Francisco Javier Muñoz-Nolasco a, b, *, Diego Miguel Arenas-Moreno c, Daniel Cruz-Sáenz d, Fausto Roberto Méndez-de la Cruz a
a Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Zoología, Laboratorio de Herpetología, Apartado postal 70515, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
b Universidad Nacional Autónoma de México, Instituto de Ecología, Laboratorio de Interacción Planta-Atmósfera, Circuito Exterior s/n, Ciudad Universitaria, Coyoacán, 04510 Ciudad de México, Mexico
c Universidad Autónoma de Guerrero, Facultad de Ciencias Químico-Biológicas, Av. Lázaro Cárdenas s/n, Cuidad Universitaria Sur, 39086 Chilpancingo de los Bravo, Guerrero, Mexico
d Universidad de Guadalajara, Centro Universitario de Ciencias Biológicas y Agropecuarias, Centro de Estudios en Zoología, Km 15.5 Carretera Guadalajara-Nogales, Predio Las Agujas, 45200 Zapopan, Jalisco, Mexico
*Corresponding author: fjmn36@gmail.com (F.J. Muñoz-Nolasco)
Received: 14 April 2021; accepted: 16 August 2022
Abstract
The relative allocation of energy organisms devote to reproductive processes (aka reproductive effort) constitutes a central topic in life history theory. We document some aspects of the reproductive ecology and investment of 2 viviparous skinks of the genus Plestiodon from Mexico: an undescribed species previously assigned to P. brevirostris (Plestiodon sp., La Malinche, Tlaxcala, 2,700-3,000 m) and P. dugesii (2 populations, Mazamitla and Tapalpa, Jalisco; 2,400-2,500 m). We used relative litter mass (RLM) as a measure of reproductive effort and calculated a body condition index (BCI) as a proxy of the energy status of non-pregnant females, post-parturient females, and neonates. There were no differences in any of the reproductive characteristics between the 2 populations of P. dugesii. Both species lacked sexual size dimorphism, and female body size did not influence RLM, litter size nor litter mass. However, female BCI influenced RLM and, in P. dugesii, a larger litter size correlated with a lower BCI of neonates. Despite their overall similarities in reproductive investment, there seem to be differential selective pressures shaping offspring characteristics of these 2 lizard species.
Keywords: Life-history traits; Relative litter mass; Trade-off; Body condition index
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Esfuerzo reproductor en dos especies vivíparas de esquincos de cola azul (Squamata: Scincidae: Plestiodon) de México
Resumen
La asignación relativa de energía que los organismos destinan a los procesos reproductores (esfuerzo reproductor) constituye un tema central en la teoría de historias de vida. Documentamos algunos aspectos de la ecología e inversión reproductora de 2 especies de esquincos vivíparos del género Plestiodon de México, una especie no descrita previamente asignada a P. brevirostris (Plestiodon sp., La Malinche, Tlaxcala, 2,700-3,000 m) y P. dugesii (2 poblaciones, Mazamitla y Tapalpa, Jalisco, 2,400-2,500 m). Usamos la masa relativa de camada (MRC) como medida del esfuerzo reproductor y calculamos un índice de condición corporal (ICC) como aproximación del estado energético de hembras no gestantes, posparturientas y neonatos. No hubo diferencias en las características reproductoras de las 2 poblaciones de P. dugesii. Ninguna especie presentó dimorfismo sexual en la talla y la talla de la hembra no influyó sobre la MRC ni en el tamaño o en la masa de la camada. El ICC de las hembras influyó sobre la MRC y, en P. dugesii, el tamaño de camada se correlacionó negativamente con el ICC de neonatos. Pese a las semejanzas generales en inversión reproductora, parece haber presiones selectivas diferenciales que moldean las características de las crías de estas 2 especies.
Palabras clave: Características de historia de vida; Masa relativa de camada; Trade-off; Índice de condición corporal
© 2023 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
A central concept in life history theory deals with the relative allocation of energy or resources that organisms devote to reproduction to maximize the production of offspring (Hirshfield & Tinkle, 1975; Shine & Schwarzkopf, 1992). However, as the available energy is limited, this investment in reproduction (aka reproductive effort) is governed by a balance between reproductive expenditure, on the one hand, and the survival and future fecundity of the organisms, on the other (Shine & Schwarzkopf, 1992; Williams, 1966). For squamates, relative clutch/litter mass (RCM, RLM) has been widely used as a measure of female reproductive effort because most species lack parental care, and so energy allocation can be summarized in terms of the relative proportion of mass offspring production accounts for (Bastiaans et al., 2013). During reproduction nutrients that otherwise might be directed to body growth are used instead for developing eggs and embryos, thus reducing the potential of success for future reproductive events (Landwer, 1994; Shine & Schwarzkopf, 1992). Furthermore, gravid/pregnant individuals might be more susceptible to predation due to the physical burden it represents and to the increase in exposure during maternal basking (Miles et al., 2000; Swain & Jones, 2000). Factors known to influence reproductive effort in lizards include phylogeny, morphology, foraging and predator-escape strategies, habitat use, resource availability, and even parasite load (Hirshfield & Tinkle, 1975; Sorci et al., 1996; Uller & While, 2015; Vitt & Price, 1982). Moreover, sexual size dimorphism might evolve in lizards in response not only to intraspecific competition among males, but also in cases in which a larger size in females allows a higher clutch/litter size, or even as a way to reduce intersexual competition through resource partitioning (Cox et al., 2007).
Whereas selection for different levels of RCM or RLM is dependent upon adult mortality (due to the physical and physiological burden gravidity or pregnancy imposes to females), other reproductive characteristics, such as clutch/litter size and mass or egg/neonate size are shaped by the progeny mortality (Messina & Fox, 2001; Vitt & Price, 1982). Indeed, the probability of parents having at least 1 surviving offspring increases with clutch or litter size (Messina & Fox, 2001); also, it is assumed that larger offspring are under less risk of predation (Uller & Olsson, 2010). These characteristics tend to be positively correlated to body size, with larger size females producing either more eggs/neonates or fewer larger ones (Uller & While, 2015; Vitt & Price, 1982).
Whatever their reproductive strategy, investment is usually higher in viviparous than in oviparous species because of their longer gestation times and extended physical and physiological burden (Dupoué et al., 2015; Recknagel & Elmer, 2019). Because of this, viviparous lizards are usually limited to no more than 1 brood per year and have lower litter sizes, both of which translate in a reduced reproductive output (Meiri et al., 2011). However, relatively few studies on lizard reproductive investment have been conducted on viviparous species (Bastiaans et al., 2013; Recknagel & Elmer, 2019; Rodríguez-Romero et al., 2005), this in spite of the higher extinction risk they face due to climate change (Sinervo et al., 2010). Even fewer studies have investigated the relationship between pregnancy and body condition in lizards, despite their potentially serious implications in terms of costs of reproduction (Boretto et al., 2014; Itonaga et al., 2012). Given that the estimation of life-history parameters is useful to evaluate the intrinsic vulnerability of reptiles to population declines and extirpations (Govindarajulu et al., 2011), information on reproductive investment of live bearing lizards could be useful for conservation purposes.
Within the genus Plestiodon, viviparity has evolved once, in the brevirostris group (Watson et al., 2014), a clade comprised by 15 currently recognized species distributed in Mexico and part of Central America (García-Vázquez et al., 2021). Most of them are highland inhabitants and share a viviparous mode of reproduction (Goldberg, 2002; Feria-Ortiz et al., 2007; López-Hernández et al., 2008). Here, we present information on the reproductive ecology of 2 species of this group: an undescribed species previously assigned to Plestiodon brevirostris (1 population from Tlaxcala) and P. dugesii (2 populations from Jalisco). Our goals were to determine whether the studied species and populations follow different strategies of reproductive investment, the existence of sexual size dimorphism and trade-offs in reproduction, as well as to elucidate the underlying causes of these patterns.
Materials and methods
The undescribed species belongs to a taxon previously assigned to Plestiodon brevirostris, which is now known to be a species complex comprised of 4 lineages, 1 distributed in the states of Tlaxcala and Puebla; a second one, in Guerrero, and 2 more from Oaxaca (Pavón-Vázquez et al., 2018). However, none of these taxa have been so far properly described. Given that the type locality of P. brevirostris is indicated by Günther (1860) as “Oaxaca (Mexico)”, and as the studied population belongs to the Tlaxcala-Puebla lineage, we will refer hereafter to this taxon as Plestiodon sp.
Plestiodon sp. is a skink up to 75.5 mm snout-vent length (SVL) (see supplementary data 5 in Pavón-Vázquez et al. [2018]). It has an asynchronous fall reproductive cycle, with vitellogenesis occurring between June and October, and ovulation and fertilization in November, so that gestation extends from then until spring (Chávez-López, 2012). The population of Plestiodon sp. studied was from La Malinche, a 4,461 m dormant volcano located in the states of Tlaxcala and Puebla, in central eastern Mexico. Fieldwork at this locality was carried out on the eastern slope of the volcano (19°14’05” N, 97°56’24” W; 2,700 to 3,000 m elevation) in an abrupt transition zone between the Pinus–Quercus forest and induced grasslands. Climate at this altitudinal level is temperate sub-humid with day temperatures ranging from 12 to 18 °C throughout the year (Villers-Ruiz, 2006). Despite its status as a protected natural area, La Malinche has been heavily affected in the last years by land use changes, illegal logging, and forest fires (Piñaza-Soto & Hernández-Hernández, 2011), and more recently by the subterranean channelling of some natural streams, which together have eroded the microhabitat opportunities for Plestiodon sp.
Plestiodon dugesii is a skink of up to 69 mm SVL, distributed in part of the states of Guanajuato, Michoacán and Jalisco in western Mexico, at elevations above 2,000 m. (Dixon, 1969; Flores-Villela & Santos-Barrera, 2007). Information about the biology of this species is scarce. However, it is under special protection according to the Mexican law NOM-059-SEMARNAT-2010 (Semarnat, 2019), and is included as vulnerable in the Red List of the International Union for Conservation of Nature (IUCN) (Flores-Villela & Santos-Barrera, 2007). We sampled 2 populations of this species, both from the state of Jalisco. The first one was from the surroundings of the cliff El Tecolote, in the municipality of Mazamitla (19°54’43” N, 102°59’30” W; 2,400 m elevation); the second population was from the locality of Ferrería de Tula, in the municipality of Tapalpa (20°03’46” N, 103°42’39” W; 2,500 m elevation). Both study sites have a temperate sub-humid climate with summer rains (Ruiz-Corral et al., 2012), with a mean daytime temperature of 19.6 °C (Mazamitla) and 16 °C (Tapalpa). Although vegetation type at these sites corresponds to pine-oak forest, we primarily encountered lizards using rotten logs at Mazamitla and rocks at Tapalpa, even though both microhabitat types were present at both localities.
We conducted samplings between March and April 2013 and 2014; however, for Plestiodon sp. we only obtained data for 2014. Fieldwork consisted in the active search and capture by hand of lizards in potential microhabitats, such as leaf litter and under rotten logs and rocks. We focused on pregnant females, which we recognized by noticing abdominal distension and through palpation (Shine, 1980), but during the same samplings we also collected non-pregnant females and males for in situ measurements to make comparisons on body size dimorphism (see below). We sexed individuals through manual hemipenial eversion.
We temporarily placed caught pregnant females in separate plastic containers (20 × 12 × 12.5 cm) to which we drilled holes for aeration. Then we transported the lizards to nearby laboratory facilities to conduct the captivity period: the Laboratorio de Herpetología II of the Instituto de Biología of the Universidad Nacional Autónoma de México (Mexico City), in the case of Plestiodon sp., and the Centro de Estudios en Zoología of the Universidad de Guadalajara (Zapopan, Jalisco) in the case of P. dugesii. Once there, we housed them in larger individual plastic enclosures (43.18 × 16.51 × 28.3 cm), with peat moss as substrate and pine bark as shelter and provided them with water and food (mealworm larvae and domestic crickets) ad libitum. Based on the available information of the thermal biology of other Plestiodon species (Youssef et al., 2008), the enclosures were kept at 25-30 °C and under natural photoperiod. For each female, we recorded SVL and axilla-groin length (AGL), as these morphometric variables are known to influence litter or offspring characteristics in lizards (Griffith, 1994; Recknagel & Elmer, 2019), using a digital calliper (± 0.01 mm; AutoTec®). We checked, at least twice a day, every container until parturition, and immediately weighed the female using a digital scale (± 0.01 g; Grobet®) and proceeded to count, weigh, and measure each of the neonates (using the same calliper and scale).
The field and experimental protocols were reviewed and approved by the Dirección General de Vida Silvestre (DGVS), Secretaría de Medio Ambiente y Recursos Naturales (Semarnat) through the scientific research permit FAUT-0074, granted to Fausto R. Méndez-de la Cruz. After completion of data collection in laboratory, we released all the females, together with their offspring, at the exact sites of capture.
To calculate RLM, we divided litter mass (excluding amniotic fluids and extraembryonic membranes) by post-parturition female mass (PPFM), as other traditional methods of calculation that include the female mass in the numerator and denominator, such as that proposed by Tinkle (1972; clutch or litter mass/mass of the gravid or pregnant female), are prone to statistical bias (Bastiaans et al., 2013; Rodríguez-Romero et al., 2005; Shine, 1980). Previous studies have used body condition estimates to assess the effect of the energy status of pregnant females on their offspring (Recknagel & Elmer, 2019). Here we calculated the body condition index of non-pregnant females, post-parturient females, and neonates, for which we used the scaled mass index proposed by Peig and Green (2009), according to the following equation:
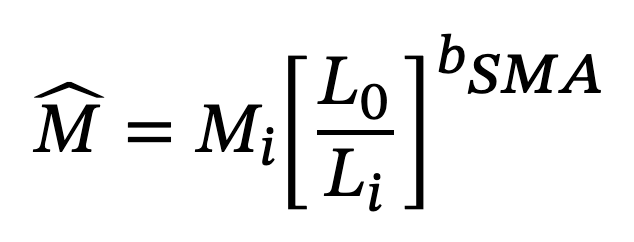
where Mi and Li were defined as body mass (g) and SVL (mm) of the individual i, respectively; L0 was the mean value of SVL of the species (using mean adult SVL for non-pregnant and post-parturient females and mean neonate SVL for the offspring; data from this study); and bSMA, the scaling exponent obtained through dividing the slope of an ordinary least squares linear regression of M on L by the Pearson’s correlation coefficient. This index is size-independent and has been shown to be more suitable than other body condition indexes based on ratios or on residuals from regressions (Labocha et al., 2014). We assumed that lower values of body condition index (BCI) in post-parturient females or offspring would indicate a detrimental effect of pregnancy or a reduced capacity to allocate enough resources to developing embryos, respectively (Bonnet et al., 2002; Recknagel & Elmer, 2019).
Before any statistical procedure, we tested each variable for normality and homoscedasticity through Kolmogorov-Smirnov and Levene’s tests, respectively, considering an α level = 0.05 (Zar, 2010). When data did not satisfy the statistical assumptions for parametric tests, we performed the non-parametric equivalent analyses (Dytham, 2011). In the case of P. dugesii, data came from 2 different populations and sampling years, so we first compared them through Student’s t tests and Mann-Whitney U tests to discard intraspecific or temporal effects on the female and litter characteristics. As sexual dimorphism in body size might reveal the existence of fecundity selection (Cox et al., 2007), we compared SVL between males and females through Student’s t tests and Mann-Whitney U tests. Also, we compared the BCI of post-parturient versus non-pregnant females of each species through Mann-Whitney U tests.
We analysed the litter and neonate characteristics through a series of general linear models. First, to compare RLM and litter size between the 2 species, we performed analyses of covariance (ANCOVA) with female SVL, AGL, and BCI as covariables, discarding non-significant factors in a stepwise procedure (Bastiaans et al., 2013). Secondly, we performed Pearson’s linear correlations to assess the relation between litter characteristics (litter size, litter mass, and RLM) with the female characteristics (SVL, AGL, PPFM, and BCI). Finally, to determine the existence of trade-offs in offspring size versus number, we assessed the relation between litter size and mean SVL, mean mass, and mean BCI of neonates with female characteristics (SVL, AGL, PPFM, and BCI) as covariables, through an ANCOVA (Bastiaans et al., 2013). We carried out the statistical tests in SPSS 15.0.1 (SPSS Inc., 2006) and SigmaPlot version 11.0 (Systat Software, 2008).
Results
We collected a total of 25 individuals of Plestiodon sp. (11 males, 7 non-pregnant females, 7 pregnant females) and 54 of P. dugesii, of which 33 were collected in 2013 (14 males, 7 non-pregnant females, 12 pregnant females) and 21 in 2014 (5 males, 7 non-pregnant females, 9 pregnant females). In laboratory, parturition of Plestiodon sp. occurred from mid-April through the end of May; in the case of P. dugesii, births occurred only during May. Minimum size (SVL) of pregnant females was 55.01 mm in Plestiodon sp. and 55.79 mm in P. dugesii, while the size of non-pregnant females ranged from 44.04 to 63.39 mm in Plestiodon sp. and 42.04 to 53.61 mm in P. dugesii. As there were no statistical differences in SVL, litter size, RLM, and BCI of P. dugesii by population or year, we pooled together the data: SVL by population: (males: t = -0.501, df = 13, p = 0.625; females U = 48.000, p = 0.448); litter size by population: (U = 24, p = 0.100); RLM by population: (U = 39, p = 0.669); RLM by year: (Mazamitla: t = 1.258, df = 4, p = 0.277; Tapalpa: t = 1.171, df = 13, p = 0.263); BCI of post-parturient females by population: (t = -0.009, df = 19, p = 0.993). Reproductive characteristics of the 2 species are summarized in Table 1.
Table 1
Reproductive characteristics of Plestiodon sp. (La Malinche, Tlaxcala; n = 7) and P. dugesii (Mazamitla and Tapalpa, Jalisco; n = 21). SVL = Snout-vent length; BCI = scaled mass body condition index (Peig & Green, 2009); RLM = relative litter mass. Data is shown as mean ± SD (minimum value-maximum value).
| Characteristic | Plestiodon sp. | P. dugesii |
| Female SVL (mm) | 62.347 ± 4.569 (51.01-68.58) | 61.268 ± 4.506 (50.56-72.2) |
| Post-parturition female mass (g) | 3.993 ± 0.983 (3.06-5.81) | 3.479 ± 0.594 (2.50-5.05) |
| Post-parturition female BCI | 3.914 ± 0.928 (3.02-5.63) | 3.422 ± 0.553 (2.49-4.88) |
| Litter size | 2.5 ± 1.4, (1-4) | 3.5 ± 1.2 (1-5) |
| Litter mass (g) | 0.864 ± 0.508 (0.33-1.68) | 0.997 ± 0.374 (0.25-1.7) |
| RLM | 0.231 ± 0.142 (0.05-0.45) | 0.292 ± 0.111 (0.08-0.5) |
| Mean neonate SVL (mm) | 26.192 ± 1.248 (24.14-28.1) | 24.449 ± 0.838 (22.88-26.11) |
| Mean neonate mass (g) | 0.336 ± 0.045 (0.29-0.42) | 0.292 ± 0.032 (0.22-0.34) |
| Mean neonate BCI | 0.336 ± 0.044 (0.28-0.41) | 0.292 ± 0.032 (0.22-0.34) |
Table 2
Results of the analysis of covariance (ANCOVA) for the effect of body condition index (BCI) of post-parturient females on the relative litter mass of Plestiodon sp. and P. dugesii.
| Source | Sum of squares | Degrees of freedom | Quadratic mean | F | p |
| Corrected model | 0.82 | 2 | 0.041 | 3.357 | 0.051 |
| Intercept | 0.231 | 1 | 0.231 | 18.921 | < 0.001 |
| BCI | 0.063 | 1 | 0.063 | 5.133 | 0.032 |
| Species | 0.003 | 1 | 0.003 | 0.223 | 0.641 |
| Error | 0.305 | 25 | 0.012 |
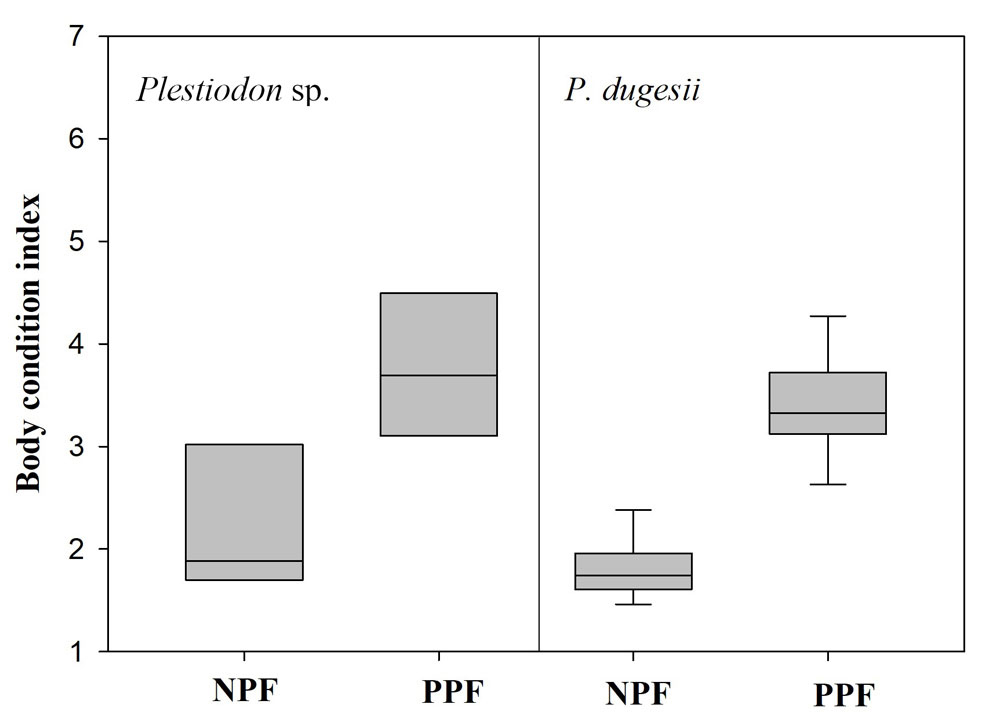
Males of Plestiodon sp. (SVL x = 61.486 mm, SD = 4.462 mm, n = 11) were larger than males of P. dugesii (SVL x = 52.983 mm, SD = 5.503 mm, n = 19) (t = 4.353, df = 28, p < 0.001); however, there were no differences in SVL between females of the 2 species (Plestiodon sp.: x = 57.151 mm, SD = 7.660 mm, n = 14; P. dugesii: x = 56.536 mm, SD = 15.282 mm, n = 35; U = 210, p = 0.445). We found no evidence of sexual size dimorphism in either of the 2 species (Plestiodon sp.: t = 1.664, df = 24, p = 0.110; P. dugesii: t = 0.333, df = 53, p = 0.097). Post-parturient females had a higher BCI than non-pregnant females in both species (Plestiodon sp.: U = 7, p = 0.026; P. dugesii: U = 1, p < 0.001; Fig. 1).
Regarding the comparison of litter size, there were no differences by species (F = 2.529, df = 1, p = 0.124) after discarding SVL, AGL, and female BCI, whose effects were non-significant (SVL: F = 0.088, df = 1, p = 0.769; AGL: F = 1.414, df = 1, p = 0.247; female BCI: F = 3.135, df = 1, p = 0.090). In the case of RLM, the effects of female SVL and AGL were also not significant (SVL: F = 0.755, df = 1, p = 0.394; AGL: F = 0.158, df = 1, p = 0.695), so we discarded them. However, in the final model female BCI had a significant effect on RLM (F = 5.133, df = 1, p = 0.032), with higher levels of BCI resulting in lower RLM, although without statistical differences between species (F = 0.223, df = 1, p = 0.641) (Table 2).
Table 3
| Source | Sum
of squares |
Degrees
of freedom |
Quadratic mean | F | p |
| Corrected model | 20.115 | 5 | 4.023 | 4.661 | 0.007 |
| Intercept | 15.715 | 1 | 15.715 | 18.207 | < 0.001 |
| SVL | 3.688 | 1 | 3.688 | 4.273 | 0.053 |
| AGL | 0.028 | 1 | 0.028 | 0.033 | 0.858 |
| PPFM | 0.528 | 1 | 0.528 | 0.611 | 0.444 |
| BCI | 0.023 | 1 | 0.023 | 0.027 | 0.871 |
| Species | 7.449 | 1 | 7.449 | 8.63 | 0.009 |
| Error | 15.536 | 18 | 0.863 |
Results of the analysis of covariance (ANCOVA) for the effects of female body characteristics on neonate mean snout-vent length of Plestiodon sp. and P. dugesii. SVL = Snout-vent length, AGL = axilla-groin length, PPFM = postpartum female mass, BCI = body condition index.
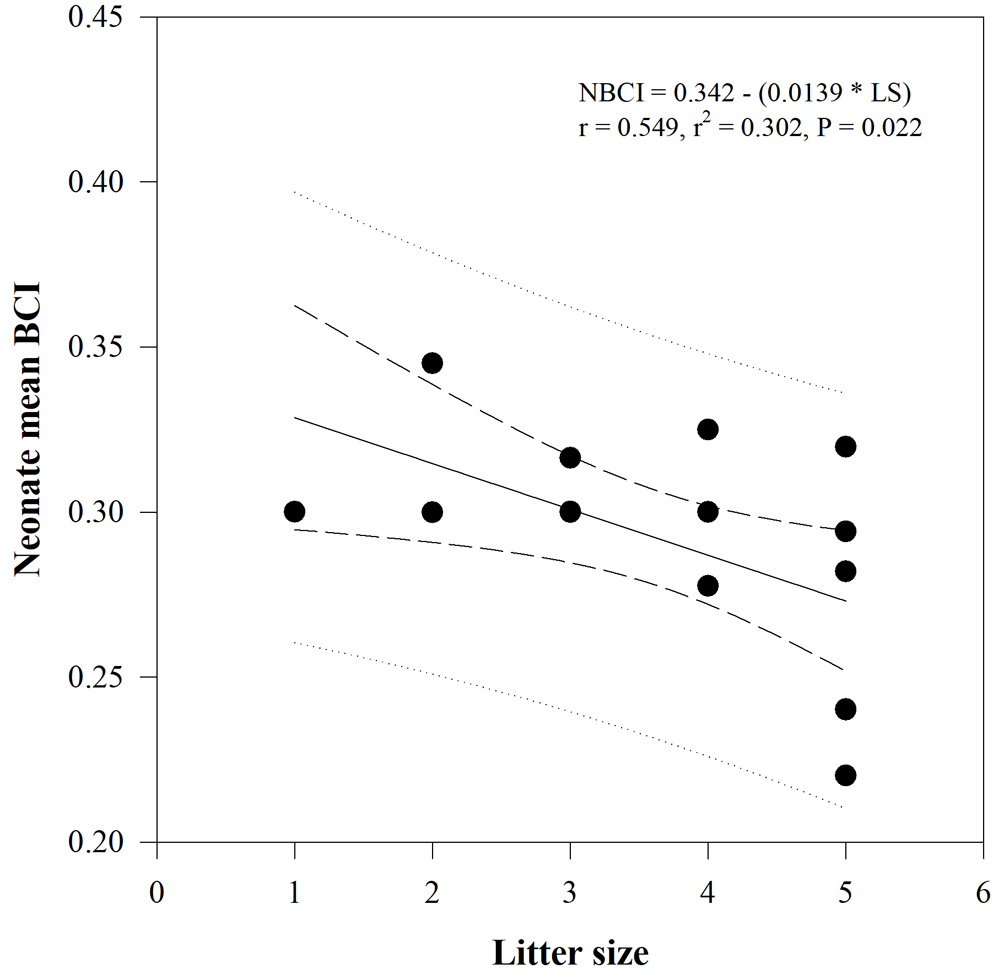
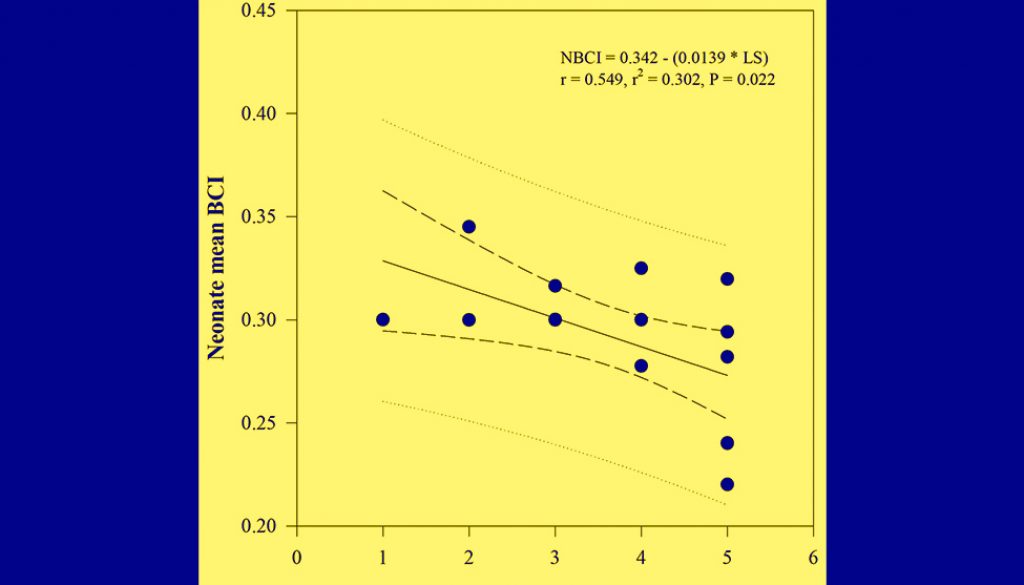
Female SVL has a marginally significant positive effect on neonate mean SVL (F = 4.273, df = 1, p = 0.053) but not female AGL, PPFM, and female BCI; after correcting for their effects, there were interspecific differences in neonate mean SVL, indicating a larger SVL in neonates of Plestiodon sp. than in neonates of P. dugesii (F = 8.630, df = 1, p = 0.009). There were no significant differences in neonate mean mass or BCI by species after accounting for female morphometric measurements (Table 3). Similarly, there were no differences in mean neonate SVL (F = 0.023, df = 1, p = 0.543), mass (F = 0.005, df = 1, p = 0.187), or BCI (F = 1.987, df = 1, p = 0.186) by population in P. dugesii. For Plestiodon sp. there were no significant relationships between any of the reproductive and female characteristics; for P. dugesii, on the other hand, we found negative relationships between litter size and neonate mean mass (r = -0.548, t = -2.541, p = 0.023) and neonate mean BCI (r = 0.549, t = -2.544, p = 0.022; Fig. 2).
Discussion
In lizards, sexual size dimorphism has evolved in cases in which: 1) larger size in males increases mating success through an advantage in intrasexual competition, 2) larger size in females determines a higher number of offspring (i.e., clutch or litter size), or 3) differences in body size allow resource partitioning to reduce intersexual competition (Cox et al., 2007). Some oviparous species of the genus Plestiodon, including P. elegans and P. laticeps, exhibit male-biased sexual dimorphism (Vitt & Cooper Jr., 1985; Wei-Guo & Xiang, 2001). Lack of sexual size dimorphism (i.e., monomorphism) in Plestiodon sp. and P. dugesii might indicate that neither sexual nor fecundity selection or differential use of resources have driven the evolution of size in these species. However, it is known that sexual size monomorphism can occur because of the mutual cancellation of sexual and fecundity selection, a pattern that has been found in other scincomorph lizards (e.g., Dubey et al., 2011; Ma et al., 2018). Whatever the cause of this phenomenon, it is likely to be shared by other members of the Plestiodon brevirostris group. This is also consistent with the fact that litter size and mass were not related to female SVL in these species.
Species of the P. brevirostris group exhibit variable patterns of maternal SVL in relation to litter characteristics. For instance, in another population of Plestiodon sp. from Puebla litter size seems not to be influenced by female SVL (Chávez-López, 2012), but in P. copei there is a positive relationship between these traits (Guillette, 1983). Furthermore, Chávez-López (2012) found, for the referred population of Plestiodon sp. from Puebla, a positive relationship between litter mass and female SVL, a pattern that we did not find for the same species. However, this may be because of our small sample size for Plestiodon sp. Mean values of RLM in Plestiodon sp. and P. dugesii are consistent with those reported for other species of skinks (Qualls & Shine, 1997; Shine, 1980; Vitt & Price, 1982), although in some females RLM accounted for about 50% of the PPFM, a high level of reproductive effort considering the active foraging strategy and elongated body plan of these lizards (Griffith, 1994; Vitt & Price, 1982).
Variation in RLM in both species was influenced by BCI but not by other female characteristics, such as SVL and AGL, which supports its usefulness as an indicator of the energy status of the organisms in a reproductive context. Although in many squamates female body condition tends to decrease because of energy expenditure during reproduction (e.g., Bonnet et al., 2002), there are cases in which reproductive investment does not impair maternal body condition (Itonaga et al., 2012). In Niveoscincus ocellatus, another viviparous skink, abdominal fat body mass increases with pregnancy (Wapstra & Swain, 2001). However, while N. ocellatus is pregnant in summer, when food resources are more abundant, members of the Plestiodon brevirostris group have a fall reproductive cycle, and therefore gestation occurs during a period of limited food availability (Chávez-López, 2012; Guillette, 1983). Furthermore, the narrowing of the activity periods in winter due to lower temperatures may cause constraints both in foraging and digestion efficiency of females (Hare & Cree, 2016; Méndez-de la Cruz et al., 2015). Given the above, it seems unlikely that the higher BCI of post-parturient Plestiodon sp. and P. dugesii is due to an ability to store fat during pregnancy (which would imply increasing either the quantity or quality of the food consumed). Another, more reasonable, explanation would be that the onset of reproduction in females of these species is conditioned to some degree by their body condition, for example, to mobilize the required maternal reserves to sustain vitellogenesis (Ramírez-Pinilla et al., 2015). Nonetheless, we cannot rule out the possibility that the higher BCI of postparturient females is an artifact of the ad libitum feeding in captivity, so these results must be taken cautiously.
Numerous studies have addressed the existence of trade-offs between clutch/litter size and offspring size or mass (e.g., Bastiaans et al., 2013; Olsson & Shine, 1997; Recknagel & Elmer, 2019). The negative relationship between litter size and mean neonate mass and BCI in P. dugesii, although moderate, reveals that a larger litter size may reduce in some degree the probability of offspring survival due to an impairment in body condition (Uller & Olsson, 2010). In Plestiodon sp. on the contrary, a larger litter size does not imply a reduction in neonate mass or BCI, and neonates of this species had a larger SVL than in P. dugesii. The above suggests differential pressures on offspring survival between the 2 species, which might be related to the environmental conditions of their habitat (Díaz et al., 2005). For instance, while in both localities of P. dugesii habitat provides plenty of shelter places and a reasonably high offer of food resources, the habitat at the locality of Plestiodon sp. is more degraded, and therefore offspring might be more prone to predation and less able to find prey.
Meiri et al. (2011) proposed to analyse several components of the life histories of lizards in terms of their reproductive output and biomass production (i.e., productivity). They defined “productivity” as the total mass of offspring produced in a year (hatchling or neonate mass × clutch or litter size × number of clutches or litters per year), and “specific productivity” as productivity divided by female mass. Since it is to be expected Plestiodon sp. and P. dugesii to produce 1 litter per year, as occurs with other members of the brevirostris group (Chávez-López, 2012; Guillette, 1983), productivity of Plestiodon sp. would be 0.85 g/year with a specific productivity of 0.21. In P. dugesii productivity would be 1.001 g/year and specific productivity 0.28. These values are consistent with those reported by Meiri et al. (2011) for other viviparous lizards from temperate environments. Furthermore, the authors found that both measures of biomass production tend to decrease in colder environments because mean annual temperatures constrain the duration of the reproductive season. The latter point is also consistent with the lower values estimated for Plestiodon sp., as this species lives at higher altitude and in a more thermally challenging environment.
Coupled with environmental factors, evolutionary history of lizards might determine patterns of reproductive investment (Uller & While, 2015). Plestiodon sp. and P. dugesii diverged about 10 Ma ago (Bryson et al., 2017), possibly a short time to produce substantial differences in their reproductive ecology, but enough to influence neonate characteristics. It remains to be assessed the extent to which certain evolutionary or environmental factors are responsible for shaping these patterns in species of the Plestiodon brevirostris group. Another aspect that deserves to be addressed is the evolution of parity mode in Plestiodon. Currently, it is considered that viviparity arose only once within the genus, in the P. brevirostris group (Watson et al., 2014); however, recent phylogenies have placed P. sumichrasti, a Central American, lowland oviparous species (Miller, 1997), nested within this clade (Brandley et al., 2012; Bryson et al., 2017). Whether this represents 2 distinct origins of viviparity in the genus or an evolutionary regression to oviparity, in P. sumichrasti has not yet been tackled.
In summary, our results indicate that in spite of the overall similarities in the reproductive characteristics of Plestiodon sp. and P. dugesii, differences in their reproductive investment, and particularly in the relationship between offspring size and number, exist between these 2 species. Further studies are needed to deepen into the reproductive biology of other viviparous members of the genus Plestiodon, which in comparison to their oviparous congeners have received less attention.
Acknowledgments
FJMN and DMAM thank the Posgrado en Ciencias Biológicas of the Universidad Nacional Autónoma de México. We thank Oscar J. Rodríguez Ruvalcaba, Isaac E. Terrones Ferreiro, Santiago Cortés Vázquez, Daniel Pérez Lozano, Jonathan E. Beas Zepeda, Jonathan Justo Prieto, Iván A. Coffeen Calvario, Miguel Veliz Pérez, and Daniel Muñoz Nolasco for support in the field. We are especially grateful to Oscar Alberto Rodríguez Aguilar and Luis Alberto Rodríguez Miranda for valuable assistance in field and laboratory. To Margarita Martínez Gómez, from the Centro Tlaxcala de Biología de la Conducta (CTBC), for allowing us the use of the facilities of the Estación Científica La Malinche, and to Luis Oceguera Arana and Santiago Cortés Vázquez, for the attentions given us during fieldwork at the Mazamitla and Tapalpa study sites. We also thank Claudia Uribe Mú, for providing helpful advice during the development of this research. Finally, we thank two anonymous reviewers for providing valuable and insightful comments and suggestions that greatly improved this manuscript. Organisms were collected under permit number FAUT-0074, granted to Fausto R. Méndez de la Cruz by the Dirección General de Vida Silvestre (DGVS) of the Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT).
References
Bastiaans, E., Méndez-de la Cruz, F., Rodríguez Hernández, K., Flores Aguirre, C., & Sinervo, B. (2013). Female reproductive investment in the mesquite lizard (Sceloporus grammicus) species complex (Squamata: Phrynosomatidae). Southwestern Naturalist, 58, 335–343. https://doi.org/10.1894/0038-4909-58.3.335
Bonnet, X., Lourdais, O., Shine, R., & Naulleau, G. (2002). Reproduction in a typical capital breeder: costs, currencies, and complications in the aspic viper. Ecology, 83, 2124–2135. https://doi.org/10.1890/0012-9658(2002)083[2124:RIATCB]2.0.CO;2
Boretto, J. M., Cabezas, F., Kubisch, E. L., Sinervo, B., & Ibarguengoytía, N. (2014). Changes in female reproduction and body condition in an endemic lizard, Phymaturus spectabilis, following the Puyehue volcanic ashfall event. Herpetological Conservation & Biology, 9, 181–191.
Brandley, M. C., Ota, H., Hikida, T., Nieto-Montes de Oca, A., Feria-Ortiz, M., Guo, X. et al. (2012). The phylogenetic systematics of blue-tailed skinks (Plestiodon) and the family Scincidae. Zoological Journal of the Linnean Society, 165, 163–189. https://doi.org/10.1111/j.1096-3642.2011.00801.x
Bryson Jr., R. W., Linkem, C. W., Pavón-Vázquez, C. J., Nieto-Montes de Oca, A., Klicka, J., & McCormack, J. E. (2017). A phylogenomic perspective on the biogeography of skinks in the Plestiodon brevirostris group inferred from target enrichment of ultraconserved elements. Journal of Biogeography, 44, 2033–2044. https://doi.org/10.1111/jbi.12989
Chávez-López, L. (2012). Biología reproductiva de Plestiodon brevirostris (Squamata: Scincidae) en el centro-este del estado de Puebla (Bachelor’s Thesis). Facultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México, México D.F.
Cox, R. M., Butler, M. A., & John-Alder, H. B. (2007). The evolution of sexual size dimorphism in reptiles. In D. J. Fairbairn, W. U. Blanckenhorn, & T. Székely (Eds.), Sex, size and gender roles: evolutionary studies of sexual size dimorphism (pp. 38–49). Oxford, Michigan: Oxford University Press.
Díaz, J. A., Pérez-Tris, J., Telleria, J. L., Carbonell, R., & Santos, T. (2005). Reproductive investment of a lacertid lizard in fragmented habitat. Conservation Biology, 19, 1578–1585. https://doi.org/10.1111/j.1523-1739.2005.00197.x
Dixon, J. R. (1969). Taxonomic review of the Mexican skinks of the Eumeces brevirostris group. Los Angeles County Museum Contributions in Science, 168, 1–29.
Dubey, S., Chevalley, M., & Shine, R. (2011). Sexual dimorphism and sexual selection in a montane scincid lizard (Eulamprus leuraensis). Austral Ecology, 36, 68–75. https://doi.org/10.1111/j.1442-9993.2010.02119.x
Dupoué, A., Brischoux, F., Angelier, F., DeNardo, D. F., Wright, C. D., & Lourdais, O. (2015). Intergenerational trade-off for water may induce a mother–offspring conflict in favour of embryos in a viviparous snake. Functional Ecology, 29, 414–422. https://doi.org/10.1111/1365-2435.12349
Dytham, C. (2011). Choosing and using statistics: a biologist’s guide. 3rd edition, Oxfordshire, Oxford: Wiley-Blackwell.
Feria-Ortiz, M., García-Vázquez, U. O., & Aguilar-López, J. L. (2007). Plestiodon brevirostris (Short-nosed Skink). Reproduction. Herpetological Review, 38, 81–82.
Flores-Villela, O., & Santos-Barrera, G. (2007). Plestiodon dugesii. The IUCN Red List of Threatened Species 2007: e.T64225A12755550. https://dx.doi.org/10.2305/IUCN.UK.2007.RLTS.T64225A12755550.en. Downloaded on 14 April 2021.
García-Vázquez, U. O., Pavón-Vázquez, C. J., Feria-Ortiz, M., & Montes-de Oca, A. (2021). A new species of blue-tailed skink (Scincidae: Plestiodon) from the Sierra Madre del Sur, Mexico. Herpetologica, 77, 85–93. https://doi.org/10.1655/0018-0831-77.1.85
Goldberg, S. R. (2002). Eumeces brevirostris (Short Nosed Skink). Reproduction. Herpetological Review, 33, 134.
Govindarajulu, P., Isaac, L. A., Engelstoft, C., & Ovaska, K. (2011). Relevance of life-history parameter estimation to conservation listing: case of the Sharp-Tailed snake (Contia tenuis). Journal of Herpetology, 45, 300–307. https://doi.org/10.1670/10-086.1
Griffith, H. (1994). Body elongation and decreased reproductive output within a restricted clade of lizards (Reptilia: Scincidae). Journal of Zoology, 233, 541–550. https://doi.org/10.1111/j.1469-7998.1994.tb05364.x
Guillette Jr., L. J. (1983). Notes concerning reproduction of the montane skink, Eumeces copei. Journal of Herpetology, 17, 144–148. https://doi.org/10.2307/1563455
Günther, A. (1860). On new reptiles and fishes from Mexico. Proceedings of the Zoological Society of London, 28, 316–322.
Hare, K. M., & Cree, A. (2016) Thermal and metabolic physiology of New Zealand lizards. In D. Chapple (Eds.), New Zealand lizards (pp. 239–267). Cham, Zug: Springer.
Hirshfield, M. F., & Tinkle, D. W. (1975). Natural selection and the evolution of reproductive effort. Proceedings of the National Academy of Sciences, 72, 2227–2231. https://doi.org/10.1073/pnas.72.6.2227
Itonaga, K., Edwards, A., Wapstra, E., & Jones, S. M. (2012). Interpopulational variation in costs of reproduction related to pregnancy in a viviparous lizard. Ethology, Ecology & Evolution, 24, 367–376. https://doi.org/10.1080/03949370.2012.702686
Labocha, M. K., Schutz, H., & Hayes, J. P. (2014). Which condition index is best? Oikos, 123, 111–119. https://doi.org/10.1111/j.1600-0706.2013.00755.x
Landwer, A. J. (1994). Manipulation of egg production reveals costs of reproduction in the tree lizard (Urosaurus ornatus). Oecologia, 100, 243–249. https://doi.org/10.1007/BF00316951
López-Hernández, I. D., Feria-Ortiz, M., & Martínez-Coronel, M. (2008). Tamaño de camada en Plestiodon brevirostris. Boletín de la Sociedad Herpetológica Mexicana, 16, 15–17.
Ma, L., Pei, J., Zhou, C., Du, Y., Ji, X., & Shen, W. (2018). Sexual dimorphism, female reproductive characteristics and egg incubation in an oviparous forest skink (Sphenomorphus incognitus) from South China. Asian Herpetological Research, 9, 119–128. https://doi.org/10.16373/j.cnki.ahr.180011
Méndez-de la Cruz, F. R., Manríquez-Morán, N. L., Arenas-Ríos, E., & Ibargüengoytía, N. (2015). Male reproductive cycles in lizards. In J. L. Rheubert, D. S. Siegel, & S. E. Trauth (Eds.), Reproductive biology and phylogeny of lizards and tuatara (pp. 302–339). Boca Raton: CRC Press.
Messina, F. J., & Fox, C. W. (2001). Offspring size and number. In C. W. Fox, D. A, Roff, & D. J. Fairbairn (Eds.), Evolutionary ecology: concepts and case studies (pp. 113–127) New York: Oxford University Press.
Meiri, S., Brown, J. H., & Sibly, R. M. (2011). The ecology of lizard reproductive output. Global Ecology and Biogeography, 21, 592–602. https://doi.org/10.1111/j.1466-8238.2011.00700.x
Miles, D. B., Sinervo, B., & Frankino, W. A. (2000). Reproductive burden, locomotor performance, and the cost of reproduction in free ranging lizards. Evolution, 54, 1386–1395. https://doi.org/10.1111/j.0014-3820.2000.tb00570.x
Miller, C. M. (1997). Eumeces sumichrasti. Brood. Herpetological Review, 28, 151–152.
Olsson, M., & Shine, R. (1997). The limits to reproductive output: offspring size versus number in the sand lizard (Lacerta agilis). The American Naturalist, 149, 179–188. https://doi.org/10.1086/285985
Pavón-Vázquez, C. J., García-Vázquez, U. O., Bryson Jr., R. W., Feria-Ortiz, M., Manríquez-Morán, N. L., & Nieto-Montes de Oca, A. (2018). Integrative species delimitation in practice: revealing cryptic lineages within the short-nosed skink Plestiodon brevirostris (Squamata: Scincidae). Molecular Phylogenetics and Evolution, 129, 242–257. https://doi.org/10.1016/j.ympev.2018.08.020
Peig, J., & Green, A. J. (2009). New perspectives for estimating body condition from mass/length data: the scaled mass index as an alternative method. Oikos, 118, 1883–1891. https://doi.org/10.1111/j.1600-0706.2009.17643.x
Piñaza-Soto, J. C., & Hernández-Hernández, C. L. (2011). Protección y conservación de la biodiversidad del estado de Puebla. In CONABIO (Ed.), La biodiversidad en Puebla: estudio de estado (pp. 311–356). México D.F: Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad (CONABIO).
Qualls, F. J., & Shine, R. (1997). Geographic variation in ‘costs of reproduction’ in the scincid lizard Lampropholis guichenoti. Functional Ecology, 11, 757–763. https://doi.org/10.1046/j.1365-2435.1997.00150.x
Ramírez-Pinilla, M. P., de Pérez, G. R., & Alvarado-Ramírez, C. (2015). Oogenesis and the ovarian cycle. In J. L. Rheubert, D. S. Siegel, & S. E. Trauth (Eds.), Reproductive biology and phylogeny of lizards and tuatara (pp. 213–252). Boca Raton: CRC Press.
Recknagel, H., & Elmer, K. R. (2019). Differential reproductive investment in co-occurring oviparous and viviparous common lizards (Zootoca vivipara) and implications for life-history trade-offs with viviparity. Oecologia, 190, 85–98. https://doi.org/10.1007/s00442-019-04398-w
Rodríguez-Romero, F., Méndez, F. R., & López-González, L. (2005). Análisis comparado del esfuerzo reproductor en algunos lacertilios mexicanos de ambiente tropical y templado. Revista de la Sociedad Mexicana de Historia Natural, 2, 168–177.
Ruiz-Corral, J. A., Flores-López, H. E., Regalado-Ruvalcaba, J. R., & Ramírez-Ojeda, G. (2012). Estadísticas climáticas normales del estado de Jalisco. México D.F: Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias.
Semarnat (Secretaría de Medio Ambiente y Recursos Naturales). (2019). Modificación del Anexo Normativo III, Lista de especies en riesgo de la Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental-Especies nativas de México de flora y fauna silvestres-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-Lista de especies en riesgo, publicada el 30 de diciembre de 2010. Diario Oficial de la Federación, Ciudad de México.
Shine, R. (1980). “Costs” of reproduction in reptiles. Oecologia, 46, 92–100. https://doi.org/10.1007/BF00346972
Shine, R., & Schwarzkopf, L. (1992). The evolution of reproductive effort in lizards and snakes. Evolution, 46, 62–75. https://doi.org/10.1111/j.1558-5646.1992.tb01985.x
Sinervo, B., Méndez-de la Cruz, F., Miles, D. B., Heulin, B., Bastiaans, E., Villagrán-Santa Cruz, M. et al. (2010). Erosion of lizard diversity by climate change and altered thermal niches. Science, 328, 894–899. https://doi.org/10.1126/science.1184695
Sorci, G., Clobert, J., & Michalakis, Y. (1996). Cost of reproduction and cost of parasitism in the Common Lizard, Lacerta vivipara. Oikos, 76, 121–130. https://doi.org/10.2307/3545754
Swain, R., & Jones, S.M. (2000). Maternal effects associated with gestation conditions in a viviparous lizard, Niveoscincus metallicus. Herpetological Monographs, 14, 432–440. https://doi.org/10.2307/1467056
Tinkle, D. W. (1972): The dynamics of a Utah population of Sceloporus undulatus. Herpetologica, 28, 351–359.
Uller, T., & Olsson, M. (2010). Offspring size and timing of hatching determine survival and reproductive output in a lizard. Oecologia, 162, 663–671. https://doi.org/10.1007/s00442-009-1503-x
Uller, T., & While, G. M. (2015). The evolutionary ecology of lizard reproductive investment. In J. L. Rheubert, D. S. Siegel, & S. E. Trauth (Eds.), Reproductive biology of lizards and tuatara (pp. 425–447) Boca Raton: CRC Press.
Villers-Ruiz, L., Rojas-García, F., & Tenorio-Lezama, P. (2006). Guía botánica del Parque Nacional Malinche, Tlaxcala-Puebla. México D.F: Centro de Ciencias de la Atmósfera, UNAM/ Instituto de Biología, UNAM.
Vitt, L. J., & Cooper Jr, W. E. (1985). The evolution of sexual dimorphism in the skink Eumeces laticeps: an example of sexual selection. Canadian Journal of Zoology, 63, 995–1002. https://doi.org/10.1139/z85-148
Vitt, L. J., & Price, H. J. (1982). Ecological and evolutionary determinants of relative clutch mass in lizards. Herpetologica, 38, 237–255.
Wapstra, E., & Swain, R. (2001). Reproductive correlates of abdominal fat body mass in Niveoscincus ocellatus, a skink with an asynchronous reproductive cycle. Journal of Herpetology, 35, 403–409. https://doi.org/10.2307/1565958
Watson, C. M., Makowsky, R., & Bagley, J. C. (2014). Reproductive mode evolution in lizards revisited: updated analyses examining geographic, climatic and phylogenetic effects support the cold-climate hypothesis. Journal of Evolutionary Biology, 27, 2767–2780. https://doi.org/10.1111/jeb.12536
Wei-Guo, D. U., & Xiang, J. I. (2001). Growth, sexual size dimorphism and female reproduction of blue tailed skinks, Eumeces elegans. Zoological Research, 22, 279–286.
Williams, G. C. (1966). Natural selection, the costs of reproduction, and a refinement of Lack’s principle. The American Naturalist, 100, 687–690. https://doi.org/10.1086/282461
Youssef, M. K., Adolph, S. C., & Richmond, J. Q. (2008). Evolutionarily conserved thermal biology across continents: the North American lizard Plestiodon gilberti (Scincidae) compared to Asian Plestiodon. Journal of Thermal Biology, 33, 308–312. https://doi.org/10.1016/j.jtherbio.2008.02.007
Zar, J. H. (2010): Biostatistical analysis, 5th edition. Saddle River, New Jersey: Prentice Hall.