib.20078706e.2020.91.3190
Von Thaden, J. J., Laborde, J., Guevara, S., & Venegas-Barrera, C. S. (2018). Forest cover change in the Los Tuxtlas Biosphere Reserve and its future: The contribution of the 1998 protected natural area decree. Land Use Policy, 72, 443–450. https://doi.org/10.1016/j.landusepol.2017.12.040
Weber, R. G., & Connell, W. A. (1975). Stelidota geminata (Say): studies of its biology (Coleoptera: Nitidulidae). Annals of the Entomological Society of America, 68, 649–653. https://doi.org/10.1093/aesa/68.4.649
Weiss, M. J., & Williams, R. N. (1980). An annotated bibliography of the genus Stelidota Erichson (Coleoptera: Nitidulidae, Nitidulinae). Wooster, Ohio: Ohio Agricultural Research and Development Center.
Weisser, W. W., & Siemann, E. (Eds.). (2008). Insects and ecosystem function. Ecological studies 173. Berlin, Heidelberg: Springer
Wharton, R. A., Marsh, P. M., & Sharky, M. J. (1997). Manual of the New World Genera of the Family Braconidae (Hymenoptera). Washigton D.C.: International Society of Hymenopterists.
Wheeler, T. A. (2010). Chloropidae (frit flies, grass flies, eye gnats). In B. V. Brown, A. Borkent, J. M. Cumming, D. M. Wood, N. E. Woodley, & M. A. Zumbado (Eds.), Manual of Central American Diptera (pp. 1137–1153). Ottawa: NRC Research Press.
Whitehead, D. R. (1979). Recognition characters and distribution records for species of Conotrachelus (Coleoptera, Curculionidae) that damage avocado fruits in Mexico And Central America. Proceedings of the Entomological Society of Washington, 81, 105–107.
Whitfield, J. B. (1995). Annotated checklist of the Microgastrinae of North America North of Mexico (Hymenoptera: Braconidae). Journal of the Kansas Entomological Society, 68, 245–262.
Wibmer, G. J., & O’Brien, C. W. (1986). Annotated checklist of the weevils (Curculionidae sensu lato) of South America (Coleoptera, Curculionoidea). Gainesville: The American Entomological Institute.
Wild, A. L. (2007). A catalogue of the ants of Paraguay (Hymenoptera: Formicidae). Zootaxa, 1622, 1–55. https://doi.org/10.5281/zenodo.179222
Williams, R. N., Acosta, N. M., & Price, M. B. (2008). Nitidulidae (Coleoptera) found in flowers of Calycanthus floridus Linnaeus (Laurales: Calycanthaceae) in northeastern Ohio, U.S.A. Entomological News, 119, 397–402. https://doi.org/10.3157/0013-872X-119.4.397
Williams, R. N., Blackmer, J. L., Richmond, D. S., & Ellis, M. S. (1992). Nitidulidae (Coleoptera) diversity in three natural preserves in Portage county, Ohio. The Ohio Journal of Science, 92, 82–87.
Williams, R. N., Fickle, D. S., Kehat, M., Blumberg, D., & Klein, M. G. (1983). Bibliography of the Genus Carpophilus Stephens (Coleoptera: Nitidulidae). Wooster, Ohio: Ohio Agricultural Research and Development Center.
Williams, R. N., Peng, C., & Galford, J. R. (1989). Male genitalia for Stelidota (Coleoptera: Nitidulidae) Identification in America North of Mexico. The Florida Entomologist, 72, 537–540.
Williams, R. N., Weiss, M. J., Kehat, M., & Blumberg, D. (1984). The hymenopterous parasites of Nitidulidae. Phytoparasitica, 12, 53–64. https://doi.org/10.1007/BF02980798
Wilson, E. O. (1984). The relation between caste ratios and division of labor in the ant genus Pheidole (Hymenoptera: Formicidae). Behavioral Ecology and Sociobiology, 16, 89–98. https://doi.org/10.1007/BF00293108
Wilson, E. O. (2003). Pheidole in the New World. Cambridge: Harvard University Press,.
Wolda, H., & Sabrosky, C. W. (1986). Insect visitors to two forms of Aristolochia pilosa in Las Cumbres, Panama. Biotropica, 18, 295–299. https://doi.org/10.2307/2388572
Wong-Corral, F. J., Cortez-Rocha, M. O., Barbosa-Flores, J., & Bustamente-Andrade, F. (1992). Insect species infesting grain stored in rural communities in the northeast of Sonora, Mexico. Southwestern Entomologist, 17, 327–331.
Wood, D. M. (1987). Tachinidae. In J. F. McAlpine (Ed.), Manual of Nearctic Diptera, Volume 2 (pp. 1193–1270). Quebec: Research Branch Agriculture Canada.
Wood, S. L. (1982). The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae), a taxonomic monograph. Great Basin Naturalist Memoirs, 6, 1–1359.
Wood, S. L., & Bright, D. E. (1992). Hosts of Scolytidae and Platypodidae. Great Basin Naturalist Memoirs, 13, 1241–1348.
Woodley, N. E. (2009). Stratiomyidae (soldier flies). In B. V. Brown, A. Borkent, J. M. Cumming, D. M. Wood, N. E. Woodley, & M. A. Zumbado (Eds.), Manual of Central America Diptera. Volume 1 (pp. 521–550). Ottawa: NRC Research Press.
Wysoki, M., van den Berg, M. A., Ish-Am, G., Gazit, S., Peña, J. E., & Waite, G. K. (2002). Pests and pollinators of avocado. In J. E. Peña, J. L. Sharp, & M. Wysoki (Eds.), Tropical fruit pests and pollinators: biology, economic importance, natural enemies and control. (pp. 223–294). New York: CABI Publishing.
Young, A. M. (1982). Distribution patterns of insects in tropical habitats. In A. M. Young (Ed.), Population biology of tropical insects (pp. 173–216). Boston: Springer. https://doi.org/10.1007/978-1-4684-1113-3_5
Yu, D. S. K. (2016). Taxapad Ichneumonoidea. Nepean, Ontario: Catalogue of Life.
Zerova, M. D., & Fursov, V. N. (1991). The palaearctic species of Eurytoma (Hymenoptera: Eurytomidae) developing in stone fruits (Rosaceae: Prunoideae). Bulletin of Entomological Research, 81, 209–219. https://doi.org/10.1017/S00074853
00051294
Zhang, Q. H., & Aldrich, J. R. (2004). Attraction of scavenging chloropid and milichiid flies (Diptera) to metathoracic scent gland compounds of plant bugs (Heteroptera: Miridae). Environmental Entomology, 33, 12–20. https://doi.org/10.
1603/0046-225X-33.1.12
Zhang, Z., & Huang, H. (2007). Turing’s Formula Revisited. Journal of Quantitative Linguistics, 14, 222–241. https://doi.org/10.1080/09296170701514189.
Zimmerman, E. C. (1964). Anchonus Duryi in Southeastern Polynesia (Coleoptera: Curculionidae: Hylobiinae: Anchonini). Psyche: A Journal of Entomology, 71, 53–56. https://doi.org/10.1155/1964/52090