Elisa Sandoval-Serés a, b, *, Khiavett Sánchez-Pinzón c, Rafael Reyna-Hurtado c
a Asociación Biomas, Avenida de América 64, 28028 Madrid, Spain
b University of Oxford, Department of Zoology, Wildlife Conservation Research Unit, Recanati-Kaplan Centre, Tubney House, Abingdon Road, Tubney, OX13 5QL, UK
c El Colegio de la Frontera Sur, Unidad Campeche, Av. Rancho, Polígono 2ª Lerma, 24500 Ciudad de Campeche, Campeche, Mexico
*Corresponding author: esandovalseres@gmail.com (E. Sandoval-Serés)
Received: 22 February 2021; accepted: 3 November 2021
Abstract
We aimed to evaluate the temporal and spatial use of waterholes by ocelots in Calakmul Biosphere Reserve (CBR), Mexico. From 2014 to 2017, we monitored 11 waterholes with camera traps. We compared diel activity patterns with circular statistics depending on waterholes’ level of human intensity and distance to the Calakmul road, seasonality, and sex. We identified 40 different ocelots. Four waterholes were the most important ones, being 2 of them close to the road. Individuals took on average 19 days to return to waterholes. The diel activity of ocelots was 63.67% nocturnal, 20.70% crepuscular and 15.60% diurnal, and they were more diurnal in waterholes distant from the road. Their activity pattern was bimodal and it did not change between any of the categories tested. This is the first study to determine the spatial and temporal activity of ocelots in waterholes of Mexico. Ocelots are mainly nocturnal, and this pattern is conserved throughout CBR, however, they are able to adjust slightly their activity depending on extrinsic factors, such as an increased human presence. In the Calakmul region, all waterholes are crucial, and we particularly emphasize the conservation of the most important waterholes for ocelots, especially the ones close to the road.
Keywords: Circular statistics; Mesopredator; Sustainable tourism; Temporal overlap
© 2022 Universidad Nacional Autónoma de México, Instituto de Biología. This is an open access article under the CC BY-NC-ND license
Uso de aguadas y patrón de actividad de ocelotes en la selva de Calakmul, México
Resumen
Nuestro objetivo fue evaluar el uso de aguadas por ocelotes y su patrón de actividad en la Reserva de la Biosfera Calakmul, México. Del 2014 al 2017, monitoreamos 11 aguadas con cámaras trampa. Con estadística circular, comparamos los patrones de actividad dependiendo del nivel de intensidad humana de las aguadas y su distancia al camino de Calakmul, temporada, y sexo. Identificamos 40 ocelotes, los cuales cada ~19 días visitaban las aguadas. Cuatro aguadas (2 cerca del camino) fueron las más importantes. Los ocelotes fueron 63.67% nocturnos, 20.70% crepusculares y 15.60% diurnos, y fueron más diurnos en las aguadas lejos del camino. Su patrón de actividad fue bimodal y éste no cambió entre ninguna categoría evaluada. Éste es el primer trabajo en identificar a ocelotes y determinar su patrón de actividad en aguadas de México. Los ocelotes son mayoritariamente nocturnos y este patrón se conserva a lo largo de Calakmul, sin embargo, son capaces de ajustar su actividad dependiendo de las circunstancias (por ejemplo, para evitar a humanos). En Calakmul, la conservación de las aguadas es crucial y particularmente, enfatizamos la conservación de las aguadas identificadas como las más importantes para los ocelotes, en especial las que están cerca del camino.
Palabras clave: Estadística circular; Mesodepredador; Turismo sustentable; Superposición temporal
© 2022 Universidad Nacional Autónoma de México, Instituto de Biología. Este es un artículo Open Access bajo la licencia CC BY-NC-ND
Introduction
Water is crucial to animals’ survival, especially in tropical dry forests, where high temperatures and extreme rainfall seasonality prevail (Stoner & Timm, 2010). In some seasonal environments, the only water sources available for wild animals during the dry season are waterholes (Montalvo et al., 2019; O’Farrill et al., 2014; Valeix et al., 2010). Hence, waterholes are extremely relevant for animal population persistence (Martinez et al., 2021; Moreira-Ramírez et al., 2016; O’Farrill et al., 2014), as well as, for richness, abundance, and diversity of animal species (Martínez-Kú et al., 2008; Ponce, 2018; Sánchez-Pinzón et al., 2020). As not all waterholes conserve water during the dry season (Sánchez-Pinzón et al., 2020), the distribution and temporal variation of waterholes can influence animal movements (Martínez-Kú et al., 2008; Montalvo et al., 2019, Sánchez-Pinzón et al., 2020). For example, a higher aggregation of animals occurs around those waterholes that conserve water during the dry season (Martínez-Kú et al., 2008; Sánchez-Pinzón et al., 2020; Valeix et al., 2010).
Animals must balance trade-offs between foraging or hydratation with risk predation (Monterroso et al., 2013; Nasanbat et al. 2021); as such, the time of the day when they are active in sites with key resources, such as water, has important implications for their survival (Stoner & Timm, 2010). Environmental —rainfall, temperature— and ecological —prey, predation risk— characteristics can affect mammals’ activity patterns (Beltran & Delibes, 1994; Di Bitetti et al., 2010). For instance, nocturnality in mammalian carnivores can be attributed to coincide with the activity period of their prey (Emmons, 1988; Goulart, Graipel et al., 2009; Tang et al., 2019), and/or to avoid high temperatures during the day (Stoner & Timm, 2010). Moreover, temporal segregation can facilitate coexistence between carnivores to reduce competition and predation (Di Bitetti et al., 2010; Hayward & Slotow, 2009).
Human impact can also modify carnivores’ activity; therefore, it can be used as an indication of environmental degradation (Boydston et al., 2003; Massara, De Oliveira-Paschoal, Bailey, Doherty, de Frias-Barreto et al., 2018). In fact, many mammals, including carnivores, increase their nocturnality to avoid humans (Cruz et al., 2018; Di Bitetti et al., 2010; Gaynor et al., 2018).
In ecosystems with limited water availability, the knowledge of the activity pattern of carnivores, including felines, can assist conservation to propose optimal management decisions (Massara, De Oliveira-Paschoal, Bailey, Doherty, de Frias-Barreto et al., 2018). The ocelot (Leopardus pardalis) is a medium-sized, solitary, Neotropical feline frequently associated with water sources (Bagilet et al., 2017; Dias, Lima-Massara et al., 2019; Di Bitetti et al., 2006; Goulart, Cáceres et al., 2009; Wang et al., 2019). It is mainly nocturnal but with some activity during the day (Di Bitetti et al., 2006; Maffei et al., 2005; Torres-Romero et al., 2017). It is distributed from the southeast USA to northern Argentina (Murray & Gardner, 1997; Paviolo, 2015), occupying habitats ranging from scrublands to tropical rainforests (Caso, 1994; Wang et al., 2019). Ocelots’ diet varies from site to site (Murray & Gardner, 1997). In tropical forests, their main prey are iguanas, small rodents and agoutis, but also, they can eat opossums, armadillos, monkeys, and deer (Emmons, 1988; Moreno et al., 2006). Habitat quality, rainfall and prey availability are crucial for the presence and abundance of ocelots (Di Bitetti et al., 2008; Oliveira et al., 2010; Santos et al., 2019).
Although ocelots’ activity pattern is widely known (Di Bitetti et al., 2006; Murray & Gardner, 1997; Torres-Romero et al., 2017), its activity around waterholes had never been studied before. Additionally, understanding wildlife use of key resources (e.g., water) is necessary for adequate conservation planning (O’Farrill et al., 2014; Ponce, 2018). Hence, we aimed to evaluate the waterhole use of ocelots in Calakmul, Mexico, and the activity pattern of ocelots in these waterholes depending on extrinsic factors (waterholes’ level of human intensity and distance to the Calakmul road, and seasonality), and intrinsic factors (ocelots’ sex). We expected that the most important waterholes for ocelots were the ones with less human presence. Moreover, it was expected that ocelots visited the waterholes more often during the dry season when there is less water availability. Regarding activity patterns, we expected that ocelots were more nocturnal in waterholes with high human presence to avoid human encounters, and in the dry season to avoid high temperatures during daylight. We predicted that between sexes, daily activity did not differ as we assumed that both sexes preyed upon similar species and that there was an intersexual temporal tolerance. Waterholes’ use by ocelots and their activity pattern in an ecosystem where animals’ survival depend completely on waterholes can be relevant for ocelots’ conservation and ecology.
Materials and methods
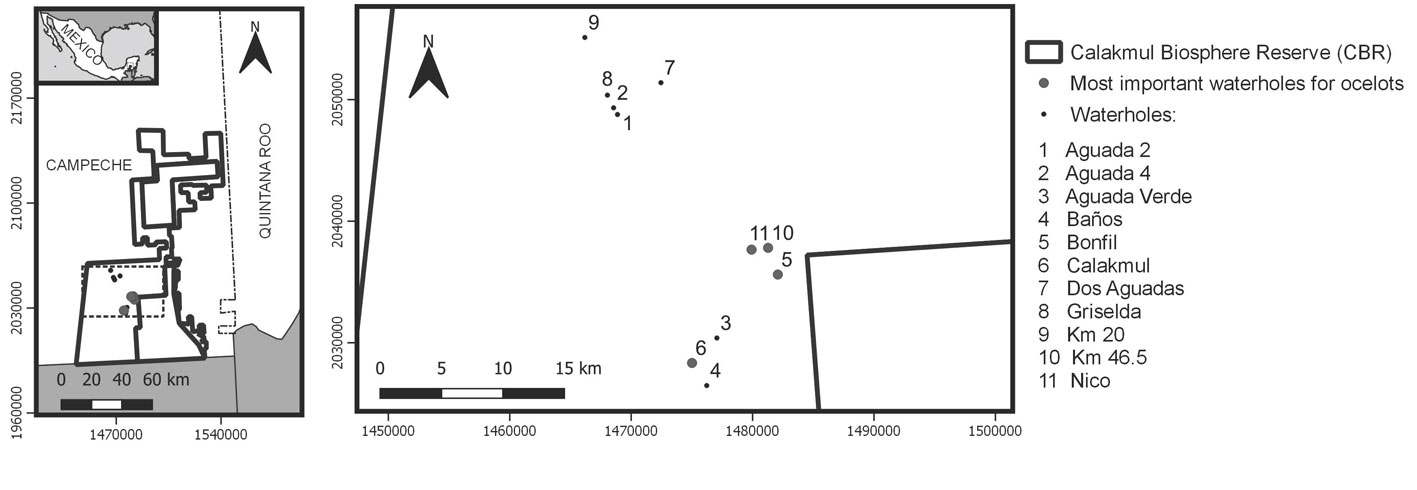
This study was conducted in the Calakmul Biosphere Reserve (CBR) (18°19’19.19” N, 89°51’38.02” W), located in the Yucatán Peninsula at the southeast of Campeche State, Mexico (Fig. 1). CBR was established in 1989 and comprises 7,231 km2, including the Calakmul archaeological ruins (which can be reached through a low-speed paved road) (INE, 1999). Together with other reserves of Belize and Guatemala, it is part of the Maya Forest, the largest tropical rainforest of Mesoamerica (Reyna-Hurtado et al., 2019). In CBR, elevation ranges from 200 to 340 m asl. The climate is warm sub-humid with rainfall occurring in the summer (wet season: from June to November). The annual rainfall is between 500 to 2,500 mm, with 60 mm of rain on the driest month, and the mean temperature is 24.6°C (INE, 1999). The only human activity in the southern area is restricted to tourism in the archaeological city with a very low visitors rate (less than 50 per day) along the years and into a single road that leads to the archaeological city. In the area, there is 1 waterhole every 10 km2 on average, and during the dry season most of them dry up completely (Reyna-Hurtado et al., 2019). The vegetation is tropical dry forest, and waterholes surrounding vegetation is low-flooded forest (Martínez & Galindo-Leal, 2002). Main ocelots’ potential prey found in CBR are reptiles (e.g., iguana Ctenosaura defensor), deer (Odocoileus virginianus, Mazama spp.), armadillo (Dasypus novemcinctus), opossums (e.g., Didelphis spp., Philander opossum, Marmosa mexicana), rabbit (Sylvilagus floridanus), and rodents such as mice, rats, squirrels, paca (Cuniculus paca), and agouti (Dasyprocta punctata) (Colston et al., 2015; Reid, 2009).
From 2014 to 2017, we monitored between 8 and 11 waterholes during all months of the years. We deployed camera traps (RECONYX: Hyperfire HC600, HC800) at the border of each waterhole, 50 cm above the floor. Each waterhole had 1 camera trap. The monitored waterholes were 7 to 120 m long (average = 38 m) and 5 to 100 width (average = 29 m) (Table 1) and were located at least 1 km away from each other. All the camera traps were set to operate continuously for 24 h each day with infrared LED flash. To avoid timing bias, we set all cameras on the same time schedule, using the summer timing (UTC-5) for the whole year. For the 4 years, the trapping effort was 11,126 camera trap/day.
Table 1
Characteristics of waterholes monitored from 2014 to 2017 in Calakmul Biosphere Reserve.
| Waterhole (*most important waterholes for ocelots) | Length (m) (approximate) | Width (m)
(approximate) |
Human activity
(High: visited from daily to once a week. Medium: visited every 2 weeks. Low: visited once a month.) |
Trapping effort (days) | Distance to the Calakmul road (m) |
| Aguada 2 | 12 | 10 | Low | 620 | 1,000 |
| Aguada 4 | 7 | 5 | Low | 1,138 | 1,000 |
| Baños | 45 | 30 | Medium | 745 | 590 |
| Bonfil* | 35 | 35 | High | 846 | 200 |
| Calakmul* | 120 | 100 | Medium | 1,748 | 1,550 |
| Dos Aguadas | 80 | 70 | Medium | 554 | 3,523 |
| Griselda | 10 | 5 | Medium | 1,412 | 711 |
| Km 20 | 25 | 25 | High | 1,079 | 800 |
| Km 46.5* | 35 | 35 | High | 861 | 100 |
| Nico* | 20 | 10 | Low | 951 | 1,400 |
| Verde | 25 | 20 | Low | 1,172 | 2,430 |
To quantify the ocelots visiting each waterhole, we recognized individuals by rosette and stripe patterns (Trolle & Kéry, 2003) and sex when possible. As there were no double stations per waterhole, we counted the minimum number of individuals as those individuals known by the right side of the body pattern (unless individuals were different due to different sex). We used the right side because there were more ocelot records on that side. To test how often (in days) individuals visited the same waterhole (latency) during each season, we only used those individuals recognized within the same season. For latency analyses, we recognized each individual by the right side, and we used both sides only when the individual turned around. We excluded latency samples between seasons. We performed a Mann-Whitney-U test to detect differences in latency between seasons (Zar, 2010).
For activity patterns, we assumed that the likelihood of obtaining a photo-capture of an ocelot was the same throughout night and day. To diminish the repetition bias of detecting the same animal, we considered 1 record of each individual per hour per camera trap as an independent detection event for each 24-h period. In CBR, sunrise is between 6:00 to 7:00 h and sunset between 17:30 to 19:30 h. Therefore, we categorized nocturnal activity as 20:00 to 05:00 h, diurnal as 08:00 to 17:00 h, and crepuscular (first and last hours of daylight) as 05:01-07:59 h and 17:01-19:59 h (UTC-5). These categories are widely used in other activity pattern studies (Azevedo et al., 2019; Gómez et al., 2005; Ross et al., 2013). We considered the wet season from June to November and the dry season from December to May.
For diurnality index calculation, we divided the total activity counts during the diurnal timing by the activity counts during the 24-h period and subsequently multiplied the result by 100 (Bagilet et al., 2017). We calculated nocturnal and crepuscular activity indexes in a similar way. We performed all these indexes per season and sex.
Furthermore, we calculated diurnality indexes for waterholes with high, medium, and low human activity, as well as, for waterholes close to (< 500 m) and distant (> 500 m) from the Calakmul road, which is a paved low-traffic road. Although human activity was not quantified in this study, we categorized the level of human activity as the level of human intensity of each waterhole receiving human visitors according to our experience in the area (~20 years). We considered ‘Bonfil’, ‘Km 46.5’, and ‘Km 20’ waterholes with high human activity (being visited from daily to once a week); ‘Griselda’, ‘Baños’, ‘Calakmul’, and ‘Dos Aguadas’ waterholes with intermediate human activity (being visited every 2 weeks approximately); and ‘Aguada Verde’, ‘Aguada 2’, ‘Aguada 4’, and ‘Nico’ waterholes with low human activity (being visited once a month approximately and only for researchers). Waterholes close to the road are only ‘Bonfil’ and ‘Km 46.5’. We tested differences between diurnal and nocturnal-crepuscular indexes with a chi-square test (χ²) (Zar, 2010).
To know whether the activity pattern of ocelots was uniformly distributed around the 24 h cycle, we performed a Rayleigh test of uniformity, in which the null hypothesis is that the activity is uniformly distributed during the day (Rayleigh, 1880). For the test, we used the R package ‘circular’ (Lund et al., 2017).
In addition, we performed statistical models of activity patterns using circular distributions (Zar, 2010). We constructed models based on observed data to predict daily activity patterns as a function of continuous trigonometric predictor variables describing 1 and 2 complete trigonometric cycles within a 24-h period (SinΘ, CosΘ, and Sin2Θ, Cos2Θ with Θ πt/24, ‘t’ being time in hours), where the best model was defined by the lowest significant p-value and with significant cycles (Appendix). These models help to predict and illustrate activity patterns, as well as, to test statistically activity differences between categories (Ross et al., 2013). We tested activity patterns between seasons and between sexes. We tested differences between the activity patterns of season and sex categories (wet vs. dry; female vs. male) using a periodic regression model (De Bruyn & Meeuwig, 2001; Ross et al., 2013). To test for differences in ocelots’ activity pattern for season and sex categories, as well as for categories of waterholes with high and medium-low human activity and waterholes close to and far from the road, we performed a Mardia-Watson-Wheeler test using the R package ‘circular’ (Lund et al., 2017). Moreover, we calculated coefficients of overlap (Δ1 when there were less than 75 observations, otherwise Δ4 was used) between all categories; and we calculated 95% confidence intervals (CI) with 1000 bootstrap samples (Meredith & Ridout, 2018; Ridout & Linkie, 2009) using the R package ‘overlap’ (Meredith & Ridout, 2020). We visualized activity patterns using the R package ‘plotrix’ (Lemon et al., 2015). All analyses were performed in R (R CORE TEAM, 2020).
Results
In 4 years and across eleven waterholes, we identified 40 different ocelots: 13 females, 21 males, and 5 of unknown sex. The most important waterholes (in terms of receiving more individuals) for ocelots were ‘Calakmul’, ‘Bonfil’, ‘Nico’, and ‘Km 46.5’ (Table 2). In each of these 4 waterholes, we identified at least 5 different individuals throughout 4 years. Furthermore, ‘Calakmul’ and ‘Bonfil’ waterholes were the ones that received the most visits by ocelots (Table 3). In ‘Km 46.5’ waterhole, an offspring with its mom was photographed. We identified between 1 and 3 ocelots visiting and sharing the same waterhole per year, and no individuals visited 2 different waterholes.
In addition, each individual visited the same waterhole in a very similar way between dry and wet seasons, on average every 19 days in the dry season [N= 43 latency samples; first quartile (25%) = 8 days; third quartile (75%) = 45 days] and on average every 20 days in the wet season (N = 43; first-quartile = 6; third quartile = 35), and this difference was not significant (U = 795; p = 0.839). Moreover, more than half of the individuals did not return to the same waterhole after a year, but the ones who returned did it for 2 or 3 years. ‘Calakmul’ waterhole had the most individuals re-visiting it (e.g., 2 males returned every year from 2015 to 2017).
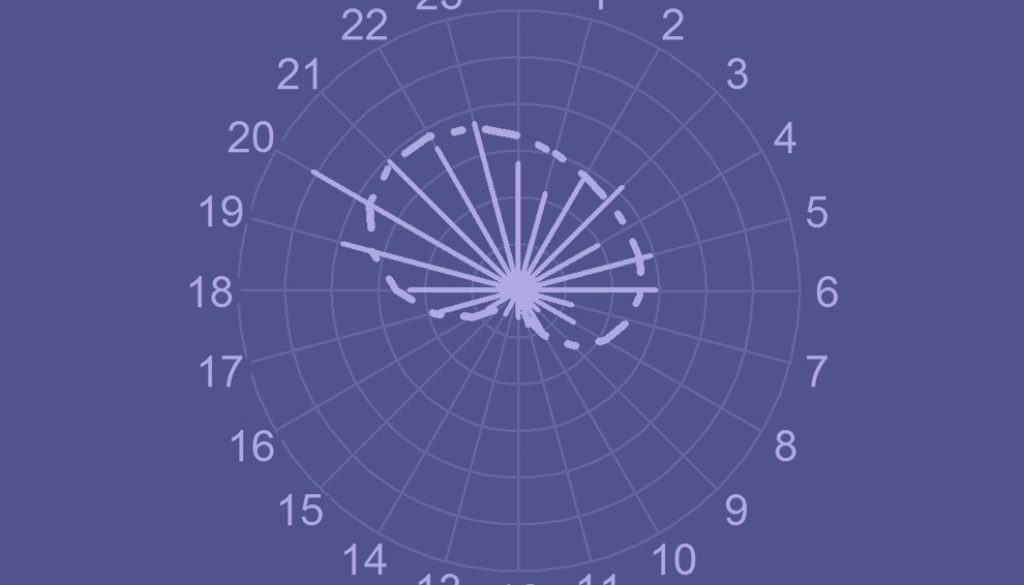
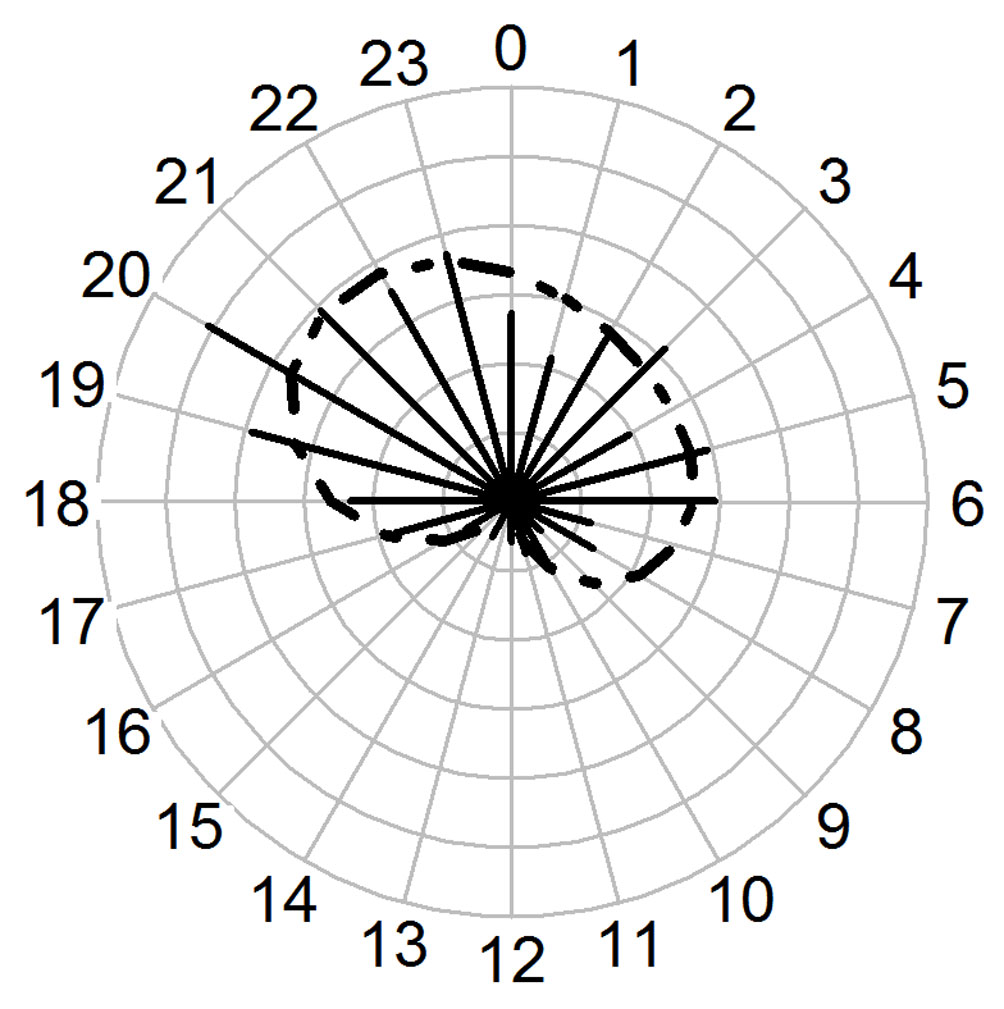
There were 256 independent detections in total (122 in the wet season, and 134 in the dry one; 92 detections for females, 114 for males and 50 for unknown sex). As the best model was accurate to predict and illustrate the activity pattern of ocelots around waterholes (sinx2+cosx1+cosx2: adjusted-R2 = 0.78, F3, 20 = 28.88; p < 0.001; Appendix), we were able to use it to illustrate ocelots’ activity pattern around waterholes (Fig. 2). In general, activity for ocelots was concentrated between 19:00 and 00:00 h, and then between 3:00 and 6:00 h. Almost no activity was found between 10:00 and 16:00 h (Fig. 2). Ocelots were 63.67% nocturnal, 20.7% crepuscular and 15.6% diurnal (Table 4). The null hypothesis of the Rayleigh test was rejected, indicating that the activity pattern of ocelots was not uniform (Z = 37.062; p < 0.001). As primary ocelots’ activity peak was between 20:00 and 00:00 h (median = 22:42), ocelots can be categorized as mostly nocturnal around waterholes (Fig. 2).
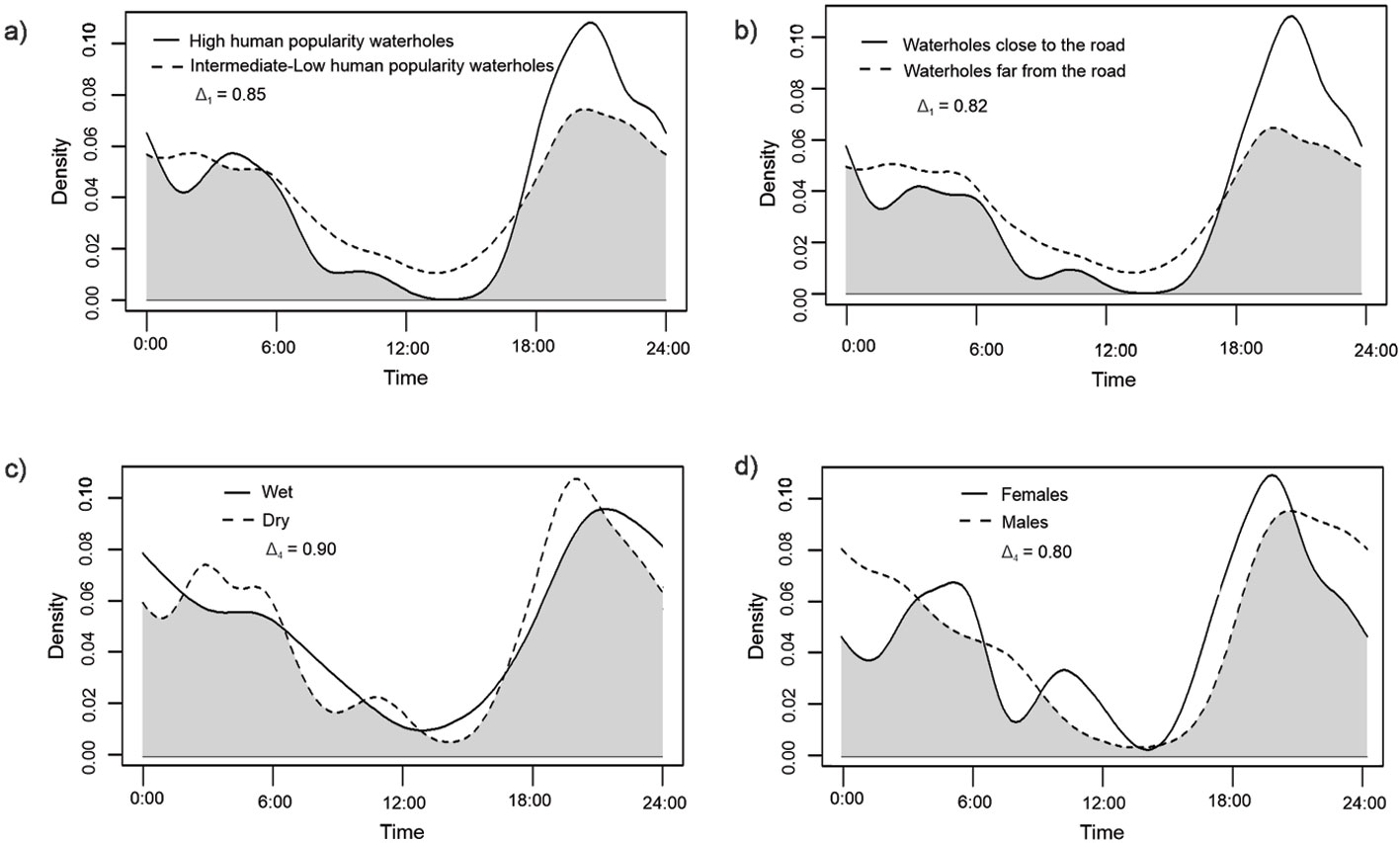
In waterholes with the highest human visits rate, 5% of ocelot records were diurnal (N = 79); and in waterholes with intermediate (N = 137) and little human activity (N = 38), 20% and 16% of ocelot records were diurnal, respectively. Only the percentages of waterholes between a high and intermediate number of visits differed significantly (χ² = 7.59, df = 1, p = 0.006). In waterholes close to the road, 5% of ocelot records were diurnal, whereas in waterholes far away from the road 18% of ocelot records were diurnal. These percentages were significantly different (χ² = 5.75, df = 1, p = 0.016). In waterholes with high human intensity and close to the road, there were barely any records (only 3) of ocelots between 7:00 to 17:00 h (Fig. 3a, b).
However, there were no significant differences in the activity pattern of ocelots neither between waterholes with high and intermediate-low human intensity (W = 0.67, df = 2, p = 0.716), nor between waterholes close to and far from the road (W = 1.18, df = 2, p = 0.554). The coefficient of overlap between waterholes with high and moderate-low human intensity was high (Δ1 = 0.85, CI: 0.74 – 0.94), as well as the overlap with waterholes between different road distances (Δ1 = 0.82, CI: 0.71 – 0.91) (Fig. 3a, b).
No significant differences were detected in ocelots’ activity pattern between seasons (W = 0.012, df = 2, p > 0.5; and cosx1*season: F 1, 44 = 0.014; p = 0.905; Appendix), and their activity overlapped extensively (Δ4 = 0.90, CI: 0.87 – 0.98) (Fig. 3c). In the wet season ocelots tended to be less crepuscular and more diurnal and nocturnal (Table 4), but these differences were non-significant (χ² = 0.82, df = 2, p = 0.663).
No significant differences were detected in the activity pattern between sexes (W = 0.021, df = 2, p > 0.5; and cosx1*sex: F1, 44 = 2.066; p = 0.158; Appendix). Activity between females and males overlapped extensively (Δ4 = 0.80, CI: 0.76-0.92) (Fig. 3d), and although females tended to be more diurnal and crepuscular than males (Table 3), these differences were non-significant (χ² = 2.33, df = 2, p = 0.312).
Table 2
Number of ocelots identified visiting the waterholes in Calakmul Biosphere Reserve from 2014 to 2017.
| Waterhole (*most important waterholes for ocelots) | Females | Males | Unknown sex | Total individuals | Average of individuals per year |
| Aguada 2 | 0 | 0 | 1 | 1 | 0.5 |
| Aguada 4 | 0 | 4 | 0 | 4 | 2 |
| Aguada verde | 1 | 2 | 0 | 3 | 0.8 |
| Baños | 1 | 1 | 0 | 2 | 0.5 |
| Bonfil* | 1 | 3 | 1 | 5 | 1.3 |
| Calakmul* | 3 | 4 | 0 | 7 | 1.75 |
| Dos aguadas | 2 | 1 | 0 | 3 | 0.8 |
| Griselda | 1 | 1 | 1 | 3 | 0.8 |
| Km 20 | 1 | 1 | 0 | 2 | 0.5 |
| Km 46.5* | 2 | 2 | 1 offspring | 5 | 1.3 |
| Nico* | 2 | 2 | 1 | 5 | 1.7 |
Table 3
Number of independent detections of ocelots visiting the waterholes in Calakmul Biosphere Reserve per year from 2014 to 2017.
| Waterhole | 2014 | 2015 | 2016 | 2017 | Total | Average of independent detections per year |
| Aguada 2 | NA | NA | 0 | 1 | 1 | 0.5 |
| Aguada 4 | NA | NA | 5 | 3 | 8 | 4 |
| Aguada verde | 1 | 3 | 2 | 1 | 7 | 1.8 |
| Baños | 0 | 3 | 1 | 3 | 7 | 1.8 |
| Bonfil | 8 | 15 | 5 | 6 | 34 | 8.5 |
| Calakmul | 28 | 6 | 6 | 22 | 62 | 15.5 |
| Dos aguadas | 14 | 0 | 11 | 6 | 31 | 7.8 |
| Griselda | 5 | 1 | 0 | 4 | 10 | 2.5 |
| Km 20 | 4 | 2 | 4 | 5 | 15 | 3.8 |
| Km 46.5 | 15 | 3 | 4 | 4 | 26 | 6.5 |
| Nico | NA | 8 | 4 | 7 | 19 | 6.3 |
NA = not applicable because there was no camera-trap station.
Table 4
Activity indexes of ocelots in Calakmul Biosphere Reserve.
| Activity | Total (%) | Season | Sex | ||
| Wet (%) | Dry (%) | Females (%) | Males (%) | ||
| Diurnal
08:00-17:00 |
15.63 | 17.21 | 14.18 | 18.48 | 10.53 |
| Nocturnal
20:00-05:00 |
63.67 | 64.75 | 62.69 | 56.52 | 68.42 |
| Crepuscular
05:01-07:59 and 17:01-19:59 |
20.70 | 18.03 | 23.13 | 25.00 | 21.05 |
Discussion
Despite not estimating density in this study, it is worthwhile mentioning that at least 40 individuals visited 11 waterholes separated a maximum distance of 40 km in CBR within 4 years. Such a minimal number of individuals could be comparable to the density found in the north of the Yucatán Peninsula, where the density is estimated to be ~14 ocelots/100 km2 (Torres-Romero et al., 2017).
The 4 most important waterholes are located further south and could probably receive more rainfall than the other ones. This is probably because in the southern part of CBR rainfall is more abundant and the medium semi-perennial forest is also more abundant than in the northern areas (Martínez & Galindo-Leal, 2002). Furthermore, ‘Calakmul’ waterhole was the one visited by most individuals and where most individuals returned, as this is the largest one monitored and next to a water stream, it could possibly sustain more ocelots visiting it. Different ocelots were able to share the same waterhole, indicating that one waterhole could be found inside different ocelot territories, as ocelot home ranges can overlap significantly (Azevedo et al., 2019; Caso, 2013; Murray & Gardner, 1997).
Contrary to expectations, 2 of the waterholes most visited by ocelots (‘Bonfil’ and ‘Km 46.5’) were the ones close to the road and some of the ones most visited by humans. Even though ocelots normally avoid sites with high human accessibility (Cruz et al., 2018) and human settlements (Dias, Lima-Massara et al., 2019), the road in CBR and the human visitors did not seem to stop ocelots from visiting the waterholes. This could be because human settlements are more invasive than a low-traffic road and low ecotourism activities.
Opposite to what we predicted, there were no differences between seasons in the latency of ocelots visiting each waterhole. This could be explained because season is not related to ocelots visiting a waterhole, but instead, it could be related to another trait, such as patrolling or hunting behaviours. On the other hand, seasonality can affect the latency of waterholes visits to other species more dependent on water, such as to tapirs (Tapirella bairdii), which did visit more often the waterholes during the dry season in the Calakmul region (Sandoval-Serés et al., 2016).
Ocelots were mainly nocturnal with some records during sunset and few records during the day, which coincides with other studies in Neotropical forests (Caso, 2013; Goulart, Graipel et al., 2009; Moreno & Bustamante, 2009).
Comparing ocelots’ nocturnality from different studies, the result in this study (63.67%) was similar to the one in tropical forests of Tamaulipas, Mexico (62%) (Caso, 1994), Pantanal, Brazil (51.6%) (Crawshaw & Quigley, 1989), and tropical forests of Argentina and Brazil (50%) (Bagilet et al., 2017). Ocelots were less nocturnal in CBR than in the Amazonia (79%; Gómez et al., 2005), savannah and dry forests of Bolivia (89%; Maffei et al., 2005), and tropical forests in Panamá (90%; Moreno & Bustamante, 2009). The similarity of ocelots’ activity pattern in CBR waterholes with different forest areas could indicate that ocelots have an entrenched activity pattern, as well as, that ocelots use the waterholes as an everyday behavioural movement rather than a focalized visit of waterholes in a specific time of the day.
Ocelots’ activity in this study had a bimodal activity pattern analogous to the one found in other studies (Caso, 2013; Goulart, Graipel et al., 2009; Moreno & Bustamante, 2009). Their activity peak in CBR (3:00-6:00 h and 19:00-23:00 h) was similar to the one of other ocelots in tropical dry forests of Mexico (Caso, 2013; Hernández-Saintmartín et al., 2013; Torres-Romero et al., 2017), tropical perennial forests of Mexico (Lira-Torres & Briones-Salas, 2012) and Panamá (Moreno & Bustamante, 2009). Ocelots were more active during the evening and early night in humid tropical forests of Brazil and Belize (Crawshaw, 1995; Goulart, Graipel et al., 2009; Harmsen et al., 2011) and coastal plain forests of Brazil (Wolff et al., 2019). In shrub dry vegetation in Brazil, ocelots main activity was at early night (Dias et al., 2018; Penido et al., 2017).
Activity pattern varies slightly from region to region (Di Bitetti et al., 2006; Goulart, Graipel et al., 2009; Oliveira et al., 2010), probably because ocelots can adapt their temporal activity to local characteristics, such as avoidance to humans and large predators (Massara, De Oliveira-Paschoal, Bailey, Doherty, de Frias-Barreto et al., 2018), mirror prey activity to increase their encounter probability (Emmons, 1988; Goulart, Graipel et al., 2009; Porfirio et al., 2016), or high-temperature avoidance during daylight with the trade-off of not coinciding with the activity pattern of their prey (Penido et al., 2017).
Despite ocelots’ activity pattern being similar throughout waterholes, diurnality was significantly lower in the most popular waterholes for tourists and the ones close to the road than in the other waterholes more isolated and less visited, this behaviour could be to avoid humans during the day. This adaptability was also found in other tropical forests (Cruz et al., 2018; García-R et al., 2019). Calakmul archaeological ruins are open to the public from 8:00 to 17:00 h (INAH, 2021), which can explain why there were very few ocelots’ records during those times in the most popular waterholes for tourists. Ocelots can avoid humans through temporal segregation (Massara, De Oliveira-Paschoal, Bailey, Doherty, de Frias-Barreto et al., 2018).
In addition, carnivores can adapt their activity to avoid intraguild competitors (Hayward & Slotow, 2009; Oliveira et al., 2010; Oliveira-Santos et al., 2012). Ocelots, jaguarundis (Puma yagouaroundi) and margays (Leopardus wiedii) have a large overlap in their diet (Silva-Pereira et al., 2011; Wang, 2002). Ocelots can be attacked occasionally by pumas (Puma concolor) and jaguars (Panthera onca) (Emmons, 1988; Murray & Gardner, 1997; Perera-Romero et al. 2020). Thus, to avoid intraguild competition (Massara, De Oliveira-Paschoal, Bailey, Doherty, de Frias-Barreto et al., 2018), there are some differences between ocelots’ temporal pattern with the one of other felines found in CBR: being jaguarundis diurnal, pumas nocturnal-crepuscular or cathemeral, margays nocturnal, jaguars nocturnal or cathemeral, and ocelots nocturnal or nocturnal-crepuscular (Bagilet et al., 2017; Hernández-Saintmartín et al., 2013; Weckel et al., 2006). In CBR, jaguars are cathemeral, pumas mainly nocturnal-crepuscular with their highest activity peak being at dawn and early night (5:00 and 00:00 h) (Estrada-Hernández, 2008), whereas ocelots were mainly nocturnal with their main activity peak being in the late evening (20:00 h). This, together with spatial evasion (Goulart, Cáceres et al., 2009), may explain their coexistence.
Some felines have a mirroring effect of their prey´s activity (Harmsen et al., 2011; Hernández-Saintmartín et al., 2013; Weckel et al., 2006); in particular, ocelots’ nocturnal activity coincides with much of their nocturnal prey (Dias et al., 2018; Nagy-Reis et al., 2019). In this study, ocelots were nocturnal-crepuscular with some activity during the day. This could mirror the activity of some of their potential prey found in CBR: opossums, small rodents and pacas being nocturnal, armadillos nocturnal-crepuscular, and squirrels, agoutis, and deer diurnal (Lira-Torres & Briones-Salas, 2012; Wang, 2002).
Contrary to expectations, seasonality did not affect ocelots’ activity pattern, this result was also found in other studies (Pérez-Irineo & Santos-Moreno, 2014; Porfirio et al., 2016). Ocelots in the dry season were more crepuscular. This slight difference might have been to overlap their activity to the one of their potential preys. For example, pacas were also more crepuscular during the dry season (Dias, Oliveira Souza-Almeida et al., 2019), and in CBR agoutis during the dry season had crepuscular activity peaks (Borges-Zapata et al., 2020). The non-significant differences of ocelots’ activity between seasons could be because, in the last years of this study, rainfall in CBR has been delayed and unevenly distributed throughout the year (Mardero et al., 2020), and some waterholes have stayed completely dry even in the wet season.
As in other studies (Crawshaw, 1995; Di Bitetti et al., 2006), sex did not influence ocelot’s activity, probably because both sexes prey on the same species (Emmons 1988). Nevertheless, in CBR, females tended to be more diurnal and crepuscular than males. As males travel more (Crawshaw, 1995; Emmons, 1988) they need to avoid high temperatures of daylight, which could have been why they were slightly more nocturnal than females.
As the vegetation of waterholes is almost homogeneous, and ocelots´ activity can highly overlap with the one of jaguars and pumas being unlikely modulated by them, ocelots’ activity could be influenced and adjusted to mirror their prey´s activity and/or to avoid humans (Martínez & Galindo-Leal, 2002; Penido et al., 2017; Santos et al., 2019). The evidence that the activity pattern of ocelots between all categories tested (waterholes’ level of human intensity and distance to roads, seasonality, and sex) was not significantly different could be attributed to the fact that ocelots have an innate well-entrenched diel activity pattern (being mostly nocturnal with a bimodal activity), but which can sometimes vary slightly to adapt to different circumstances, such as for human avoidance.
Ocelots are endangered in Mexico (SEMARNAT, 2019); thus, it is relevant to acknowledge their potential threats. Ocelots are associated with waterholes presence (Ponce, 2018; Wang et al., 2019). However, waterholes in Calakmul region are highly threatened by climate change and their inappropriate use by humans (Martínez-Kú et al., 2008; O’Farrill et al., 2014). Every year there are more waterholes that dry up completely (Sánchez-Pinzón et al., 2020). Water availability is not the only potential threat for ocelots in Calakmul region, but also the potential decrease in the abundance of their prey due to overhunting. Although in the surrounding villages of CBR, ocelots are rarely poached, their potential prey is frequently hunted (Escamilla et al., 2000). The decrease in their prey availability could exacerbate intraguild competition (Palomares & Caro, 1999). Thus, poaching ocelot’s prey should not be an underestimated threat for ocelots in Calakmul region. Ocelots’ potential prey are highly dependent on water (Borges-Zapata et al., 2020; Kumul et al., 2020), and ocelots could be using waterholes as hunting sites, especially because there can be more prey availability (Goulart, Cáceres et al., 2009). This emphasizes the importance of waterholes for ocelots’ main prey, and consequently, for ocelots themselves.
In CBR, up to now, ecotourism did not appear to be a threat for ocelots, as they were able to occur in waterholes with human activity, by adapting their temporal pattern to avoid people. However, it is important to emphasize that ‘Bonfil’ and ‘Km 46.5’ waterholes need to receive special conservation attention, as they are the ones closest to the road and, in turn, receive more tourist visits. In addition, they were 2 of the ones most visited by different ocelot individuals. Now ocelots have been able to adapt and visit these 2 waterholes at night, however, a potential increase of tourists visiting them could affect ocelots and even prevent them from using these 2 waterholes. Not to mention that activity pattern behavioural changes could potentially affect ocelots negatively by potentially increasing their energetic costs (Massara, De Oliveira-Paschoal, Bailey, Doherty, de Frias-Barreto et al., 2018).
Although we did not evaluate the importance of waterholes for ocelots, we identified 4 waterholes (‘Calakmul’, ‘Bonfil’, ‘Nico’, and ‘Km 46.5’) to be the most important ones for this species (both in terms of receiving more visits and individuals), and therefore should be prioritized for conservation efforts. The conservation of all waterholes in CBR is extremely important (O’Farrill et al., 2014; Sánchez-Pinzón et al., 2020), more specifically it is crucial to conserve the most used waterholes by ocelots in Calakmul region, especially the ones close to the road (‘Bonfil’, and ‘Km 46.5’), as they might be the most vulnerable ones.
To complement this study, ocelot movements among waterholes could be monitored to estimate the number of waterholes found in an ocelot’s home range. In addition, a comparison of ocelot’s activity with their prey’s activity within and outside protected areas could be performed to detect the impact of prey poaching in ocelots’ temporal pattern. All this is to elaborate conservation actions for this species.
Acknowledgements
To Conanp (Comisión Nacional de Áreas Naturales Protegidas) of the Calakmul Biosphere Reserve for granting permission to perform fieldwork. To El Colegio de la Frontera Sur (Mexico), and Consejo Nacional de Ciencia y Tecnología (Conacyt)-Ciencia Básica 182386 grant given to RRH for logistics and funding for fieldwork. To McGill University (Canada) for camera-trap equipment support. To Conacyt-scholarship 470731 for supporting KSP. To the Recanati-Kaplan Foundation and Wildlife Conservation Research Unit for supporting ESS continuing education (University of Oxford, UK). To N. Arias-Domínguez and P. Canul for helping during fieldwork. To C. Tan for analyses advise. To S. Gutiérrez for English advise.
Appendix. Activity pattern models to predict ocelots’ activity in the waterholes of Calakmul Rainforest. Best models to predict ocelots’ activity pattern in bold.
| Model | Cycles (significant cycles in bold) | d.f. | Adjusted R2 | F | p |
| All model Ocelots | sinx1 + sinx2 + cosx1 + cosx2 | 4 and 19 | 0.80 | 24.63 | 2.72e-07 |
| Final model Ocelots | sinx1 + sinx2 + cosx1 | 3 and 20 | 0.78 | 28.88 | 1.816e-07 |
| All model Dry Season | sinx1 + sinx2 + cosx1 + cosx2 | 4 and 19 | 0.61 | 10.01 | 0.000157 |
| Final model Dry Season | cosx1 + cosx2 | 2 and 21 | 0.56 | 15.78 | 6.561e-05 |
| All model Wet Season | sinx1 + sinx2 + cosx1 + cosx2 | 4 and 19 | 0.82 | 27.95 | 9.998e-08 |
| Final model Wet Season | sinx2 + cosx1 | 2 and 21 | 0.80 | 47.62 | 1.573e-08 |
| All model Females | sinx1 + sinx2 + cosx1 + cosx2 | 4 and 19 | 0.44 | 5.46 | 0.00423 |
| Final model Females | cosx1 + cosx2 | 2 and 21 | 0.36 | 7.46 | 0.00356 |
| All model Males | sinx1 + sinx2 + cosx1 + cosx2 | 4 and 19 | 0.69 | 14.1 | 1.667e-05 |
| Final model Males | sinx2 + cosx1 | 2 and 21 | 0.71 | 29.16 | 8.712e-07 |
References
Azevedo, F. C. C., Mähler, J. K. F., Indrusiak, C. B., Scognamillo, D., Conforti, V. A., Morato, R. G. et al. (2019). Spatial organization and activity patterns of ocelots (Leopardus pardalis) in a protected subtropical forest of Brazil. Mammal Research, 64, 503–510. https://doi.org/10.1007/s13364-019-00430-9
Bagilet, S., Delmar, R., & Cristina, M. (2017). Daily rhythmicity of behavior of nine species of South American feral felids in captivity. Physiology and Behavior, 180, 107–112. https://doi.org/10.1016/j.physbeh.2017.08.015
Beltran, J., & Delibes, M. (1994). Environmental determinants of circadian activity of free-ranging Iberian lynxes. Journal of Mammalogy, 75, 382–393. https://doi.org/10.2307/1382557
Borges-Zapata, J. Y., Contreras-Moreno, F. M., Serrano-Mac-Gregor, I., Sima-Pantí, D. E., Coutiño-Cal y Mayor, C., Zúñiga-Morales, J. A. et al. (2020). Uso de bebederos artificiales por el sereque centroamericano (Dasyprocta punctata) en la Reserva de la biosfera Calakmul, México. Agroproductividad, 13, 51–58. https://doi.org/https://doi.org/10.32854/agrop.vi0.1575
Boydston, E. E., Kapheim, K. M., Watts, H. E., Szykman, M., & Holekamp, K. E. (2003). Altered behaviour in spotted hyenas associated with increased human activity. Animal Conservation, 6, 207–219. https://doi.org/10.1017/S1367943003003263
Caso, A. (1994). Home range and habitat use of three neotropical carnivores in Northeast Mexico (M.Sc. Thesis). Texas A & M University, Kingsville, Texas, USA.
Caso, A. (2013). Spatial differences and local avoidance of ocelot (Leopardus pardalis) and jaguarundi (Puma yagouaroundi) in northeast Mexico (Ph.D. Thesis). Texas A & M University, USA, Texas.
Colston, T. J., Barão-Nóbrega, J. A. L., Manders, R., Lett, A., Willmott, J., Cameron, G. et al. (2015). Amphibians and rep-
tiles of the Calakmul Biosphere Reserve, México, with new records. Check List, 11, 1–7. https://doi.org/10.15560/11.5.1759
Crawshaw, P. G. (1995). Comparative ecology of ocelot (Felis pardalis) and jaguar (Panthera onca) in a protected subtropical forest in Brazil and Argentina (Ph.D. Thesis). University of Florida, USA, Florida.
Crawshaw, P. G., & Quigley, H. (1989). Notes on ocelot movement and activity in the Pantanal region, Brazil. Biotropica, 21, 377–379.
Cruz, P., Iezzi, M. E., De Angelo, C., Varela, D., Di Bitetti, M. S., & Paviolo, A. (2018). Effects of human impacts on habitat use, activity patterns and ecological relationships among medium and small felids of the Atlantic Forest. Plos One, 13, 1–21. https://doi.org/10.1371/journal.pone.0200806
De Bruyn, A. M. H., & Meeuwig, J. J. (2001). Detecting lunar cycles in marine ecology: periodic regression versus categorical ANOVA. Marine Ecology Progress Series, 214, 307–310. https://doi:10.3354/meps214307
Di Bitetti, M. S., De Angelo, C. D., Di Blanco, Y. E., & Paviolo, A. (2010). Niche partitioning and species coexistence in a Neotropical felid assemblage. Acta Oecologica, 36, 403–412. https://doi.org/10.1016/j.actao.2010.04.001
Di Bitetti, M. S., Paviolo, A., & De Angelo, C. (2006). Density, habitat use and activity patterns of ocelots (Leopardus pardalis) in the Atlantic Forest of Misiones, Argentina. Journal of Zoology, 270, 153–163. https://doi.org/10.1111/j.1469-7998.2006.00102.x
Di Bitetti, M. S., Paviolo, A., De Angelo, C. D., & Di Blanco, Y. E. (2008). Local and continental correlates of the abundance of a neotropical cat, the ocelot (Leopardus pardalis). Journal of Tropical Ecology, 24, 189–200. https://doi.org/10.1017/S0266467408004847
Dias, D. M., Campos, C. B., & Guimarães-Rodrigues, F. H. (2018). Behavioural ecology in a predator-prey system. Mammalian Biology, 92, 30–36. https://doi.org/10.1016/j.mambio.2018.04.005
Dias, D. M., Lima-Massara, R., Campos, C. B., & Guimarães-Rodrigues, F. H. (2019). Human activities influence the occupancy probability of mammalian carnivores in the Brazilian Caatinga. Biotropica, 51, 253–265. https://doi.org/10.1111/btp.12628
Dias, D. M., Oliveira Souza-Almeida, M., de Araújo-Piovezan, T. G., & Dantas, J. O. (2019). Spatiotemporal ecology of two neotropical herbivorous mammals. Papeis Avulsos de Zoologia, 59, 0–4. https://doi.org/10.11606/1807-0205/2019.59.10
Emmons, L. H. (1988). A field study of ocelots (Felis pardalis) in Peru. Revue d’écologie – la Terre et la Vie, 43, 133–157.
Escamilla, A., Sanvicente, M., Sosa, M., & Galindo-Leal, C. (2000). Habitat mosaic, wildlife availability, and hunting in the tro-
pical forest of Calakmul, Mexico. Conservation Biology, 14,
1592–1601. https://doi.org/10.1111/j.1523-1739.2000.99069.x
Estrada-Hernández, C. G. (2008). Dieta, uso de hábitat y patrones de actividad del puma (Puma concolor) y el jaguar (Panthera onca) en la selva Maya Centromerica. Revista Mexicana de Mastozoología, 12, 113–130. https://doi.org/10.1075/target.22.1.12mey
García-R, S., Botero-Cañola, S., Sánchez-Giraldo, C., & Solari, S. (2019). Habitat use and activity patterns of Leopardus pardalis (Felidae) in the Northern Andes, Antioquia, Colombia. Biodiversity, 20, 5–19. https://doi.org/10.1080/14888386.2019.1590235
Gaynor, K. M., Hojnowski, C. E., Carter, N. H., & Brashares, J. S. (2018). The influence of human disturbance on wildlife nocturnality. Science, 360, 1232–1235. https://doi.org/10.1126/science.aar7121
Gómez, H., Wallace, R. B., Ayala, G., & Tejada, R. (2005). Dry season activity periods of some Amazonian mammals. Studies on Neotropical Fauna and Environment, 40, 91–95. https://doi.org/10.1080/01650520500129638
Goulart, F. V. B., Cáceres, N. C., Graipel, M. E., Tortato, M. A., Ghizoni, I. R., & Oliveira-Santos, L. G. R. (2009). Habitat selection by large mammals in a southern Brazilian Atlantic Forest. Mammalian Biology, 74, 182–190. https://doi.org/10.1016/j.mambio.2009.02.006
Goulart, F. V. B., Graipel, M. E., Tortato, M., Ghizoni-Jr., I., Oliveira-Santos, L. G., & Cáceres, N. (2009). Ecology of the ocelot (Leopardus pardalis) in the Atlantic Forest of Southern Brazil. Neotropical Biology and Conservation, 4, 137–143. https://doi.org/10.4013/nbc.2009.43.03
Harmsen, B. J., Foster, R. J., Scott, C. S., Ostro, L. E. T., & Doncaster, C. (2011). Jaguar and puma activity patterns in relation to their main prey. Mammalian Biology, 76, 320–324. https://doi.org/10.1016/j.mambio.2010.08.007
Hayward, M. W., & Slotow, R. (2009). Temporal partitioning of activity in large African carnivores: Tests of multiple hypotheses. South African Journal of Wildlife Research, 39, 109–125. https://doi.org/10.3957/056.039.0207
Hernández-Saintmartín, A. D., Rosas-Rosas, O. C., Palacio-Núñez, J., Tarango-Arámbula, L. A., Clemente-Sánchez, F., & Hoogesteijn, A. (2013). Activity patterns of jaguar, puma and their potential prey in San Luis Potosí, Mexico. Acta Zoológica Mexicana, 29, 520–533. https://doi.org/10.1515/mammalia-2015-0175
INAH (Instituto Nacional de Antropología e Historia). (2021). Zona Arqueológica de Calakmul. Accessed on 07th, January, 2021 from: https://www.inah.gob.mx/zonas/62-zona-arqueologica-de-calakmul
INE (Instituto Nacional de Ecología). (1999). Programa de Manejo Reserva de la Biosfera Calakmul. Mexico City: INE.
Lemon, J., Bolker, B., Oom, S., Klein, E., Rowlingson, B., Wickham, H. et al. (2015). ‘plotrix’: Various Plotting Functions (version 3.6-1). R Package. Accessed on 10th, January, 2020 from: https://cran.r-project.org/web/packages/plotrix/index.html
Lira-Torres, I., & Briones-Salas, M. (2012). Abundacia relativa y patrones de actividad de los mamíferos de los Chimalapas, Oaxaca, México. Acta Zoológica Mexicana, 28, 566–585. https://www.redalyc.org/articulo.oa?id=57524600006
Lund, U., Agostinelli, C., Arai, H., Gagliardi, A., García-Portuges, E., Giunchi, D. et al. (2017). ‘circular’: Circular Statistics (version 0.4-93). R Package. Accessed on 10th, January, 2020 from: https://cran.r-project.org/web/packages/circular/index.html
Maffei, L., Noss, A. J., Cuéllar, E., & Rumiz, D. I. (2005). Ocelot (Felis pardalis) population densities, activity, and ranging behaviour in the dry forests of eastern Bolivia: data from camera trapping. Journal of Tropical Ecology, 21, 349–353. https://doi.org/10.1017/S0266467405002397
Mardero, S., Schmook, B., Christman, Z., Metcalfe, S. E., & De la Barreda-Bautista, B. (2020). Recent disruptions in the timing and intensity of precipitation in Calakmul, Mexico. Theoretical and Applied Climatology, 140, 129–144. https://doi.org/10.1007/s00704-019-03068-4
Martínez, E., & Galindo-Leal, C. (2002). La vegetación de Calakmul, Campeche, México: clasificación, descripción y distribución. Boletín de la Sociedad Botánica de México, 71, 7–32. https://doi.org/10.17129/botsci.1660
Martinez, W. E., Reyna-Hurtado, R. A., Naranjo, E. J., Thornton, D., Cal, R. N., & Figueroa, O. A. (2021). Occupancy rate and observations of Baird’s tapir (Tapirella Bairdii) near waterholes in the Maya forest corridor, Belize. Therya, 12, 37–43. https://doi.org/10.12933/THERYA-21-969
Martínez-Kú, D., Escalona-Segura, G., & Vargas-Contreras, J. (2008). Importancia de las aguadas para los mamíferos de talla mediana y grande en Calakmul, Campeche, México. In C. Lorenzo, E. Espinoza, & J. Ortega (Eds.), Avances en el estudio de los mamíferos de México. Especiales, Vol. II (pp. 449-468). Mexico City: Asociación Mexicana de Mastozoología, A.C.
Massara, R., De Oliveira-Paschoal, A., Bailey, L. L., Doherty, P. F., de Frias-Barreto, M., & Chiarello, A. G. (2018). Effect of humans and pumas on the temporal activity of ocelots in protected areas of Atlantic Forest. Mammalian Biology, 92, 86–93. https://doi.org/10.1016/j.mambio.2018.04.009
Massara, R., De Oliveira-Paschoal, A., Bailey, L. L., Doherty, P. F., Hirsch, A., & Chiarello, A. G. (2018). Factors influencing ocelot occupancy in Brazilian Atlantic Forest reserves. Biotropica, 50, 125–134. https://doi.org/10.1111/btp.12481
Meredith, M., & Ridout, M. (2018). Overview of the overlap package. R Project.
Meredith, M., & Ridout, M. (2020). ‘overlap’ (version 0.3.3). R Package. Accessed on 10th, January, 2020 from: https://cran.r-project.org/web/packages/overlap/index.html
Montalvo, V. H., Saénz-Bolanõs, C., Alfaro, L. D., Cruz, J. C., Guimarães-Rodrigues, F. H., Carrillo, E. et al. (2019). Seasonal use of waterholes and pathways by macrofauna in the dry forest of Costa Rica. Journal of Tropical Ecology, 35, 68–73. https://doi.org/10.1017/S0266467418000457
Monterroso, P., Alves, P. C., & Ferreras, P. (2013). Catch me if you can: Diel activity patterns of mammalian prey and predators. Ethology, 119, 1044–1056. https://doi.org/10.1111/eth.12156
Moreira-Ramírez, J. F., Reyna-Hurtado, R., Hidalgo-Mihart, M., Naranjo, E., Ribeiro, M. C., García-Anleu, R. et al. (2016). Importancia de las aguadas para el pecarí de labios blancos (Tayassu pecari) en la Selva Maya, Guatemala. Therya, 7, 51–64. https://doi.org/10.12933/therya-16-344
Moreno, R. S., Kays, R. W., & Samudio, R. (2006). Competitive release in diets of ocelot (Leopardus pardalis) and puma (Puma concolor) after jaguar (Panthera onca) decline. Journal of Mammalogy, 87, 808–816. https://doi.org/10.1644/05-MAMM-A-360R2.1
Moreno, R., & Bustamante, A. (2009). Datos ecológicos del ocelote (Leopardus pardalis) en Cana, Parque Nacional Darien, Panamá; utilizando el método de cámaras trampa. Tecnociencia, 11, 91–102. https://www.revistas.up.ac.pa/index.php/tecnociencia/article/view/859
Murray, B. J. L., & Gardner, G. L. (1997). Leopardus pardalis. Mammalian Species, 548, 1–10. https://doi.org/10.2307/3504082
Nagy-Reis, M. B., Iwakami, V. H. S., Estevo, C. A., & Setz, E. Z. F. (2019). Temporal and dietary segregation in a neotropical small-felid assemblage and its relation to prey activity. Mammalian Biology, 95, 1–8. https://doi.org/10.1016/j.mambio.2018.12.005
Nasanbat, B., Ceacero, F., & Ravchig, S. (2021). A small neighborhood well-organized: seasonal and daily activity patterns of the community of large and mid-sized mammals around waterholes in the Gobi Desert, Mongolia. Frontiers in Zoology, 18, 1–11. https://doi.org/10.1186/s12983-021-00412-1
O’Farrill, G., Schampaert, K. G., Rayfield, B., Bodin, Ö., Calmé, S., Sengupta, R. et al. (2014). The potential connectivity of waterhole networks and the effectiveness of a protected area under various drought scenarios. Plos One, 9, e95049. https://doi.org/10.1371/journal.pone.0095049
Oliveira, T. G., Tortato, M. A., Silveira, L., Mazim, D., Lucherini, M., Ja, A. T. et al. (2010). Ocelot ecology and its effect on the small-felid guild in the lowland neotropics. In D. W. Macdonald, & A. J. Loveridge, (Eds.), Biology and conservation of wild felids (pp. 559–580). New York: Oxford University Press.
Oliveira-Santos, L. G. R., Graipel, M. E., Tortato, M. A., Zucco, C. A., Cáceres, N. C., & Goulart, F. V. B. (2012). Abundance changes and activity flexibility of the oncilla, Leopardus tigrinus (Carnivora: Felidae), appear to reflect avoidance of conflict. Zoologia, 29, 115–120. https://doi.org/10.1590/S1984-46702012000200003
Palomares, F., & Caro, T. M. (1999). Interspecific killing among mammalian carnivores. American Naturalist, 153, 492-508. https://doi.org/10.1086/303189
Paviolo, A., Crawshaw, P., Caso, A., Oliveira, T., López-González, C. A., Kelly, M. et al. (2015). Leopardus pardalis (errata version published in 2016). The IUCN Red List of Threatened Species 2015: e.T11509A97212355. Accessed on 10th, January, 2020 from: http://dx.doi.org/10.2305/IUCN.UK.2015-4.RLTS.T11509A50653476.en.
Penido, G., Astete, S., Jácomo, A. T. A., Sollmann, R., Tôrres, N., Silveira, L. et al. (2017). Mesocarnivore activity patterns in the semiarid Caatinga: Limited by the harsh environment or affected by interspecific interactions? Journal of Mammalogy, 98, 1732–1740. https://doi.org/10.1093/jmammal/gyx119
Perera-Romero, L., García-Anleu, R., McNab, R. B., & Thornton, D. H. (2020). When waterholes get busy, rare interactions thrive: Photographic evidence of a jaguar (Panthera onca) killing an ocelot (Leopardus pardalis). Biotropica, 53, 367–371. https://doi.org/10.1111/btp.12916
Pérez-Irineo, G., & Santos-Moreno, A. (2014). Density, distribution, and activity of the ocelot Leopardus pardalis (Carnivora: Felidae) in Southeast Mexican rainforests. Revista de Biología Tropical, 62, 1421–1432. https://doi.org/10.15517/rbt.v62i4.12941
Ponce, G. (2018). Monitoreo de cuerpos de agua y fauna asociada en la Selva Maya. Reporte 2018. Aguadas y fauna asociada Selva Maya.
Porfirio, G., Foster, V. C., Fonseca, C., & Sarmento, P. (2016). Activity patterns of ocelots and their potential prey in the Brazilian Pantanal. Mammalian Biology, 81, 511–517. https://doi.org/10.1016/j.mambio.2016.06.006
R Core Team. (2020). R: A language and environment for statistical computing. Accessed on 10th, January, 2020 from: https://www.R-project.org/
Rayleigh, L. (1880). On the resultant of a large number of vibrations of the same pitch and of arbitrary phase. Philosophical Magazine, 10, 73–78. https://doi.org/10.1080/
14786448008626893
Reid, F. A. (2009). A field guide to the mammals of Central America and Southeast Mexico. New York: Oxford University Press.
Reyna-Hurtado, R., Sima-Pantí, D., Andrade, M., Padilla, A., Retana-Guaiscon, O., Sánchez-Pinzón, K. et al. (2019). Tapir population patterns under the disappearance of freestanding water. Therya, 10, 353–358. https://doi.org/10.12933/therya-
19-902
Ridout, M. S., & Linkie, M. (2009). Estimating overlap of daily activity patterns from camera trap data. Journal of Agricultural, Biological, and Environmental Statistics, 14, 322–337. https://doi.org/10.1198/jabes.2009.08038
Ross, J., Hearn, A. J., Johnson, P. J., & Macdonald, D. W. (2013). Activity patterns and temporal avoidance by prey in response to Sunda clouded leopard predation risk. Journal of Zoology, 290, 96–106. https://doi.org/10.1111/jzo.12018
Sánchez-Pinzón, K., Reyna-Hurtado, R., Naranjo, E. J., & Alexine, K. (2020). Peccaries and their relationship with water availability and their predators in Calakmul, México. Therya, 11, 1–8. https://doi.org/10.12933/therya-20-878
Sandoval-Serés, E., Reyna-Hurtado, R., Briceño-Mendes, M., & De
La Cerda-Vega, R. (2016). Pond use and relative abundance of Tapirus bairdii in the Calakmul region, Campeche, Mexi-
co. Therya, 7, 39–50. https://doi.org/10.12933/therya-16-349
Santos, F., Carbone, C., Wearn, O. R., Rowcliffe, J. M., Espinosa, S., Moreira, M. G. et al. (2019). Prey availability and temporal partitioning modulate felid coexistence in Neotropical forests. Plos One, 14, 1–23. https://doi.org/10.1371/journal.pone.0213671
Semarnat (Secretaría de Medio Ambiente y Recursos Naturales). 2019. Modificación del Anexo Normativo III, Lista de especies en riesgo de la Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental-Especies nativas de México de flora y fauna silvestre- Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-Lista de especies en riesgo. Diario Oficial de la Federación, DOF: 14/11/2019 (pp. 32–136).
Silva-Pereira, J. E., Moro-Ríos, R. F., Bilski, D. R., & Passos, F. C. (2011). Diets of three sympatric Neotropical small cats: food niche overlap and interspecies differences in prey consumption. Mammalian Biology, 76, 308–312. https://doi.org/10.1016/j.mambio.2010.09.001
Stoner, K. E., & Timm, R. M. (2010). Seasonally dry tropical forests mammals: adaptations and seasonal patterns. In R. Dirzo, H. S. Young, H. A. Mooney, & G. Ceballos (Eds.), Seasonally dry tropical forests: Ecology and Conservation (pp. 85–106). Washington D.C.: Island Press.
Tang, X., Tang, S., Li, X., Menghe, D., Bao, W., & Xiang, C. (2019). A study of population size and activity patterns and their relationship to the prey species of the Eurasian. Animals, 9, 864. https://doi.org/10.3390/ani9110864
Torres-Romero, E. J., Espinoza-Medinilla, E., Lazcano-Barrero, M., & Maffei, L. (2017). Ecology and conservation of ocelot (Leopardus pardalis) in Northern Quintana Roo, Mexico. Therya, 8, 11–18. https://doi.org/10.12933/therya-17-439
Trolle, M., & Kéry, M. (2003). Estimation of ocelot density in the pantanal using capture-recapture analysis of camera-trapping data. Journal of Mammalogy, 84, 607–614. https://doi.org/10.1644/1545-1542(2003)084<0607:EOODIT>2.0.CO;2
Valeix, M., Loveridge, A. J., Davidson, Z., Madzikanda, H., Fritz, H., & Macdonald, D. W. (2010). How key habitat features influence large terrestrial carnivore movements: waterholes and African lions in a semi-arid savanna of north-western Zimbabwe. Landscape Ecology, 25, 337–351. https://doi.org/10.1007/s10980-009-9425-x
Wang, B., Rocha, D. G., Abrahams, M. I., Antunes, A. P., Costa, H. C. M., Gonçalves, A. L. S. et al. (2019). Habitat use of the ocelot (Leopardus pardalis) in Brazilian Amazon. Ecology and Evolution, 9, 5049–5062. https://doi.org/10.1002/ece3.5005
Wang, E. (2002). Diets of ocelots (Leopardus pardalis), margays (L . wiedii), and oncillas (L . tigrinus) in the Atlantic rainforest in Southeast Brazil. Studies on Neotropical Fauna and Environment, 37, 207–212. http://dx.doi.org/10.1076/snfe.37.3.207.8564
Weckel, M., Giuliano, W., & Silver, S. (2006). Jaguar (Panthera onca) feeding ecology: distribution of predator and prey through time and space. Journal of Zoology, 270, 25–30. https://doi.org/10.1111/j.1469-7998.2006.00106.x
Wolff, N. M., Ferreguetti, A. C., Tomas, W. M., & Bergallo, H. G. (2019). Population density, activity pattern and habitat use of the ocelot Leopardus pardalis in an Atlantic Forest protected area, Southeastern Brazil. Hystrix, the Italian Journal of Mammalogy, 30, 120–125. https://doi.org/doi:10.4404/hystrix–00214-2019
Zar, J. (2010). Biostatistical analysis. New Jersey: Pearson.